
Polymorphisms in the LPL and CETP Genes and Haplotype in the ESR1 Gene Are Associated with Metabolic Syndrome in Women from Southwestern Mexico

 ,
,
Abstract
:1. Introduction
2. Results

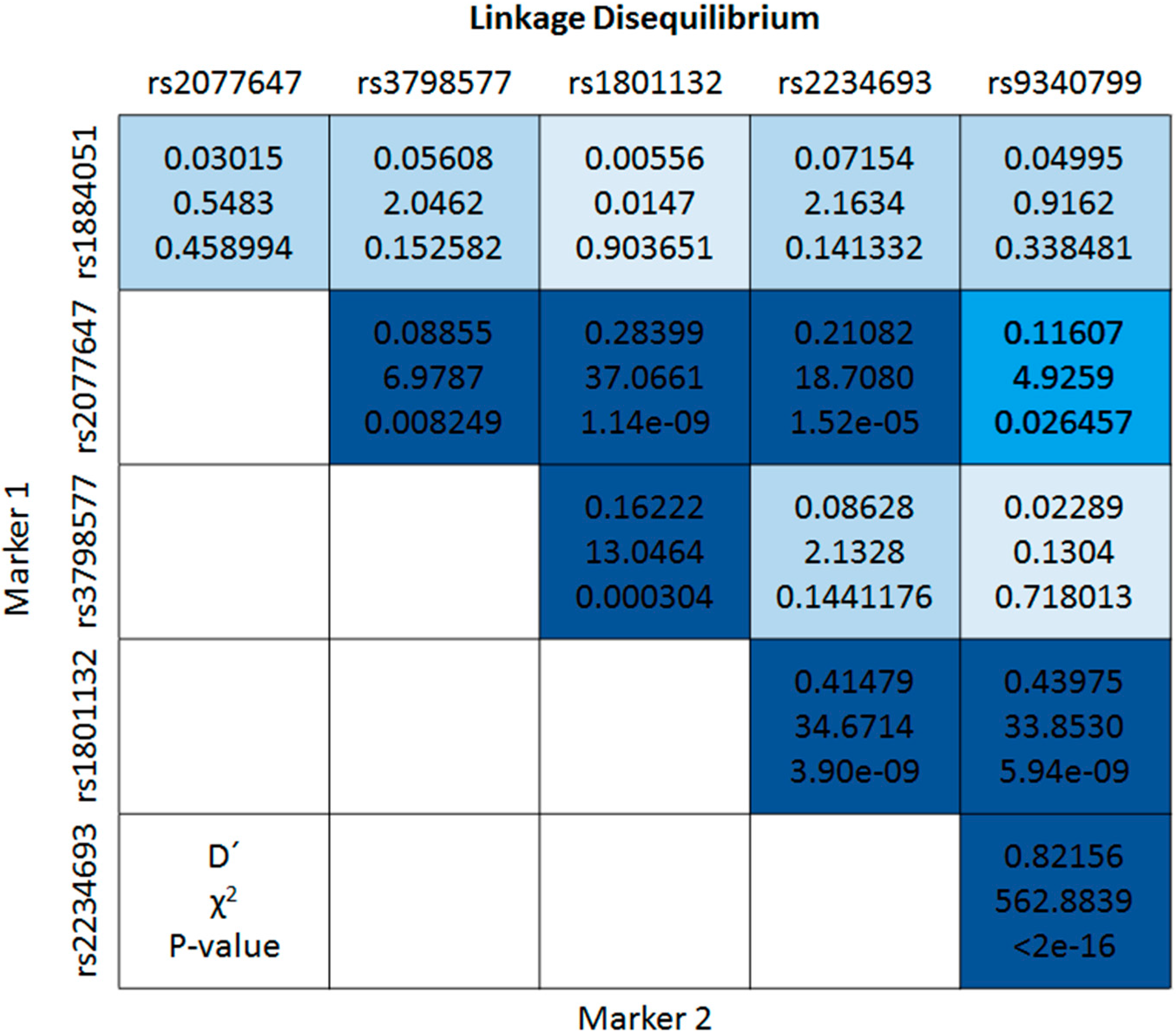{kind=link}
| Characteristic | n = 480 | MetS n = 159 (33.1%) | Without MetS n = 321 (66.9%) | p |
|---|---|---|---|---|
| Age (year) | 46 (38–53) | 51 (44–56) | 43 (36–50) | <0.001 † |
| BMI (kg/m2) | 27.3 (24.8–30.3) | 29.2 (27–33.4) | 26.3 (24–29) | <0.001 † |
| Abdominal obesity, n (%) | 271 (56.5) | 141 (88.7) | 130 (40.5) | <0.001 ‡ |
| % Body fat | 36.6 (31.4–40.9) | 39.5 (35.4–43.6) | 35.4 (29.7–38.8) | <0.001 † |
| % Water | 44.4 (41.8–47.9) | 42.2 (39.9–45.1) | 45.3 (43–48.9) | <0.001 † |
| BP systolic, (mm·Hg) | 116 (107–127) | 129 (117–137) | 112 (105–120) | <0.001 † |
| BP diastolic, (mm·Hg) | 73.5 (67–81) | 79 (71–86) | 71 (66–78) | <0.001 † |
| Glucose (mg/dL) | 79.7 (71.5–89) | 86 (77–107) | 76.9 (70–83.4) | <0.001 † |
| ≥110, n (%) | 54 (11.3) | 43 (27) | 11 (3.4) | <0.001 ‡ |
| Total cholesterol (mg/dL) | 172 (146.7–197) | 184.3 (159.9–207) | 165 (142–191.1) | <0.001 † |
| ≥200, n (%) | 115 (24) | 49 (30.8) | 66 (20.6) | 0.013 ‡ |
| Triglycerides, (mg/dL) | 130.5 (93.4–170.8) | 170 (145–212.5) | 110 (80.5–141) | <0.001 † |
| ≥150, n (%) | 175 (36.5) | 114 (71.7) | 61 (19) | <0.001 ‡ |
| HDL-c, (mg/dL) | 39.9 (32.3–49.5) | 37.6 (31.5–44.2) | 40.4 (33.5–53.5) | <0.001 † |
| <50, n (%) | 366 (76.3) | 147 (92.5) | 219 (68.2) | <0.001 ‡ |
| LDL-c, mg/dL | 119 (90.5–157.2) | 126.3 (95.3–168.3) | 112.9 (88.8–155) | 0.04 † |
| ≥160, n (%) | 114 (23.9) | 42 (26.8) | 72 (22.5) | 0.306 ‡ |
| Exercise, n (%) | 235 (49.1) | 80 (50.6) | 155 (48.3) | 0.629 ‡ |
| Years of schooling | 17 (12–18) | 17 (12–18) | 17 (12–19) | 0.155 |
| Ancestry, % | ||||
| Native American | 69.2 | 68.8 | 69.4 | 0.548 † |
| European | 27.1 | 27.7 | 26.7 | 0.275 † |
| African | 3.7 | 3.5 | 3.9 | 0.602 † |
| SNP | Total | Allele Frequencies | Allele Frequencies (dbSNP-NCBI) | MetS | Without MetS | p ‡ |
|---|---|---|---|---|---|---|
| ESR1 | n (%) | n (%) | n (%) | |||
| rs1884051 | ||||||
| AA | 95 (19.8) | A: 0.442 | A = 0.509 | 29 (18.2) | 66 (20.6) | 0.251 |
| AG | 234 (48.7) | G: 0.558 | G = 0.491 | 86 (54.1) | 148 (46.1) | |
| GG | 151 (31.5) | 44 (27.7) | 107 (33.3) | |||
| rs3798577 | ||||||
| TT | 142 (29.6) | T: 0.539 | T = 0.536 | 44 (27.7) | 98 (30.5) | 0.740 |
| CT | 233 (48.5) | C: 0.461 | C = 0.464 | 81 (50.9) | 152 (47.4) | |
| CC | 105 (21.9) | 34 (21.4) | 71 (22.1) | |||
| rs2077647 | ||||||
| TT | 146 (30.4) | T: 0.557 | T = 0.533 | 49 (30.8) | 97 (30.2) | 0.991 |
