Development of Biomarkers for Inhibition of SLC6A19 (B0AT1)—A Potential Target to Treat Metabolic Disorders



Abstract
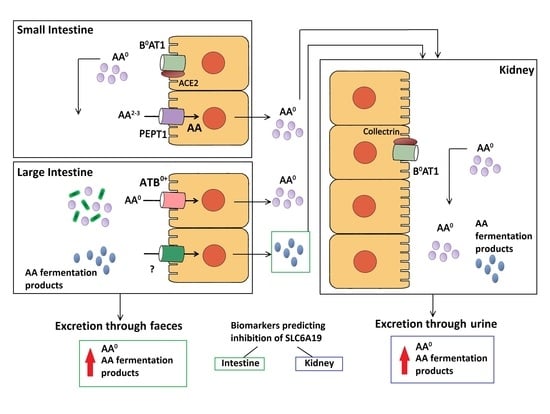
:
1. Introduction
2. Results
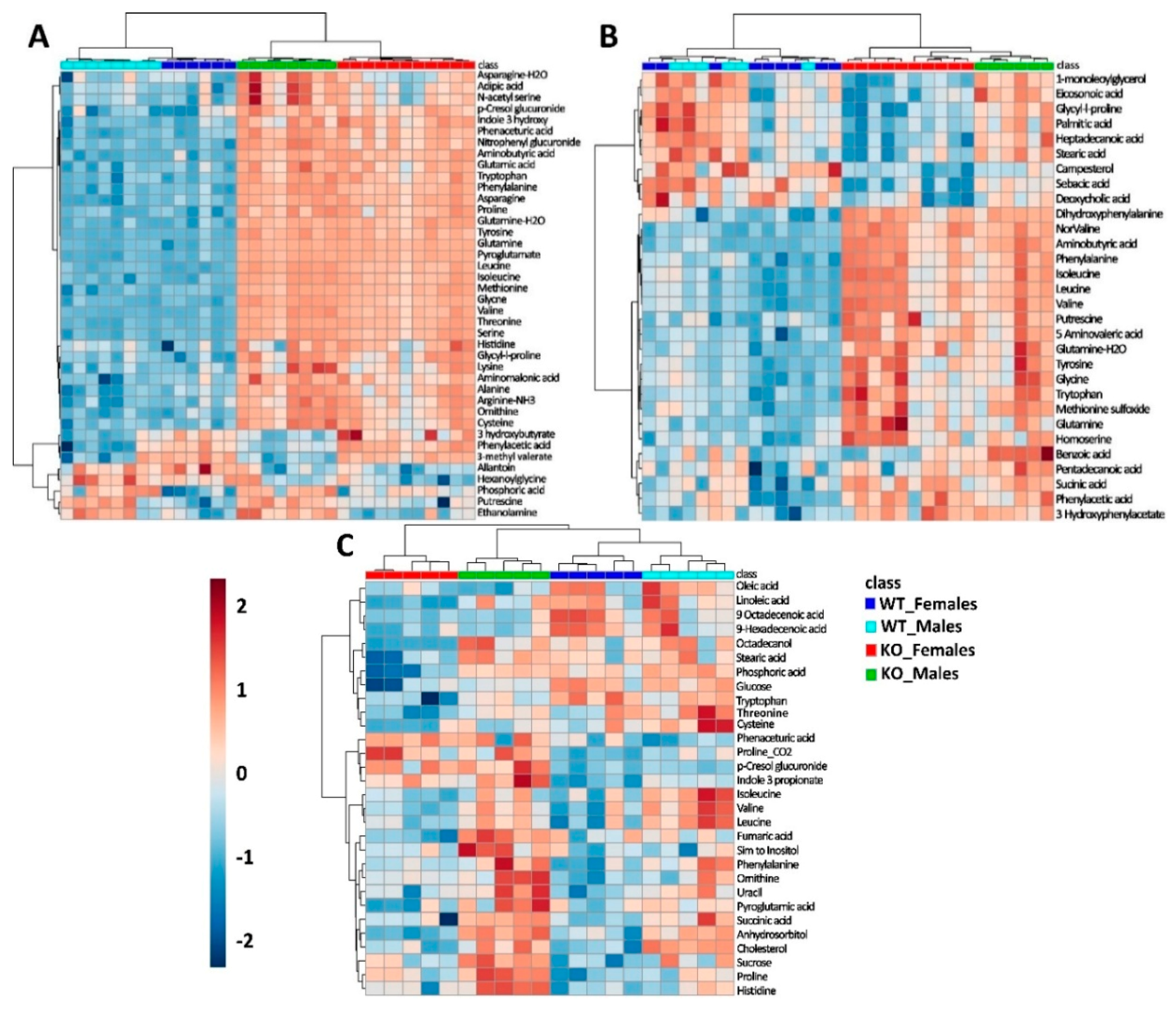
2.1. Overview of Untargeted Metabolomics of SLC6A19ko and SLC6A19wt Mice
2.2. Metabolic Profiling in Urine Samples
2.3. Metabolic Profiles in Faecal Samples
2.4. Metabolic Profiles of Plasma Samples
2.5. Assessment of Biomarkers for the In Vivo Detection of SLC6A19 Inhibition
2.6. Titration of SLC6A19 Transporter Activity
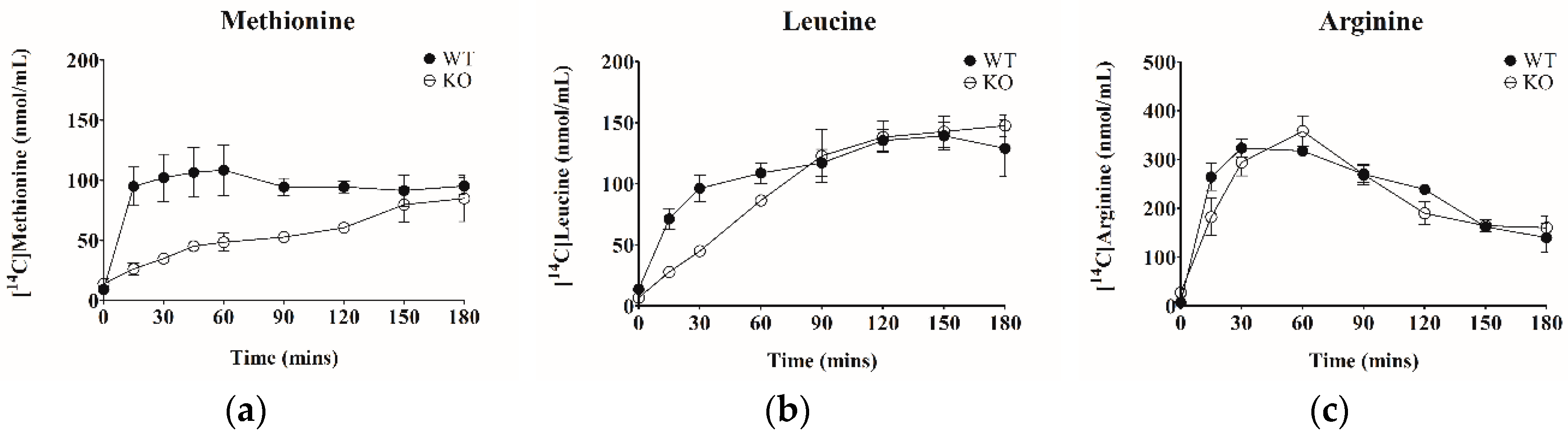
2.7. Evaluation of Biomarkers In vivo
3. Discussion
4. Materials and Methods
4.1. Animals and Sample Collection
4.2. Pharmacological Animal Studies
4.3. Radioactive Uptake Assay
4.4. Ammonia and Urea Assay
4.5. Untargeted Metabolomics of Urine, Faecal and Plasma Samples
4.6. Microarray Analysis
4.7. Data Analysis
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ACE2 | Angiotension converting enzyme 2 |
| AUC | Area under curve |
| B0AT1 | Broad neutral amino acid transporter |
| BCAA | Branched chain amino acids |
| FGF21 | Fibroblast growth factor 1 |
| GC-MS | Gas chromatography mass spectrometry |
| GCN2 | General control non derepressible 2 |
| GLP-1 | Glucagon like peptide 1 |
| GMD | Golm Metabolome Database |
| HCA | Heirarchial clustering analysis |
| mTORC1 | Mammalian target of rapamycin 1 |
| NIST | National Institute of Standards and Technology |
| NZO | Newzealand obese |
| PCA | Principal component analysis |
| QC | Quality control |
| RI | Retention index |
| ROC | Receiver operating characteristic |
References
- Mirzaei, H.; Suarez, J.A.; Longo, V.D. Protein and amino acid restriction, aging and disease: From yeast to humans. Trends Endocrinol. Metab. 2014, 25, 558–566. [Google Scholar] [CrossRef] [PubMed]
- Gallinetti, J.; Harputlugil, E.; Mitchell, J.R. Amino acid sensing in dietary-restriction-mediated longevity: Roles of signal-transducing kinases GCN2 and TOR. Biochem. J. 2013, 449, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Solon-Biet, S.M.; McMahon, A.C.; Ballard, J.W.; Ruohonen, K.; Wu, L.E.; Cogger, V.C.; Warren, A.; Huang, X.; Pichaud, N.; Melvin, R.G.; et al. The Ratio of Macronutrients, Not Caloric Intake, Dictates Cardiometabolic Health, Aging, and Longevity in Ad Libitum-Fed Mice. Cell Metab. 2014, 19, 418–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newgard, C.B.; An, J.; Bain, J.R.; Muehlbauer, M.J.; Stevens, R.D.; Lien, L.F.; Haqq, A.M.; Shah, S.H.; Arlotto, M.; Slentz, C.A.; et al. A branched-chain amino acid-related metabolic signature that differentiates obese and lean humans and contributes to insulin resistance. Cell. Metab. 2009, 9, 311–326. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.J.; Larson, M.G.; Vasan, R.S.; Cheng, S.; Rhee, E.P.; McCabe, E.; Lewis, G.D.; Fox, C.S.; Jacques, P.F.; Fernandez, C.; et al. Metabolite profiles and the risk of developing diabetes. Nat. Med. 2011, 17, 448–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giesbertz, P.; Daniel, H. Branched-chain amino acids as biomarkers in diabetes. Curr. Opin. Clin. Nutr. Metab. Care 2016, 19, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Wurtz, P.; Soininen, P.; Kangas, A.J.; Ronnemaa, T.; Lehtimaki, T.; Kahonen, M.; Viikari, J.S.; Raitakari, O.T.; Ala-Korpela, M. Branched-chain and aromatic amino acids are predictors of insulin resistance in young adults. Diabetes Care 2013, 36, 648–655. [Google Scholar] [CrossRef] [PubMed]
- Fontana, L.; Cummings, N.E.; Arriola Apelo, S.I.; Neuman, J.C.; Kasza, I.; Schmidt, B.A.; Cava, E.; Spelta, F.; Tosti, V.; Syed, F.A.; et al. Decreased Consumption of Branched-Chain Amino Acids Improves Metabolic Health. Cell Rep. 2016, 16, 520–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cummings, N.E.; Williams, E.M.; Kasza, I.; Konon, E.N.; Schaid, M.D.; Schmidt, B.A.; Poudel, C.; Sherman, D.S.; Yu, D.; Arriola Apelo, S.I.; et al. Restoration of metabolic health by decreased consumption of branched-chain amino acids. J. Physiol. 2017, 596, 623–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laeger, T.; Henagan, T.M.; Albarado, D.C.; Redman, L.M.; Bray, G.A.; Noland, R.C.; Münzberg, H.; Hutson, S.M.; Gettys, T.W.; Schwartz, M.W.; et al. FGF21 is an endocrine signal of protein restriction. J. Clin. Investig. 