Abstract
Overexpression of a constitutively active truncated form of OsCDPK1 (OEtr) in rice produced smaller seeds, but a double-stranded RNA gene-silenced form of OsCDPK1 (Ri) yielded larger seeds, suggesting that OsCDPK1 plays a functional role in rice seed development. In the study presented here, we propose a model in which OsCDPK1 plays key roles in negatively controlling the grain size, amylose content, and endosperm appearance, and also affects the physicochemical properties of the starch. The dehulled transgenic OEtr grains were smaller than the dehulled wild-type grains, and the OEtr endosperm was opaque and had a low amylose content and numerous small loosely packed polyhedral starch granules. However, the OEtr grain sizes and endosperm appearances were not affected by temperature, which ranged from low (22 °C) to high (31 °C) during the grain-filling phase. In contrast, the transgenic Ri grains were larger, had higher amylose content, and had more transparent endosperms filled with tightly packed polyhedral starch granules. This demonstrates that OsCDPK1 plays a novel functional role in starch biosynthesis during seed development and affects the transparent appearance of the endosperm. These results improve our understanding of the molecular mechanisms through which the grain-filling process occurs in rice.
1. Introduction
The quality of rice (Oryza sativa L.) grain is defined in terms of several main factors, including (i) eating and cooking qualities, and (ii) milling qualities and appearance [1]. Eating and cooking qualities are determined by the amylose content, amylopectin structure, gelatinization temperature, and pasting viscosity [2], and milling qualities and appearance correlate strongly with the transparency, flouriness, and chalkiness of the endosperm [3]. The filling and accumulation of starch granules in developing rice endosperms can accelerate at high temperatures, causing the starch in the endosperm cells to be packed loosely and the kernel to be chalky. Such grains crack easily during milling, yielding poor eating and cooking qualities [4,5,6].
It has been shown in many studies that chalky and less-transparent kernels contain more amylopectin and less amylose in the endosperm than do less-chalky and more-transparent kernels [7,8,9]. Eliminating chalkiness by regulating the amylopectin and amylose content ratios (by affecting biosynthesis) in the endosperm during the grain-filling phase is therefore a key way of improving grain quality. Two enzymes involved in amylose biosynthesis are ADP-glucose pyrophosphorylase and the Waxy gene-encoded granule-bound starch synthase I (GBSSI) [10,11]. The enzymes involved in the biosynthesis and modification of amylopectin are ADP-glucose pyrophosphorylase, soluble starch synthase, the starch-branching enzyme (BE), and the starch-debranching enzyme [10,11,12]. In higher plants, BE plays an essential role in amylopectin biosynthesis because it is the only enzyme that can add α-1,6-glucosidic linkages to polyglucans [13]. Three BE isoforms—BEI, BEIIa, and BEIIb—have been found in rice [10,14]. The rice mutant amylose-extender, which has a null mutation in BEIIb, has been found to alter the degree of polymerization (DP) of amylopectin, giving fewer short chains (DP ≤ 17) and more long chains (DP ≥ 18), the changes being related to the dose on the amylose-extender locus in the triploid endosperm cells. These results suggest that BEIIb might have critical effects on the amylopectin structure and the rheological properties of the starch [12]. Several floury endosperm rice mutants (flo1–flo7) have been isolated. Treating fertilized rice egg cells with the chemical mutagen N-methyl-N-nitrosourea yielded mutants flo1 and flo2, which had floury endosperms [15,16]. The flo2 mutant has been found, through map-based cloning, to be a member of the tetratricopeptide repeat-motif protein family. The gene mutation decreases the grain size and decreases the starch quality (by decreasing the amylose content) and also changes the fine structure of the amylopectin [17]. The flo3 mutant was produced through applying gamma-irradiation and ethyl methansulfonate treatment, and had a low 16 kDa globulin content in the endosperm [18]. The white-core floury endosperm mutants flo4 and flo5 were produced through T-DNA insertional mutagenesis and were found to have pyruvate orthophosphate dikinase B and starch synthase IIIa (SSIIIa) gene mutations, respectively [19,20]. The flo4 mutant endosperm had a low amylose content, suggesting that pyruvate orthophosphate dikinase B might play a role in regulating carbon metabolism during the grain filling process [19]. DP analysis of amylopectin in flo5 was performed, and the amounts of DP 6–8 and DP 16–20 in the mutant endosperm were found to be decreased but the amounts of DP 9–15 and DP 22–29 had increased, indicating that SSIIIa strongly affects the chain-length distribution of amylopectin biosynthesized in developing rice grains [20]. The floury endosperm mutant flo6 had a completely floury white endosperm, but the flo7 endosperm was floury and white only at the peripheries, and not in the interior [21,22]. Map-based cloning demonstrated that flo6 was an insertion mutation in the unknown function gene Os03g0686900 [21] and that flo7 was a deletion mutation in the unknown function gene Os10g0463800 [22], suggesting that these genes may play vital roles in starch biosynthesis and granule formation in the endosperm during the grain-filling process.
Calcium ions (Ca2+) are secondary messengers in plant cells, and are used in response to various environmental and developmental stimuli through temporal and spatial fluctuations of the cytosolic Ca2+ concentration [23,24]. Calcium-dependent protein kinases (CDPKs) are a major family of calcium sensors that have been characterized in various plant species. CDPKs are Ser/Thr protein kinases that are encoded by multigene families [25,26]. CDPKs have four functional domains: an N-terminal variable domain, a catalytic kinase domain, an autoinhibitory domain, and a calcium-binding EF-hands regulatory domain [27]. Under normal growth conditions (i.e., absence of Ca2+ signals), the autoinhibitory domain can interact with the kinase domain thereby inhibits kinase activity [24]. Deletion of the autoinhibitory domain and the Ca+2 binding domains from the coding region could bypass Ca2+ signal stimulation and resulted in produced a constitutively active enzyme of CDPKs [26,27]. It has been shown in many studies that CDPKs play important physiological roles in response to various environmental stresses and developmental processes [23,24,28,29,30]. Only in a few studies have CDPKs been shown to play a role in starch biosynthesis during the grain-filling process in rice. The rice SPK (which shares 79% of its amino acid sequence with OsCDPK1) has been found to encode a sucrose synthase kinase. Expression of antisense SPK in transgenic rice produced watery grains because large amounts of sucrose had accumulated in the endosperm due to low sucrose synthase activity, resulting in inefficient sucrose degradation [31]. This indicates that SPK may be a regulator in the starch biosynthesis pathway.
In the study presented here, OsCDPK1 was found to play pivotal roles in rice-seed development, in the physicochemical properties of the starch produced, and in the appearance of the endosperm. The pleiotropic effects on various agronomic traits in loss- and gain-of-function of OsCDPK1 transgenic plants are also characterized.
