Pretreatment with NaCl Promotes the Seed Germination of White Clover by Affecting Endogenous Phytohormones, Metabolic Regulation, and Dehydrin-Encoded Genes Expression under Water Stress


Abstract
:1. Introduction
2. Results
2.1. Effects of Seed Soaking with NaCl on Germination Characteristics
2.2. Effect of Seed Soaking with NaCl on the Changes of Endogenous Phytohormones
2.3. Effect of Seed Soaking with NaCl on Starch Metabolism and Osmotic Adjustment
2.4. Effect of Seed Soaking with NaCl on Oxidative Damage
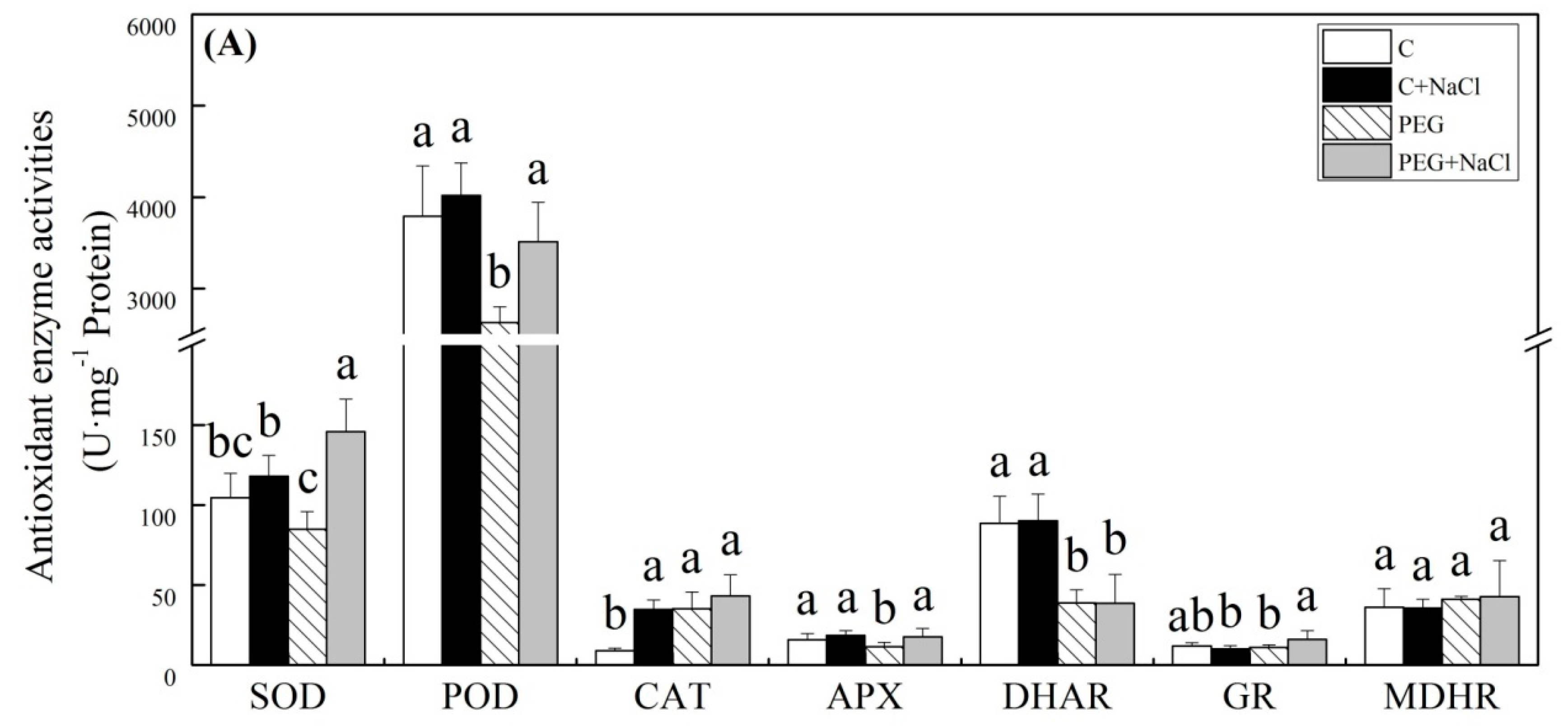
2.5. Effect of Seed Soaking with NaCl on Antioxidant Metabolism
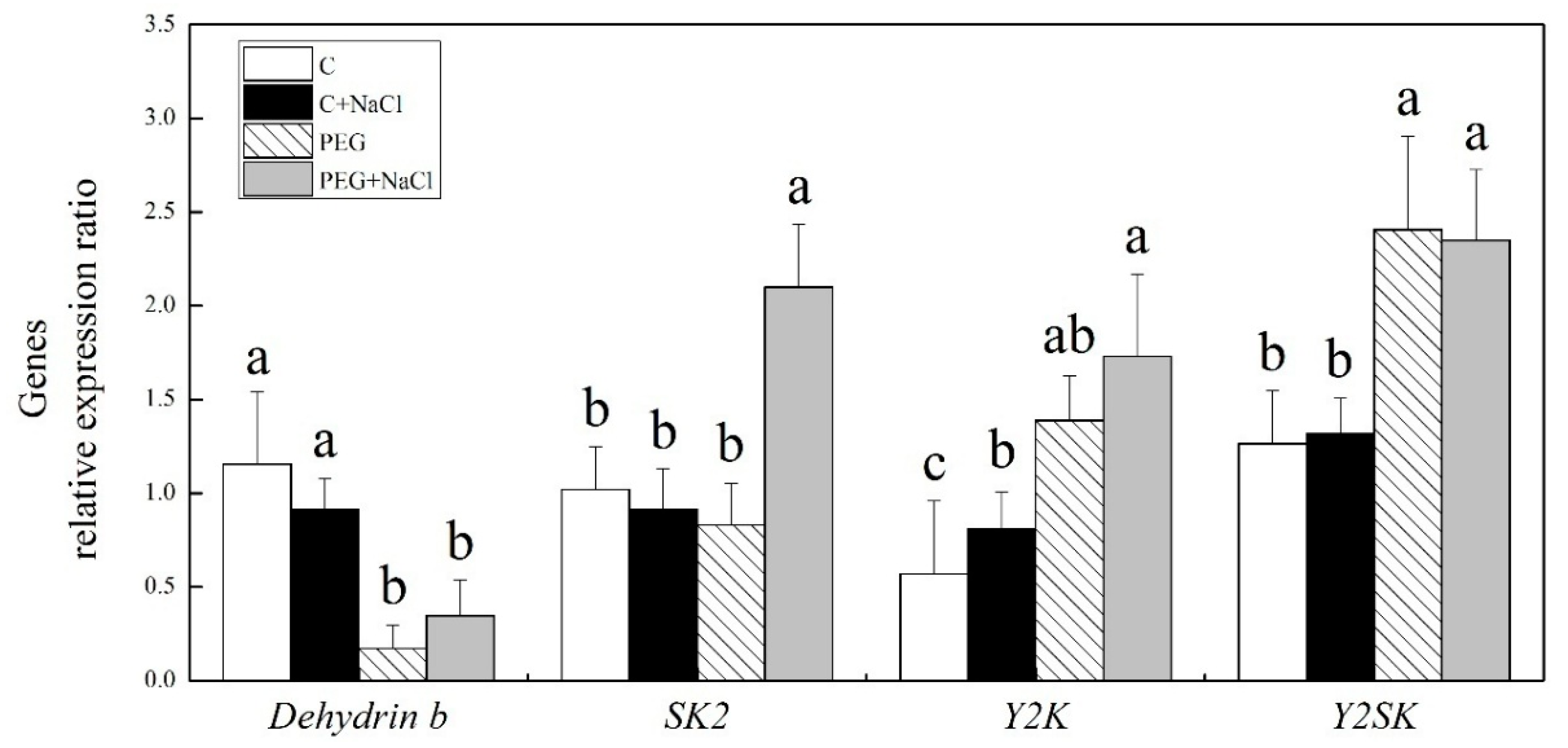
2.6. Effect of Seed Soaking with NaCl on Dehydrin-Encoded Gene Expression
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Treatments
4.2. Determination of Seed Germination Characteristics and Starch Metabolism
4.3. Determination of Endogenous Phytohormones
4.4. Determination of Soluble Sugars, Amino Acids, Proline, and Osmotic Potential
4.5. Determination of Antioxidant Metabolism and Electrolyte Leakage
4.6. Genes Expression Analyses
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Xu, L.; Han, L.; Huang, B. Antioxidant enzyme activities and gene expression patterns in leaves of kentucky bluegrass in response to drought and post-drought recovery. J. Am. Soc. Hort. Sci. 2011, 136, 247–255. [Google Scholar]
- Rajasekaran, L.; RBlake, T.J. New plant growth regulators protect photosynthesis and enhance growth under drought of jack pine seedlings. J. Plant Growth Regul. 1999, 18, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Tsvetkov, I.J.; Atanassov, A.I.; Tsolova, V.M.; Bouquet, A.; Boursiquot, J.M. Gene transfer for stress resistance in grapes. Acta Hort. 2000, 528, 389–396. [Google Scholar] [CrossRef]
- Wang, X.; Huang, W.; Liu, J.; Yang, Z.; Huang, B. Molecular regulation and physiological functions of a novel FaHsfA2c cloned from tall fescue conferring plant tolerance to heat stress. Plant Biotechnol. J. 2017, 15, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Savvides, A.; Ali, S.; Tester, M.; Fotopoulos, V. Chemical priming of plants against multiple abiotic stresses: Mission possible? Trends Plant Sci. 2016, 21, 329–340. [Google Scholar] [CrossRef] [PubMed]
- Burgess, P.; Huang, B. Effects of sequential application of plant growth regulators and osmoregulants on drought tolerance of creeping bentgrass (Agrostis stolonifera). Crop Sci. 2013, 54, 837–844. [Google Scholar] [CrossRef]