2014, 124, 3913–3922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maida, A.; Chan, J.S.K.; Sjoberg, K.A.; Zota, A.; Schmoll, D.; Kiens, B.; Herzig, S.; Rose, A.J. Repletion of branched chain amino acids reverses mTORC1 signaling but not improved metabolism during dietary protein dilution. Mol. Metab. 2017, 6, 873–881. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.M.; Morrison, C.D. Dietary branched chain amino acids and metabolic health: When less is more. J. Physiol. 2018, 596, 555–556. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.C.; Kaya, A.; Gladyshev, V.N. Methionine restriction and life-span control. Ann. N. Y. Acad. Sci. 2016, 1363, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Latimer, M.N.; Freij, K.W.; Cleveland, B.M.; Biga, P.R. Physiological and Molecular Mechanisms of Methionine Restriction. Front. Endocrinol. 2018, 9, 217. [Google Scholar] [CrossRef] [PubMed]
- Miller Richard, A.; Buehner, G.; Chang, Y.; Harper James, M.; Sigler, R.; Smith-Wheelock, M. Methionine-deficient diet extends mouse lifespan, slows immune and lens aging, alters glucose, T4, IGF-I and insulin levels, and increases hepatocyte MIF levels and stress resistance. Aging Cell 2005, 4, 119–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wanders, D.; Stone, K.P.; Forney, L.A.; Cortez, C.C.; Dille, K.N.; Simon, J.; Xu, M.; Hotard, E.C.; Nikonorova, I.A.; Pettit, A.P.; et al. Role of GCN2-Independent Signaling Through a Noncanonical PERK/NRF2 Pathway in the Physiological Responses to Dietary Methionine Restriction. Diabetes 2016, 65, 1499. [Google Scholar] [CrossRef] [PubMed]
- Bass, J. Forever (FGF) 21. Nat. Med. 2013, 19, 1090–1092. [Google Scholar] [CrossRef] [PubMed]
- Maida, A.; Zota, A.; Vegiopoulos, A.; Appak-Baskoy, S.; Augustin, H.G.; Heikenwalder, M.; Herzig, S.; Rose, A.J. Dietary protein dilution limits dyslipidemia in obesity through FGF21-driven fatty acid clearance. J. Nutr. Biochem. 2018, 57, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Talukdar, S.; Zhou, Y.; Li, D.; Rossulek, M.; Dong, J.; Somayaji, V.; Weng, Y.; Clark, R.; Lanba, A.; Owen, B.M.; et al. A Long-Acting FGF21 Molecule, PF-05231023, Decreases Body Weight and Improves Lipid Profile in Non-human Primates and Type 2 Diabetic Subjects. Cell Metab. 2016, 23, 427–440. [Google Scholar] [CrossRef] [PubMed]
- Broer, S.; Broer, A. Amino acid homeostasis and signalling in mammalian cells and organisms. Biochem. J. 2017, 474, 1935–1963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laferrere, B.; Reilly, D.; Arias, S.; Swerdlow, N.; Gorroochurn, P.; Bawa, B.; Bose, M.; Teixeira, J.; Stevens, R.D.; Wenner, B.R.; et al. Differential metabolic impact of gastric bypass surgery versus dietary intervention in obese diabetic subjects despite identical weight loss. Sci. Transl. Med. 2011, 3, 80re82. [Google Scholar] [CrossRef] [PubMed]
- Friedman, M.N.; Sancetta, A.J.; Magovern, G.J. The amelioration of diabetes mellitus following subtotal gastrectomy. Surg. Gynecol. Obstet. 1955, 100, 201–204. [Google Scholar] [PubMed]
- Magkos, F.; Bradley, D.; Schweitzer, G.G.; Finck, B.N.; Eagon, J.C.; Ilkayeva, O.; Newgard, C.B.; Klein, S. Effect of Roux-en-Y gastric bypass and laparoscopic adjustable gastric banding on branched-chain amino acid metabolism. Diabetes 2013, 62, 2757–2761. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Rose, A.J.; Sijmonsma, T.P.; Broer, A.; Pfenninger, A.; Herzig, S.; Schmoll, D.; Broer, S. Mice lacking neutral amino acid transporter B(0)AT1 (Slc6a19) have elevated levels of FGF21 and GLP-1 and improved glycaemic control. Mol. Metab. 2015, 4, 406–417. [Google Scholar] [CrossRef] [PubMed]
- Reimann, F.; Ward, P.S.; Gribble, F.M. Signaling Mechanisms Underlying the Release of Glucagon-Like Peptide 1. Diabetes 2006, 55, S78. [Google Scholar] [CrossRef]
- Kyrou, I.; Weickert, M.O.; Gharanei, S.; Randeva, H.S.; Tan, B.K. Fibroblast growth factors: New insights, new targets in the management of diabetes. Minerva Endocrinol. 2017, 42, 248–270. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Q.; Johnsson, K.; Grandy, S.; Kostev, K. Treatment Outcomes and Tolerability Following Initiation of GLP-1 Receptor Agonists Among Type 2 Diabetes Patients in Primary Care Practices in Germany. J. Diabetes Sci. Technol. 2016, 11, 272–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broer, S. Amino Acid Transporters as Disease Modifiers and Drug Targets. SLAS Discov. 2018, 23, 303–320. [Google Scholar] [CrossRef] [PubMed]