2. Results
2.1. Phenotypic Changes in Transgenic Rice Plants with Overexpressing or Silenced OsCDPK1
In previous studies, to understand the physiological roles of OsCDPK1, the gene-overexpression and gene-silencing approaches were implemented. For the truncated form of OsCDPK1 (OEtr), the coding sequences in which the autoinhibitory region and the calcium-binding domains had been removed were expressed under the control of maize ubiquitin gene promoter to generate the constitutively active form of the OEtr transgenic plants [32]. The transgenic plants in OEtrs yielded smaller seeds, whereas RNA-interference gene knockdown mutants (Ris) yielded larger seeds [32]. In this study, the various agronomic traits in the T4 transgenic lines of OEtrs (OEtr-1, -3 and -4) and Ris (Ri-1, -2 and -3) were analyzed. The results (Table 1) were consistent with our previous studies [32] and showed that the Ris (Ri-1, -2 and -3) lines had, on average, 7.1% and 10.8% increases in plant height and 1000-grain weight, respectively, compared to those of WT (wild type). However, both examined traits decreased in the OEtrs (OEtr-1, -3 and -4) lines compared with WT, with 5.8% and 21.1% reductions in plant height and 1000-grain weight, respectively. Furthermore, compared to WT, the heading date and growth duration were shorter in Ris, while OEtrs showed no significant difference. The average of the heading date of Ris (83.2 day) was 6 days shorter than that of WT (89.3 day) and the growth duration of Ris (104.8 day) was 10 days shorter than that of WT (115.2 day). The dehulled grain weight, starch content, and amylose content were higher in Ris and lower in OEtrs than those of WT; for example, the average weights of dehulled grain of Ris and OEtrs were 111.2% and 84.1%, respectively, of that of WT. The average percentage of starch content versus dehulled grain weight in WT, OEtrs, and Ris was 73.4%, 64.2%, and 78.1%, respectively. Moreover, the average amylose content in WT, OEtrs, and Ris was 23.2%, 15.1% and 26.2%, respectively.

Table 1.
Comparison of agronomic traits between WT and transgenic lines.
2.2. Ectopic Overexpression and Silencing of OsCDPK1 in Transgenic Rice Plants Yielded Opaque and Transparent Endosperms, Respectively
We examined the 55-day-old plant phenotypes in the T4 transgenic lines further, and the results were consistent with Table 1. The Ri-1, -2 and -3 plants were higher (mean value is 76.6 ± 6.18 cm) and the OEtr-1, -3 and -4 plants were shorter (52.4 ± 3.58 cm) than the WT plants (63.3 ± 4.63 cm) (Figure 1A,B). At the grain-filling stage, the WT and transgenic plants were transferred to the growth room at an optimal temperature of a cycle of 25 °C for 16 h light and 20 °C for 8 h dark. Fifteen seeds were randomly selected from each individual line and dehulled. We found that the dehulled Ri-1 grains were longer and that the OEtr-1 grains were shorter than the WT grains (Figure 1E,F). We also found that the dehulled OEtr-1 grains were all of the floury-kernel phenotype (Figure 1C,E).
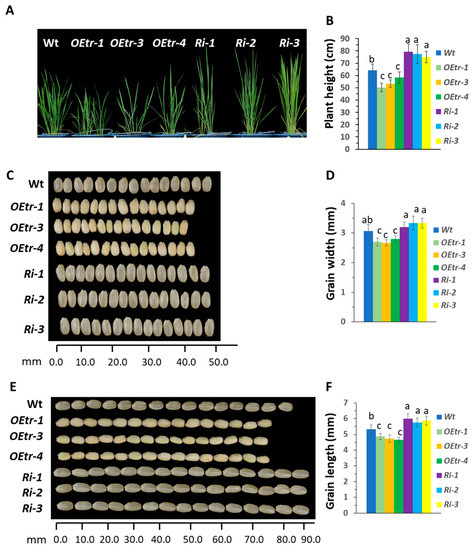
Figure 1.
Plant heights and grain morphologies in the WT, the OEtr-1, -3 and -4 (OEtrs), and the Ri-1, -2 and -3 (Ris) plants. After 5 d of flowering, rice plants were transferred to a growth chamber and grown at an optimal temperature (25 °C for 16 h light, 20 °C for 8 h dark). (A) Heights of the 55-day-old WT, OEtrs, and Ris plants. (B) Quantification of the plant heights. Each error bar is the standard error for 15 individual plants. (C) Endosperm appearances and grain widths for the WT, OEtrs, and Ris plants. Fifteen grains per line were positioned in a row and measured. (D) Quantification of the grain widths. Each error bar is the standard error for 50 individual grains. (E) Endosperm appearances and grain lengths for the WT, OEtrs, and Ris plants. Fifteen grains per line were positioned in a row and measured. (F) Quantification of the grain lengths. Each error bar is the standard deviation (n = 50). Different letters above the bars indicate significant differences, identified by performing ANOVAs (p < 0.01).
We determined whether the temperature affected the endosperm appearance in the transgenic lines by growing rice plants at relatively low temperatures (LT; 22 °C for 16 h light, 20 °C for 8 h dark) and at relatively high temperatures (HT; 31 °C for 16 h light, 28 °C for 8 h dark) during the grain-filling process. The mature seeds were collected from the WT, the Ri-1, Ri-2 and Ri-3 (Ris), and the OEtr-1, OEtr-3 and OEtr-4 (OEtrs) plants. As shown in Figure 2, under LT and HT conditions, all the dehulled OEtrs grains had smaller and floury endosperms (100% in the OEtr-1, OEtr-3 and OEtr-4 lines). In contrast, under LT condition, most of the grains in the WT and Ris lines displayed transparency phenotype; the ratios of chalky grains were 16.2% in the WT grains, and 6.0%, 6.7%, and 7.1% in the Ri-1, Ri-2 and Ri-3 grains, respectively (Figure 2A and Table 2). However, the higher temperature caused a significant increase in the WT and Ris lines’ endosperms of the chalky phenotype; the ratios of chalky grains were 63.6% in the WT grains, and 50.1%, 51.2% and 45.1% in the Ri-1, Ri-2 and Ri-3 grains, respectively (Figure 2A and Table 2). Illuminating the kernels with a backlight showed that the OEtrs grains all had opaque endosperms regardless of the temperature at which the plants were grown at LT or HT (Figure 2B). However, most of the WT and Ris grains displayed transparent endosperms under the LT condition (the Ris endosperms being more transparent than those of WT), the proportion of opaque endosperms increased to more than 50% in both the WT and Ris lines under HT growth condition (Figure 2B and Figure S1). Cross-sections of the endosperms (Figure 2C) indicated that the kernels in the WT and Ris displayed transparent phenotype under LT treatment and showed partial opaque phenotype under HT condition. However, in the OEtrs, all endosperms displayed completely floury appearance under both LT and HT conditions, regardless of the different temperature treatments used. These results suggest that OsCDPK1 affects rice endosperm appearance in a temperature-independent manner.