| CT | 243 (50.6) | C: 0.443 | C = 0.467 | 80 (50.3) | 163 (50.8) | |
| CC | 91 (19.0) | 30 (18.9) | 61 (19.0) | |||
| rs1801132 | ||||||
| CC | 190 (39.6) | C: 0.624 | C = 0.718 | 64 (40.2) | 126 (39.2) | 0.975 |
| CG | 219 (45.6) | G: 0.376 | G = 0.282 | 72 (45.3) | 147 (45.8) | |
| GG | 71 (14.8) | 23 (14.5) | 48 (15.0) | |||
| rs2234693 | ||||||
| TT | 270 (56.3) | T = 0.742 * | T = 0.554 * | 93 (58.5) | 177 (55.1) | 0.254 |
| CT | 172 (35.8) | C = 0.258 | C = 0.446 | 58 (36.5) | 114 (35.5) | |
| CC | 38 (7.9) | 8 (5.0) | 30 (9.4) | |||
| rs9340799 | ||||||
| AA | 295 (61.5) | A = 0.768 | A = 0.719 | 102 (64.2) | 193 (60.1) | 0.309 |
| AG | 147 (30.6) | G = 0.232 | G = 0.281 | 42 (26.4) | 105 (32.7) | |
| GG | 38 (7.9) | 15 (9.4) | 23 (7.2) | |||
| LPL | ||||||
| rs320 | ||||||
| TT | 325 (67.7) | T = 0.821 | T = 0.738 | 108 (67.9) | 217 (67.6) | 0.366 |
| TG | 138 (28.8) | G = 0.179 | G = 0.262 | 48 (30.2) | 90 (28.0) | |
| GG | 17 (3.5) | 3 (1.9) | 14 (4.4) | |||
| rs328 | ||||||
| CC | 444 (92.5) | C = 0.961 | C = 0.907 | 152 (95.6) | 292 (91.0) | 0.043 |
| CG | 35 (7.3) | G = 0.039 | G = 0.093 | 6 (3.8) | 29 (9.0) | |
| GG | 1 (0.2) | 1 (0.6) | 0 | |||
| CETP | ||||||
| rs708272 | ||||||
| GG | 115 (24.0) | G = 0.493 * | G = 0.622 * | 32 (20.1) | 83 (25.9) | 0.032 |
| GA | 243 (50.6) | A = 0.507 | A = 0.378 | 94 (59.1) | 149 (46.4) | |
| AA | 122 (25.4) | 33 (20.8) | 89 (27.7) |
| Factor | Inheritance Model ‡ | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Co-Dominant | Dominant | Over-Dominant | |||||||||||||
| GG | AG | OR (95% CI) | p | AA | OR (95% CI) | p | GG | AG + AA | OR (95% CI) | p | GG + AA | AG | OR (95% CI) | p | |
| MetS, n (%) | |||||||||||||||
| No | 107 (33.3) | 148 (46.1) | 1.0 | 66 (20.6) | 1.0 | 107 (33.3) | 214 (66.7) | 1.0 | 173 (53.9) | 148 (46.1) | 1.0 | ||||
| Yes | 44 (27.7) | 86 (54.1) | 1.3 (0.8–2.1) | 0.237 | 29 (18.2) | 0.9 (0.5–1.7) | 0.768 | 44 (27.7) | 115 (72.3) | 1.2 (0.7–1.9) | 0.430 | 73 (45.9) | 86 (54.1) | 1.4 (0.9–2.1) | 0.125 |
| AO > 88 cm, n (%) | |||||||||||||||
| No | 65 (31.1) | 98 (46.9) | 1.0 | 46 (22.0) | 1.0 | 65 (31.1) | 144 (68.9) | 1.0 | 111 (53.1) | 98 (46.9) | 1.0 | ||||
| Yes | 86 (31.7) | 136 (50.2) | 1.0 (0.6–1.5) | 0.890 | 49 (18.1) | 0.7 (0.4–1.3) | 0.768 | 86 (31.7) | 185 (68.3) | 0.9 (0.6–1.3) | 0.599 | 135 (49.8) | 136 (50.2) | 1.1 (0.8–1.6) | 0.653 |
| BP ≥ 130/85 mm·Hg, n (%) | |||||||||||||||
| No | 119 (33.1) | 172 (47.8) | 1.0 | 69 (19.2) | 1.0 | 119 (33.1) | 241 (66.9) | 1.0 | 188 (52.2) | 172 (47.8) | 1.0 | ||||