- Belanger, A.M.; Przybylska, M.; Gefteas, E.; Furgerson, M.; Geller, S.; Kloss, A.; Cheng, S.H.; Zhu, Y.; Yew, N.S. Inhibiting neutral amino acid transport for the treatment of phenylketonuria. JCI Insight 2018, 3. [Google Scholar] [CrossRef] [PubMed]
- Broer, S. The role of the neutral amino acid transporter B0AT1 (SLC6A19) in Hartnup disorder and protein nutrition. IUBMB Life 2009, 61, 591–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Q.; Shah, N.; Broer, A.; Fairweather, S.; Jiang, Y.; Schmoll, D.; Corry, B.; Broer, S. Identification of novel inhibitors of the amino acid transporter B(0) AT1 (SLC6A19), a potential target to induce protein restriction and to treat type 2 diabetes. Br. J. Pharmacol. 2017, 174, 468–482. [Google Scholar] [CrossRef] [PubMed]
- Pochini, L.; Seidita, A.; Sensi, C.; Scalise, M.; Eberini, I.; Indiveri, C. Nimesulide binding site in the B0AT1 (SLC6A19) amino acid transporter. Mechanism of inhibition revealed by proteoliposome transport assay and molecular modelling. Biochem. Pharmacol. 2014, 89, 422–430. [Google Scholar] [CrossRef] [PubMed]
- Floegel, A.; Stefan, N.; Yu, Z.; Muhlenbruch, K.; Drogan, D.; Joost, H.G.; Fritsche, A.; Haring, H.U.; Hrabe de Angelis, M.; Peters, A.; et al. Identification of serum metabolites associated with risk of type 2 diabetes using a targeted metabolomic approach. Diabetes 2013, 62, 639–648. [Google Scholar] [CrossRef] [PubMed]
- Chan, E.C.; Pasikanti, K.K.; Nicholson, J.K. Global urinary metabolic profiling procedures using gas chromatography-mass spectrometry. Nat. Protoc. 2011, 6, 1483–1499. [Google Scholar] [CrossRef] [PubMed]
- Broer, A.; Juelich, T.; Vanslambrouck, J.M.; Tietze, N.; Solomon, P.S.; Holst, J.; Bailey, C.G.; Rasko, J.E.; Broer, S. Impaired nutrient signaling and body weight control in a Na+ neutral amino acid cotransporter (Slc6a19)-deficient mouse. J. Biol. Chem. 2011, 286, 26638–26651. [Google Scholar] [CrossRef] [PubMed]
- Broer, S.; Cavanaugh, J.A.; Rasko, J.E. Neutral amino acid transport in epithelial cells and its malfunction in Hartnup disorder. Biochem. Soc. Trans. 2005, 33, 233–236. [Google Scholar] [CrossRef] [PubMed]
- Dulloo, A.G.; Miller, D.S. The effect of parasympathetic drugs on energy expenditure: Relevance to the autonomic hypothesis. Can. J. Physiol. Pharmacol. 1986, 64, 586–591. [Google Scholar] [CrossRef] [PubMed]
- Brachs, S.; Grune, J.; Kintscher, U.; Brachs, M.; Bröer, S.; Spranger, J. Slc6a19 deficiency improves insulin sensitivity and preserves cardiac function in mice during diet-induced obesity. Diabetol. Stoffwechs. 2018, 13, FV 4. [Google Scholar] [CrossRef]
- Matthews, D.M. Intestinal absorption of amino acids and peptides. Proc. Nutr. Soc. 2007, 31, 171–177. [Google Scholar] [CrossRef]
- Toden, S.; Bird, A.R.; Topping, D.L.; Conlon, M.A. Resistant starch attenuates colonic DNA damage induced by higher dietary protein in rats. Nutr. Cancer 2005, 51, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Singer, D.; Camargo, S.M.; Ramadan, T.; Schafer, M.; Mariotta, L.; Herzog, B.; Huggel, K.; Wolfer, D.; Werner, S.; Penninger, J.M.; et al. Defective intestinal amino acid absorption in Ace2 null mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 303, G686–G695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scriver, C.R.; Mahon, B.; Levy, H.L.; Clow, C.L.; Reade, T.M.; Kronick, J.; Lemieux, B.; Laberge, C. The Hartnup phenotype: Mendelian transport disorder, multifactorial disease. Am. J. Hum. Genet. 1987, 40, 401–412. [Google Scholar] [PubMed]