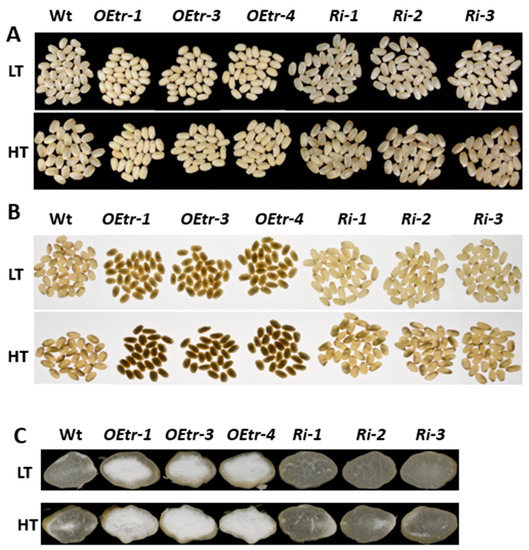
Figure 2.
Effects of temperature on the appearances of the WT, OEtrs, and Ris endosperms. After 5 d of flowering, rice plants were transferred to a growth chamber and grown at a lower temperature (22 °C for 16 h light, 20 °C for 8 h dark) or a higher temperature (31 °C for 16 h light, 28 °C for 8 h dark). (A,B) Seeds harvested from the plants grown at the lower and higher temperatures, respectively, illuminated using (A) normal lighting and (B) backlighting. (C) Cross-sections of the endosperms of the seeds from plants grown at either lower or higher temperature. LT: lower temperature; HT: higher temperature.
Table 2.
The ratios of chalky grains in wild type (TNG67), OEtr-1, and Ri-1 lines growth under lower temperature (LT; 22 °C for 16 h light, 20 °C for 8 h dark) or higher temperature (HT; 31 °C for 16 h light, 28 °C for 8 h dark) during the rice grain-filling process.
2.3. Effect of OsCDPK1 on Starch Granule Morphology in Rice Endosperms
Due to the similar transgenic plant phenotypes and grain morphology (Figure 1 and Figure 2), the OsCDPK1-overexpressing line, OEtr-1, and the OsCDPK1-silencing line, Ri-1, were therefore selected for further study. We examined the starch granule morphology in the WT, Ri-1, and OEtr-1 transgenic seeds by analyzing the endosperm cross-sections acquired from scanning electron microscopy. As shown in Figure 3, the three-dimensional structures of the starch granules in the endosperms were irregularly polygonal and polyhedral in all three grains types. The starch granules were large and tightly packed in the WT and Ri-1 endosperms but small and loosely packed in the OEtr-1 endosperm (Figure 3B), suggesting that OsCDPK1 affects the starch granule size and packing density in developing rice seed.

Figure 3.
Scanning electron microscopy images of the structures of the starch granules in the rice endosperms. (A) The central areas of the cross-sections of mature endosperms from the WT plants (top panel), OEtr-1 plants (middle panel), and Ri-1 plants (bottom panel) were acquired using a scanning electron microscope. (B) A zoomed-in image of the 20 μm photo of OEtr-1. Scale bar colors: blue as 1 mm, red as 100 μm, green as 40 μm, purple as 20 μm.
2.4. Effects of OsCDPK1 on Starch Properties and Gelatinization in the Endosperm
We examined the apparent amylose content in the endosperm by reacting 20 mg of each rice endosperm powder sample with 1 N NaOH to gelatinize the starch. The amylose content was measured using a colorimetric method using an I2/KI solution [12,33]. The OEtr-1 samples had less affinity than the other samples for iodine and were light purple, whereas the Ri-1 and WT samples were dark blue and light blue, respectively (Figure 4A and Figure S2). As described in Table 1, the average amylose content in WT, OEtrs, and Ris was 23.2%, 15.1% and 26.2%, respectively. Here, the apparent amylose content in WT, OEtr-1 and Ri-1 were examined in detail to dissect the roles of OsCDPK1 in starch biosynthesis. The absorption spectra of the I2/KI stained solutions and found strong absorbance between 480 and 720 nm for both the Ri-1 and WT samples, with maxima at 620 nm, but stronger absorbance was found for Ri-1 than for WT at the same wavelength (Figure 4B). OEtr-1 absorbance was weaker and decreased as the wavelength increased. We compared the results to the absorbances of potato amylose standards to allow the apparent amylose contents to be determined. The apparent amylose contents of the WT, OEtr-1, and Ri-1 samples were 23.35%, 14.74%, and 26.15%, respectively (Table S2). These results demonstrate that the Ri-1 seed endosperms had higher amylose content than the WT seed endosperms and that the OEtr-1 seed endosperms had lower amylose content than the WT seed endosperms. Because the WT, OEtr-1, and Ri-1 amylose contents were different, we investigated starch gelatinization at different urea concentrations (0–9 M). A 20-mg aliquot of a rice endosperm powder was mixed with 1 mL of urea solution, and the mixture was allowed to react for 24 h. The mixture was then centrifuged and the degree of gelatinization determined by measuring the sediment volume. Starch gelatinization started at urea concentrations of 3.0–4.0 M (Figure 4C and Figure S3). In 4.0 M urea, the Ri-1 sediment volume was 5.3% higher than the WT sediment volume, whereas the OEtr-1 sediment volume was 36.4% lower than the WT sediment volume (Figure 4D). These results indicate that OsCDPK1 affects the physicochemical properties of the starch in rice endosperms.
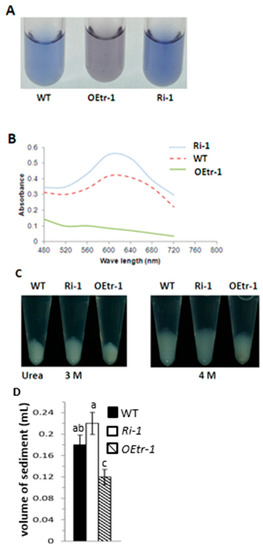
Figure 4.
Iodine-staining and gelatinization properties of the starch in the rice endosperms. A 20-mg aliquot of endosperm powder was treated with 1 N NaOH as described in Section 4. (A) Supernatants of the iodine-stained WT, OEtr-1, and Ri-1 samples. (B) Starch–iodine absorbance spectra of the supernatants. (C) Effects of using 3.0 and 4.0 M urea solutions on the gelatinization characteristics of the WT, OEtr-1, and Ri-1 endosperm starch. A 20-mg aliquot of rice powder was mixed in an Eppendorf tube with 1 mL of urea solution and the mixture was shaken for 24 h at 25 °C. The mixture was centrifuged, and the volume of the gelatinized starch sediment was measured. (D) Quantification of the gelatinization volume. Different letters above the bars indicate significant differences, identified by performing ANOVAs (p < 0.05). Each value is the mean ± SD of three independent measurements.
