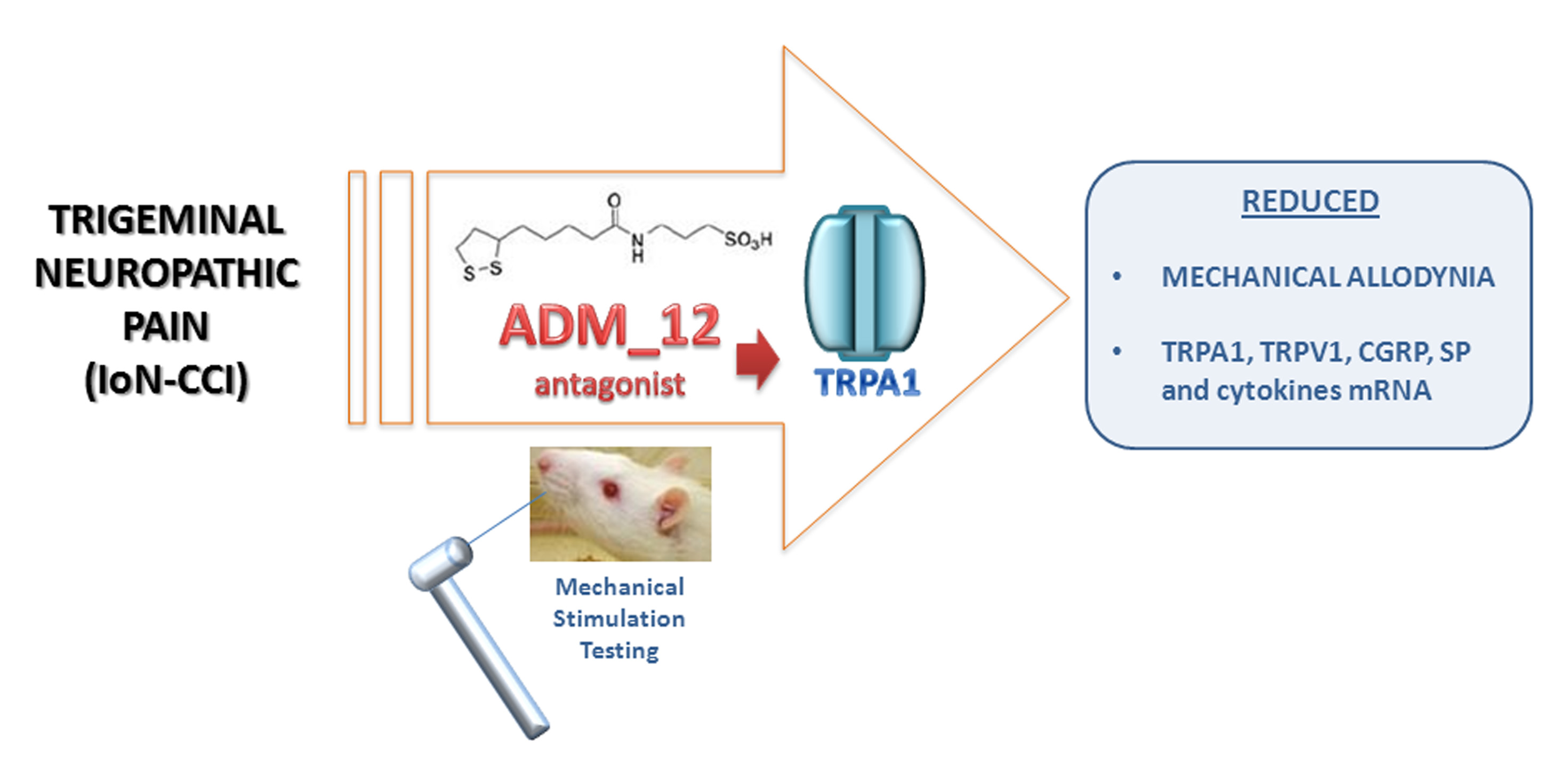
Antagonism of Transient Receptor Potential Ankyrin Type-1 Channels as a Potential Target for the Treatment of Trigeminal Neuropathic Pain: Study in an Animal Model

 ,
,
Abstract
:
1. Introduction
2. Results
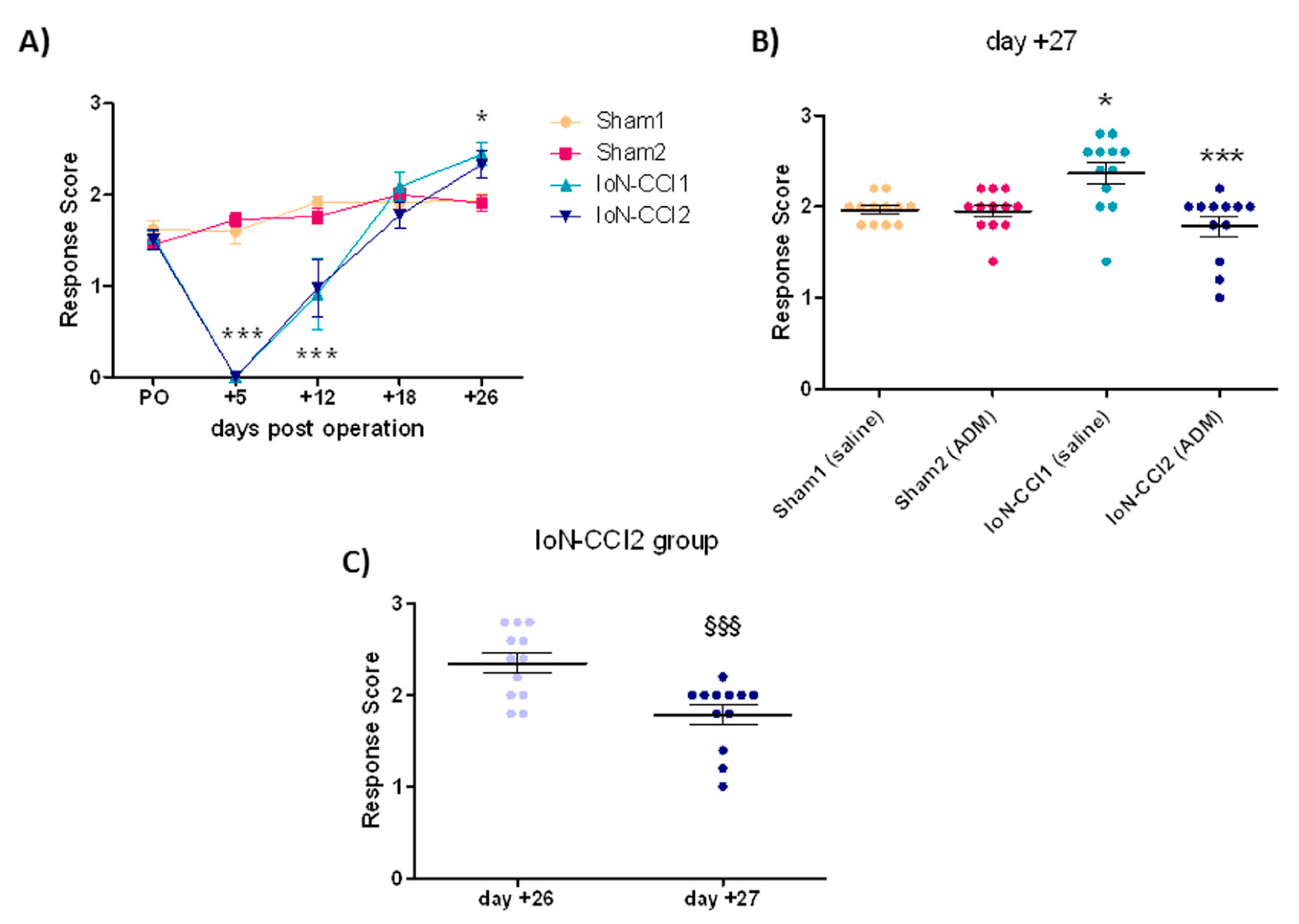
2.1. ADM_12 Effect on Behavioral Response
2.2. ADM_12 Effect on Gene Expression
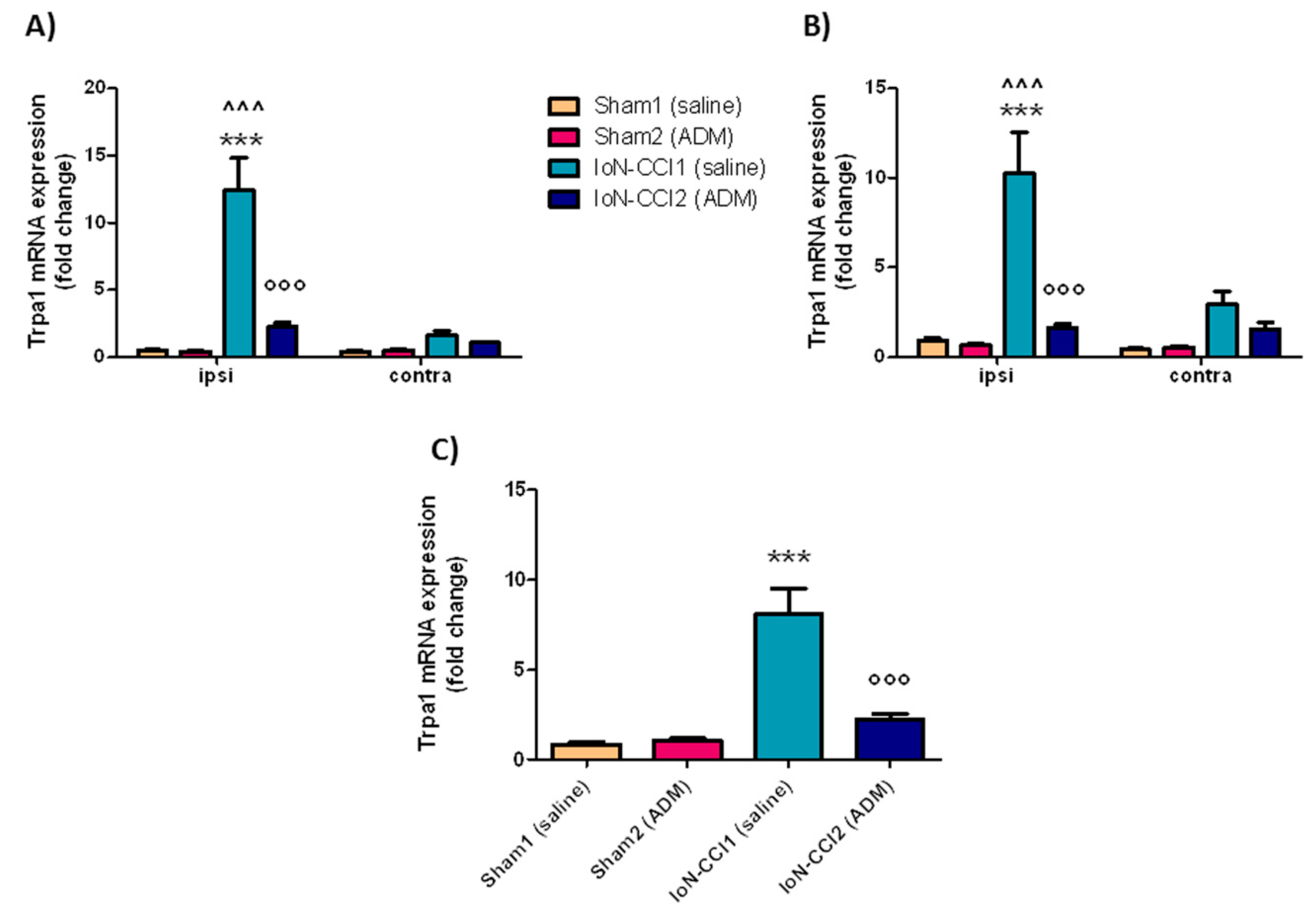
2.2.1. Trpa1 mRNA Expression
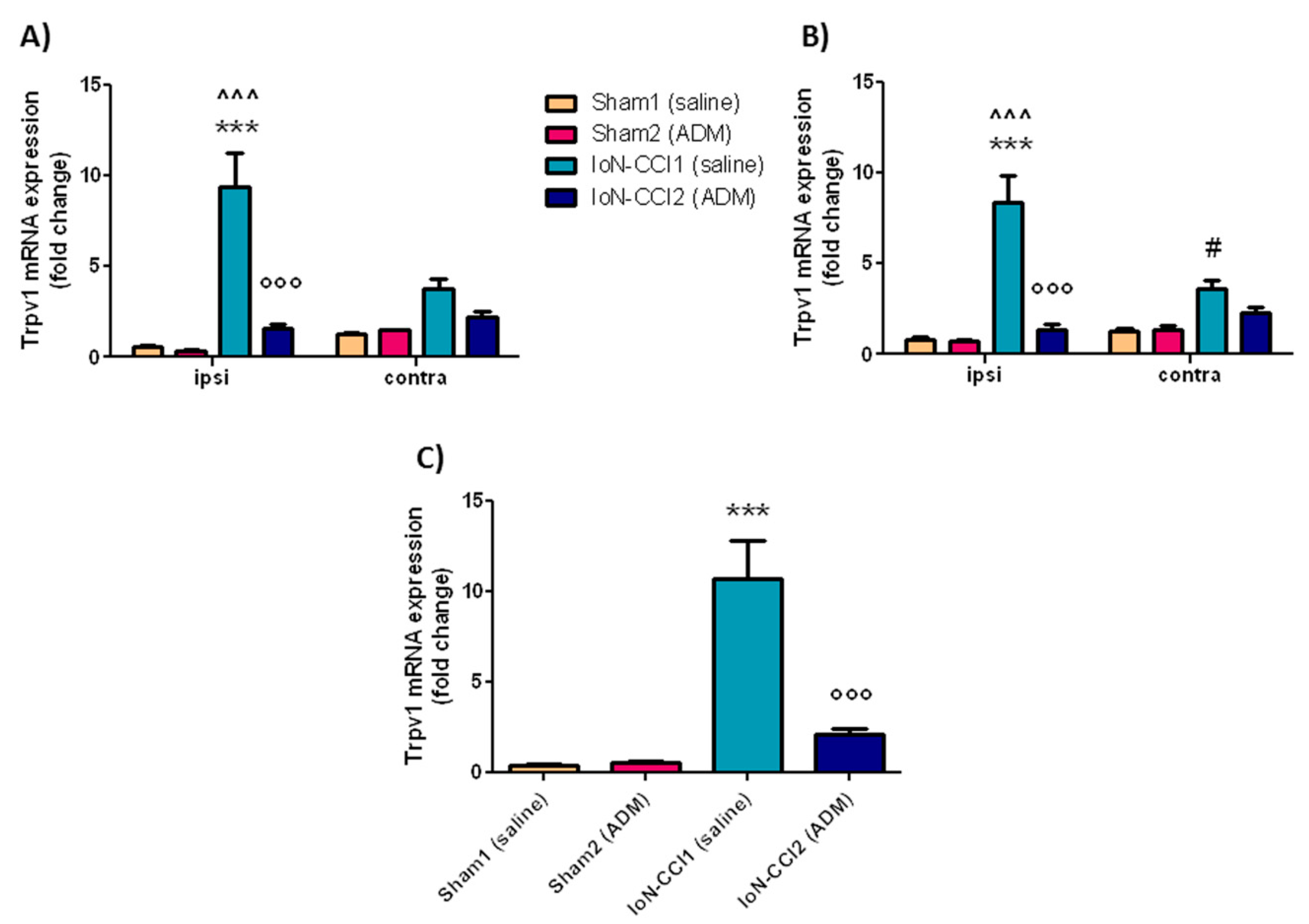
2.2.2. Trpv1 mRNA Expression
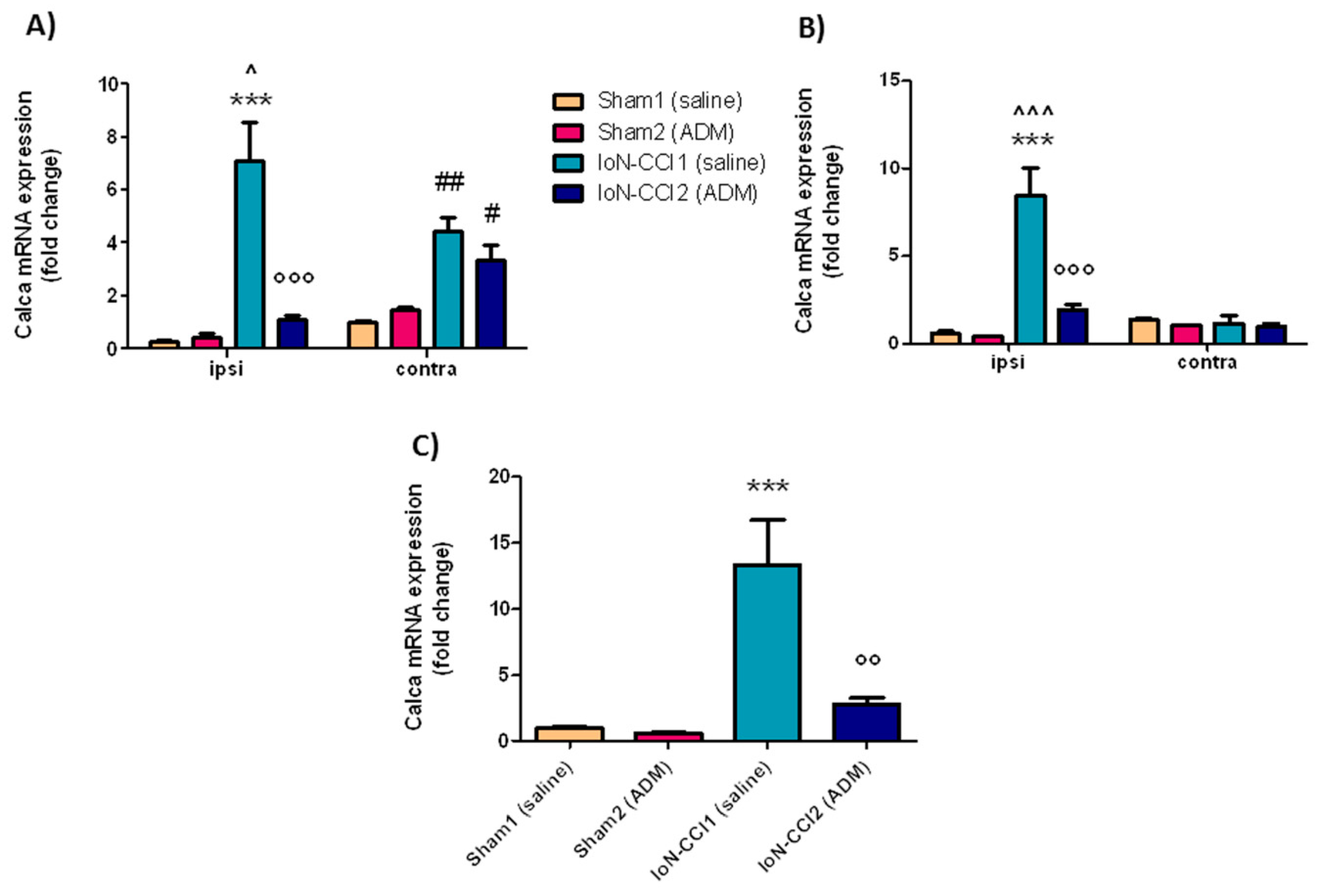
2.2.3. Calca mRNA Expression
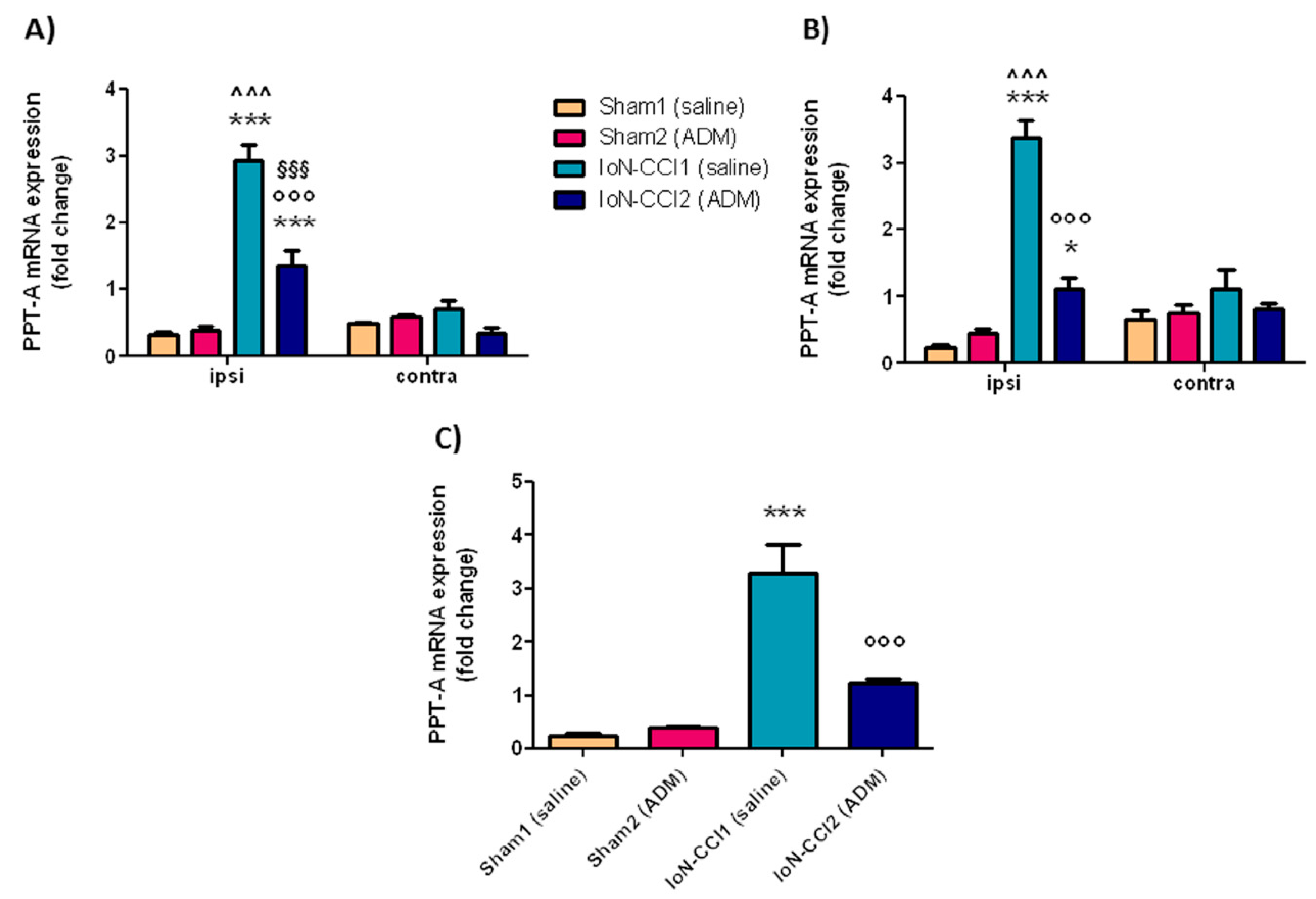
2.2.4. PPT-A mRNA Expression
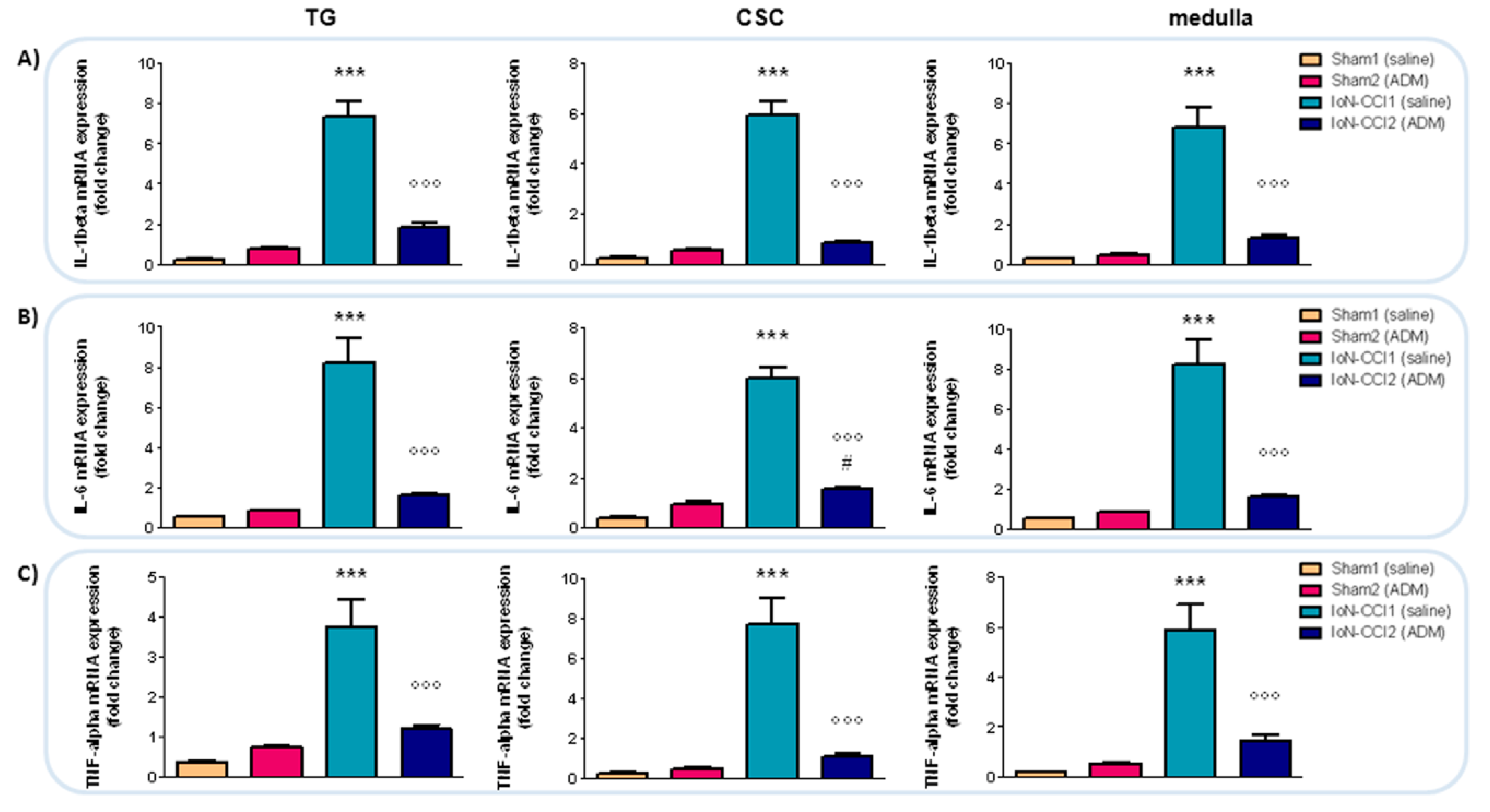
2.2.5. IL-1beta, IL-6, and TNF-alpha mRNA Expression
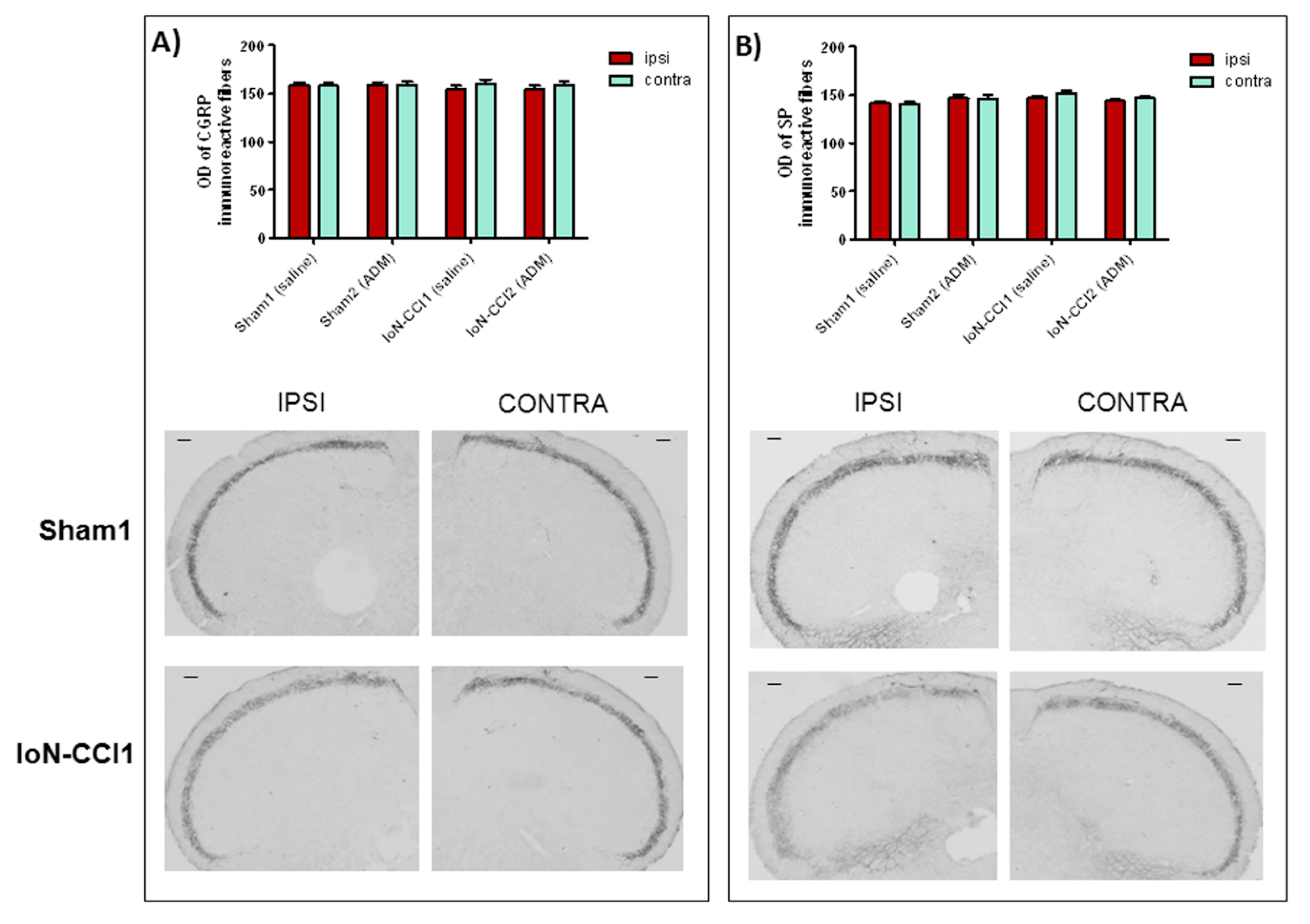
2.3. ADM_12 Effect on Neuropeptide Protein Expression
3. Discussion
3.1. Behavioral Response
3.2. Trpa1 and Trpv1 mRNA Expression
3.3. Neuropeptide Expression
3.4. Limitations of the Study and Future Perspectives
4. Materials and Methods
4.1. Animals
4.2. Surgery
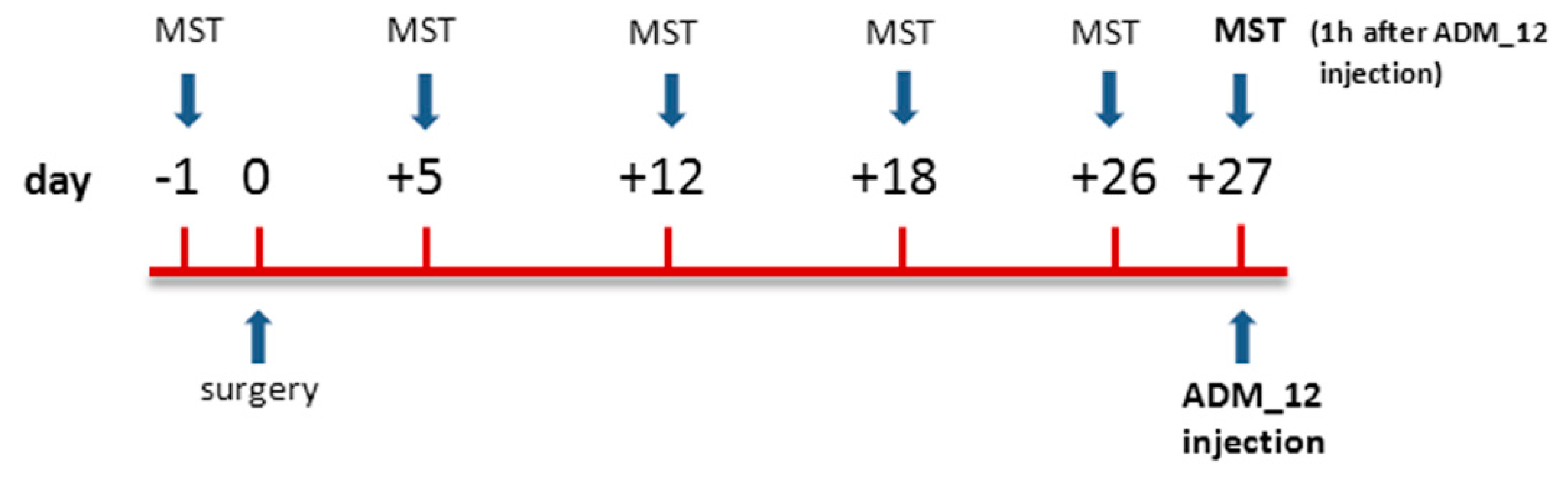
4.3. Mechanical Stimulation Testing (MST)
4.4. Drug and Experimental Plan
4.5. Real Time-PCR
4.6. Immunohistochemistry
4.7. Statistical Evaluation
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CaMKII | Ca(2+)/calmodulin-dependent protein kinase II |
| Calca | calcitonin-related polypeptide alpha |
| CGRP | calcitonin gene-related peptide |
| CREB | cAMP response element-binding protein |
| CSC | cervical spinal cord |
| DRG | dorsal root ganglia |
| GAPDH | glyceraldehyde 3-phosphate dehydrogenase |
| IL | interleukin |
| IoN | infraorbital nerve |
| IoN-CCI | chronic constriction injury of the infraorbital nerve |
| MST | mechanical stimulation testing |
| OD | optical density |
| PPT-A | preprotachykinin-A |
| RT-PCR | real time polymerase chain reaction |
| SP | substance P |
| Sp5C | spinal nucleus of trigeminal nerve pars caudalis |
| TG | trigeminal ganglia |
| TN | trigeminal neuralgia |
| TNF-alpha | tumor necrosis factor alpha |
| TRP | transient receptor potential |
References
- Cruccu, G. Trigeminal Neuralgia. Continuum 2017, 23, 396–420. [Google Scholar] [CrossRef] [PubMed]
- Headache Classification Committee of the International Headache Society (IHS). The International Classification of Headache Disorders, 3rd edition. Cephalalgia 2018, 38, 1–211. [Google Scholar] [CrossRef] [PubMed]
- Zakrzewska, J.M.; Wu, J.; Mon-Williams, M.; Phillips, N.; Pavitt, S.H. Evaluating the impact of trigeminal neuralgia. Pain 2017, 158, 1166–1174. [Google Scholar] [CrossRef] [PubMed]
- Obermann, M.; Katsarava, Z. Update on trigeminal neuralgia. Expert Rev. Neurother. 2009, 9, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Zakrzewska, J.M.; Akram, H. Neurosurgical interventions for the treatment of classical trigeminal neuralgia. Cochrane Database Syst. Rev. 2011, 9, CD007312. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.P.; Wang, X.; Xu, H. TRP channels of intracellular membranes. J. Neurochem. 2010, 113, 313–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Story, G.M.; Peier, A.M.; Reeve, A.J.; Eid, S.R.; Mosbacher, J.; Hricik, T.R.; Earley, T.J.; Hergarden, A.C.; Andersson, D.A.; Hwang, S.W.; et al. ANKTM1, a TRP-like channel expressed in nociceptive neurons, is activated by cold temperatures. Cell 2003, 112, 819–829. [Google Scholar] [CrossRef]
- Bhattacharya, M.R.; Bautista, D.M.; Wu, K.; Haeberle, H.; Lumpkin, E.A.; Julius, D. Radial stretch reveals distinct populations of mechanosensitive mammalian somatosensory neurons. Proc. Natl. Acad. Sci. USA 2008, 105, 20015–20020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quartu, M.; Serra, M.P.; Boi, M.; Poddighe, L.; Picci, C.; Demontis, R.; Del Fiacco, M. TRPV1 receptor in the human trigeminal ganglion and spinal nucleus: Immunohistochemical localization and comparison with the neuropeptides CGRP and SP. J. Anat. 2016, 229, 755–767. [Google Scholar] [CrossRef] [PubMed]
- Jardín, I.; López, J.J.; Diez, R.; Sánchez-Collado, J.; Cantonero, C.; Albarrán, L.; Woodard, G.E.; Redondo, P.C.; Salido, G.M.; Smani, T.; et al. TRPs in Pain Sensation. Front Physiol. 2017, 8, 392. [Google Scholar] [CrossRef] [PubMed]
- Obata, K.; Katsura, H.; Mizushima, T.; Yamanaka, H.; Kobayashi, K.; Dai, Y.; Fukuoka, T.; Tokunaga, A.; Tominaga, M.; Noguchi, K. TRPA1 induced in sensory neurons contributes to cold hyperalgesia after inflammation and nerve injury. J. Clin. Investig. 2005, 115, 2393–2401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katsura, H.; Obata, K.; Mizushima, T.; Yamanaka, H.; Kobayashi, K.; Dai, Y.; Fukuoka, T.; Tokunaga, A.; Sakagami, M.; Noguchi, K. Antisense knock down of TRPA1, but not TRPM8, alleviates cold hyperalgesia after spinal nerve ligation in rats. Exp. Neurol. 2006, 200, 112–123. [Google Scholar] [CrossRef] [PubMed]