| Yes | 32 (26.7) | 62 (51.6) | 1.3 (0.8–2.1) | 0.362 | 26 (21.7) | 1.3 (0.7–2.4) | 0.458 | 32 (26.7) | 88 (73.3) | 1.3 (0.8–2.0) | 0.335 | 58 (48.3) | 62 (51.6) | 1.1 (0.7–1.8) | 0.530 |
| Glucose ≥110 mg/dL or T2D, n (%) | |||||||||||||||
| No | 140 (32.9) | 209 (49.0) | 1.0 | 77 (18.1) | 1.0 | 140 (32.9) | 286 (67.1) | 1.0 | 217 (50.9) | 209 (49.0) | 1.0 | ||||
| Yes | 11 (20.4) | 25 (46.3) | 1.4 (0.7–3.1) | 0.351 | 18 (33.3) | 2.9 (1.2–6.6) | 0.013 | 11 (20.4) | 43 (79.6) | 1.8 (0.9–3.7) | 0.102 | 29 (53.7) | 25 (46.3) | 0.9 (0.5–1.5) | 0.606 |
| TG ≥ 150 mg/dL, n (%) | |||||||||||||||
| No | 97 (31.8) | 140 (45.9) | 1.0 | 68 (22.3) | 1.0 | 97 (31.8) | 208 (68.2) | 1.0 | 165 (54.1) | 140 (45.9) | 1.0 | ||||
| Yes | 54 (30.9) | 94 (53.7) | 1.2 (0.8–1.8) | 0.476 | 27 (15.4) | 0.6 (0.4–1.1) | 0.134 | 54 (30.9) | 121 (69.1) | 1.0 (0.7–1.5) | 0.980 | 81 (46.3) | 94 (53.7) | 1.4 (0.9–2.0) | 0.099 |
| HDL-c < 50 mg/dL, n (%) | |||||||||||||||
| No | 34 (29.8) | 49 (43.0) | 1.0 | 31 (27.2) | 1.0 | 34 (29.8) | 80 (70.2) | 1.0 | 65 (57.0) | 49 (43.0) | 1.0 | ||||
| Yes | 117 (32.0) | 185 (50.5) | 1.1 (0.7–1.8) | 0.764 | 64 (17.5) | 0.6 (0.3–1.0) | 0.067 | 117 (32.0) | 249 (68.0) | 0.9 (0.6–1.4) | 0.607 | 181 (49.5) | 185 (50.5) | 1.3 (0.9–2.1) | 0.168 |
| Factor | Inheritance Model ‡ | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Co-Dominant | Dominant | Over-Dominant | ||||||||||||
| CC | CG | OR (95% CI) | p | GG | OR (95% CI) | CC | CG + GG | OR (95% CI) | p | CC + GG | CG | OR (95% CI) | p | |
| MetS, n (%) | ||||||||||||||
| No | 292 (91.0) | 29 (9.0) | 1.0 | 0 | 1.0 | 292 (91.0) | 29 (9.0) | 1.0 | 292 (91.0) | 29 (9.0) | 1.0 | |||
| Yes | 152 (95.6) | 6 (3.8) | 0.3 (0.1–0.8) | 0.018 | 1 (0.6) | ND | 152 (95.6) | 7 (4.4) | 0.4 (0.1–0.9) * | 0.030 | 153 (96.2) | 6 (3.8) | 0.3 (0.1–0.8) | 0.02 |
| AO > 88 cm, n (%) | ||||||||||||||
| No | 192 (91.9) | 16 (7.6) | 1.0 | 1 (0.5) | 1.0 | 192 (91.9) | 17 (8.1) | 1.0 | 193 (92.3) | 16 (7.6) | 1.0 | |||
| Yes | 252 (93.0) | 19 (7.0) | 0.9 (0.4–1.7) | 0.664 | 0 | ND | 252 (93.0) | 19 (7.0) | 1.3 (0.6–2.5) | 0.529 | 252 (93.0) | 19 (7.0) | 0.9 (0.4–1.8) | 0.670 |
| BP ≥ 130/85 mm·Hg, n (%) | ||||||||||||||
| No | 329 (91.4) | 31 (8.6) | 1.0 | 0 | 1.0 | 329 (91.4) | 31 (8.6) | 1.0 | 329 (91.4) | 31 (8.6) | 1.0 | |||
| Yes | 115 (95.8) | 4 (3.4) | 0.3 (0.1–1.0) | 0.041 | 1 (0.8) | ND | 115 (95.8) | 5 (4.2) | 2.5 (0.9–7.0) | 0.080 | 116 (96.7) | 4 (3.3) | 0.3 (0.1–0.9) | 0.040 |
| Glucose ≥ 110 mg/dL or T2D, n (%) | ||||||||||||||
| No | 391 (91.8) | 34 (8.0) | 1.0 | 1 (0.2) | 1.0 | 391 (91.8) | 35 (8.2) | 1.0 | 392 (92.0) | 34 (8.0) | 1.0 | |||