2.5. OsCDPK1 Expression Profiles in Developing Rice Seeds
Our results have demonstrated that OsCDPK1 affects rice seed development. It is necessary to track changes in OsCDPK1 gene expression during rice seed development. The OsCDPK1::GUS transgenic line was generated using a GUS (β-glucuronidase) reporter gene controlled by the OsCDPK1 promoter (−1706 to +301 bp, i.e., a total of 2007 bp upstream of the translational start site) containing the first intron (607 bp, in the 5′-untranslated region) (Figure 5A). As shown in Figure 5B, strong GUS staining was observed in the ovaries and anthers before flowering, but weaker staining was observed in the styles and lemma. At 1 DAF (days after flowering), strong GUS activity was found only in the ovaries and styles, weaker activity was found in the lemma, and no GUS activity was found in the anthers and stigma. Between 2 and 5 DAF, concentrated GUS staining was found in the rachilla and both ends of the developing seeds and weak staining was found in the lemma. The blue color gradually expanded from both ends toward the central parts of the developing seeds between 6 and 7 DAF, and at 8 DAF the entire seeds were thoroughly stained blue. Staining gradually decreased afterwards, but remained strong between 10 and 14 DAF, then decreased quickly after 14 DAF and had completely gone by 18 DAF. These results suggest that OsCDPK1 was expressed in a particular temporal and spatial way, predominantly in the middle stage of rice seed development.
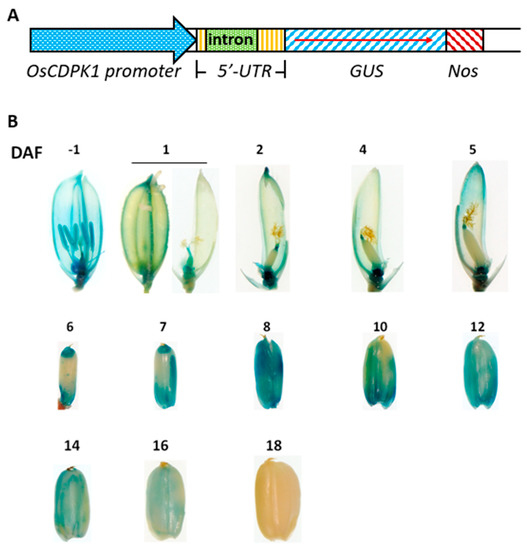
Figure 5.
Histochemical GUS activity staining in flowers and developing rice seeds. (A) Map of the OsCDPK1::GUS expression construct. (B) Rice spikelets were collected before flowering and 1–18 days after flowering (DAF). The lemma and palea were partially or completed removed from each spikelet or developing seed before staining. The stained spikelets or immature grains were preserved in 70% ethanol and photographed. −1 DAF means before flowering.
2.6. Effects of OsCDPK1 on the Levels of Starch-Biosynthesis-Related Genes in Developing Rice Seeds
We further investigated the roles of OsCDPK1 in rice-seed development by analyzing the expression patterns of 12 genes involved in starch biosynthesis. These genes were granule-bound starch synthase (OsGBSSI), starch synthase (OsSSI, OsSSIIa, OsSSIIb, OsSSIIc, OsSSIIIa, and OsSSIIIb), branching enzyme (OsBEI), ADP-glucose pyrophosphorylase large subunit (OsAGPLI, OsAGPLII, and OsAGPLIII), and a small subunit of ADP-glucose pyrophosphorylase (OsAGPSIIb). The developing WT, Ri-1, and OEtr-1 rice seeds at 5 and 12 DAF were analyzed. Total RNA was isolated from the dehulled embryo-less half seeds and subjected to quantitative RT-PCR. The relative expression levels of OsAGPLI, OsAGPSIIb, OsGBSSI, OsSSIIc, and OsSSIIIa were significantly up-regulated in Ri-1 and down-regulated in OEtr-1 at 12 DAF, but there was no significant difference at 5 DAF compared with the genes in WT plants (Figure 6). Expression of the seven other genes in the transgenic lines was not significantly different at 5 or 12 DAF from expression in the WT plants. These results suggest that OsCDPK1 might be involved in regulating starch-biosynthesis-related genes in the mid-development stage of the rice seed.
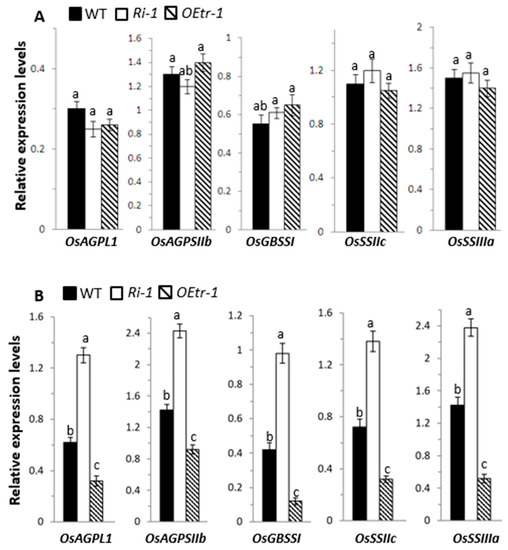
Figure 6.
Expression of starch-biosynthesis-related genes during seed development in the WT, Ri-1, and OEtr-1 plants. Total RNA was isolated from developing endosperms (A) 5 and (B) 12 DAF from the WT, Ri-1, and OEtr-1 plants, and subjected to RT-PCR analysis. The relative expression levels of each gene were normalized to the expression level of the internal control OsActin. Different letters above the bars indicate significant differences, identified by performing ANOVAs (p < 0.01). Each value is the mean ± SD of three independent measurements. OsAGPLI: ADP-glucose pyrophosphorylase large subunit I; OsAGPSIIb: ADP-glucose pyrophosphorylase small subunit IIb; OsGBSSI: granule-bound starch synthase I; OsSSIIc: starch synthase IIc; OsSSIIIa: starch synthase IIIa. Primer sets and gene accession numbers are listed in Table S1.
3. Discussion
The amylose content of endosperms is an important parameter determining the eating quality of rice, which is negatively related with stickiness but positively related to rice grain hardness [34,35]. The key enzymes (genes) involved in the starch (amylopectin and amylose) biosynthesis during rice grain filling are well known. However, the control mechanisms by which the amylose and amylopectin biosynthesis is orchestrated are still not fully understood. Our results indicate for the first time that the protein kinase OsCDPK1 is functionally negatively correlated with the amylose content, endosperm transparency, and seed size in developing rice seed.
The OEtr-1 and Ri-1 seeds had some distinct features compared with the WT seeds. For example, the OEtr-1 grains were smaller, had lower amylose contents, and had more floury endosperms than the WT grains, and the Ri-1 grains were larger, had higher amylose contents, and had more transparent endosperms than the WT grains (Figure 1, Figure 2, Figure 3 and Figure 4). This indicates that the OsCDPK1 function is closely associated with the rice endosperm starch properties. The OsCDPK1::GUS expression profile in the developing rice grains gradually increased immediately after flowering and reached a maximum between 7 and 14 DAF (Figure 5). The OsCDPK1::GUS expression timing in the developing rice seeds was similar to that found in a study by Ohdan et al. [36], in which 27 different genes involved in starch biosynthesis were examined during rice-seed development. In that study, all the genes had been differentially expressed before 15 DAF. In this study, the GUS staining was found throughout the endosperm at 12 and 14 DAF of developing seeds, and was more intensified in the interior region at 12 DAF (Figure S4), suggesting that OsCDPK1 first affects the expression of starch biosynthesis-related genes and later affects the starch composition and endosperm appearance. The starchy endosperm in the OEtr-1 grains had a low amylose content and an opaque floury appearance, and the starch granules were small and loosely packed (Figure 1, Figure 2 and Figure 3), indicating that the OsCDPK1 roles were closely associated with the structures and qualities of the starch granules during the grain filling process. Moreover, OsCDPK1::GUS staining was found throughout the developing endosperm at 7–16 DAF (Figure 5B and Figure S4). These results suggest that the expression of some starch-biosynthesis-related genes in the endosperm cells may be affected by OsCDPK1, followed by changing the amylose content and resulting in the opaque endosperm in the OEtr-1 grains.