- Newgard, C.B. Metabolomics and Metabolic Diseases: Where Do We Stand? Cell Metab. 2017, 25, 43–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashimoto, T.; Perlot, T.; Rehman, A.; Trichereau, J.; Ishiguro, H.; Paolino, M.; Sigl, V.; Hanada, T.; Hanada, R.; Lipinski, S.; et al. ACE2 links amino acid malnutrition to microbial ecology and intestinal inflammation. Nature 2012, 487, 477–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bone, E.; Tamm, A.; Hill, M. The production of urinary phenols by gut bacteria and their possible role in the causation of large bowel cancer. Am. J. Clin. Nutr. 1976, 29, 1448–1454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lesaffer, G.; De Smet, R.; Belpaire, F.M.; Van Vlem, B.; Van Hulle, M.; Cornelis, R.; Lameire, N.; Vanholder, R. Urinary excretion of the uraemic toxin p-cresol in the rat: Contribution of glucuronidation to its metabolization. Nephrol. Dial. Transplant. 2003, 18, 1299–1306. [Google Scholar] [CrossRef] [PubMed]
- Bammens, B.; Verbeke, K.; Vanrenterghem, Y.; Evenepoel, P. Evidence for impaired assimilation of protein in chronic renal failure. Kidney Int. 2003, 64, 2196–2203. [Google Scholar] [CrossRef] [PubMed]
- Lopes, T.; Geloneze, B.; Pareja, J.; Calixto, A.; Ferreira, M.; Marsaioli, A. Blood Metabolome Changes before and After Bariatric Surgery: A 1 H NMR-Based Clinical Investigation. OMICS 2015, 19, 318–327. [Google Scholar] [CrossRef] [PubMed]
- Li, J.V.; Ashrafian, H.; Bueter, M.; Kinross, J.; Sands, C.; le Roux, C.W.; Bloom, S.R.; Darzi, A.; Athanasiou, T.; Marchesi, J.R.; et al. Metabolic surgery profoundly influences gut microbial-host metabolic cross-talk. Gut 2011, 60, 1214–1223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koppe, L.; Alix, P.M.; Croze, M.L.; Chambert, S.; Vanholder, R.; Glorieux, G.; Fouque, D.; Soulage, C.O. p-Cresyl glucuronide is a major metabolite of p-cresol in mouse: In contrast to p-cresyl sulphate, p-cresyl glucuronide fails to promote insulin resistance. Nephrol. Dial. Transplant. 2017, 32, 2000–2009. [Google Scholar] [CrossRef] [PubMed]
- Dunn, W.B.; Broadhurst, D.; Begley, P.; Zelena, E.; Francis-McIntyre, S.; Anderson, N.; Brown, M.; Knowles, J.D.; Halsall, A.; Haselden, J.N.; et al. Procedures for large-scale metabolic profiling of serum and plasma using gas chromatography and liquid chromatography coupled to mass spectrometry. Nat. Protoc. 2011, 6, 1060–1083. [Google Scholar] [CrossRef] [PubMed]
- Goodacre, R. Metabolomics of a Superorganism. J. Nutr. 2007, 137, 259S–266S. [Google Scholar] [CrossRef] [PubMed]
- Metges, C.C. Contribution of microbial amino acids to amino acid homeostasis of the host. J. Nutr. 2000, 130, 1857S–1864S. [Google Scholar] [CrossRef] [PubMed]
- Lynch, C.J.; Adams, S.H. Branched-chain amino acids in metabolic signalling and insulin resistance. Nat. Rev. Endocrinol. 2014, 10, 723–736. [Google Scholar] [CrossRef] [PubMed]
- Tochikubo, O.; Nakamura, H.; Jinzu, H.; Nagao, K.; Yoshida, H.; Kageyama, N.; Miyano, H. Weight loss is associated with plasma free amino acid alterations in subjects with metabolic syndrome. Nutr. Diabetes 2016, 6, e197. [Google Scholar] [CrossRef] [PubMed]
- Larson, K.R.; Russo, K.A.; Fang, Y.; Mohajerani, N.; Goodson, M.L.; Ryan, K.K. Sex Differences in the Hormonal and Metabolic Response to Dietary Protein Dilution. Endocrinology 2017, 158, 3477–3487. [Google Scholar] [CrossRef] [PubMed]
- Bachmanov, A.A.; Reed, D.R.; Beauchamp, G.K.; Tordoff, M.G. Food intake, water intake, and drinking spout side preference of 28 mouse strains. Behav. Genet. 2002, 32, 435–443. [Google Scholar] [CrossRef] [PubMed]
- Creedon, C.T.; Verhulst, P.J.; Choi, K.M.; Mason, J.E.; Linden, D.R.; Szurszewski, J.H.; Gibbons, S.J.; Farrugia, G. Assessment of gastric emptying in non-obese diabetic mice using a [13C]-octanoic acid breath test. J. Vis. Exp. 2013, 23, e50301. [Google Scholar] [CrossRef]
- Carroll, A.J.; Badger, M.R.; Harvey Millar, A. The MetabolomeExpress Project: Enabling web-based processing, analysis and transparent dissemination of GC/MS metabolomics datasets. BMC Bioinform. 2010, 11, 376. [Google Scholar] [CrossRef] [PubMed]
- Chetwynd, A.J.; Abdul-Sada, A.; Holt, S.G.; Hill, E.M. Use of a pre-analysis osmolality normalisation method to correct for variable urine concentrations and for improved metabolomic analyses. J. Chromatogr. A 2016, 1431, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.A.; Want, E.J.; O'Maille, G.; Abagyan, R.; Siuzdak, G. XCMS: Processing mass spectrometry data for metabolite profiling using nonlinear peak alignment, matching, and identification. Anal. Chem. 2006, 78, 779–787. [Google Scholar] [CrossRef] [PubMed]
- Tautenhahn, R.; Patti, G.J.; Rinehart, D.; Siuzdak, G. XCMS Online: A web-based platform to process untargeted metabolomic data. Anal. Chem. 2012, 84, 5035–5039. [Google Scholar] [CrossRef] [PubMed]
