Fatty Acid Elongation in Non-Alcoholic Steatohepatitis and Hepatocellular Carcinoma


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
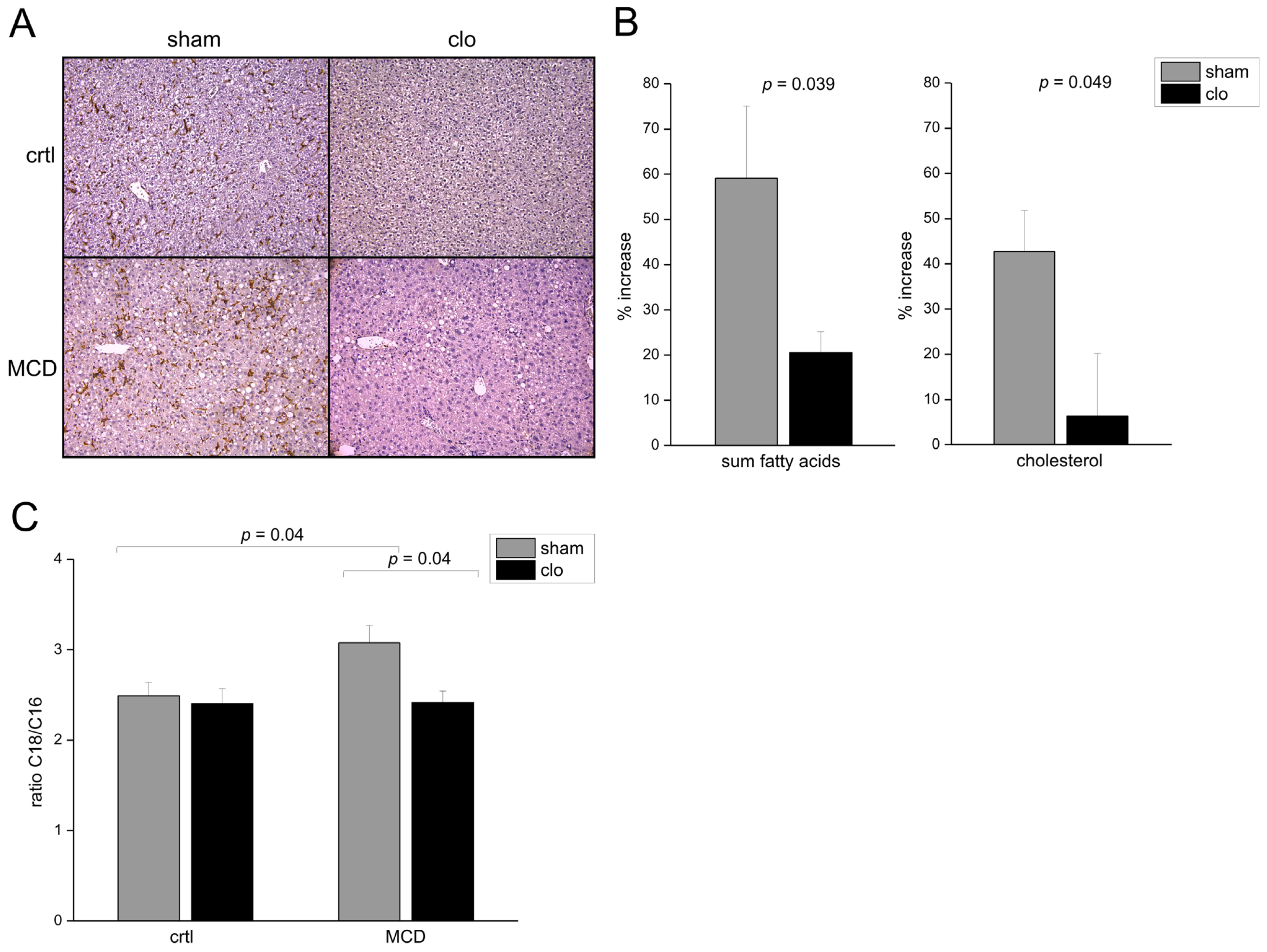
2.1. Fatty Acid Elongation in Murine Non-Alcoholic Steatohepatitis (NASH) Is Kupffer Cell Dependent
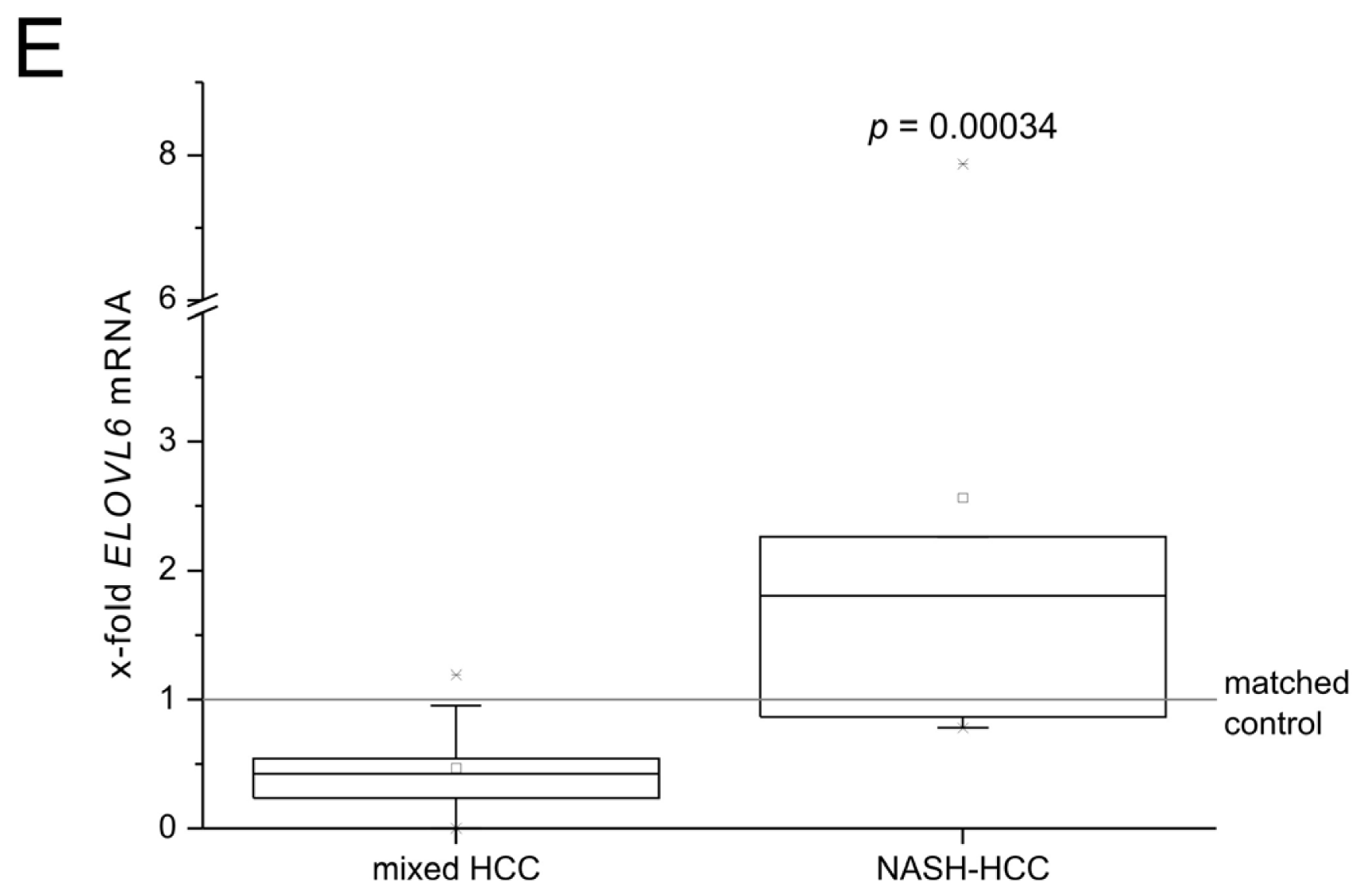
2.2. Role of Fatty Acid Elongation in NASH-Related Hepatocellular Carcinoma (HCC) and Human NASH
3. Experimental Section
3.1. Animals
3.2. Human Liver Tissue
3.3. Fatty Acid Measurement by Gas Chromatography-Mass Spectrometry (GC-MS)
3.4. Histochemistry and Immunohistochemistry
3.5. Analysis of the Public Gene Omnibus (GEO) Datasets
3.6. Statistical Analysis
4. Conclusions
Acknowledgments
Conflicts of Interest
- Author ContributionsS.M.K., Y.S., and A.K.K. designed experiments, analysed data and wrote the manuscript. A.K.K. initiated and directed the study. The others designed experiments and participated in data acquisition.