Several mutations of rice genes involved in starch biosynthesis have been found to alter the structures and properties of the starch produced. For example, the SSIIIa mutation (flo5) was found to increase the amylose content, alter the amylopectin structure, and cause the endosperm to have a white core [20]. The waxy mutant (a mutation in GBSSI) produced an amylose-free, floury endosperm [11,12]. The amylose-extender mutation in BEIIb altered the fine structure of amylopectin and gave a floury endosperm [12]. A mutation in BEI (starch-branching enzyme I) caused the amylopectin fine structure to change but did not appear to affect the endosperm appearance [10]. Our data show that the expression of some starch-biosynthesis-related genes was affected by OsCDPK1, similar to the results of previous studies. During the middle phase (12 DAF) of endosperm development, examples of genes affected were OsAGPLI, OsAGPSIIb, OsGBSSI, OsSSIIc, and OsSSIIIa, which were significantly up-regulated in the Ri-1 and down-regulated in OEtr-1 (Figure 6). Changes in the expression of these genes caused the OEtr-1 endosperm to have a lower amylose content and a floury appearance, whereas the Ri-1 endosperm had a high amylose content and was more transparent (Figure 1, Figure 2 and Figure 4 and Table 1 and Table 2). Calcium ions (Ca2+) are secondary messengers in plant cells, with plant CDPK being a sensor to relay calcium signals via binding with calcium via the calcium-binding domains. We therefore suggest that OsCDPK1 acts as an upstream regulator that is closely associated with starch biosynthesis. Some regulators have been found to regulate the expression of genes encoding key starch-biosynthesis enzymes, and null mutations in these regulators usually alter the amylopectin fine structure, the starch composition, the starch granule morphology and size, and the endosperm appearance. For instance, some starch-biosynthesis-related genes were found to be affected in five independent mutants, flo2 (a tetratricopeptide repeat motif protein) [17], flo4 (pyruvate orthophosphate dikinase B) [19], flo7 (an unknown protein) [22], osbzip58 (a bZIP transcription factor) [37], and osbt1 (an ADP-glucose transporter) [38]. All these effects decreased the amylose content and changed the amylopectin composition. The endosperm appearance was affected differently in different parts; e.g., flo2 had endosperm with a floury kernel, flo4 and osbzip58 and osbt1 gave white-core phenotypes, and chalkiness was only found in the peripheral endosperm of flo7. Similarly, the low amylose content of the OEtr-1 endosperm was expressed in a floury morphology (Figure 1 and Figure 4). The results of previous studies and this study together indicate that a low-amylose content of rice endosperm gives a chalky or floury phenotype, suggesting that the amount of amylose present is an important factor affecting the quality and appearance of the starchy endosperm. This raises the question of whether rice can be engineered to have an endosperm with a high amylose content and, therefore, a transparent appearance. Here, we have provided direct evidence that silencing OsCDPK1 (Ri-1) increases the amylose content of the endosperm, making the starchy grain more transparent (Figure 1 and Figure 4 and Table 1 and Table 2). These results will be useful in developing rice-breeding strategies aimed at maintaining (or even improving) grain quality in rice to cope with global warming. The transgenic lines OEtrs and Ris could also be ideal materials for investigating the mechanisms that control rice seed size and starch biosynthesis.
Temperature also strongly affects amylose synthesis during rice grain development. In previous studies, a lower temperature increased the expression of the waxy gene and protein and increased the amylose content in developing rice endosperms [39]; a higher temperature had the opposite effects [39,40]. Moreover, during the rice grain-filling process, a temperature higher than the optimum usually causes impaired starch accumulation, resulting in loosely packed starch granules with small air spaces between them, giving high proportions of opaque chalky or floury grains. These effects decrease the market value because the rice will have poor milling qualities (being easily broken), poor cooking and eating qualities, and a poor appearance [41,42]. It has recently been found that a high temperature (33 °C for 12 h light, 28 °C for 12 h dark) also induced the expression of three α-amylase genes—Amy1A, Amy3C, and Amy3D—in developing endosperms, causing the grains to be chalky, probably because of starch degradation and the accumulation of soluble sugars in the endosperm [43]. In contrast, we found that the Ri-1 endosperm was more transparent at both a low temperature (22 °C for 16 h light, 20 °C for 8 h dark) and a high temperature (31 °C for 16 h light, 28 °C for 8 h dark) than that of the WT plant during the grain-filling process (Figure 2 and Table 2). Our results will be useful in developing rice-breeding strategies aimed at maintaining (or even improving) grain quality in rice selected to cope with global warming. Our results also improve our understanding of the molecular mechanisms involved in amylose biosynthesis.
It has been found in several studies that chalky or floury endosperms might be caused by loosely packed small, round starch granules formed in developing rice seeds [9,19,20,38]. The scanning electron microscopy images indicate that the WT and Ri-1 endosperms contained closely packed polyhedral starch granules but that the white core of the OEtr-1 endosperm contained loosely packed small starch granules. The starch granules in the OEtr-1 endosperm were polyhedral (like in the WT and Ri-1 endosperm) rather than small and round as in most chalky or floury mutants (Figure 3B). It is, therefore, likely that OsCDPK1 plays a role in starch biosynthesis and negatively affects the sizes but not the shapes of the starch granules.
We previously found that OsCDPK1 inhibits the feedback of GA biosynthesis through down-regulating GA3ox2 and GA20ox1 [32]. In this study, we put forward a model in which OsCDPK1 plays key roles in negatively controlling the grain size, amylose content, and endosperm appearance, and also affects the physicochemical properties of the starch (Figure 7). Milled OsCDPK1-gene-silenced Ri-1 grains were larger than WT grains, and there were numerous densely packed polyhedral starch granules accompanied by a high amylose content and a transparent endosperm. In contrast, the OEtr-1 grains were smaller and contained loosely packed small, but still polyhedral, starch granules. The OEtr-1 grains had lower amylose content and opaque white-cored endosperms. Notably, the phenotypes of the grain size and the floury endosperm in OEtr-1, -2, and -3 were consistent and were unaffected by temperature during the grain-filling process (Figure 1 and Figure 2). OsCDPK1 therefore plays pleiotropic roles in rice seed development. Our results indicate that Ri-1 and OEtr-1 could be ideal materials for investigating the mechanisms controlling rice seed size and starch biosynthesis, and could also be valuable reference samples for rice breeding aimed at simultaneously improving grain yield and quality.