- Kopka, J.; Schauer, N.; Krueger, S.; Birkemeyer, C.; Usadel, B.; Bergmuller, E.; Dormann, P.; Weckwerth, W.; Gibon, Y.; Stitt, M.; et al. [email protected]: The Golm Metabolome Database. Bioinformatics 2005, 21, 1635–1638. [Google Scholar] [CrossRef] [PubMed]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | Body Weight (g) | Urea (Plasma) mmol/L | Urea (Urine) mmol/L | Ammonia (Plasma) mg/mL | Ammonia (Urine) µg/mL | Ammonia (Faeces) mg/mL | |
|---|---|---|---|---|---|---|---|
| Females | ko | 26.1 ± 2.5 | 6.5 ± 1.5 | 568 ± 23.6 | 0.7 ± 0.2 | 7.5 ± 0.2 | 11.2 ± 2.8 |
| wt | 29.4 ± 5.2 | 8.5 ± 2.1 | 423 ± 56.8 | 0.7 ± 0.1 | 7.4 ± 0.1 | 5.8 ± 1.5 | |
| Males | ko | 22.9 ± 4.2 | 7.1 ± 2.6 | 618 ± 30.3 | 0.3 ± 0.4 | 4.9 ± 0.1 | 9.2 ± 1.8 |
| wt | 27.9 ±4.6 | 8.2 ± 2.9 | 566 ± 19.9 | 0.3 ± 0.1 | 4.8 ± 0.2 | 3.3 ± 0.8 |
| Metabolite | RI | Quant Ion (m/z) | Retention Time (min) | ko/wt | Females | Males | QC %RSD | ID Level | |||
|---|---|---|---|---|---|---|---|---|---|---|---|
| FC | p Value | FC | p Value | FC | p Value | ||||||
| Serine | 1350 | 204.1 | 7.82 | 211.44 | 1.95 × 10−7 | 173.01 | 1.97 × 10−4 | 264.67 | 9.31 × 10−4 | 11.6 | 1 |
| Isoleucine | 1288 | 158.1 | 7.27 | 194.89 | 6.21 × 10−8 | 148.17 | 1.02 × 10−4 | 258.11 | 4.17 × 10−4 | 13.3 | 1 |
| Threonine | 1375 | 291.2 | 8.04 | 192.66 | 1.84 × 10−9 | 178.06 | 1.77 × 10−5 | 213.14 | 3.99 × 10−4 | 8.84 | 1 |
| Leucine | 1261 | 158.1 | 7.05 | 191.17 | 2.17 × 10−8 | 195.86 | 2.06 × 10−5 | 219.21 | 9.81 × 10−5 | 33.4 | 1 |
| Methionine | 1515 | 176.1 | 9.25 | 184.73 | 1.05 × 10−8 | 173.84 | 4.87 × 10−5 | 196.27 | 6.61 × 10−4 | 9.4 | 1 |
| Glutamine | 1723 | 156.0 | 10.81 | 146.47 | 3.63 × 10−11 | 228.76 | 1.19 × 10−6 | 121.57 | 2.09 × 10−4 | 12.1 | 1 |
| Valine | 1206 | 144.1 | 6.57 | 107.94 | 1.82 × 10−7 | 81.72 | 4.17 × 10−5 | 143.93 | 7.04 × 10−4 | 10.5 | 1 |
| Glycine | 1303 | 174.1 | 7.41 | 91.06 | 1.06 × 10−8 | 71.84 | 4.41 × 10−6 | 118.73 | 4.38 × 10−5 | 25.5 | 1 |
| Phenylalanine | 1630 | 192.1 | 10.13 | 84.99 | 4.24 × 10−7 | 57.00 | 5.02 × 10−5 | 123.75 | 6.26 × 10−4 | 9.0 | 1 |
| Tyrosine | 1931 | 218.1 | 12.34 | 70.60 | 2.04 × 10−8 | 61.11 | 3.35 × 10−5 | 83.72 | 6.62 × 10−4 | 9.9 | 1 |
| Proline | 1298 | 142.1 | 7.28 | 57.90 | 2.34 × 10−6 | 57.05 | 4.11 × 10−4 | 63.12 | 3.40 × 10−3 | 35.7 | 1 |
| Aminobutyric acid | 1161 | 130.1 | 6.17 | 56.53 | 3.81 × 10−8 | 31.51 | 1.03 × 10−4 | 120.84 | 1.11 × 10−3 | 9.5 | 1 |
| Asparagine | 1668 | 231.1 | 10.40 | 42.83 | 2.29 × 10−9 | 33.20 | 2.73 × 10−4 | 53.77 | 3.84 × 10−4 | 6.4 | 1 |
| Cystine | 2289 | 266.1 | 14.51 | 36.43 | 3.56 × 10−4 | 22.89 | 5.40 × 10−3 | 54.72 | 1.04 × 10−2 | 30.8 | 2 |
| Nitrophenyl Glucuronide | 2344 | 375.2 | 14.80 | 35.02 | 2.99 × 10−6 | 30.66 | 2.61 × 10−4 | 43.74 | 2.19 × 10−3 | 7.9 | 3 |
| Pyroglutamate | 1523 | 156.0 | 9.31 | 32.84 | 2.00 × 10−15 | 30.31 | 7.63 × 10−9 | 35.14 | 4.62 × 10−6 | 13.6 | 2 |
| Tryptophan | 2216 | 202.1 | 14.13 | 26.01 | 2.82 × 10−7 | 29.54 | 3.95 × 10−4 | 26.53 | 1.50 × 10−3 | 16.2 | 2 |
| Aminomalonic acid | 1461 | 320.1 | 8.79 | 17.40 | 1.19 × 10−2 | 10.32 | 1.90 × 10−2 | 27.11 | 5.15 × 10−3 | 27.6 | 2 |
| Alanine | 1360 | 262.1 | 7.83 | 17.39 | 8.68 × 10−7 | 10.94 | 6.45 × 10−4 | 27.21 | 9.58 × 10−4 | 14.2 | 2 |
| Ornithine | 1609 | 142.1 | 9.96 | 17.05 | 7.26 × 10−7 | 11.50 | 3.04 × 10−4 | 24.80 | 1.17 × 10−3 | 31.7 | 2 |
| Phenaceturic acid | 1896 | 250.1 | 12.10 | 14.05 | 1.48 × 10−8 | 10.33 | 4.10 × 10−5 | 18.88 | 4.06 × 10−4 | 10.0 | 3 |
| Asparagine ester | 1618 | 216.1 | 10.03 | 12.80 | 4.86 × 10−4 | 16.42 | 1.98 × 10−2 | 12.93 | 3.24 × 10−2 | 31.1 | 2 |
| Cresol glucuronide | 2405 | 375.2 | 15.21 | 8.80 | 9.91 × 10−6 | 5.51 | 8.41 × 10−4 | 13.93 | 1.75 × 10−3 | 5.9 | 3 |
| Glutamic acid | 1614 | 246.1 | 9.99 | 7.38 | 2.94 × 10−7 | 5.74 | 1.49 × 10−5 | 9.44 | 6.22 × 10−3 | 9.6 | 1 |
| 5 Hydroxy Indole | 1719 | 277.1 | 10.79 | 5.89 | 1.02 × 10−7 | 5.70 | 2.88 × 10−5 | 5.79 | 6.50 × 10−3 | 26.1 | 2 |
| Lysine | 1908 | 230.1 | 12.19 | 5.18 | 3.24 × 10−4 | 3.50 | 2.19 × 10−3 | 7.49 | 1.74 × 10−2 | 6.3 | 1 |
| Histidine | 1915 | 154.1 | 12.23 | 4.05 | 6.41 × 10−5 | 5.71 | 1.40 × 10−3 | 3.18 | 2.66 × 10−2 | 31.9 | 1 |