- Farouk, S.F.S.; Qados, A.M.S.A. Osmotic adjustment and yield of cowpea in response to drought stress and chitosan. Indian J. Appl. Res. 2011, 3, 1–6. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, Y.; Zhang, X.; Merewitz, E.; Peng, Y.; Ma, X. Metabolic pathways regulated by chitosan contributing to drought resistance in white clover. J. Proteome Res. 2017, 16, 3039–3052. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, P.M.; Bressan, R.A.; Zhu, J.K.; Bohnert, H.J. Plant cellular and molecular responses to high salinity. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2000, 51, 463–499. [Google Scholar] [CrossRef] [PubMed]
- Kawa, D.; Julkowska, M.; Montero, S.H.; Horst, A.T.; Haring, M.A.; Testerink, C. Phosphate-dependent root system architecture responses to salt stress. Plant Physiol. 2016, 172, 690–706. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Chong, J.; Li, C.; Kim, C.; Shi, D.; Wang, D. Osmotic adjustment and ion balance traits of an alkali resistant halophyte Kochia sieversiana during adaptation to salt and alkali conditions. Plant Soil. 2007, 294, 263–276. [Google Scholar] [CrossRef]
- Wang, S.; Wan, C.; Wang, Y.; Chen, H.; Zhou, Z.; Fu, H. The characteristics of Na, K and free proline distribution in several drought-resistant plants of the Alxa Desert, China. J. Arid Environ. 2004, 56, 525–539. [Google Scholar] [CrossRef]
- Martínez, J.P.; Kinet, J.M.; Bajji, M.; Lutts, S. NaCl alleviates polyethylene glycol-induced water stress in the halophyte species Atriplex halimus L. J. Exp. Bot. 2005, 56, 2421–2431. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Peng, D.D.; Zhang, X.Q.; Peng, Y.; Chen, M.; Ma, X.; Huang, L.K.; Yan, Y.H. Na+ induces the tolerance to water stress in white clover associated with osmotic adjustment and aquaporins-mediated water transport and balance in root and leaf. Environ. Exp. Bot. 2017, 144, 11–24. [Google Scholar] [CrossRef]
- Annicchiarico, P.; Piano, E. Indirect selection for root development of white clover and implications for drought tolerance. J. Agron. Crop Sci. 2010, 190, 28–34. [Google Scholar] [CrossRef]
- Sanderson, M.A.; Elwinger, G.F. Grass species and cultivar effects on establishment of grass–white clover mixtures. Agron. J. 1999, 91, 889–897. [Google Scholar] [CrossRef]