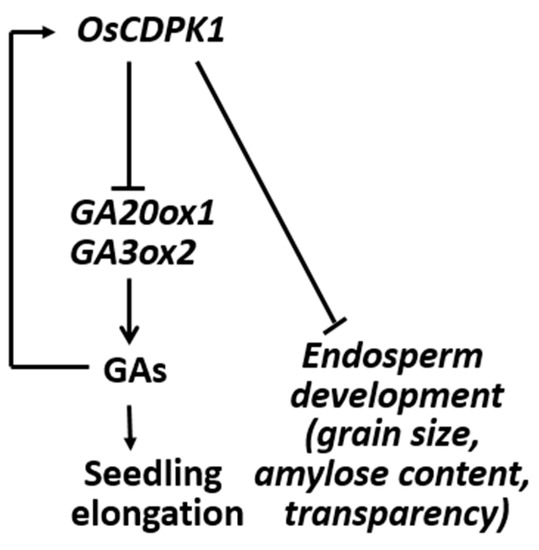
Figure 7.
Proposed roles of OsCDPK1 in the interconnecting GA biosynthesis and signaling pathways and endosperm developmental processes. The model is described in detail in the text. The arrows indicate activation and the blunt ends indicate inhibition.
4. Materials and Methods
4.1. Plant Materials
In our previous studies [32], 8 independent T1 transgenic lines that carried a single transgene in rice were subjected to ectopic overexpression of a constitutively active truncated form of OsCDPK1 (OEtr) in rice seedlings, which all yielded a semi-dwarf phenotype that produced small seeds. By contrast, 5 independent T1 transgenic plants exhibiting a single copy of transgene were subject to OsCDPK1 gene silencing (Ri) by RNA interference, which all gave a slender-like phenotype during seedling development and subsequently produced large seeds. According to the consistency of the phenotypic traits in OEtr lines of which all displayed semi-dwarf phenotype and produced small seeds. In Ri lines, which all gave a slender-like phenotype and produced large seeds [32]. Therefore, in this study, the homozygous transgenes of the T4 transgenic plants from OEtr-1, OEtr-3, and OEtr-4, and Ri-1, Ri-2, and Ri-3 were selected to explore the roles of OsCDPK1 in rice grain development.
4.2. Callus Induction
Rice Oryza sativa L. cv. Tainung 67 was used in the study. Immature seeds were de-hulled, sterilized with 2.4% NaOCl for 30 min, then washed thoroughly with sterile water. The seeds were then incubated on N6 agar medium [44] containing 10 µM 2,4-D to induce calli formation [45]. After about 30 days, the calli derived from the scutella were transferred to the fresh N6 agar medium containing 2,4-D for another 15 days and were subjected to Agrobacterium-mediated gene transformation [45].
4.3. Primers
The nucleotide sequences of all the primers used in the real-time polymerase chain reaction (qRT-PCR) amplification are shown in Table S1.
4.4. Construction of OsCDPK1::GUS Expression Vectors
The OsCDPK1::GUS expression vector was constructed by amplifying a 2007 bp DNA fragment containing the OsCDPK1 promoter and its 5′-untranslated region (which contained a 607 bp intron) (Figure 1A) by PCR using the forward primer OsCDPK1-5P (5′-ATACTGCAGTGGTCTTATT AGGTAAGGCC-3′) and the reverse primer OsCDPK1-3B (5′-ATAGGATCC TCCAAGAACTCCTTATGCAA-3′). The DNA fragment was cleaved using PstI and BamHI, and then cloned into vector pBX-2 as described previously [45]. The OsCDPK1::GUS construct was linearized by digesting it with PstI, then inserted into the PstI site of the pSMY1H binary vector [45], then it was subjected to Agrobacterium-mediated gene transformation.
4.5. Plant Transformation
Recombinant binary plasmids were introduced into Agrobacterium tumefaciens strain EHA101 by electroporation, and rice calli were transformed as described previously [45].
4.6. Histochemical Staining of GUS Activity in Developing Rice Grains
Rice spikelets collected before flowering and 1–18 DAF were subjected to GUS activity staining to assess OsCDPK1 gene expression profiles in developing rice grains. The lemma and palea were partially or completely removed from each spikelet (developing seed) before the staining process, and the spikelets were then incubated in a 1-mM 5-bromo-4-chloro-3-indolyl β-D-glucuronide solution (in 100 mM sodium phosphate containing 10 mM EDTA, 0.5 mM potassium ferrocyanide, 0.5 mM potassium ferricyanide, and 0.1% Triton X-100, at pH 7.0) at 37 °C in the dark for 4 h. The stained spikelets or immature grains were then preserved in 70% ethanol and rinsed with water before being photographed.
4.7. Quantitative RT-PCR
Developing seeds were collected from wild-type (WT), Ri-1, and OEtr-1 at 5 and 12 DAF, respectively. Total RNA was isolated from developing endosperms by using TRIzol reagent (Invitrogen, Carlsbad, CA, USA), and DNA contamination was then removed using a TURBO DNA-free kit (Ambion, Foster City, CA, USA). A 5-µg aliquot of the total RNA was used to synthesize first strand cDNA using M-MuLV reverse transcriptase (New England Biolabs, Ipswich, MA, USA) and oligo (dT) primer. Quantitative RT-PCR was performed using an Eco Real-Time PCR System (Illumina, San Diego, CA, USA) following the manufacturer’s instructions. Gene-specific primer sets (Table S1) localized at the 3′-untranslated regions for each gene examined were selected to allow assessment of the extent to which the starch-biosynthesis-related genes in WT, OEtr-1, and Ri-1 were expressed. The relative expression levels were normalized to expression in the internal control, OsActin 1. All reactions were performed in triplicate.
4.8. Analysis of the Gelatinization Properties of the Starch
A 20-mg aliquot of rice endosperm powder derived from de-embryonic seeds was mixed with 1 mL of urea solution at a concentration of between 0 and 9 M, and the mixture was shaken vigorously for 24 h at room temperature [12]. All the tested mixtures were then centrifuged at 25,000× g for 60 min (Eppendorf, model 5427R, North Ryde, Australia) at a same time, and the volume of the gelatinized starch sediment was measured by the volume scale on the Eppendorf tube.
4.9. Apparent Amylose Content Analysis
A 20-mg aliquot of rice endosperm powder was gelatinized by adding 2 mL of 1 N NaOH and incubating the mixture at 25 °C for 24 h. Then, 4 mL of 1 N CH3COOH was added, the mixture was mixed well, and 4 mL H2O was added. A 0.8-mL aliquot of the solution was mixed with 0.2 mL I2/KI (0.2%/2%), then 4 mL H2O was added. The apparent amylose content was measured using the colorimetric method described by [33]. Absorbance at 620 nm was measured, and the apparent amylose content was determined by comparing the absorbance to a calibration curve prepared using potato amylose standards.
4.10. Scanning Electron Microscopy
Dehusked rice grains were cut transversely and analyzed using a scanning electron microscope (Quanta 200; FEI, Hillsboro, OR, USA) following the manufacturer’s instructions.