| Glutaric acid 3 hydroxy | 1596 | 247.1 | 9.86 | 2.82 | 2.01 × 10−3 | 2.30 | 2.05 × 10−2 | 3.17 | 1.27 × 10−1 | 12.7 | 2 |
| Serine N-acetyl | 1499 | 261.1 | 9.13 | 2.22 | 2.30 × 10−3 | 1.82 | 1.74 × 10−2 | 2.86 | 1.10 × 10−2 | 11.1 | 2 |
| Adipic acid | 1498 | 186.0 | 9.13 | 2.19 | 2.79 × 10−3 | 1.94 | 1.33 × 10−2 | 2.70 | 1.46 × 10−2 | 8.7 | 2 |
| Alanine beta | 1423 | 174.1 | 8.46 | 1.65 | 4.35 × 10−2 | 1.10 | 8.15 × 10−1 | 2.48 | 3.17 × 10−2 | 34.3 | 1 |
| 3hydroxyphenyl acetic acid | 1614 | 164.0 | 9.98 | 1.63 | 1.27 × 10−2 | 1.38 | 3.18 × 10−1 | 1.90 | 3.30 × 10−2 | 13.1 | 2 |
| Putrescine | 1739 | 174.1 | 10.93 | 0.98 | 9.64 × 10−1 | 2.36 | 7.55 × 10−3 | 1.12 | 8.10 × 10−1 | 12.5 | 1 |
| Succinic acid | 1311 | 247.1 | 7.47 | 0.73 | 2.00 × 10−1 | 0.80 | 5.86 × 10−1 | 0.54 | 1.80 × 10−2 | 20.8 | 1 |
| Quinolone 2 carboxylic acid | 2262 | 406.2 | 14.37 | 0.55 | 1.17 × 10−2 | 0.57 | 1.50 × 10−1 | 0.62 | 3.86 × 10−2 | 21.9 | 2 |
| Butyrylglycine | 1447 | 158.1 | 8.66 | 0.52 | 1.12 × 10−2 | 0.38 | 6.22 × 10−3 | 0.70 | 8.70 × 10−2 | 15.8 | 2 |
| Thymine | 1397 | 270.1 | 8.24 | 0.48 | 5.57 × 10−3 | 0.47 | 1.46 × 10−1 | 0.56 | 4.23 × 10−2 | 30.2 | 2 |
| Hexanoylglycine | 1644 | 158.1 | 10.23 | 0.39 | 2.96 × 10−3 | 0.40 | 1.26 × 10−2 | 0.45 | 1.72 × 10−2 | 22.9 | 2 |
| Allantoin | 1876 | 188.0 | 11.96 | 0.26 | 5.08 × 10−3 | 0.19 | 7.09 × 10−2 | 0.37 | 4.23 × 10−2 | 29.2 | 2 |
| Taurine | 1672 | 326.1 | 10.44 | 0.24 | 6.57 × 10−3 | 0.28 | 1.84 × 10−1 | 0.13 | 2.07 × 10−3 | 14.7 | 2 |
| Transporter | Acronym | SLC6A19ko | SLC6A19wt | Fold Change (ko/wt) | p Value |
|---|---|---|---|---|---|
| SLC7A9 | b0,+AT | 14.1 | 14 | 1.1 | 0.5 |
| SLC7A7 | y+LAT1/4F2hc | 14.1 | 15 | 0.94 | 0.06 |
| SLC1A1 | EAAT3 | 12.9 | 13.5 | 0.96 | 0.05 |
| SLC3A1 | rBAT | 14.3 | 14.5 | 0.99 | 0.37 |
| Metabolite ID | RI | Quant Ion (m/z) | Retention Tine (min) | ko/wt | Females | Males | QC %RSD | ID Level | |||
|---|---|---|---|---|---|---|---|---|---|---|---|
| FC | p Value | FC | p Value | FC | p Value | ||||||
| Acetylglutamine | 1720 | 227.07 | 10.81 | 25.86 | 1.34 × 10−2 | 37.20 | 2.26 × 10−2 | 7.29 | 4.34 × 10−2 | 40.2 | 3 |
| Aminobutanoic acid | 1163 | 130.05 | 6.15 | 24.46 | 1.60 × 10−5 | 31.85 | 1.14 × 10−3 | 18.80 | 1.23 × 10−2 | 11.9 | 1 |
| Leucine | 1264 | 158.10 | 7.05 | 20.34 | 1.55 × 10−5 | 27.80 | 5.86 × 10−4 | 13.35 | 2.01 × 10−2 | 9.3 | 1 |
| Norvaline | 1230 | 144.05 | 6.76 | 16.67 | 1.85 × 10−6 | 17.77 | 1.97 × 10−4 | 14.70 | 6.58 × 10−3 | 8.4 | 3 |
| Valine | 1207 | 144.10 | 6.55 | 13.98 | 9.24 × 10−5 | 17.99 | 1.74 × 10−3 | 9.94 | 4.13 × 10−2 | 6.7 | 1 |
| Putrescine | 1740 | 174.04 | 10.93 | 11.90 | 6.43 × 10−3 | 16.48 | 2.64 × 10−2 | 6.58 | 1.18 × 10−1 | 15.7 | 1 |
| Phenylalanine | 1633 | 192.07 | 10.13 | 11.52 | 6.21 × 10−5 | 16.57 | 8.63 × 10−4 | 7.44 | 5.16 × 10−2 | 13.5 | 1 |
| Dihydroxyphenyl-alanine | 2086 | 368.05 | 13.32 | 11.37 | 9.60 × 10−6 | 9.41 | 1.38 × 10−3 | 17.68 | 2.63 × 10−3 | 39.8 | 3 |
| Isoleucine | 1287 | 158.10 | 7.25 | 10.07 | 2.43 × 10−4 | 12.81 | 3.10 × 10−3 | 7.37 | 5.94 × 10−2 | 10.8 | 1 |
| Tyrosine | 1931 | 218.08 | 12.34 | 9.63 | 3.06 × 10−3 | 10.78 | 6.90 × 10−3 | 8.56 | 1.51 × 10−1 | 19.1 | 1 |
| Tryptophan | 2216 | 202.06 | 14.13 | 7.99 | 2.65 × 10−3 | 10.25 | 2.71 × 10−2 | 5.72 | 4.38 × 10−2 | 34.8 | 1 |
| Glycine | 1303 | 174.06 | 7.39 | 6.89 | 3.49 × 10−3 | 8.42 | 2.76 × 10−2 | 5.67 | 8.87 × 10−2 | 13.8 | 1 |
| Tyramine | 1912 | 338.03 | 12.21 | 6.20 | 1.42 × 10−3 | 10.40 | 3.05 × 10−2 | 4.40 | 1.35 × 10−2 | 15.2 | 2 |
| Methionine sulfoxide | 1782 | 128.00 | 11.25 | 5.98 | 1.25 × 10−3 | 7.56 | 2.49 × 10−2 | 4.52 | 8.16 × 10−3 | 9.9 | 2 |
| Homoserine | 1441 | 218.10 | 8.60 | 5.79 | 4.49 × 10−4 | 7.56 | 4.30 × 10−3 | 3.56 | 2.66 × 10−2 | 39.3 | 2 |
| Homocystine ester | 1857 | 128.00 | 11.80 | 5.62 | 3.14 × 10−3 | 7.26 | 3.42 × 10−2 | 3.99 | 1.11 × 10−2 | 11.3 | 2 |
| 5 amino valeric acid | 1633 | 174.07 | 10.11 | 4.44 | 4.11 × 10−6 | 5.70 | 7.35 × 10−5 | 3.14 | 3.38 × 10−2 | 9.1 | 2 |
| 3 hydroxyphenyl-acetic acid | 1613 | 163.97 | 9.98 | 4.05 | 2.02 × 10−4 | 4.26 | 9.13 × 10−3 | 3.72 | 2.59 × 10−3 | 12.5 | 2 |
| Threonine | 1377 | 291.09 | 8.03 | 3.68 | 2.69 × 10−2 | 4.45 | 1.02 × 10−1 | 2.81 | 1.18 × 10−1 | 18.7 | 1 |
| Ornithine | 1810 | 142.02 | 11.46 | 3.59 | 2.41 × 10−3 | 4.79 | 5.85 × 10−3 | 1.97 | 2.08 × 10−1 | 4.9 | 1 |