- Caspani, O.; Zurborg, S.; Labuz, D.; Heppenstall, P.A. The contribution of TRPM8 and TRPA1 channels to cold allodynia and neuropathic pain. PLoS ONE 2009, 4, e7383. [Google Scholar] [CrossRef] [PubMed]
- Staaf, S.; Oerther, S.; Lucas, G.; Mattsson, J.P.; Ernfors, P. Differential regulation of TRP channels in a rat model of neuropathic pain. Pain 2009, 144, 187–199. [Google Scholar] [CrossRef] [PubMed]
- Eid, S.R.; Crown, E.D.; Moore, E.L.; Liang, H.A.; Choong, K.C.; Dima, S.; Henze, D.A.; Kane, S.A.; Urban, M.O. HC-030031, a TRPA1 selective antagonist, attenuates inflammatory- and neuropathyinduced mechanical hypersensitivity. Mol. Pain 2008, 4, 48. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Joshi, S.K.; DiDomenico, S.; Perner, R.J.; Mikusa, J.P.; Gauvin, D.M.; Segreti, J.A.; Han, P.; Zhang, X.F.; Niforatos, W.; et al. Selective blockade of TRPA1 channel attenuates pathological pain without altering noxious cold sensation or body temperature regulation. Pain 2011, 152, 1165–1172. [Google Scholar] [CrossRef] [PubMed]
- Trevisan, G.; Benemei, S.; Materazzi, S.; De Logu, F.; De Siena, G.; Fusi, C.; Fortes Rossato, M.; Coppi, E.; Marone, I.M.; Ferreira, J.; et al. TRPA1 mediates trigeminal neuropathic pain in mice downstream of monocytes/macrophages and oxidative stress. Brain 2016, 139, 1361–1377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deseure, K.; Hans, G.H. Chronic Constriction Injury of the Rat’s Infraorbital Nerve (IoN-CCI) to Study Trigeminal Neuropathic Pain. J. Vis. Exp. 2015, 103. [Google Scholar] [CrossRef] [PubMed]
- Staruschenko, A.; Jeske, N.A.; Akopian, A.N. Contribution of TRPV1-TRPA1 interaction to the single channel properties of the TRPA1 channel. J. Biol. Chem. 2010, 285, 15167–15177. [Google Scholar] [CrossRef] [PubMed]
- Akopian, A.N. Regulation of nociceptive transmission at the periphery via TRPA1-TRPV1 interactions. Curr. Pharm. Biotechnol. 2011, 12, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Fischer, M.J.; Balasuriya, D.; Jeggle, P.; Goetze, T.A.; McNaughton, P.A.; Reeh, P.W.; Edwardson, J.M. Direct evidence for functional TRPV1/TRPA1 heteromers. Pflugers Arch. 2014, 466, 2229–2241. [Google Scholar] [CrossRef] [PubMed]
- Fragai, M.; Comito, G.; Di Cesare Mannelli, L.; Gualdani, R.; Calderone, V.; Louka, A.; Richichi, B.; Francesconi, O.; Angeli, A.; Nocentini, A.; et al. Lipoyl-Homotaurine Derivative (ADM_12) Reverts Oxaliplatin-Induced Neuropathy and Reduces Cancer Cells Malignancy by Inhibiting Carbonic Anhydrase IX (CAIX). J. Med. Chem. 2017, 60, 9003–9011. [Google Scholar] [CrossRef] [PubMed]
- Gualdani, R.; Ceruti, S.; Magni, G.; Merli, D.; Di Cesare Mannelli, L.; Francesconi, O.; Richichi, B.; la Marca, G.; Ghelardini, C.; Moncelli, M.R.; et al. Lipoic-based TRPA1/TRPV1 antagonist to treat orofacial pain. ACS Chem. Neurosci. 2015, 6, 380–385. [Google Scholar] [CrossRef] [PubMed]
- Demartini, C.; Tassorelli, C.; Zanaboni, A.M.; Tonsi, G.; Francesconi, O.; Nativi, C.; Greco, R. The role of the transient receptor potential ankyrin type-1 (TRPA1) channel in migraine pain: Evaluation in an animal model. J. Headache Pain 2017, 18, 9. [Google Scholar] [CrossRef] [PubMed]
- Vos, B.P.; Strassman, A.M.; Maciewicz, R.J. Behavioral evidence of trigeminal neuropathic pain following chronic constriction injury to the rat’s infraorbital nerve. J. Neurosci. 1994, 14, 2708–2723. [Google Scholar] [CrossRef] [PubMed]
- Costa, G.M.F.; de Oliveira, A.P.; Martinelli, P.M.; da Silva Camargos, E.R.; Arantes, R.M.E.; de Almeida-Leite, C.M. Demyelination/remyelination and expression of interleukin-1β, substance P, nerve growth factor, and glial-derived neurotrophic factor during trigeminal neuropathic pain in rats. Neurosci. Lett. 2016, 612, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Deseure, K.; Hans, G. Behavioral study of non-evoked orofacial pain following different types of infraorbital nerve injury in rats. Physiol. Behav. 2015, 138, 292–296. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, Y.; Une, Y.; Miyano, K.; Abe, H.; Hisaoka, K.; Morioka, N.; Nakata, Y. Activation of transient receptor potential ankyrin 1 evokes nociception through substance P release from primary sensory neurons. J. Neurochem. 2012, 120, 1036–1047. [Google Scholar] [CrossRef] [PubMed]