References
- Kanuri, G.; Bergheim, I. In vitro and in vivo models of non-alcoholic fatty liver disease (NAFLD). Int. J. Mol. Sci 2013, 14, 11963–11980. [Google Scholar]
- Adams, L.A.; Angulo, P.; Lindor, K.D. Nonalcoholic fatty liver disease. CMAJ 2005, 172, 899–905. [Google Scholar]
- Browning, J.D.; Szczepaniak, L.S.; Dobbins, R.; Nuremberg, P.; Horton, J.D.; Cohen, J.C.; Grundy, S.M.; Hobbs, H.H. Prevalence of hepatic steatosis in an urban population in the United States: Impact of ethnicity. Hepatology 2004, 40, 1387–1395. [Google Scholar]
- De Alwis, N.M.W.; Day, C.P. Non-alcoholic fatty liver disease: The mist gradually clears. J. Hepatol 2008, 48, S104–S112. [Google Scholar]
- Adams, L.A.; Waters, O.R.; Knuiman, M.W.; Elliott, R.R.; Olynyk, J.K. NAFLD as a risk factor for the development of diabetes and the metabolic syndrome: An eleven-year follow-up study. Am. J. Gastroenterol 2009, 104, 861–867. [Google Scholar]
- Ascha, M.S.; Hanouneh, I.A.; Lopez, R.; Tamimi, T.A.R.; Feldstein, A.F.; Zein, N.N. The incidence and risk factors of hepatocellular carcinoma in patients with nonalcoholic steatohepatitis. Hepatology 2010, 51, 1972–1978. [Google Scholar]
- Fabbrini, E.; Sullivan, S.; Klein, S. Obesity and nonalcoholic fatty liver disease: Biochemical, metabolic, and clinical implications. Hepatology 2010, 51, 679–689. [Google Scholar]
- Angulo, P. Medical progress: Nonalcoholic fatty liver disease. N. Engl. J. Med 2002, 346, 1221–1231. [Google Scholar]
- Day, C.P. Genetic and environmental susceptibility to non-alcoholic fatty liver disease. Dig. Dis 2006, 28, 255–260. [Google Scholar]
- Malhi, H.; Gores, G.J. Molecular mechanisms of lipotoxicity in nonalcoholic fatty liver disease. Semin. Liver Dis 2008, 28, 360–369. [Google Scholar]
- Cohen, J.C.; Horton, J.D.; Hobbs, H.H. Human fatty liver disease: Old questions and new insights. Science 2011, 332, 1519–1523. [Google Scholar]
- Alkhouri, N.; Dixon, L.J.; Feldstein, A.E. Lipotoxicity in nonalcoholic fatty liver disease: Not all lipids are created equal. Expert Rev. Gastroenterol. Hepatol 2009, 3, 445–451. [Google Scholar]
- Takaki, A.; Kawai, D.; Yamamoto, K. Multiple hits, including oxidative stress, as pathogenesis and treatment target in non-alcoholic steatohepatitis (NASH). Int. J. Mol. Sci 2013, 14, 20704–20728. [Google Scholar]
- Puri, P.; Baillie, R.A.; Wiest, M.M.; Mirshahi, F.; Choudhury, J.; Cheung, O.; Sargeant, C.; Contos, M.J.; Sanyal, A.J. A lipidomic analysis of nonalcoholic fatty liver disease. Hepatology 2007, 46, 1081–1090. [Google Scholar]
- Puri, P.; Wiest, M.M.; Cheung, O.; Mirshahi, F.; Sargeant, C.; Min, H.K.; Contos, M.J.; Sterling, R.K.; Fuchs, M.; Zhou, H.; et al. The plasma lipidomic signature of nonalcoholic steatohepatitis. Hepatology 2009, 50, 1827–1838. [Google Scholar]
- Kim, K.H.; Shin, H.; Kim, K.; Choi, H.M.; Rhee, S.H.; Moon, H.; Kim, H.H.; Yang, U.S.; Yu, D.; Cheong, J. Hepatitis B virus X protein induces hepatic steatosis via transcriptional activation of SREBP1 and PPARγ. Gastroenterology 2007, 132, 1955–1967. [Google Scholar]