| Phenylacetic acid | 1302 | 163.99 | 7.41 | 3.12 | 5.82 × 10−4 | 4.26 | 2.26 × 10−3 | 2.13 | 1.45 × 10−1 | 20.2 | 3 |
| Lysine | 1908 | 174.05 | 12.19 | 2.96 | 1.03 × 10−2 | 4.20 | 3.53 × 10−2 | 1.75 | 8.27 × 10−2 | 13.1 | 1 |
| Serine | 1352 | 204.08 | 7.81 | 2.94 | 3.34 × 10−2 | 3.84 | 9.41 × 10−2 | 2.16 | 2.35 × 10−1 | 15.8 | 1 |
| Succinic acid | 1309 | 247.02 | 7.45 | 2.66 | 6.86 × 10−6 | 3.39 | 2.22 × 10−4 | 1.96 | 2.40 × 10−2 | 20.7 | 1 |
| Indole 5 hydroxy | 1791 | 277.05 | 11.32 | 2.50 | 6.51 × 10−4 | 2.95 | 2.48 × 10−3 | 1.96 | 1.39 × 10−1 | 14.8 | 2 |
| Proline | 1297 | 142.06 | 7.34 | 2.09 | 7.96 × 10−3 | 2.49 | 1.04 × 10−2 | 1.79 | 2.15 × 10−1 | 14.8 | 1 |
| 4 hydroxyphenyl-acetic acid | 1643 | 163.98 | 10.23 | 2.08 | 6.74 × 10−4 | 1.66 | 4.33 × 10−2 | 2.89 | 1.36 × 10−2 | 16.6 | 2 |
| Alanine | 1137 | 116.03 | 5.86 | 1.88 | 2.92 × 10−3 | 1.82 | 2.63 × 10−2 | 1.96 | 8.15 × 10−2 | 31.1 | 1 |
| Myoinositol | 2077 | 318.09 | 13.26 | 1.88 | 2.49 × 10−2 | 1.86 | 1.73 × 10−1 | 1.93 | 2.24 × 10−2 | 17.3 | 1 |
| Threitol | 1488 | 128.01 | 9.03 | 1.60 | 2.45 × 10−2 | 1.85 | 2.84 × 10−2 | 1.40 | 2.84 × 10−1 | 14.7 | 2 |
| Valero 1,5-lactam | 1143 | 156.00 | 6.01 | 1.43 | 1.21 × 10−2 | 1.76 | 1.18 × 10−3 | 1.10 | 7.14 × 10−1 | 32.5 | 2 |
| Glyceric acid | 1318 | 292.06 | 7.54 | 1.35 | 2.27 × 10−1 | 1.77 | 2.07 × 10−2 | 1.07 | 8.71 × 10−1 | 40.2 | 2 |
| Benzoic acid | 1254 | 178.98 | 6.97 | 1.11 | 3.98 × 10−2 | 1.03 | 5.37 × 10−1 | 1.24 | 4.56 × 10−3 | 18.1 | 2 |
| alpha tocopherol | 3155 | 502.41 | 18.76 | 0.85 | 2.70 × 10−1 | 0.59 | 1.34 × 10−2 | 1.51 | 2.15 × 10−2 | 13.6 | 2 |
| 1-monoleoylglycerol | 2748 | 397.33 | 16.39 | 0.79 | 3.35 × 10−2 | 0.66 | 1.44 × 10−2 | 0.99 | 9.32 × 10−1 | 10.8 | 2 |
| Sitosterol | 3371 | 357.32 | 19.71 | 0.78 | 2.80 × 10−2 | 0.66 | 1.08 × 10−2 | 1.01 | 9.41 × 10−1 | 14.2 | 3 |
| Stigmastan-3-ol | 3386 | 215.11 | 19.77 | 0.77 | 2.59 × 10−2 | 0.63 | 3.49 × 10−3 | 1.08 | 5.77 × 10−1 | 14.3 | 3 |
| Heptdecanoic acid | 2141 | 327.24 | 13.68 | 0.76 | 1.67 × 10−2 | 0.64 | 1.45 × 10−3 | 0.94 | 6.35 × 10−1 | 23.8 | 3 |
| Tetradecanoic acid | 1845 | 285.20 | 11.75 | 0.74 | 1.30 × 10−2 | 0.64 | 1.05 × 10−2 | 0.90 | 3.97 × 10−1 | 35.9 | 2 |
| Allose | 1914 | 299.20 | 12.23 | 0.69 | 1.12 × 10−2 | 0.54 | 5.79 × 10−3 | 0.90 | 5.42 × 10−1 | 37.4 | 2 |
| Campesterol | 3291 | 343.28 | 19.35 | 0.46 | 3.96 × 10−4 | 0.43 | 8.48 × 10−3 | 0.50 | 3.42 × 10−2 | 15.1 | 3 |
| Deoxycholic acid | 3296 | 266.10 | 19.39 | 0.41 | 1.63 × 10−2 | 0.24 | 1.06 × 10−2 | 1.08 | 7.90 × 10−1 | 20.4 | 3 |
| Sebacic acid | 2424 | 215.12 | 15.22 | 0.33 | 6.33 × 10−4 | 0.19 | 1.37 × 10−3 | 0.60 | 2.53 × 10−1 | 8.6 | 3 |
| Metabolite ID | RI | Quant Ion (m/z) | Retention Time (min) | ko/wt | Females | Males | QC %RSD | ID Level | |||
|---|---|---|---|---|---|---|---|---|---|---|---|
| FC | p Value | FC | p Value | FC | p Value | ||||||
| Unknown | 2606 | 254.1 | 16.07 | 23.59 | 3.65 × 10−2 | 10.39 | 1.06 × 10−2 | 29.42 | 6.05 × 10−2 | 13.7 | 4 |
| p-cresol glucuronide | 2343 | 375.1 | 14.70 | 17.34 | 9.88 × 10−3 | 14.86 | 2.63 × 10−2 | 19.37 | 1.03 × 10−1 | 33.7 | 3 |
| Indole 3 propionic acid | 2095 | 202.1 | 13.28 | 4.41 | 2.19 × 10−2 | 4.46 | 1.74 × 10−2 | 4.39 | 1.01 × 10−1 | 12.2 | 2 |
| Phenaceturic acid | 1895 | 250.1 | 12.02 | 2.92 | 4.68 × 10−4 | 2.41 | 2.37 × 10−3 | 3.77 | 4.51 × 10−2 | 19.8 | 3 |
| Proline | 1298 | 142.1 | 7.28 | 2.50 | 9.79 × 10−3 | 2.14 | 4.71 × 10−2 | 2.68 | 8.88 × 10−3 | 12.5 | 1 |
| Xanthine | 2017 | 353.1 | 12.79 | 1.65 | 9.01 × 10−2 | 0.80 | 5.87 × 10−1 | 3.99 | 2.38 × 10−3 | 10.7 | 2 |
| Histidine | 1914 | 254.1 | 12.14 | 1.47 | 3.17 × 10−2 | 1.21 | 2.19 × 10−1 | 1.67 | 1.18 × 10−2 | 14.4 | 1 |
| Malate | 1474 | 233.1 | 8.83 | 1.47 | 4.16 × 10−2 | 1.33 | 3.54 × 10−1 | 1.57 | 2.51 × 10−2 | 11.6 | 1 |
| Glutamic acid | 1613 | 246.1 | 9.90 | 1.43 | 9.44 × 10−2 | 0.89 | 7.04 × 10−1 | 2.12 | 6.80 × 10−3 | 7.2 | 1 |
| Lysine | 1907 | 174.1 | 12.11 | 1.42 | 1.07 × 10−3 | 1.29 | 1.77 × 10−3 | 1.55 | 3.12 × 10−2 | 16.7 | 1 |
| Ornithine | 1809 | 142.1 | 11.37 | 1.38 | 6.81 × 10−2 | 1.34 | 7.76 × 10−3 | 1.41 | 1.54 × 10−1 | 14.8 | 1 |
| Glutamine | 1771 | 156.1 | 11.09 | 1.36 | 2.65 × 10−2 | 1.08 | 6.17 × 10−1 | 1.65 | 1.75 × 10−2 | 7.3 | 1 |