- Benemei, S.; Fusi, C.; Trevisan, G.; Geppetti, P. The TRPA1 channel in migraine mechanism and treatment. Br. J. Pharmacol. 2014, 171, 2552–2567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gold, M.S.; Gebhart, G.F. Nociceptor sensitization in pain pathogenesis. Nat. Med. 2010, 16, 1248–1257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.; Xie, N.; Lian, Y.; Xu, H.; Chen, C.; Zheng, Y.; Chen, Y.; Zhang, H. Central antinociceptive activity of peripherally applied botulinum toxin type A in lab rat model of trigeminal neuralgia. SpringerPlus 2016, 5, 431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Latrémolière, A.; Mauborgne, A.; Masson, J.; Bourgoin, S.; Kayser, V.; Hamon, M.; Pohl, M. Differential implication of proinflammatory cytokine interleukin-6 in the development of cephalic versus extracephalic neuropathic pain in rats. J. Neurosci. 2008, 28, 8489–8501. [Google Scholar] [CrossRef] [PubMed]
- Choi, B.M.; Lee, S.H.; An, S.M.; Park, D.Y.; Lee, G.W.; Noh, G.J. The time-course and RNA interference of TNF-α, IL-6, and IL-1β expression on neuropathic pain induced by L5 spinal nerve transection in rats. Korean J. Anesthesiol. 2015, 68, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Yu, Y.; Aori, G.; Wang, Q.; Kong, D.; Yang, W.; Guo, Z.; Zhang, L. Tanshinone IIA Attenuates Diabetic Peripheral Neuropathic Pain in Experimental Rats via Inhibiting Inflammation. Evid.-Based Complement. Altern. Med. 2018, 2018, 2789847. [Google Scholar] [CrossRef] [PubMed]
- Atoyan, R.; Shander, D.; Botchkareva, N.V. Non-neuronal expression of transient receptor potential type A1 (TRPA1) in human skin. J. Invest. Dermatol. 2009, 129, 2312–2315. [Google Scholar] [CrossRef] [PubMed]
- Thacker, M.A.; Clark, A.K.; Marchand, F.; McMahon, S.B. Pathophysiology of peripheral neuropathic pain: Immune cells and molecules. Anesth. Analg. 2007, 105, 838–847. [Google Scholar] [CrossRef] [PubMed]
- Mika, J.; Zychowska, M.; Popiolek-Barczyk, K.; Rojewska, E.; Przewlocka, B. Importance of glial activation in neuropathic pain. Eur. J. Pharmacol. 2013, 716, 106–119. [Google Scholar] [CrossRef] [PubMed]
- Ledeboer, A.; Sloane, E.M.; Milligan, E.D.; Frank, M.G.; Mahony, J.H.; Maier, S.F.; Watkins, L.R. Minocycline attenuates mechanical allodynia and proinflammatory cytokine expression in rat models of pain facilitation. Pain 2005, 115, 71–83. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Z.Y.; Gerner, P.; Woolf, C.J.; Ji, R.R. ERK is sequentially activated in neurons, microglia, and astrocytes by spinal nerve ligation and contributes to mechanical allodynia in this neuropathic pain model. Pain 2005, 114, 149–159. [Google Scholar] [CrossRef] [PubMed]
- Lees, J.G.; Duffy, S.S.; Moalem-Taylor, G. Immunotherapy targeting cytokines in neuropathic pain. Front. Pharmacol. 2013, 4, 142. [Google Scholar] [CrossRef] [PubMed]
- Zanjani, T.M.; Sabetkasaei, M.; Karimian, B.; Labibi, F.; Farokhi, B.; Mossafa, N. The attenuation of pain behaviour and serum interleukin-6 concentration by nimesulide in a rat model of neuropathic pain. Scand. J. Pain 2010, 1, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Piao, Z.G.; Cho, I.H.; Park, C.K.; Hong, J.P.; Choi, S.Y.; Lee, S.J.; Lee, S.; Park, K.; Kim, J.S.; Oh, S.B. Activation of glia and microglial p38 MAPK in medullary dorsal horn contributes to tactile hypersensitivity following trigeminal sensory nerve injury. Pain 2006, 121, 219–231. [Google Scholar] [CrossRef] [PubMed]
- Kerstein, P.C.; del Camino, D.; Moran, M.M.; Stucky, C.L. Pharmacological blockade of TRPA1 inhibits mechanical firing in nociceptors. Mol. Pain 2009, 5, 19. [Google Scholar] [CrossRef] [PubMed]
- Kwan, K.Y.; Glazer, J.M.; Corey, D.P.; Rice, F.L.; Stucky, C.L. TRPA1 modulates mechanotransduction in cutaneous sensory neurons. J. Neurosci. 2009, 29, 4808–4819. [Google Scholar] [CrossRef] [PubMed]
- Lehto, S.G.; Weyer, A.D.; Youngblood, B.D.; Zhang, M.; Yin, R.; Wang, W.; Teffera, Y.; Cooke, M.; Stucky, C.L.; Schenkel, L.; et al. Selective antagonism of TRPA1 produces limited efficacy in models of inflammatory- and neuropathic-induced mechanical hypersensitivity in rats. Mol. Pain 2016, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, H.; Koivisto, A.; Saarnilehto, M.; Chapman, H.; Kuokkanen, K.; Hao, B.; Huang, J.L.; Wang, Y.X.; Pertovaara, A. Spinal transient receptor potential ankyrin 1 channel contributes to central pain hypersensitivity in various pathophysiological conditions in the rat. Pain 2011, 152, 582–591. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Zheng, L.; Acosta, G.; Vega-Alvarez, S.; Chen, Z.; Muratori, B.; Cao, P.; Shi, R. Acrolein contributes to TRPA1 up-regulation in peripheral and central sensory hypersensitivity following spinal cord injury. J. Neurochem. 2015, 135, 987–997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Materazzi, S.; Fusi, C.; Benemei, S.; Pedretti, P.; Patacchini, R.; Nilius, B.; Prenen, J.; Creminon, C.; Geppetti, P.; Nassini, R. TRPA1 and TRPV4 mediate paclitaxel-induced peripheral neuropathy in mice via a glutathione-sensitive mechanism. Pflugers Arch. 2012, 463, 561–569. [Google Scholar] [CrossRef] [PubMed]