5. Conclusions
Our results demonstrated that OsCDPK1 plays key roles in negatively controlling the grain size, amylose content, and endosperm appearance, and also affects the physicochemical properties of the starch. Milled OsCDPK1-gene-silenced Ri-1 grains were larger than WT grains, and there were numerous densely packed polyhedral starch granules accompanied by a high amylose content and a transparent endosperm. In contrast, the OEtr-1 grains were smaller and contained loosely packed small, but still polyhedral, starch granules. The OEtr-1 grains had lower amylose content and opaque white-cored endosperms. Moreover, the grain phenotypes in OEtrs were unaffected by temperature during the grain-filling process. OsCDPK1 therefore plays pleiotropic roles in rice reproductive developmental processes, in a negative sense. Our results indicate that Ri-1 and OEtr-1 could be ideal materials for investigating the mechanisms that control rice seed size and starch biosynthesis, and for rice breeding to improve grain yield and quality.
Supplementary Materials
The following are available online at http://www.mdpi.com/1422-0067/19/10/3247/s1.
Author Contributions
Conceptualization: S.-L.H.; Data curation: J.-Z.J. and S.-L.H.; Investigation: J.-Z.J., C.-H.K., B.-H.C. and M.-K.C.; Methodology: C.-S.L.; Supervision: S.-L.H.; Writing—original draft: S.-L.H.
Funding
This work was supported by grants from the Ministry of Science and Technology of the Republic of China (grant no. MOST 105-2313-B-415-009-, MOST 106-2313-B-415-006- and MOST 107-2313-B-415-006-).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Rabiei, B.; Valizadeh, M.; Ghareyazie, B.; Moghaddam, M.; Ali, A.J. Identification of QTLs for rice grain size and shape of Iranian cultivars using SSR markers. Euphytica 2004, 137, 325–332. [Google Scholar] [CrossRef]
- Bao, J.; Jin, L.; Xiao, P.; Shen, S.; Sun, M.; Corke, H. Starch physicochemical properties and their associations with microsatellite alleles of starch synthesizing genes in a rice RIL population. J. Agric. Food Chem. 2008, 56, 1589–1594. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, M.A.; Lisle, A.J.; Martin, M. Chalky and translucent rice grains differ in starch composition and structure and cooking properties. Cereal Chem. 2000, 77, 627–632. [Google Scholar]
- Jiang, H.; Dian, W.; Wu, P. Effect of high temperature on fine structure of amylopectin in rice endosperm by reducing the activity of the starch branching enzyme. Phytochemistry 2003, 63, 53–59. [Google Scholar] [CrossRef]
- Morita, S.; Shiratsuchi, H.; Takanashi, J.; Fujita, K. Effect of high temperature on grain ripening in rice plants: Analysis of the effects of high night and high day temperatures applied to the panicle and other parts of the plant. Jpn. J. Crop Sci. 2004, 73, 77–83. [Google Scholar] [CrossRef]
- Counce, P.A.; Bryant, R.J.; Bergman, C.J.; Bautista, R.C.; Wang, Y.J.; Siebenmorgen, T.J.; Moldenhauer, K.A.; Meullenet, J.F.C. Rice milling quality, grain dimensions, and starch branching as affected by high night temperatures. Cereal Chem. 2005, 82, 645–648. [Google Scholar] [CrossRef]
- Patindol, J.; Wang, Y.J. Fine structures and physicochemical properties of starches from chalky and translucent rice kernels. J. Agric. Food Chem. 2003, 51, 2777–2784. [Google Scholar] [CrossRef] [PubMed]
- Yamakawa, H.; Hirose, T.; Kuroda, M.; Yamaguchi, T. Comprehensive expression profiling of rice grain filling-related genes under high temperature using DNA microarray. Plant Physiol. 2007, 144, 258–277. [Google Scholar] [CrossRef] [PubMed]
- Fu, F.F.; Xue, H.W. Coexpression analysis identifies rice starch regulator1, a rice AP2/EREBP family transcription factor, as a novel rice starch biosynthesis regulator. Plant Physiol. 2010, 154, 927–938. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, K.; Kimura, K.; Arai, Y.; Kawasaki, T.; Shimada, H.; Baba, T. Starch branching enzymes from immature rice seeds. J. Biochem. 1992, 112, 643–651. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.M.; Denyer, K.; Martin, C. The synthesis of the starch granule. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1997, 48, 67–87. [Google Scholar] [CrossRef] [PubMed]
- Nishi, A.; Nakamura, Y.; Tanaka, N.; Satoh, H. Biochemical and genetic analysis of the effects of amylose-extender mutation in rice endosperm. Plant Physiol. 2001, 127, 459–472. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.K.; Preiss, J. Starch branching enzymes from maize. Plant Physiol. 1985, 79, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, Y.; Takeichi, T.; Kawaguchi, K.; Yamanouchi, H. Purification of two forms of starch branching enzyme (Q-enzyme) from developing rice endosperm. Physiol. Plant. 1992, 84, 329–335. [Google Scholar] [CrossRef]
- Satoh, H.; Omura, T. New endosperm mutations induced by chemical mutagens in rice, Oryza sativa L. Jpn. J. Breed. 1981, 31, 316–326. [Google Scholar] [CrossRef]
- Kaushik, R.P.; Khush, G.S. Genetic analysis of endosperm mutants in rice Oryza sativa L. Theor. Appl. Genet. 1991, 83, 146–152. [Google Scholar] [CrossRef] [PubMed]
- She, K.C.; Kusano, H.; Koizumi, K.; Yamakawa, H.; Hakatae, M.; Imamura, T.; Fukuda, M.; Naito, N.; Tsurumaki, Y.; Yaeshima, M.; et al. A novel factor FLOURY ENDOSPERM 2 is involved in regulation of rice grain size and starch quality. Plant Cell 2010, 22, 3280–3294. [Google Scholar] [CrossRef] [PubMed]
- Nishio, T.; Iida, S. Mutant having a low content of 16-kDa allergenic protein in rice (Oryza sativa L. ). Theor. Appl. Genet. 1993, 86, 317–321. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.G.; Park, S.; Matsuoka, M.; An, G. White-core endosperm floury endosperm-4 in rice is generated by knockout mutations in the C4-type pyruvate orthophosphate dikinase gene (OsPPDKB). Plant J. 2005, 42, 901–911. [Google Scholar] [CrossRef] [PubMed]