- Sincik, M.; Acikgoz, E. Effects of white clover inclusion on turf characteristics, nitrogen fixation, and nitrogen transfer from white clover to grass species in turf mixtures. Commun. Soil Sci. Plant Anal. 2007, 38, 1861–1877. [Google Scholar] [CrossRef]
- Cattivelli, L.; Rizza, F.; Badeck, F.W.; Mazzucotelli, E.; Mastrangelo, A.M.; Francia, E. Drought tolerance improvement in crop plants: An integrated view from breeding to genomics. Field Crop. Res. 2008, 105, 1–14. [Google Scholar] [CrossRef]
- Hameed, A.; Iqbal, N. Chemo-priming with mannose, mannitol and H2O2 mitigate drought stress in wheat. Cereal Res. Commun. 2014, 42, 450–462. [Google Scholar] [CrossRef]
- Li, Z.; Peng, Y.; Zhang, X.Q.; Ma, X.; Hang, L.K.; Yan, Y.H. Exogenous spermidine improves seed germination of white clover under water stress via involvement in starch metabolism, antioxidant defenses and relevant gene expression. Molecules 2014, 19, 18003. [Google Scholar] [CrossRef] [PubMed]
- Farooq, M.; Irfan, M.; Aziz, T.; Ahmad, I.; Cheema, S.A. Seed priming with ascorbic acid improves drought resistance of wheat. J. Agron. Crop Sci. 2013, 199, 12–22. [Google Scholar] [CrossRef]
- Biju, S.; Fuentes, S.; Gupta, D. Silicon improves seed germination and alleviates drought stress in lentil crops by regulating osmolytes, hydrolytic enzymes and antioxidant defense system. Plant Physiol. Biochem. 2017, 119, 250–264. [Google Scholar] [CrossRef] [PubMed]
- Gazzarrini, S.; Tsai, A.Y. Hormone cross-talk during seed germination. Essays Biochem. 2015, 58, 151–164. [Google Scholar] [CrossRef] [PubMed]
- Vanstraelen, M.; Benkovã, E. Hormonal interactions in the regulation of plant development. Annu. Rev. Cell Dev. Biol. 2012, 28, 463–487. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, S.; Davies, W.J. ABA-based chemical signalling: The co-ordination of responses to stress in plants. Plant Cell Environ. 2010, 25, 195–210. [Google Scholar] [CrossRef]
- Liu, N.; Shen, Y.; Huang, B. Osmoregulants involved in osmotic adjustment for differential drought tolerance in different bentgrass genotypes. J. Am. Soc. Hort. Sci. 2015, 140, 605–613. [Google Scholar]
- Robyt, J.F. Chapter IV-Enzymes in the hydrolysis and synthesis of starch. In Starch: Chemistry and Technology, 2nd ed.; Whistler, R.L., Bemiller, J.N., Paschall, E.F., Eds.; Elsevier: Amsterdam, The Netherlands, 1984; pp. 87–123. [Google Scholar]