- Miyoshi, H.; Moriya, K.; Tsutsumi, T.; Shinzawa, S.; Fujie, H.; Shintani, Y.; Fujinaga, H.; Goto, K.; Todoroki, T.; Suzuki, T.; et al. Pathogenesis of lipid metabolism disorder in hepatitis C: Polyunsaturated fatty acids counteract lipid alterations induced by the core protein. J. Hepatol 2011, 54, 432–438. [Google Scholar]
- Moriya, K.; Todoroki, T.; Tsutsumi, T.; Fujie, H.; Shintani, Y.; Miyoshi, H.; Ishibashi, K.; Takayama, T.; Makuuchi, M.; Watanabe, K.; et al. Increase in the concentration of carbon 18 monounsaturated fatty acids in the liver with hepatitis C: Analysis in transgenic mice and humans. Biochem. Biophys. Res. Commun 2001, 281, 1207–1212. [Google Scholar]
- Yamaguchi, A.; Tazuma, S.; Nishioka, T.; Ohishi, W.; Hyogo, H.; Nomura, S.; Chayama, K. Hepatitis C virus core protein modulates fatty acid metabolism and thereby causes lipid accumulation in the liver. Dig. Dis. Sci 2005, 50, 1361–1371. [Google Scholar]
- Wood, C.B.; Habib, N.A.; Apostolov, K. Reduction in the stearic to oleic acid ratio in human malignant liver neoplasms. Eur. J. Surg. Oncol 1985, 11, 347–348. [Google Scholar]
- Matsuzaka, T.; Shimano, H.; Yahagi, N.; Kato, T.; Atsumi, A.; Yamamoto, T.; Inoue, N.; Ishikawa, M.; Okada, S.; Ishigaki, N. Crucial role of a long-chain fatty acid elongase, ELOVL6, in obesity-induced insulin resistance. Nat. Med 2007, 13, 1193–1202. [Google Scholar]
- Matsuzaka, T.; Atsumi, A.; Matsumori, R.; Nie, T.; Shinozaki, H.; Suzuki-Kemuriyama, N.; Kuba, M.; Nakagawa, Y.; Ishii, K.; Shimada, M. Elovl6 promotes nonalcoholic steatohepatitis. Hepatology 2012, 56, 2199–2208. [Google Scholar]
- Muir, K.; Hazim, A.; He, Y.; Peyressatre, M.; Kim, D.Y.; Song, X.; Beretta, L. Proteomic and lipidomic signatures of lipid metabolism in NASH-associated hepatocellular carcinoma. Cancer Res 2013, 73, 4722–4731. [Google Scholar]
- Imajo, K.; Yoneda, M.; Kessoku, T.; Ogawa, Y.; Maeda, S.; Sumida, Y.; Hyogo, H.; Eguchi, Y.; Wada, K.; Nakajima, A. Rodent models of nonalcoholic fatty liver disease/nonalcoholic steatohepatitis. Int. J. Mol. Sci 2013, 14, 21833–21857. [Google Scholar]
- Trak-Smayra, V.; Paradis, V.; Massart, J.; Nasser, S.; Jebara, V.; Fromenty, B. Pathology of the liver in obese and diabetic ob/ob and db/db mice fed a standard or high-calorie diet. Int. J. Exp. Pathol 2011, 92, 413–421. [Google Scholar]
- Xu, H.; Barnes, G.T.; Yang, Q.; Tan, G.; Yang, D.; Chou, C.J.; Sole, J.; Nichols, A.; Ross, J.S.; Tartaglia, L.A.; et al. Chronic inflammation in fat plays a crucial role in the development of obesity-related insulin resistance. J. Clin. Investig 2003, 112, 1821–1830. [Google Scholar]
- Prieur, X.; Mok, C.Y.L.; Velagapudi, V.R.; Núnez, V.; Fuentes, L.; Montaner, D.; Ishikawa, K.; Camacho, A.; Barbarroja, N.; O’Rahilly, S.; et al. Differential lipid partitioning between adipocytes and tissue macrophages modulates macrophage lipotoxicity and M2/M1 polarization in obese mice. Diabetes 2011, 60, 797–809. [Google Scholar]