- Trevisan, G.; Materazzi, S.; Fusi, C.; Altomare, A.; Aldini, G.; Lodovici, M.; Patacchini, R.; Geppetti, P.; Nassini, R. Novel Therapeutic Strategy to Prevent Chemotherapy-Induced Persistent Sensory Neuropathy By TRPA1 Blockade. Cancer Res. 2013, 73, 3120–3131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandewauw, I.; Owsianik, G.; Voets, T. Systematic and quantitative mRNA expression analysis of TRP channel genes at the single trigeminal and dorsal root ganglion level in mouse. BMC Neurosci. 2013, 14, 21. [Google Scholar] [CrossRef] [PubMed]
- Paulsen, C.E.; Armache, J.P.; Gao, Y.; Cheng, Y.; Julius, D. Structure of the TRPA1 ion channel suggests regulatory mechanisms. Nature 2015, 525, 552. [Google Scholar] [CrossRef] [PubMed]
- Jancalek, R. Signaling mechanisms in mirror image pain pathogenesis. Ann. Neurosci. 2011, 18, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Amaya, F.; Oh-hashi, K.; Naruse, Y.; Iijima, N.; Ueda, M.; Shimosato, G.; Tominaga, M.; Tanaka, Y.; Tanaka, M. Local inflammation increases vanilloid receptor 1 expression within distinct subgroups of DRG neurons. Brain Res. 2003, 963, 190–196. [Google Scholar] [CrossRef]
- Devesa, I.; Planells-Cases, R.; Fernández-Ballester, G.; González-Ros, J.M.; Ferrer-Montiel, A.; Fernández-Carvajal, A. Role of the transient receptor potential vanilloid 1 in inflammation and sepsis. J. Inflamm. Res. 2011, 4, 67–81. [Google Scholar] [CrossRef] [PubMed]
- Diogenes, A.; Akopian, A.N.; Hargreaves, K.M. NGF up-regulates TRPA1: Implications for orofacial pain. J. Dent. Res. 2007, 86, 550–555. [Google Scholar] [CrossRef] [PubMed]
- Hatano, N.; Itoh, Y.; Suzuki, H.; Muraki, Y.; Hayashi, H.; Onozaki, K.; Wood, I.C.; Beech, D.J.; Muraki, K. Hypoxia-inducible factor-1α (HIF1α) switches on transient receptor potential ankyrin repeat 1 (TRPA1) gene expression via a hypoxia response element-like motif to modulate cytokine release. J. Biol. Chem. 2012, 287, 31962–31972. [Google Scholar] [CrossRef] [PubMed]
- Hensellek, S.; Brell, P.; Schaible, H.G.; Bräuer, R.; Segond von Banchet, G. The cytokine TNFalpha increases the proportion of DRG neurones expressing the TRPV1 receptor via the TNFR1 receptor and ERK activation. Mol. Cell Neurosci. 2007, 36, 381–391. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.A.; Diogenes, A.; Jeske, N.A.; Henry, M.A.; Akopian, A.; Hargreaves, K.M. Tumor necrosis factor alpha enhances the sensitivity of rat trigeminal neurons to capsaicin. Neuroscience 2008, 155, 503–509. [Google Scholar] [CrossRef] [PubMed]
- Ikeda-Miyagawa, Y.; Kobayashi, K.; Yamanaka, H.; Okubo, M.; Wang, S.; Dai, Y.; Yagi, H.; Hirose, M.; Noguchi, K. Peripherally increased artemin is a key regulator of TRPA1/V1 expression in primary afferent neurons. Mol. Pain 2015, 11, 8. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, K.; Chiba, N.; Chiba, T.; Kambe, T.; Abe, K.; Kawakami, K.; Utsunomiya, I.; Taguchi, K. Transient receptor potential ankyrin 1 that is induced in dorsal root ganglion neurons contributes to acute cold hypersensitivity after oxaliplatin administration. Mol. Pain 2015, 11, 69. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.; Ding, Y.; Li, B.; Liu, H.; Yang, X.; Chen, M. TRPA1 mediated aggravation of allergic contact dermatitis induced by DINP and regulated by NF-κB activation. Sci. Rep. 2017, 7, 43586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; You, B.; Jo, E.K.; Han, S.K.; Simon, M.I.; Lee, S.J. NADPH oxidase 2-derived reactive oxygen species in spinal cord microglia contribute to peripheral nerve injury-induced neuropathic pain. Proc. Natl. Acad. Sci. USA 2010, 107, 14851–14856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.K.; Park, S.K.; Zhou, J.L.; Taglialatela, G.; Chung, K.; Coggeshall, R.E.; Chung, J.M. Reactive oxygen species (ROS) play an important role in a rat model of neuropathic pain. Pain 2004, 111, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Naik, A.K.; Tandan, S.K.; Dudhgaonkar, S.P.; Jadhav, S.H.; Kataria, M.; Prakash, V.R.; Kumar, D. Role of oxidative stress in pathophysiology of peripheral neuropathy and modulation by N-acetyl-L-cysteine in rats. Eur. J. Pain 2006, 10, 573–579. [Google Scholar] [CrossRef] [PubMed]