| Yes | 53 (98.1) | 1 (1.9) | 0.2 (0.1–1.5) | 0.115 | 0 | ND | 53 (98.1) | 1 (1.9) | 5.5 (0.7–42.4) | 0.102 | 53 (98.1) | 1 (1.9) | 0.2 (0.1–1.5) | 0.116 |
| TG ≥ 150 mg/dL, n (%) | ||||||||||||||
| No | 280 (91.8) | 25 (8.2) | 1.0 | 0 | 1.0 | 280 (91.8) | 25 (8.2) | 1.0 | 280 (91.8) | 25 (8.2) | 1.0 | |||
| Yes | 164 (93.7) | 10 (5.7) | 0.7 (0.3–1.4) | 0.292 | 1 (0.6) | ND | 164 (93.7) | 11 (6.3) | 1.4 (0.7–3.1) | 0.364 | 165 (94.3) | 10 (5.7) | 0.6 (0.3–1.4) | 0.289 |
| HDL-c < 50 mg/dL, n (%) | ||||||||||||||
| No | 102 (89.5) | 12 (10.5) | 1.0 | 0 | 1.0 | 102 (89.5) | 12 (10.5) | 1.0 | 102 (89.5) | 12 (10.5) | 1.0 | |||
| Yes | 342 (93.4) | 23 (6.3) | 0.5 (0.2–1.1) | 0.090 | 1 (0.3) | ND | 342 (93.4) | 24 (6.6) | 1.8 (0.9–3.8) | 0.112 | 343 (93.7) | 23 (6.3) | 0.5 (0.2–1.1) | 0.089 |
| Factor | Inheritance Model ‡ | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Co-Dominant | Dominant | Over-Dominant | |||||||||||||
| AA | GA | OR (95% CI) | p | GG | OR (95% CI) | p | AA | GA + GG | OR (95% CI) | p | AA + GG | GA | OR (95% CI) | p | |
| MetS, n (%) | |||||||||||||||
| No | 89 (27.7) | 149 (46.4) | 1.0 | 83 (25.9) | 1.0 | 89 (27.7) | 232 (72.3) | 1.0 | 172 (53.6) | 149 (46.4) | 1.0 | ||||
| Yes | 33 (20.8) | 94 (59.1) | 1.7 (1.0–2.8) | 0.042 | 32 (20.1) | 0.9 (0.9–1.7) | 0.749 | 33 (20.8) | 126 (79.2) | 1.4 (0.9–2.3) | 0.170 | 65 (40.9) | 94 (59.1) | 1.8 (1.2–2.7) | 0.006 |
| AO > 88 cm, n (%) | |||||||||||||||
| No | 53 (25.3) | 99 (47.4) | 1.0 | 57 (27.3) | 1.0 | 53 (25.3) | 156 (74.6) | 1.0 | 110 (52.6) | 99 (47.4) | 1.0 | ||||
| Yes | 69 (25.5) | 144 (53.1) | 1.1 (0.7–1.7) | 0.794 | 58 (21.4) | 0.7 (0.4–1.2) | 0.794 | 69 (25.5) | 202 (74.5) | 0.9 (0.6–1.4) | 0.769 | 127 (46.9) | 144 (53.1) | 1.2 (0.9–1.8) | 0.250 |
| BP ≥ 130/85 mm·Hg, n (%) | |||||||||||||||
| No | 103 (28.6) | 168 (46.7) | 1.0 | 89 (24.7) | 1.0 | 103 (28.6) | 257 (71.4) | 1.0 | 192 (53.3) | 168 (46.7) | 1.0 | ||||
| Yes | 19 (15.8) | 75 (62.5) | 2.5 (1.4–4.4) | 0.002 | 26 (21.7) | 1.5 (0.7–2.9) | 0.279 | 19 (15.8) | 101 (84.2 | 2.1 (1.2–3.7) | 0.009 | 45 (37.5) | 75 (62.5) | 2.0 (1.3–3.2) | 0.002 |
| Glucose ≥ 110 mg/dL or T2D, n (%) | |||||||||||||||
| No | 117 (27.5) | 211 (49.5) | 1.0 | 98 (23.0) | 1.0 | 117 (27.5) | 309 (72.5) | 1.0 | 215 (50.5) | 211 (49.5) | 1.0 | ||||
| Yes | 5 (9.3) | 32 (59.2) | 3.5 (1.3–9.3) | 0.013 | 17 (31.5) | 3.7 (1.3–10.7) | 0.016 | 5 (9.3) | 49 (90.7) | 3.6 (1.4–9.3) | 0.010 | 22 (40.7) | 32 (59.2) | 1.5 (0.8–2.8) | 0.161 |
| TG ≥ 150 mg/dL, n (%) | |||||||||||||||
| No | 81 (26.6) | 148 (48.5) | 1.0 | 76 (24.9) | 1.0 | 81 (26.6) | 224 (73.4) | 1.0 | 157 (51.5) | 148 (48.5) | 1.0 | ||||