- Cramer, G.R.; Ergül, A.; Grimplet, J.; Tillett, R.L.; Tattersall, E.A.R.; Bohlman, M.C. Water and salinity stress in grapevines: Early and late changes in transcript and metabolite profiles. Funct. Integr. Genom. 2007, 7, 111–134. [Google Scholar] [CrossRef] [PubMed]
- Szabados, L.; Savouré, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Shabala, S.; Shabala, L. Ion transport and osmotic adjustment in plants and bacteria. Biomol. Concepts 2011, 2, 407–419. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.J.; Wang, Y.J.; Zhou, Y.H.; Tao, Y.; Mao, W.H.; Shi, K. Reactive oxygen species are involved in brassinosteroid-induced stress tolerance in cucumber. Plant Physiol. 2009, 150, 801–814. [Google Scholar] [CrossRef] [PubMed]
- Shah, K.; Kumar, R.G.; Verma, S.; Dubey, R.S. Effect of cadmium on lipid peroxidation, superoxide anion generation and activities of antioxidant enzymes in growing rice seedlings. Plant Sci. 2001, 161, 1135–1144. [Google Scholar] [CrossRef]
- Ennajeh, M.; Vadel, A.M.; Khemira, H. Osmoregulation and osmoprotection in the leaf cells of two olive cultivars subjected to severe water deficit. Acta Physiol. Plant. 2009, 31, 711–721. [Google Scholar] [CrossRef]
- Upadhyaya, H.; Panda, S.K.; Dutta, B.K. Variation of physiological and antioxidative responses in tea cultivars subjected to elevated water stress followed by rehydration recovery. Acta Physiol. Plant. 2008, 30, 457–468. [Google Scholar] [CrossRef]
- Li, Z.; Li, Y.; Zhang, Y.; Cheng, B.; Peng, Y.; Zhang, X. Indole-3-acetic acid modulates phytohormones and polyamines metabolism associated with the tolerance to water stress in white clover. Plant Physiol. Biochem. 2018, 129, 251–263. [Google Scholar] [CrossRef] [PubMed]
- Lecube, M.L.; Noriega, G.O.; Cruz, D.M.S.; Tomaro, M.L.; Batlle, A.; Balestrasse, K.B. Indole acetic acid is responsible for protection against oxidative stress caused by drought in soybean plants: The role of heme oxygenase induction. Redox Rep. 2014, 19, 242–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, H.; Chen, L.; Ye, T.; Liu, X.; Ding, K.; Chan, Z. Modulation of auxin content in arabidopsis confers improved drought stress resistance. Plant Physiol. Biochem. 2014, 82, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Hara, M.; Fujinaga, M.; Kuboi, T. Metal binding by citrus dehydrin with histidine-rich domains. J. Exp. Bot. 2005, 56, 2695–2703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kruger, C.; Berkowitz, O.; Stephan, U.W.; Hell, R. A metal-binding member of the late embryogenesis abundant protein family transports iron in the phloem of Ricinus communis L. J. Biol. Chem. 2002, 277, 25062–25069. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Jing, W.; Peng, Y.; Zhang, X.Q.; Ma, X.; Hang, L.K.; Yan, Y.H. Spermine alleviates drought stress in white clover with different resistance by influencing carbohydrate metabolism and dehydrins synthesis. PLoS ONE 2015, 10, e0120708. [Google Scholar] [CrossRef] [PubMed]
- Hundertmark, M.; Buitink, J.; Leprince, O.; Hincha, D.K. Reduction of seed-specific dehydrins reduces seed longevity in Arabidopsis thaliana. Seed Sci. Res. 2011, 21, 165–173. [Google Scholar] [CrossRef]
- Cheng, B.; Li, Z.; Liang, L.; Cao, Y.; Zeng, W.; Zhang, X. The γ-aminobutyric acid (GABA) alleviates salt stress damage during seeds germination of white clover associated with Na+/K.+ transportation, dehydrins accumulation, and stress-related genes expression in white clover. Int. J. Mol. Sci. 2018, 19, 2520. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Hu, J.; Zhang, Y.; Xie, X.J.; Knapp, A. Seed priming with brassinolide improves lucerne (Medicago sativa L.) seed germination and seedling growth in relation to physiological changes under salinity stress. Aust. J. Agric. Res. 2007, 58, 811–815. [Google Scholar] [CrossRef]
- Smith, D. Removing and analyzing total nonstructural carbohydrates from plant tissue. Jpn. J. Grassl. Sci. 1981, 17, 75–82. [Google Scholar]
- Tárrago, J.F.; Nicolás, G. Starch degradation in the cotyledons of germinating lentils. Plant Physiol. 1976, 58, 618–621. [Google Scholar] [CrossRef] [PubMed]
- Kishorekumar, A.; Jaleel, C.A.; Manivannan, P.; Sankar, B.; Sridharan, R.; Panneerselvam, R. Comparative effects of different triazole compounds on growth, photosynthetic pigments and carbohydrate metabolism of Solenostemon rotundifolius. Colloid Surf. B. 2007, 60, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Maren, M.; Sergi, M.B. Rapid and sensitive hormonal profiling of complex plant samples by liquid chromatography coupled to electrospray ionization tandem mass spectrometry. Plant Methods 2011, 7, 37. [Google Scholar] [Green Version]
- Blum, A. Osmotic adjustment and growth of barley genotypes under drought stress. Crop Sci. 1989, 29, 230–233. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Giannopolitis, C.N.; Ries, S.K. Superoxide dismutases: I. occurrence in higher plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Dhindsa, R.S.; Plumbdhindsa, P.; Thorpe, T.A. Leaf senescence: Correlated with increased leaves of membrane permeability and lipid peroxidation, and decreased levels of superoxide dismutase and catalase. J. Exp. Bot. 1981, 32, 93–101. [Google Scholar] [CrossRef]