- Andersson, D.A.; Gentry, C.; Moss, S.; Bevan, S. Transient receptor potential A1 is a sensory receptor for multiple products of oxidative stress. J. Neurosci. 2008, 28, 2485–2494. [Google Scholar] [CrossRef] [PubMed]
- Hudson, L.J.; Bevan, S.; Wotherspoon, G.; Gentry, C.; Fox, A.; Winter, J. VR1 protein expression increases in undamaged DRG neurons after partial nerve injury. Eur. J. Neurosci. 2001, 13, 2105–2114. [Google Scholar] [CrossRef] [PubMed]
- Fukuoka, T.; Tokunaga, A.; Tachibana, T.; Dai, Y.; Yamanaka, H.; Noguchi, K. VR1, but not P2X(3), increases in the spared L4 DRG in rats with L5 spinal nerve ligation. Pain 2002, 99, 111–120. [Google Scholar] [CrossRef]
- Frederick, J.; Buck, M.E.; Matson, D.J.; Cortright, D.N. Increased TRPA1, TRPM8, and TRPV2 expression in dorsal root ganglia by nerve injury. Biochem. Biophys. Res. Commun. 2007, 358, 1058–1064. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Back, S.K.; Davies, A.J.; Jeong, H.; Jo, H.J.; Chung, G.; Na, H.S.; Bae, Y.C.; Kim, S.J.; Kim, J.S.; et al. TRPV1 in GABAergic interneurons mediates neuropathic mechanical allodynia and disinhibition of the nociceptive circuitry in the spinal cord. Neuron 2012, 74, 640–647. [Google Scholar] [CrossRef] [PubMed]
- Urano, H.; Ara, T.; Fujinami, Y.; Hiraoka, B.Y. Aberrant TRPV1 expression in heat hyperalgesia associated with trigeminal neuropathic pain. Int. J. Med. Sci. 2012, 9, 690–697. [Google Scholar] [CrossRef] [PubMed]
- Quartu, M.; Carozzi, V.A.; Dorsey, S.G.; Serra, M.P.; Poddighe, L.; Picci, C.; Boi, M.; Melis, T.; Del Fiacco, M.; Meregalli, C.; et al. Bortezomib treatment produces nocifensive behavior and changes in the expression of TRPV1, CGRP, and substance P in the rat DRG, spinal cord, and sciatic nerve. Biomed. Res. Int. 2014, 2014, 180428. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, E.S.; Christianson, J.A.; Chen, X.; La, J.H.; Davis, B.M.; Albers, K.M.; Gebhart, G.F. Synergistic role of TRPV1 and TRPA1 in pancreatic pain and inflammation. Gastroenterology 2011, 140, 1283–1291. [Google Scholar] [CrossRef] [PubMed]
- Nummenmaa, E.; Hämäläinen, M.; Moilanen, L.J.; Paukkeri, E.L.; Nieminen, R.M.; Moilanen, T.; Vuolteenaho, K.; Moilanen, E. Transient receptor potential ankyrin 1 (TRPA1) is functionally expressed in primary human osteoarthritic chondrocytes. Arthritis Res. Ther. 2016, 18, 185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tohda, C.; Sasaki, M.; Konemura, T.; Sasamura, T.; Itoh, M.; Kuraishi, Y. Axonal transport of VR1 capsaicin receptor mRNA in primary afferents and its participation in inflammation-induced increase in capsaicin sensitivity. J. Neurochem. 2001, 76, 1628–1635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosugi, M.; Nakatsuka, T.; Fujita, T.; Kuroda, Y.; Kumamoto, E. Activation of TRPA1 channel facilitates excitatory synaptic transmission in substantia gelatinosa neurons of the adult rat spinal cord. J. Neurosci. 2007, 27, 4443–4451. [Google Scholar] [CrossRef] [PubMed]
- Koivisto, A.; Chapman, H.; Jalava, N.; Korjamo, T.; Saarnilehto, M.; Lindstedt, K.; Pertovaara, A. TRPA1: A transducer and amplifier of pain and inflammation. Basic Clin. Pharmacol. Toxicol. 2014, 114, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Chu, Y.; Han, L.; Li, M.; Li, Z.; LaVinka, P.C.; Sun, S.; Tang, Z.; Park, K.; Caterina, M.J.; et al. Central terminal sensitization of TRPV1 by descending serotonergic facilitation modulates chronic pain. Neuron 2014, 81, 873–887. [Google Scholar] [CrossRef] [PubMed]
- Yeo, E.J.; Cho, Y.S.; Paik, S.K.; Yoshida, A.; Park, M.J.; Ahn, D.K.; Moon, C.; Kim, Y.S.; Bae, Y.C. Ultrastructural analysis of the synaptic connectivity of TRPV1-expressing primary afferent terminals in the rat trigeminal caudal nucleus. J. Comp. Neurol. 2010, 518, 4134–4146. [Google Scholar] [CrossRef] [PubMed]
- Doly, S.; Fischer, J.; Salio, C.; Conrath, M. The vanilloid receptor-1 is expressed in rat spinal dorsal horn astrocytes. Neurosci. Lett. 2004, 357, 123–126. [Google Scholar] [CrossRef] [PubMed]