- Ryoo, N.; Yu, C.; Park, C.S.; Baik, M.Y.; Park, I.M.; Cho, M.H.; Bhoo, S.H.; An, G.; Hahn, T.R.; Jeon, J.S. Knockout of a starch synthase gene OsSSIIIa/Flo5 causes white-core floury endosperm in rice (Oryza sativa L.). Plant Cell Rep. 2007, 26, 1083–1095. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.; Wang, Y.H.; Liu, F.; Ren, Y.L.; Zhou, K.N.; Lv, J.; Zheng, M.; Zhao, S.L.; Zhang, L.; Wang, C.M.; et al. FLOURY ENDOSPERM6 encodes a CBM48 domain-containing protein involved in compound granule formation and starch synthesis in rice endosperm. Plant J. 2014, 77, 917–930. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Ren, Y.L.; Lu, B.Y.; Yang, C.Y.; Feng, Z.M.; Liu, Z.; Chen, J.; Ma, W.W.; Wang, Y.H.; Liu, Y.S.; et al. FLOURY ENDOSPERM7 encodes a regulator of starch synthesis and amyloplast development essential for peripheral endosperm development in rice. J. Exp. Bot. 2016, 67, 633–647. [Google Scholar] [CrossRef] [PubMed]
- Sanders, D.; Pelloux, J.; Brownlee, C.; Harper, J.F. Calcium at the crossroads of signaling. Plant Cell 2002, 14, S401–S417. [Google Scholar] [CrossRef] [PubMed]
- Harper, J.F.; Breton, G.; Harmon, A. Decoding Ca2+ signals through plant protein kinases. Annu. Rev. Plant Biol. 2004, 55, 263–288. [Google Scholar] [CrossRef] [PubMed]
- Mori, I.C.; Murata, Y.; Yang, Y.; Munemasa, S.; Wang, Y.F.; Andreoli, S.; Tiriac, H.; Alonso, J.M.; Harper, J.F.; Ecker, J.R.; et al. CDPKs CPK6 and CPK3 function in ABA regulation of guard cell S-type anion- and Ca2+-permeable channels and stomatal closure. PLoS Biol. 2006, 4, e327. [Google Scholar] [CrossRef] [PubMed]
- Sheen, J. Ca2+-dependent protein kinases and stress signal transduction in plants. Science 1996, 274, 1900–1902. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, A.A.; Romeis, T.; Jones, J.D. CDPK-mediated signalling pathways: Specificity and cross-talk. J. Exp. Bot. 2004, 55, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Romeis, T.; Ludwig, A.A.; Martin, R.; Jones, J.D.G. Calcium-dependent protein kinases play an essential role in a plant defence response. EMBO J. 2001, 20, 5556–5567. [Google Scholar] [CrossRef] [PubMed]
- Abbasi, F.; Onodera, H.; Toki, S.; Tanaka, H.; Komatsu, S. OsCDPK13, a calcium-dependent protein kinase gene from rice, is induced by cold and gibberellin in rice leaf sheath. Plant Mol. Biol. 2004, 55, 541–552. [Google Scholar] [CrossRef] [PubMed]
- Wan, B.; Lin, Y.; Mou, T. Expression of rice Ca2+-dependent protein kinases (CDPKs) genes under different environmental stresses. FEBS Lett. 2007, 581, 1179–1189. [Google Scholar] [CrossRef] [PubMed]
- Asano, T.; Kunieda, N.; Omura, Y.; Ibe, H.; Kawasaki, T.; Takano, M.; Sato, M.; Furuhashi, H.; Mujin, T.; Takaiwa, F.; et al. Rice SPK, a calmodulin-like domain protein kinase, is required for storage product accumulation during seed development: Phosphorylation of sucrose synthase is a possible factor. Plant Cell 2002, 14, 619–628. [Google Scholar] [CrossRef] [PubMed]
- Ho, S.L.; Huang, L.F.; Lu, C.A.; He, S.L.; Wang, C.C.; Yu, S.P.; Chen, J.; Yu, S.M. Sugar starvation- and GA-inducible calcium-dependent protein kinase 1 feedback regulates GA biosynthesis and activates a 14-3-3 protein to confer drought tolerance in rice seedlings. Plant Mol. Biol. 2013, 81, 347–361. [Google Scholar] [CrossRef] [PubMed]
- Juliano, B.O. A simplified assay for milled-rice amylose. Cereal Sci. Today 1971, 16, 334–340. [Google Scholar]
- Cameron, D.K.; Wang, Y.J. A better understanding of factors that affect the hardness and stickiness of long-grain rice. Cereal Chem. 2005, 82, 113–119. [Google Scholar] [CrossRef]
- Li, H.; Prakash, S.; Nicholson, T.M.; Fitzgerald, M.A.; Gilbert, R.G. The importance of amylose and amylopectin fine structure for textural properties of cooked rice grains. Food Chem. 2016, 196, 702–711. [Google Scholar] [CrossRef] [PubMed]
- Ohdan, T.; Francisco, P.B., Jr.; Sawada, T.; Hirose, T.; Terao, T.; Satoh, H.; Nakamura, Y. Expression profiling of genes involved in starch synthesis in sink and source organs of rice. J. Exp. Bot. 2005, 56, 3229–3244. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.C.; Xu, H.; Zhu, Y.; Liu, Q.Q.; Ca, X.L. OsbZIP58, a basic leucine zipper transcription factor, regulates starch biosynthesis in rice endosperm. J. Exp. Bot. 2013, 64, 3453–3466. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Wei, X.; Ren, Y.; Qiu, J.; Jiao, G.; Guo, X.; Tang, S.; Wan, J.; Hu, P. OsBT1 encodes an ADP-glucose transporter involved in starch synthesis and compound granule formation in rice endosperm. Sci. Rep. 2017, 7, 40124. [Google Scholar] [CrossRef] [PubMed]
- Sano, Y.; Maekawa, M.; Kikuchi, H. Temperature effects on the Wx protein level and amylose content in the endosperm of rice. J. Hered. 1985, 6, 221–222. [Google Scholar] [CrossRef]
- Asaoka, M.; Okuno, K.; Sugimoto, Y.; Kawakami, J.; Fuwa, H. Effect of environmental temperature during development of rice plants on some properties of endosperm starch. Starch 1984, 36, 189–193. [Google Scholar] [CrossRef]
- Lyman, N.B.; Jagadish, S.V.K.; Nalley, L.L.; Dixon, B.L.; Siebenmorgen, T. Neglecting rice milling yield and quality underestimates economic losses from high-temperature stress. PLoS ONE 2013, 8, e72157. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.; Yin, X.; Struik, P.C.; Solis, C.; Xie, F.; Schmidt, R.C.; Huang, M.; Zou, Y.; Ye, C.; Jagadish, S.V.K. High day- and night-time temperatures affect grain growth dynamics in contrasting rice genotypes. J. Exp. Bot. 2017, 68, 5233–5245. [Google Scholar] [CrossRef] [PubMed]
- Nakata, M.; Fukamatsu, Y.; Miyashita, T.; Hakata, M.; Kimura, R.; Nakata, Y.; Kuroda, M.; Yamaguchi, T.; Yamakawa, H. High temperature-induced expression of rice α-amylases in developing endosperm produces chalky grains. Front. Plant Sci. 2017, 8, 2089. [Google Scholar] [CrossRef] [PubMed]
- Chu, C.C.; Wang, C.C.; Sun, C.S.; Hsu, C.; Yin, K.C.; Chu, C.Y.; Bi, F.Y. Establishment of an efficient medium for another culture of rice through comparative experiments on the nitrogen sources. Sci. Sin. 1975, 5, 659–668. [Google Scholar]
- Ho, S.L.; Tong, W.F.; Yu, S.M. Multiple mode regulation of a cysteine proteinase gene expression in rice. Plant Physiol. 2000, 122, 57–66. [Google Scholar] [CrossRef] [PubMed]
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).