| Yes | 41 (23.4) | 95 (54.3) | 1.2 (0.7–2.0) | 0.375 | 39 (22.3) | 0.9 (0.5–1.6) | 0.684 | 41 (23.4) | 134 (76.6) | 1.1 (0.7–1.8) | 0.630 | 80 (45.7) | 95 (54.3) | 1.3 (0.9–1.9) | 0.171 |
| HDL-c < 50 mg/dL, n (%) | |||||||||||||||
| No | 26 (22.8) | 64 (56.1) | 1.0 | 24 (21.1) | 1.0 | 26 (22.8) | 88 (77.2) | 1.0 | 50 (43.9) | 64 (56.1) | 1.0 | ||||
| Yes | 96 (26.2) | 179 (48.9) | 0.8 (0.4–1.3) | 0.285 | 91 (24.9) | 1.0 (0.6–1.9) | 0.913 | 96 (26.2) | 270 (73.8) | 0.8 (0.5–1.4) | 0.460 | 187 (51.1) | 179 (48.9) | 0.7 (0.5–1.1) | 0.164 |

3. Discussion
4. Experimental Section
4.1. Subjects
4.2. Diagnosis of Metabolic Syndrome
4.3. Biochemical Assays
4.4. Genomic DNA Isolation and Genotyping
4.5. Ancestry Informative Markers
4.6. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wilson, P.W.F.; D’Agostino, R.B.; Parise, H.; Sullivan, L.; Meigs, J.B. Metabolic syndrome as a precursor of cardiovascular disease and type 2 diabetes mellitus. Circulation 2005, 112, 3066–3072. [Google Scholar] [CrossRef] [PubMed]
- Expert Panel on Detection, Evaluation, and Treatment of High Blood Cholesterol in Adults. Executive Summary of the Third Report of The National Cholesterol Education Program (NCEP). Expert Panel on Detection, Evaluation, and Treatment of High Blood Cholesterol in Adults (Adult Treatment Panel III). JAMA 2001, 285, 2486–2497. [Google Scholar]
- Grundy, S.M.; Brewer, H.B., Jr.; Cleeman, J.I.; Smith, S.C., Jr.; Lenfant, C. Definition of metabolic syndrome: Report of the National Heart, Lung, and Blood Institute/American Heart Association conference on scientific issues related to definition. Arterioscler. Thromb. Vasc. Biol. 2004, 109, 433–438. [Google Scholar]
- Kubrusly, M.; Costa-De Oliveira, C.M.; Freire-Simões, P.S.; de Oliveira-Lima, R.; Rabelo-Galdino, P.N.; Fortaleza-Sousa, P.A.; Carneiro-Jerônimo, A.L. Prevalence of Metabolic Syndrome according to NCEP-ATP III and IDF criteria in Patients on Hemodialysis. J. Bras. Nefrol. 2015, 37, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Miller, V.M.; Duckles, S.P. Vascular actions of estrogens: Functional implications. Pharmacol. Rev. 2008, 60, 210–241. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, C.J.; Langefeld, C.D.; Gordon, C.J.; Campbell, J.K.; Mychaleckyj, J.C.; Bryer-Ash, M.; Rich, S.S.; Bowden, D.W.; Sale, M.M. Association of the estrogen receptor-alpha gene with the metabolic syndrome and its component traits in African-American families: The Insulin Resistance Atherosclerosis Family Study. Diabetes 2007, 56, 2135–2141. [Google Scholar] [CrossRef] [PubMed]