- Mandadi, S.; Armati, P.J.; Roufogalis, B.D. Protein kinase C modulation of thermo-sensitive transient receptor potential channels: Implications for pain signaling. J. Nat. Sci. Biol. Med. 2011, 2, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Zurborg, S.; Yurgionas, B.; Jira, J.A.; Caspani, O.; Heppenstall, P.A. Direct activation of the ion channel TRPA1 by Ca2+. Nat. Neurosci. 2007, 10, 277–279. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, N.; Mizuno, Y.; Kozai, D.; Yamamoto, S.; Kiyonaka, S.; Shibata, T.; Uchida, K.; Mori, Y. Molecular characterization of TRPA1 channel activation by cysteine-reactive inflammatory mediators. Channels 2008, 2, 287–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bandell, M.; Story, G.M.; Hwang, S.W.; Viswanath, V.; Eid, S.R.; Petrus, M.J.; Earley, T.J.; Patapoutian, A. Noxious cold ion channel TRPA1 is activated by pungent compounds and bradykinin. Neuron 2004, 41, 849–857. [Google Scholar] [CrossRef]
- Bautista, D.M.; Jordt, S.E.; Nikai, T.; Tsuruda, P.R.; Read, A.J.; Poblete, J.; Yamoah, E.N.; Basbaum, A.I.; Julius, D. TRPA1 mediates the inflammatory actions of environmental irritants and proalgesic agents. Cell 2006, 124, 1269–1282. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Dai, Y.; Fukuoka, T.; Yamanaka, H.; Kobayashi, K.; Obata, K.; Cui, X.; Tominaga, M.; Noguchi, K. Phospholipase C and protein kinase A mediate bradykinin sensitization of TRPA1: A molecular mechanism of inflammatory pain. Brain 2008, 131, 1241–1251. [Google Scholar] [CrossRef] [PubMed]
- Lee, L.Y.; Hsu, C.C.; Lin, Y.J.; Lin, R.L.; Khosravi, M. Interaction between TRPA1 and TRPV1: Synergy on pulmonary sensory nerves. Pulm. Pharmacol. Ther. 2015, 35, 87–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masuoka, T.; Kudo, M.; Yamashita, Y.; Yoshida, J.; Imaizumi, N.; Muramatsu, I.; Nishio, M.; Ishibashi, T. TRPA1 Channels Modify TRPV1-Mediated Current Responses in Dorsal Root Ganglion Neurons. Front. Physiol. 2017, 8, 272. [Google Scholar] [CrossRef] [PubMed]
- Matzner, O.; Devor, M. Hyperexcitability at sites of nerve injury depends on voltage-sensitive Na+ channels. J. Neurophysiol. 1994, 72, 349–359. [Google Scholar] [CrossRef] [PubMed]
- McCallum, J.B.; Wu, H.E.; Tang, Q.; Kwok, W.M.; Hogan, Q.H. Subtype-specific reduction of voltage-gated calcium current in medium-sized dorsal root ganglion neurons after painful peripheral nerve injury. Neuroscience 2011, 179, 244–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waxman, S.G.; Kocsis, J.D.; Black, J.A. Type III Na+ channel mRNA is expressed in embryonic but not adult spinal sensory neurons, and is re-expressed following axotomy. J. Neurophysiol. 1994, 72, 466–470. [Google Scholar] [CrossRef] [PubMed]
- Ruparel, N.B.; Patwardhan, A.M.; Akopian, A.N.; Hargreaves, K.M. Desensitization of transient receptor potential ankyrin 1 (TRPA1) by the TRP vanilloid 1-selective cannabinoid arachidonoyl-2 chloroethanolamine. Mol. Pharmacol. 2011, 80, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Andrade, E.L.; Meotti, F.C.; Calixto, J.B. TRPA1 antagonists as potential analgesic drugs. Pharmacol. Ther. 2012, 133, 189–204. [Google Scholar] [CrossRef] [PubMed]
- Honda, K.; Shinoda, M.; Furukawa, A.; Kita, K.; Noma, N.; Iwata, K. TRPA1 contributes to capsaicin-induced facial cold hyperalgesia in rats. Eur. J. Oral Sci. 2014, 122, 391–396. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.H.; Kim, J.I.; Han, J.A.; Choe, M.A.; Ahn, J.H. Upregulation of neuronal nitric oxide synthase in the periphery promotes pain hypersensitivity after peripheral nerve injury. Neuroscience 2011, 8, 367–378. [Google Scholar] [CrossRef] [PubMed]
- Costa, G.M.F.; Leite, C.M.A. Trigeminal neuralgia: Peripheral and central mechanisms. Rev. Dor São Paulo. 2015, 16, 297–301. [Google Scholar] [CrossRef]
- Pfaller, K.; Arvidsson, J. Central distribution of trigeminal and upper cervical primary afferents in the rat studied by anterograde transport of horseradish peroxidase conjugated to wheat germ agglutinin. J. Comp. Neurol. 1988, 268, 91–108. [Google Scholar] [CrossRef] [PubMed]
- Jacquin, M.F.; Chiaia, N.L.; Rhoades, R.W. Trigeminal projections to contralateral dorsal horn: Central extent, peripheral origins, and plasticity. Somatosens. Mot. Res. 1990, 7, 153–183. [Google Scholar] [CrossRef] [PubMed]
- Ingvardsen, B.K.; Laursen, H.; Olsen, U.B.; Hansen, A.J. Possible mechanism of c-fos expression in trigeminal nucleus caudalis following cortical spreading depression. Pain 1997, 72, 407–415. [Google Scholar] [CrossRef]