- Matsuzaka, T.; Shimano, H.; Yahagi, N.; Yoshikawa, T.; Amemiya-Kudo, M.; Hasty, A.H.; Okazaki, H.; Tamura, Y.; Iizuka, Y.; Ohashi, K.; et al. Cloning and characterization of a mammalian fatty acyl-CoA elongase as a lipogenic enzyme regulated by SREBPs. J. Lipid Res 2002, 43, 911–920. [Google Scholar]
- Leroux, A.; Ferrere, G.; Godie, V.; Cailleux, F.; Renoud, M.L.; Gaudin, F.; Naveau, S.; Prévot, S.; Makhzami, S.; Perlemuter, G.; et al. Toxic lipids stored by Kupffer cells correlates with their pro-inflammatory phenotype at an early stage of steatohepatitis. J. Hepatol 2012, 57, 141–149. [Google Scholar]
- Vonghia, L.; Michielsen, P.; Francque, S. Immunological mechanisms in the pathophysiology of non-alcoholic steatohepatitis. Int. J. Mol. Sci 2013, 14, 19867–19890. [Google Scholar]
- Saito, R.; Matsuzaka, T.; Karasawa, T.; Sekiya, M.; Okada, N.; Igarashi, M.; Matsumori, R.; Ishii, K.; Nakagawa, Y.; Iwasaki, H.; et al. Macrophage Elovl6 deficiency ameliorates foam cell formation and reduces atherosclerosis in low-density lipoprotein receptor-deficient mice. Arterioscler. Thromb. Vasc. Biol 2011, 31, 1973–1979. [Google Scholar]
- Naugler, W.E.; Sakurai, T.; Kim, S.; Maeda, S.; Kim, K.; Elsharkawy, A.M.; Karin, M. Gender disparity in liver cancer due to sex differences in MyD88-dependent IL-6 production. Science 2007, 317, 121–124. [Google Scholar]
- Park, E.J.; Lee, J.H.; Yu, G.Y.; He, G.; Ali, S.R.; Holzer, R.G.; Österreicher, C.H.; Takahashi, H.; Karin, M. Dietary and genetic obesity promote liver inflammation and tumorigenesis by enhancing IL-6 and TNF expression. Cell 2010, 140, 197–208. [Google Scholar]
- Braunbeck, T.A.; Teh, S.J.; Lester, S.M.; Hinton, D.E. Ultrastructural alterations in liver of medaka (Oryzias latipes) exposed to diethylnitrosamine. Toxicol. Pathol 1992, 20, 179–196. [Google Scholar]
- Laurén, D.J.; Teh, S.J.; Hinton, D.E. Cytotoxicity phase of diethylnitrosamine-induced hepatic neoplasia in medaka. Cancer Res 1990, 50, 5504–5514. [Google Scholar]
- Abel, S.; Smuts, C.M.; de Villiers, C.; Gelderblom, W.C.A. Changes in essential fatty acid patterns associated with normal liver regeneration and the progression of hepatocyte nodules in rat hepatocarcinogenesis. Carcinogenesis 2001, 22, 795–804. [Google Scholar]
- Canuto, R.A.; Biocca, M.E.; Muzio, G.; Dianzani, M.U. Fatty acid composition of phospholipids in mitochondria and microsomes during diethylnitrosamine carcinogenesis in rat liver. Cell Biochem. Funct 1989, 7, 11–19. [Google Scholar]
- Yoshimura, K.; Mandal, M.K.; Hara, M.; Fujii, H.; Chen, L.C.; Tanabe, K.; Hiraoka, K.; Takeda, S. Real-time diagnosis of chemically induced hepatocellular carcinoma using a novel mass spectrometry-based technique. Anal. Biochem 2013, 441, 32–37. [Google Scholar]
- Kessler, S.M.; Laggai, S.; Barghash, A.; Helms, V.; Kiemer, A.K. Lipid metabolism signatures in NASH-associated HCC. Cancer Res 2014. [Google Scholar] [CrossRef]
- Kessler, S.M.; Pokorny, J.; Zimmer, V.; Laggai, S.; Lammert, F.; Bohle, R.M.; Kiemer, A.K. IGF2 mRNA binding protein p62/IMP2-2 in hepatocellular carcinoma: Antiapoptotic action is independent of IGF2/PI3K signaling. Am. J. Physiol 2013, 304, G328–G336. [Google Scholar]