- Elstner, E.F.; Heupel, A. Inhibition of nitrite formation from hydroxylammoniumchloride: A simple assay for superoxide dismutase. Anal. Biochem. 1976, 70, 616–620. [Google Scholar] [CrossRef]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: Protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Blum, A.; Ebercon, A. Cell membrane stability as a measure of drought and heat tolerance in wheat. Crop Sci. 1981, 21, 43–47. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, Y.; Zhang, X.Q.; Peng, Y.; Merewitz, E.; Ma, X.; Hang, L.K.; Yan, Y.H. The alteration of endogenous polyamines and phytohormones induced by exogenous application of spermidine regulate antioxidant metabolism, metallothionein and relevant genes conferring drought tolerance in white clover. Environ. Exp. Bot. 2016, 124, 22–38. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, Y.; Xu, Y.; Zhang, X.Q.; Peng, Y.; Ma, X.; Huang, L.K.; Yan, Y.H. Physiological and iTRAQ-based proteomic analyses reveal the function of spermidine on improving drought tolerance in white clover. J. Proteome Res. 2016, 15, 1563–1579. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NaCl (mM) | Germination Percentage | Germination Vigor | Germination Index | Mean Germination Speed | ||||
|---|---|---|---|---|---|---|---|---|
| NC | WS | NC | WS | NC | WS | NC | WS | |
| 0.00 | 98.0 ± 2.0 a | 36.7 ± 3.1 c | 92.7 ± 3.1 a | 18.0 ± 3.5 d | 43.90 ± 1.91 a | 6.96 ± 1.35 c | 1.31 ± 0.14 a | 3.75 ± 0.53 a |
| 0.25 | 96.7 ± 3.1 a | 42.7 ± 5.0 bc | 93.0 ± 5.3 a | 32.7 ± 2.3 c | 44.11 ± 2.56 a | 11.44 ± 1.43 bc | 1.26 ± 0.11 a | 2.46 ± 0.32 b |
| 0.50 | 97.3 ± 2.3 a | 46.0 ± 6.0 b | 92.7 ± 5.0 a | 40.7 ± 3.1 b | 44.81 ± 3.82 a | 11.88 ± 1.68 b | 1.28 ± 0.19 a | 2.26 ± 0.26 b |
| 1.00 | 98.0 ± 2.0 a | 58.7 ± 2.3 a | 92.7 ± 1.2 a | 47.3 ± 4.6 a | 45.43 ± 1.00 a | 17.95 ± 2.16 a | 1.22 ± 0.02 a | 2.09 ± 0.49 b |
| 2.50 | 97.3 ± 1.2 a | 50.0 ± 5.3 b | 93.0 ± 2.0 a | 40.0 ± 2.0 b | 46.31 ± 0.50 a | 13.83 ± 2.33 b | 1.16 ± 0.08 a | 2.44 ± 0.27 b |
| 5.00 | 96.7 ± 3.1 a | 40.7 ± 4.6 bc | 93.0 ± 5.3 a | 29.3 ± 3.1 c | 45.40 ± 2.15 a | 10.68 ± 4.97 c | 1.15 ± 0.08 a | 2.71 ± 0.46 b |