- Samsam, M.; Coveñas, R.; Csillik, B.; Ahangari, R.; Yajeya, J.; Riquelme, R.; Narváez, J.A.; Tramu, G. Depletion of substance P, neurokinin A and calcitonin gene-related peptide from the contralateral and ipsilateral caudal trigeminal nucleus following unilateral electrical stimulation of the trigeminal ganglion; a possible neurophysiological and neuroanatomical link to generalized head pain. J. Chem. Neuroanat. 2001, 21, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Nakanishi, M.; Hata, K.; Nagayama, T.; Sakurai, T.; Nishisho, T.; Wakabayashi, H.; Hiraga, T.; Ebisu, S.; Yoneda, T. Acid activation of Trpv1 leads to an up-regulation of calcitonin gene-related peptide expression in dorsal root ganglion neurons via the CaMK-CREB cascade: A potential mechanism of inflammatory pain. Mol. Biol. Cell 2010, 21, 2568–2577. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Li, S.; Dhaka, A.; Story, G.M.; Cao, Y.Q. Expression of the transient receptor potential channels TRPV1, TRPA1 and TRPM8 in mouse trigeminal primary afferent neurons innervating the dura. Mol. Pain 2012, 8, 66. [Google Scholar] [CrossRef] [PubMed]
- Qin, Z.L.; Yang, L.Q.; Li, N.; Yue, J.N.; Wu, B.S.; Tang, Y.Z.; Guo, Y.N.; Lai, G.H.; Ni, J.X. Clinical study of cerebrospinal fluid neuropeptides in patients with primary trigeminal neuralgia. Clin. Neurol. Neurosurg. 2016, 143, 111–115. [Google Scholar] [CrossRef] [PubMed]
- Strittmatter, M.; Grauer, M.; Isenberg, E.; Hamann, G.; Fischer, C.; Hoffmann, K.H.; Blaes, F.; Schimrigk, K. Cerebrospinal fluid neuropeptides and monoaminergic transmitters in patients with trigeminal neuralgia. Headache 1997, 37, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Lynds, R.; Lyu, C.; Lyu, G.W.; Shi, X.Q.; Rosén, A.; Mustafa, K.; Shi, T.J.S. Neuronal plasticity of trigeminal ganglia in mice following nerve injury. J. Pain Res. 2017, 10, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Aita, M.; Chavkin, C. Partial infraorbital nerve ligation as a model of trigeminal nerve injury in the mouse: Behavioral, neural, and glial reactions. J. Pain 2008, 9, 1036–1048. [Google Scholar] [CrossRef] [PubMed]
- Chiechio, S.; Copani, A.; De Petris, L.; Morales, M.E.; Nicoletti, F.; Gereau, R.W., 4th. Transcriptional regulation of metabotropic glutamate receptor 2/3 expression by the NF-kappaB pathway in primary dorsal root ganglia neurons: A possible mechanism for the analgesic effect of L-acetylcarnitine. Mol. Pain 2006, 2, 20. [Google Scholar] [CrossRef] [PubMed]
- Ambalavanar, R.; Dessem, D.; Moutanni, A.; Yallampalli, C.; Yallampalli, U.; Gangula, P.; Bai, G. Muscle inflammation induces a rapid increase in calcitonin gene-related peptide (CGRP) mRNA that temporally relates to CGRP immunoreactivity and nociceptive behavior. Neuroscience 2006, 143, 875–884. [Google Scholar] [CrossRef] [PubMed]
- Nesic, O.; Svrakic, N.M.; Xu, G.Y.; McAdoo, D.; Westlund, K.N.; Hulsebosch, C.E.; Ye, Z.; Galante, A.; Soteropoulos, P.; Tolias, P.; et al. DNA microarray analysis of the contused spinal cord: Effect of NMDA receptor inhibition. J. Neurosci. Res. 2002, 68, 406–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmerman, M. Ethical guidelines for investigations of experimental pain in conscious animals. Pain 1983, 16, 109–110. [Google Scholar] [CrossRef] [Green Version]
- Deseure, K.; Koek, W.; Colpaert, F.C.; Adriaensen, H. The 5-HT(1A) receptor agonist F 13640 attenuates mechanical allodynia in a rat model of trigeminal neuropathic pain. Eur. J. Pharmacol. 2002, 456, 51–57. [Google Scholar] [CrossRef]
- Deseure, K.; Koek, W.; Adriaensen, H.; Colpaert, F.C. Continuous administration of the 5-hydroxytryptamine1A agonist (3-Chloro-4-fluoro-phenyl)-[4-fluoro-4-[[(5-methyl-pyridin-2-ylmethyl) -amino]-methyl]piperidin-1-yl]-methadone (F 13640) attenuates allodynia-like behavior in a rat model of trigeminal neuropathic pain. J. Pharmacol. Exp. Ther. 2003, 306, 505–514. [Google Scholar] [PubMed]
- Deseure, K.R.; Adriaensen, H.F.; Colpaert, F.C. Effects of the combined continuous administration of morphine and the high-efficacy 5-HT1A agonist, F 13640 in a rat model of trigeminal neuropathic pain. Eur. J. Pain 2004, 8, 547–554. [Google Scholar] [CrossRef] [PubMed]
- Deseure, K.; Bréand, S.; Colpaert, F.C. Curative-like analgesia in a neuropathic pain model: Parametric analysis of the dose and the duration of treatment with a high-efficacy 5-HT(1A) receptor agonist. Eur. J. Pharmacol. 2007, 568, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Greco, R.; Demartini, C.; Zanaboni, A.M.; Redavide, E.; Pampalone, S.; Toldi, J.; Fülöp, F.; Blandini, F.; Nappi, G.; Sandrini, G.; et al. Effects of kynurenic acid analogue 1 (KYNA-A1) in nitroglycerin-induced hyperalgesia: Targets and anti-migraine mechanisms. Cephalalgia 2017, 37, 1272–1284. [Google Scholar] [CrossRef] [PubMed]
- Greco, R.; Ferrigno, A.; Demartini, C.; Zanaboni, A.; Mangione, A.S.; Blandini, F.; Nappi, G.; Vairetti, M.; Tassorelli, C. Evaluation of ADMA-DDAH-NOS axis in specific brain areas following nitroglycerin administration: Study in an animal model of migraine. J. Headache Pain 2015, 16, 560. [Google Scholar] [CrossRef] [PubMed]