| Myoinositol | 2009 | 305.1 | 12.75 | 1.28 | 3.29 × 10−2 | 1.05 | 6.16 × 10−1 | 1.52 | 1.88 × 10−2 | 14.1 | 1 |
| Serine | 1350 | 218.1 | 7.74 | 1.25 | 6.11 × 10−2 | 1.11 | 4.09 × 10−1 | 1.37 | 4.94 × 10−2 | 13.5 | 1 |
| Fumaric acid | 1347 | 245.1 | 7.70 | 1.16 | 4.80 × 10−1 | 0.60 | 6.17 × 10−2 | 1.73 | 4.80 × 10−3 | 13.9 | 1 |
| Tyrosine | 1931 | 218.1 | 12.25 | 1.15 | 2.69 × 10−1 | 1.05 | 5.44 × 10−1 | 1.24 | 2.49 × 10−1 | 8.1 | 1 |
| Phenylalanine | 1632 | 192.1 | 10.04 | 1.14 | 4.24 × 10−1 | 1.18 | 1.68 × 10−1 | 1.11 | 5.73 × 10−1 | 10.9 | 1 |
| Stearic acid | 2242 | 341.3 | 14.16 | 0.95 | 4.59 × 10−1 | 0.81 | 2.63 × 10−2 | 1.08 | 3.09 × 10−1 | 7.2 | 1 |
| Octadecanol | 2143 | 327.2 | 13.58 | 0.92 | 3.47 × 10−1 | 0.77 | 1.20 × 10−2 | 1.04 | 6.70 × 10−1 | 18.1 | 2 |
| Leucine | 1265 | 158.2 | 6.99 | 0.90 | 4.80 × 10−1 | 1.09 | 6.24 × 10−1 | 0.79 | 9.38 × 10−2 | 14.1 | 1 |
| Valine | 1208 | 144.1 | 6.49 | 0.86 | 3.00 × 10−1 | 0.96 | 7.71 × 10−1 | 0.80 | 1.05 × 10−1 | 13.2 | 1 |
| Isoleucine | 1288 | 158.2 | 7.19 | 0.81 | 1.42 × 10−1 | 0.94 | 6.46 × 10−1 | 0.73 | 5.65 × 10−2 | 13.4 | 1 |
| Glucose | 1876 | 217.1 | 11.88 | 0.78 | 4.77 × 10−4 | 0.72 | 1.59 × 10−2 | 0.85 | 3.54 × 10−3 | 8.7 | 1 |
| Uric acid | 2088 | 441.2 | 13.24 | 0.77 | 8.27 × 10−2 | 0.94 | 7.73 × 10−1 | 0.67 | 1.02 × 10−2 | 8.7 | 1 |
| Threonine | 1376 | 291.1 | 7.96 | 0.73 | 1.65 × 10−2 | 0.69 | 5.60 × 10−2 | 0.76 | 8.27 × 10−2 | 16.5 | 1 |
| Glycerol | 1262 | 205.1 | 6.96 | 0.70 | 4.83 × 10−3 | 0.60 | 3.69 × 10−3 | 0.81 | 2.82 × 10−1 | 9.2 | 1 |
| Palmitic acid | 2046 | 313.2 | 12.97 | 0.69 | 7.69 × 10−3 | 0.55 | 2.10 × 10−2 | 0.84 | 3.14 × 10−1 | 11.1 | 1 |
| Linoleic acid | 2210 | 337.2 | 14.00 | 0.67 | 2.32 × 10−2 | 0.52 | 1.85 × 10−2 | 0.81 | 3.61 × 10−1 | 10.1 | 1 |
| Tryptophan | 2215 | 202.1 | 14.03 | 0.64 | 4.44 × 10−4 | 0.47 | 1.10 × 10−3 | 0.84 | 1.32 × 10−1 | 12.1 | 1 |
| Cysteine | 1550 | 220.1 | 9.42 | 0.63 | 1.8 × 10−2 | 0.71 | 1.10 × 10−1 | 0.58 | 4.76 × 10−2 | 24.1 | 1 |
| 11-cis-Octadecenoic acid | 2215 | 339.2 | 14.03 | 0.43 | 3.24 × 10−3 | 0.35 | 2.43 × 10−2 | 0.54 | 1.19 × 10−1 | 9.9 | 2 |
| Oleic acid | 2748 | 397.3 | 16.82 | 0.36 | 2.88 × 10−3 | 0.44 | 9.66 × 10−2 | 0.29 | 2.81 × 10−2 | 18.9 | 2 |
| cis-9-Hexadecenoic acid | 2026 | 311.2 | 12.85 | 0.19 | 1.10 × 10−2 | 0.16 | 2.56 × 10−2 | 0.22 | 2.05 × 10−1 | 10.2 | 2 |
| Urine | Faeces | Plasma | ||||||
|---|---|---|---|---|---|---|---|---|
| ID | AUC | p Value | ID | AUC | p Value | ID | AUC | p Value |
| Aminobutyric acid | 1 | 8.80 × 10−10 | Aminobutyric acid | 1 | 1.2 × 10−6 | Indole 3 Propionic acid | 0.99 | 1.32 × 10−2 |
| Valine | 1 | 5.50 × 10−9 | Valine | 1 | 1.50 × 10−5 | p−cresol glucuronide | 0.99 | 4.38 × 10−3 |
| Isoleucine | 1 | 3.00 × 10−11 | Leucine | 1 | 9.80 × 10−7 | Ornithine | 0.95 | 2.15 × 10−3 |
| Leucine | 1 | 5.60 × 10−10 | 5−aminovalerate | 1 | 2.50 × 10−7 | Tryptophan | 0.94 | 4.29 × 10−4 |
| Glycine | 1 | 1.40 × 10−9 | Phenylalanine | 0.992 | 2.40 × 10−6 | 11Octadecenoic acid | 0.93 | 1.22 × 10−3 |
| Serine | 1 | 4.40 × 10−9 | Tryptophan | 0.975 | 1.90 × 10−3 | Glucose | 0.92 | 4.28 × 10−4 |
| Threonine | 1 | 5.30 × 10−12 | Isoleucine | 0.975 | 6.40 × 10−5 | Phenaceturic acid | 0.91 | 1.67 × 10−4 |
| Methionine | 1 | 1.80 × 10−10 | Tyrosine | 0.975 | 0.001948 | Tetradecanoic acid | 0.91 | 5.41 × 10−3 |
| Phenylalanine | 1 | 1.40 × 10−8 | Dihydro × yphenylalanine | 0.975 | 1.37 × 10−6 | Lysine | 0.9 | 9.17 × 10−4 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Javed, K.; Cheng, Q.; Carroll, A.J.; Truong, T.T.; Bröer, S. Development of Biomarkers for Inhibition of SLC6A19 (B0AT1)—A Potential Target to Treat Metabolic Disorders. Int. J. Mol. Sci. 2018, 19, 3597. https://doi.org/10.3390/ijms19113597
Javed K, Cheng Q, Carroll AJ, Truong TT, Bröer S. Development of Biomarkers for Inhibition of SLC6A19 (B0AT1)—A Potential Target to Treat Metabolic Disorders. International Journal of Molecular Sciences. 2018; 19(11):3597. https://doi.org/10.3390/ijms19113597
Chicago/Turabian StyleJaved, Kiran, Qi Cheng, Adam J. Carroll, Thy T. Truong, and Stefan Bröer. 2018. "Development of Biomarkers for Inhibition of SLC6A19 (B0AT1)—A Potential Target to Treat Metabolic Disorders" International Journal of Molecular Sciences 19, no. 11: 3597. https://doi.org/10.3390/ijms19113597