- Ghattas, M.H.; Mehanna, E.T.; Mesbah, N.M.; Abo-Elmatty, D.M. Association of estrogen receptor alpha gene polymorphisms with metabolic syndrome in Egyptian women. Metabolism 2013, 62, 1437–1442. [Google Scholar] [CrossRef] [PubMed]
- Deroo, B.J.; Korach, K.S. Estrogen receptors and human disease. J. Clin. Investig. 2006, 116, 561–570. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, T.; Ishida, T.; Rader, D.J. Update on the role of endothelial lipase in high-density lipoprotein metabolism, reverse cholesterol transport, and atherosclerosis. Circ. J. 2010, 74, 2263–2270. [Google Scholar] [CrossRef] [PubMed]
- Nagano, M.; Yamashita, S.; Hirano, K.; Takano, M.; Maruyama, T.; Ishihara, M.; Sagehashi, Y.; Kujiraoka, T.; Tanaka, K.; Hattori, H.; et al. Molecular mechanisms of cholesteryl ester transfer protein deficiency in Japanese. J. Atheroscler. Thromb. 2004, 11, 110–121. [Google Scholar] [CrossRef] [PubMed]
- Kaur, J. A comprehensive review on metabolic syndrome. Cardiol. Res. Pract. 2014. [Google Scholar] [CrossRef] [PubMed]
- Grundy, S.M. Metabolic syndrome pandemic. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 629–636. [Google Scholar] [CrossRef] [PubMed]
- Heiss, G.; Snyder, M.L.; Teng, Y.; Schneiderman, N.; Llabre, M.M.; Cowie, C.; Carnethon, M.; Kaplan, R.; Giachello, A.; Gallo, L.; et al. Prevalence of metabolic syndrome among Hispanics/Latinos of diverse background: The Hispanic Community Health Study/Study of Latinos. Diabetes Care 2014, 37, 2391–2399. [Google Scholar] [CrossRef] [PubMed]
- Rojas, R.; Aguilar-Salinas, C.A.; Jiménez-Corona, A.; Shamah-Levy, T.; Rauda, J.; Avila-Burgos, L.; Villalpando, S.; Ponce, E.L. Metabolic syndrome in Mexican adults: Results from the National Health and Nutrition Survey 2006. Salud Publica Mex. 2010, 52, S11–S18. [Google Scholar] [CrossRef] [PubMed]
- Faulds, M.H.; Zhao, C.; Dahlman-Wright, K.; Gustafsson, J.A. The diversity of sex steroid action: Regulation of metabolism by estrogen signaling. J. Endocrinol. 2012, 212, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Magdalena, P.; Ropero, A.B.; Carrera, M.P.; Cederroth, C.R.; Baquié, M.; Gauthier, B.R.; Nef, S.; Stefani, E.; Nadal, A. Pancreatic insulin content regulation by the estrogen receptor ER alpha. PLoS ONE 2008, 3, E2039. [Google Scholar] [CrossRef] [PubMed]
- Hopp, T.A.; Fuqua, S.A. Estrogen Receptor Variants. J. Mammary Gland Biol. Neoplasia 1998, 3, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Anghel, A.; Narita, D.; Seclaman, E.; Popovici, E.; Anghel, M.; Tamas, L. Estrogen receptor alpha polymorphisms and the risk of malignancies. Pathol. Oncol. Res. 2010, 16, 485–496. [Google Scholar] [CrossRef] [PubMed]