- Terayama, R.; Nagamatsu, N.; Ikeda, T.; Nakamura, T.; Rahman, O.I.; Sakoda, S.; Shiba, R.; Nishimori, T. Differential expression of Fos protein after transection of the rat infraorbital nerve in the trigeminal nucleus caudalis. Brain Res. 1997, 768, 135–146. [Google Scholar] [CrossRef]
- Panneton, W.M.; Pan, B.; Gan, Q. Somatotopy in the Medullary Dorsal Horn As a Basis for Orofacial Reflex Behavior. Front. Neurol. 2017, 8, 522. [Google Scholar] [CrossRef] [PubMed]
- Greco, R.; Tassorelli, C.; Sandrini, G.; Di Bella, P.; Buscone, S.; Nappi, G. Role of calcitonin gene-related peptide and substance P in different models of pain. Cephalalgia 2008, 28, 114–126. [Google Scholar] [CrossRef] [PubMed]
- Green, D.; Ruparel, S.; Gao, X.; Ruparel, N.; Patil, M.; Akopian, A.; Hargreaves, K. Central activation of TRPV1 and TRPA1 by novel endogenous agonists contributes to mechanical allodynia and thermal hyperalgesia after burn injury. Mol. Pain 2016, 12. [Google Scholar] [CrossRef] [PubMed]
- Ossipov, M.H.; Lai, J.; Malan, T.P., Jr.; Porreca, F. Spinal and supraspinal mechanisms of neuropathic pain. Ann. N. Y. Acad. Sci. 2000, 909, 12–24. [Google Scholar] [CrossRef] [PubMed]
- Zakrzewska, J.M. Differential diagnosis of facial pain and guidelines for management. Br. J. Anaesth. 2013, 111, 95–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garrison, S.R.; Stucky, C.L. The dynamic TRPA1 channel: A suitable pharmacological pain target? Curr. Pharm. Biotechnol. 2011, 12, 1689–1697. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Hackos, D.H. TRPA1 as a drug target—promise and challenges. N.-S. Arch. Pharmacol. 2015, 388, 451–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SCORE | TYPE OF RESPONSE |
|---|---|
| 0 | no response |
| 1 | detection: the rat turns the head toward the stimulating object and the stimulus object is then explored |
| 2 | withdrawal reaction: the rat turns the head slowly away or pulls it briskly backward when the stimulation is applied; sometimes a single face wipe ipsilateral to the stimulated area occurs |
| 3 | escape/attack: the rat avoids further contact with the stimulus object, either passively by moving its body away from the stimulating object to assume a crouching position against the cage wall, or actively by attacking the stimulus object, making biting and grabbing movements |
| 4 | asymmetric face grooming: the rat displays an uninterrupted series of at least three face-wash strokes directed toward the stimulated facial area |
| EXPERIMENTAL GROUPS | Surgery | Treatment on Day +27 | MST | RT-PCR | IHC |
|---|---|---|---|---|---|
| Sham1 | Sham | saline | N = 12 | N = 6 | N = 6 |
| Sham2 | Sham | ADM_12 | N = 12 | N = 6 | N = 6 |
| IoN-CCI1 | IoN-CCI | saline | N = 12 | N = 6 | N = 6 |
| IoN-CCI2 | IoN-CCI | ADM_12 | N = 12 | N = 6 | N = 6 |
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| GAPDH | AACCTGCCAAGTATGATGAC | GGAGTTGCTGTTGAAGTCA |
| Trpa1 | CTCCCCGAGTGCATGAAAGT | TGCATATACGCGGGGATGTC |
| Trpv1 | CTTGCTCCATTTGGGGTGTG | CTGGAGGTGGCTTGCAGTTA |
| Calca | CAGTCTCAGCTCCAAGTCATC | TTCCAAGGTTGACCTCAAAG |
| PPT-A | GCTCTTTATGGGCATGGTC | GGGTTTATTTACGCCTTCTTTC |
| IL-1beta | CTTCCTTGTGCAAGTGTCTG | CAGGTCATTCTCCTCACTGTC |
| IL-6 | TTCTCTCCGCAAGAGACTTC | GGTCTGTTGTGGGTGGTATC |
| TNF-alpha | CCTCACACTCAGATCATCTTCTC | CGCTTGGTGGTTTGCTAC |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Demartini, C.; Greco, R.; Zanaboni, A.M.; Francesconi, O.; Nativi, C.; Tassorelli, C.; Deseure, K. Antagonism of Transient Receptor Potential Ankyrin Type-1 Channels as a Potential Target for the Treatment of Trigeminal Neuropathic Pain: Study in an Animal Model. Int. J. Mol. Sci. 2018, 19, 3320. https://doi.org/10.3390/ijms19113320
Demartini C, Greco R, Zanaboni AM, Francesconi O, Nativi C, Tassorelli C, Deseure K. Antagonism of Transient Receptor Potential Ankyrin Type-1 Channels as a Potential Target for the Treatment of Trigeminal Neuropathic Pain: Study in an Animal Model. International Journal of Molecular Sciences. 2018; 19(11):3320. https://doi.org/10.3390/ijms19113320
Chicago/Turabian StyleDemartini, Chiara, Rosaria Greco, Anna Maria Zanaboni, Oscar Francesconi, Cristina Nativi, Cristina Tassorelli, and Kristof Deseure. 2018. "Antagonism of Transient Receptor Potential Ankyrin Type-1 Channels as a Potential Target for the Treatment of Trigeminal Neuropathic Pain: Study in an Animal Model" International Journal of Molecular Sciences 19, no. 11: 3320. https://doi.org/10.3390/ijms19113320