- Ahrens, M.; Ammerpohl, O.; von Schönfels, W.; Kolarova, J.; Bens, S.; Itzel, T.; Teufel, A.; Herrmann, A.; Brosch, M.; Hinrichsen, H.; et al. DNA methylation analysis in nonalcoholic fatty liver disease suggests distinct disease—Specific and remodeling signatures after bariatric surgery. Cell Metab 2013, 18, 296–302. [Google Scholar]
- López-Vicario, C.; González-Périz, A.; Rius, B.; Morán-Salvador, E.; García-Alonso, V.; Lozano, J.J.; Bataller, R.; Cofán, M.; Kang, J.X.; Arroyo, V.; et al. Molecular interplay between D5/D6 desaturases and long-chain fatty acids in the pathogenesis of non-alcoholic steatohepatitis. Gut 2014, 63, 344–355. [Google Scholar]
- Keller, M.; Gerbes, A.L.; Kulhanek-Heinze, S.; Gerwig, T.; Grützner, U.; van Rooijen, N.; Vollmar, A.M.; Kiemer, A.K. Hepatocyte cytoskeleton during ischemia and reperfusion— Influence of ANP-mediated p38 MAPK activation. World J. Gastroenterol 2005, 11, 7418–7429. [Google Scholar]
- Bode, H.B.; Ring, M.W.; Schwär, G.; Kroppenstedt, R.M.; Kaiser, D.; Müller, R. 3-Hydroxy-3- methylglutaryl-coenzyme A (CoA) synthase is involved in biosynthesis of isovaleryl-CoA in the myxobacterium Myxococcus xanthus during fruiting body formation. J. Bacteriol 2006, 188, 6524–6528. [Google Scholar]
- Simon, Y.; Kessler, S.M.; Bohle, R.M.; Haybaeck, J.; Kiemer, A.K. The insulin-like growth factor 2 (IGF2) mRNA-binding protein p62/IGF2BP2-2 as a promoter of NAFLD and HCC? Gut 2013, in press. [Google Scholar] [CrossRef]
- Tybl, E.; Shi, F.-D.; Kessler, S.M.; Tierling, S.; Walter, J.; Bohle, R.M.; Wieland, S.; Zhang, J.; Tan, E.M.; Kiemer, A.K. Overexpression of the IGF2-mRNA binding protein p62 in transgenic mice induces a steatotic phenotype. J. Hepatol 2011, 54, 994–1001. [Google Scholar]
- Edgar, R.; Domrachev, M.; Lash, A.E. Gene expression omnibus: NCBI gene expression and hybridization array data repository. Nucleic Acids Res 2002, 30, 207–210. [Google Scholar]




© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kessler, S.M.; Simon, Y.; Gemperlein, K.; Gianmoena, K.; Cadenas, C.; Zimmer, V.; Pokorny, J.; Barghash, A.; Helms, V.; Van Rooijen, N.; et al. Fatty Acid Elongation in Non-Alcoholic Steatohepatitis and Hepatocellular Carcinoma. Int. J. Mol. Sci. 2014, 15, 5762-5773. https://doi.org/10.3390/ijms15045762
Kessler SM, Simon Y, Gemperlein K, Gianmoena K, Cadenas C, Zimmer V, Pokorny J, Barghash A, Helms V, Van Rooijen N, et al. Fatty Acid Elongation in Non-Alcoholic Steatohepatitis and Hepatocellular Carcinoma. International Journal of Molecular Sciences. 2014; 15(4):5762-5773. https://doi.org/10.3390/ijms15045762
Chicago/Turabian StyleKessler, Sonja M., Yvette Simon, Katja Gemperlein, Kathrin Gianmoena, Cristina Cadenas, Vincent Zimmer, Juliane Pokorny, Ahmad Barghash, Volkhard Helms, Nico Van Rooijen, and et al. 2014. "Fatty Acid Elongation in Non-Alcoholic Steatohepatitis and Hepatocellular Carcinoma" International Journal of Molecular Sciences 15, no. 4: 5762-5773. https://doi.org/10.3390/ijms15045762