- Klos, K.L.; Boerwinkle, E.; Ferrell, R.E.; Turner, S.T.; Morrison, A.C. ESR1 polymorphism is associated with plasma lipid and apolipoprotein levels in Caucasians of the Rochester Family Heart Study. J. Lipid Res. 2008, 49, 1701–1706. [Google Scholar] [CrossRef] [PubMed]
- Sowers, M.R.; Wilson, A.L.; Karvonen-Gutierrez, C.A.; Kardia, S.R. Sex steroid hormone pathway genes and health-related measures in women of 4 races/ethnicities: The Study of Women’s Health Across the Nation (SWAN). Am. J. Med. 2006, 119, S103–S110. [Google Scholar] [CrossRef] [PubMed]
- Goulart, A.C.; Zee, R.Y.; Pradhan, A.; Rexrode, K.M. Associations of the estrogen receptors 1 and 2 gene polymorphisms with the metabolic syndrome in women. Metab. Syndr. Relat. Disord. 2009, 7, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Rebelo, A.C.; Verlengia, R.; Kunz, V.; Tamburus, N.; Cerda, A.; Hirata, R.; Hirata, M.; Silva, E. Lack of association of estrogen receptor alpha gene polymorphisms with cardiorespiratory and metabolic variables in young women. Int. J. Mol. Sci. 2012, 13, 13691–13703. [Google Scholar] [CrossRef] [PubMed]
- Dallinga-Thie, G.M.; Franssen, R.; Mooij, H.L.; Visser, M.E.; Hassing, H.C.; Peelman, F.; Kastelein, J.J.; Péterfy, M.; Nieuwdorp, M. The metabolism of triglyceride-rich lipoproteins revisited: New players, new insight. Atherosclerosis 2010, 211, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ariza, M.J.; Sánchez-Chaparro, M.A.; Barón, F.J.; Hornos, A.M.; Calvo-Bonacho, E.; Rioja, J.; Valdivielso, P.; Gelpi, J.A.; González-Santos, P. Additive effects of LPL, APOA5 and APOE variant combinations on triglyceride levels and hypertriglyceridemia: Results of the ICARIA genetic sub-study. BMC Med. Genet. 2010, 29, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Webster, R.J.; Warrington, N.M.; Weedon, M.N.; Hattersley, A.T.; McCaskie, P.A.; Beilby, J.P.; Palmer, L.J.; Frayling, T.M. The association of common genetic variants in the APOA5, LPL and GCK genes with longitudinal changes in metabolic and cardiovascular traits. Diabetologia 2009, 52, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Emamian, M.; Avan, A.; Pasdar, A.; Mirhafez, S.R.; Sadeghzadeh, M.; Moghadam, M.; Parizadeh, S.M.; Ferns, G.A.; Ghayour-Mobarhan, M. The lipoprotein lipase S447X and cholesteryl ester transfer protein rs5882 polymorphisms and their relationship with lipid profile in human serum of obese individuals. Gene 2015, 558, 195–199. [Google Scholar] [CrossRef] [PubMed]