| 10.00 | 94.0 ± 2.0 a | 35.3 ± 2.3 c | 91.3 ± 2.3 a | 20.7 ± 4.2 d | 43.85 ± 0.46 a | 6.34 ± 0.86 c | 1.22 ± 0.06 a | 3.42 ± 0.30 a |
| Target Gene | Accession No. | Forward Primer (5′–3′) | Reverse Primer (5′–3′) | Tm (°C) |
|---|---|---|---|---|
| Cu/ZnSOD | JQ321597.1 | AACTGTGTACCACGAGGACTTC | AGACTAACAGGTGCTAACAACG | 58 |
| FeSOD | KP202173 | ACACGATTTCTCAGGGTTACGAC | GCGGCCAAGACTATCAGTTCCAT | 58 |
| MnSOD | JQ321598.1 | TAAGGGAACCTACCCGATAACT | CCAGGACCAAACGTCACCAAAG | 66 |
| CAT | JQ321596.1 | AACAGGACGGGAATAGCACG | ACCAGGTTCAGACACGGAGACA | 58 |
| POD | JQ321606.1 | CACTTGGTTTAGTTTTGTCGCC | AACACGGTCTTGTCTGCTACG | 64 |
| APX | JQ321599.1 | TAAAGATAGTCAACCCACCTCAACA | ACCAGTCTTGGGAAACAACGTA | 58 |
| MDHR | KP202172 | CCAACTGCCTAAAGCCACATCT | GAAGAAAGGAAACTAACGGAGCAT | 64 |
| DHAR | KP202171 | TGGTTACCTCCCGACCCTAT | TCTTACCAAGGAACTTTAGTCAGG | 58 |
| CytGR | JQ321602.1 | TAAACTTCCACTCCCTTTCTATCG | CTACAATATGGGTTGAGGACAGGT | 58 |
| Dehydrin b | GU443960.1 | TCCAGTCATCCAGCCTGTTG | CCAGCCACAACACTTGTCA | 60 |
| SK2 | GU443960.1 | TGGAACAGGAGTAACAACAGGTGGA | TGCCAGTTGAGAAAGTTGAGGTTGT | 58 |
| Y2K | JF748410.1 | AGCCACGCAACAAGGTTCTAA | TTGAGGATACGGGATGGGTG | 60 |
| Y2SK | GU443965.1 | GTGCGATGGAGATGCTGTTTG | CCTAATCCAACTTCAGGTTCAGC | 60 |
| β-Actin | JF968419 | TTACAATGAATTGCGTGTTG | AGAGGACAGCCTGAATGG | 58 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, Y.; Liang, L.; Cheng, B.; Dong, Y.; Wei, J.; Tian, X.; Peng, Y.; Li, Z. Pretreatment with NaCl Promotes the Seed Germination of White Clover by Affecting Endogenous Phytohormones, Metabolic Regulation, and Dehydrin-Encoded Genes Expression under Water Stress. Int. J. Mol. Sci. 2018, 19, 3570. https://doi.org/10.3390/ijms19113570
Cao Y, Liang L, Cheng B, Dong Y, Wei J, Tian X, Peng Y, Li Z. Pretreatment with NaCl Promotes the Seed Germination of White Clover by Affecting Endogenous Phytohormones, Metabolic Regulation, and Dehydrin-Encoded Genes Expression under Water Stress. International Journal of Molecular Sciences. 2018; 19(11):3570. https://doi.org/10.3390/ijms19113570
Chicago/Turabian StyleCao, Yiqin, Linlin Liang, Bizhen Cheng, Yue Dong, Jiaqi Wei, Xiaolan Tian, Yan Peng, and Zhou Li. 2018. "Pretreatment with NaCl Promotes the Seed Germination of White Clover by Affecting Endogenous Phytohormones, Metabolic Regulation, and Dehydrin-Encoded Genes Expression under Water Stress" International Journal of Molecular Sciences 19, no. 11: 3570. https://doi.org/10.3390/ijms19113570