- Sandhofer, A.; Tatarczyk, T.; Laimer, M.; Ritsch, A.; Kaser, S.; Paulweber, B.; Ebenbichler, C.F.; Patsch, J.R. The Taq1B-variant in the cholesteryl ester-transfer protein gene and the risk of metabolic syndrome. Obesity 2008, 16, 919–922. [Google Scholar] [CrossRef] [PubMed]
- López-Ríos, L.; Nóvoa, F.J.; Chirino, R.; Varillas, F.; Boronat-Cortés, M.; Wagner, A.M. Interaction between cholesteryl ester transfer protein and hepatic lipase encoding genes and the risk of type 2 diabetes: Results from the Telde study. PLoS ONE 2011, 6, e27208. [Google Scholar] [CrossRef] [PubMed]
- Elsammak, M.Y.; Al-Sharkaweey, R.M.; Fahmy, M.; Reda, A.A.; Farid, W.; Emara, A.; Hassan, H.; Kandil, M.H. Taq1B polymorphism of cholesteryl ester transfer protein (CETP) in Egyptian patients with metabolic syndrome. Diabetes Metab. Syndr. 2011, 5, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Lahiri, D.K.; Nurnberger, J.I., Jr. A rapid non-enzymatic method for the preparation of HMW DNA from blood for RFLP studies. Nucleic Acids Res. 1991, 19. [Google Scholar] [CrossRef]
- Cheverud, J.M. A simple correction for multiple comparisons in interval mapping genome scans. Heredity 2001, 87, 52–58. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cahua-Pablo, J.Á.; Cruz, M.; Méndez-Palacios, A.; Antúnez-Ortiz, D.L.; Vences-Velázquez, A.; Del Carmen Alarcón-Romero, L.; Parra, E.J.; Tello-Flores, V.A.; Leyva-Vázquez, M.A.; Valladares-Salgado, A.; et al. Polymorphisms in the LPL and CETP Genes and Haplotype in the ESR1 Gene Are Associated with Metabolic Syndrome in Women from Southwestern Mexico. Int. J. Mol. Sci. 2015, 16, 21539-21554. https://doi.org/10.3390/ijms160921539
Cahua-Pablo JÁ, Cruz M, Méndez-Palacios A, Antúnez-Ortiz DL, Vences-Velázquez A, Del Carmen Alarcón-Romero L, Parra EJ, Tello-Flores VA, Leyva-Vázquez MA, Valladares-Salgado A, et al. Polymorphisms in the LPL and CETP Genes and Haplotype in the ESR1 Gene Are Associated with Metabolic Syndrome in Women from Southwestern Mexico. International Journal of Molecular Sciences. 2015; 16(9):21539-21554. https://doi.org/10.3390/ijms160921539
Chicago/Turabian StyleCahua-Pablo, José Ángel, Miguel Cruz, Abigail Méndez-Palacios, Diana Lizzete Antúnez-Ortiz, Amalia Vences-Velázquez, Luz Del Carmen Alarcón-Romero, Esteban Juan Parra, Vianet Argelia Tello-Flores, Marco Antonio Leyva-Vázquez, Adán Valladares-Salgado, and et al. 2015. "Polymorphisms in the LPL and CETP Genes and Haplotype in the ESR1 Gene Are Associated with Metabolic Syndrome in Women from Southwestern Mexico" International Journal of Molecular Sciences 16, no. 9: 21539-21554. https://doi.org/10.3390/ijms160921539