Suppression of IFN-γ Production in Murine Splenocytes by Histamine Receptor Antagonists

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
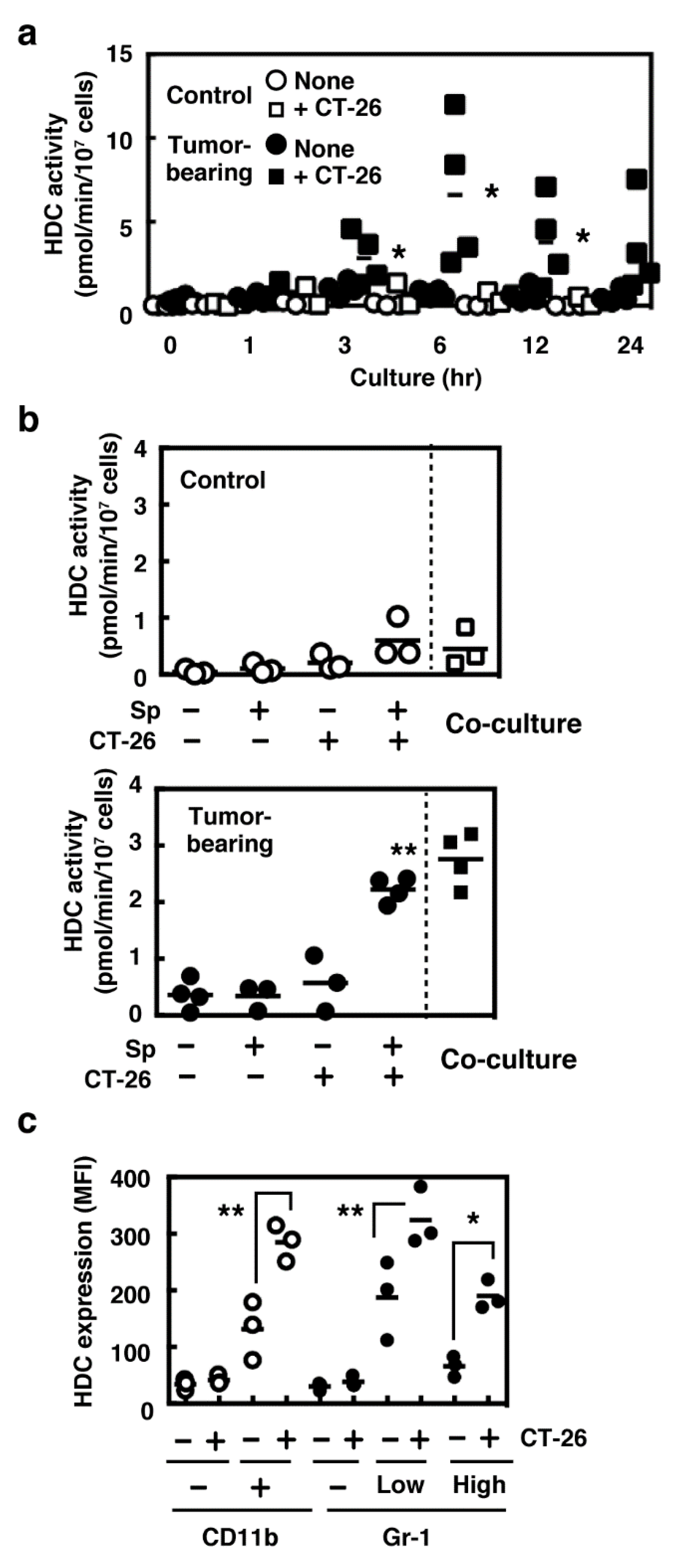
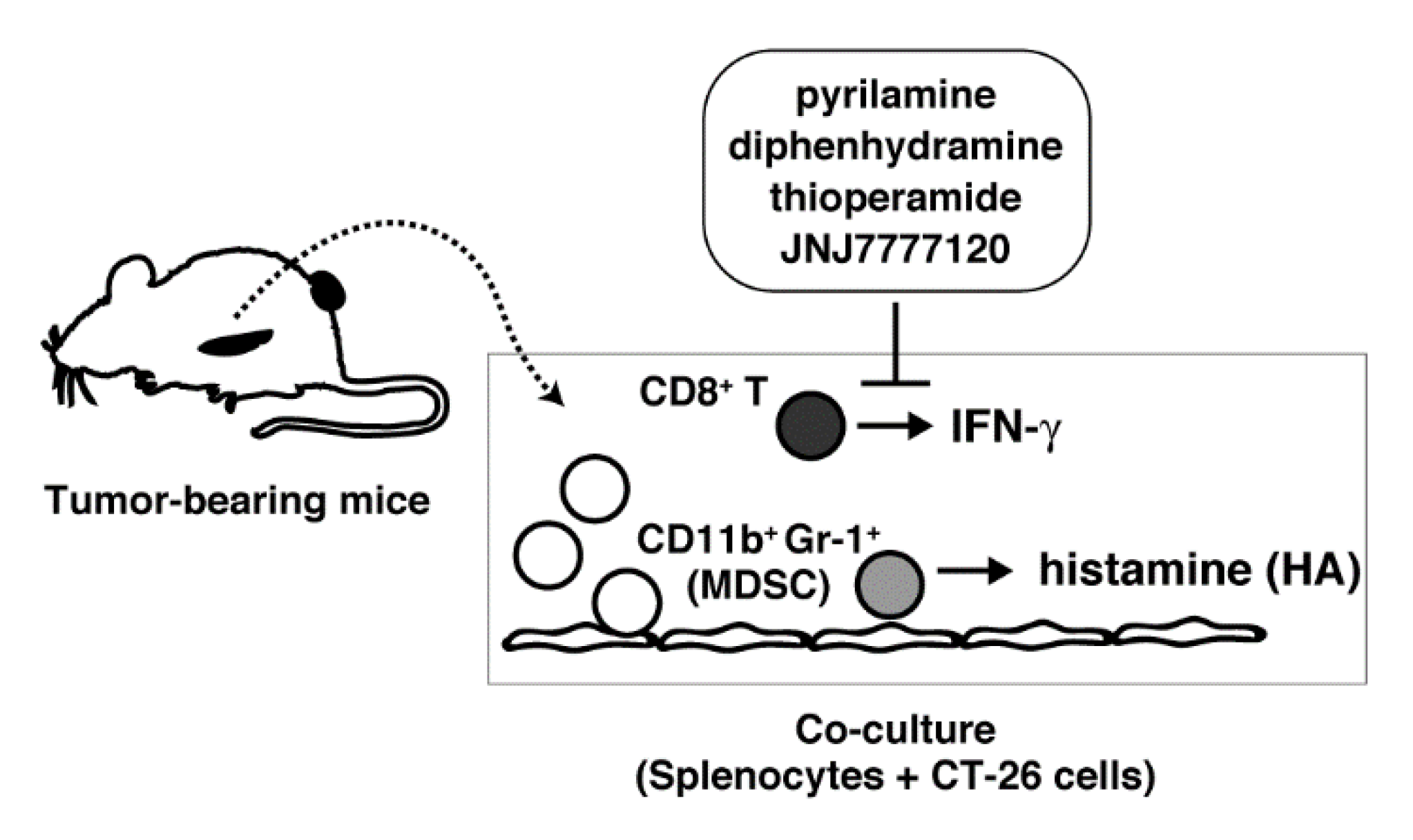
2.1. Induced Histamine Synthesis in Splenocytes Derived from Tumor-Bearing Mice
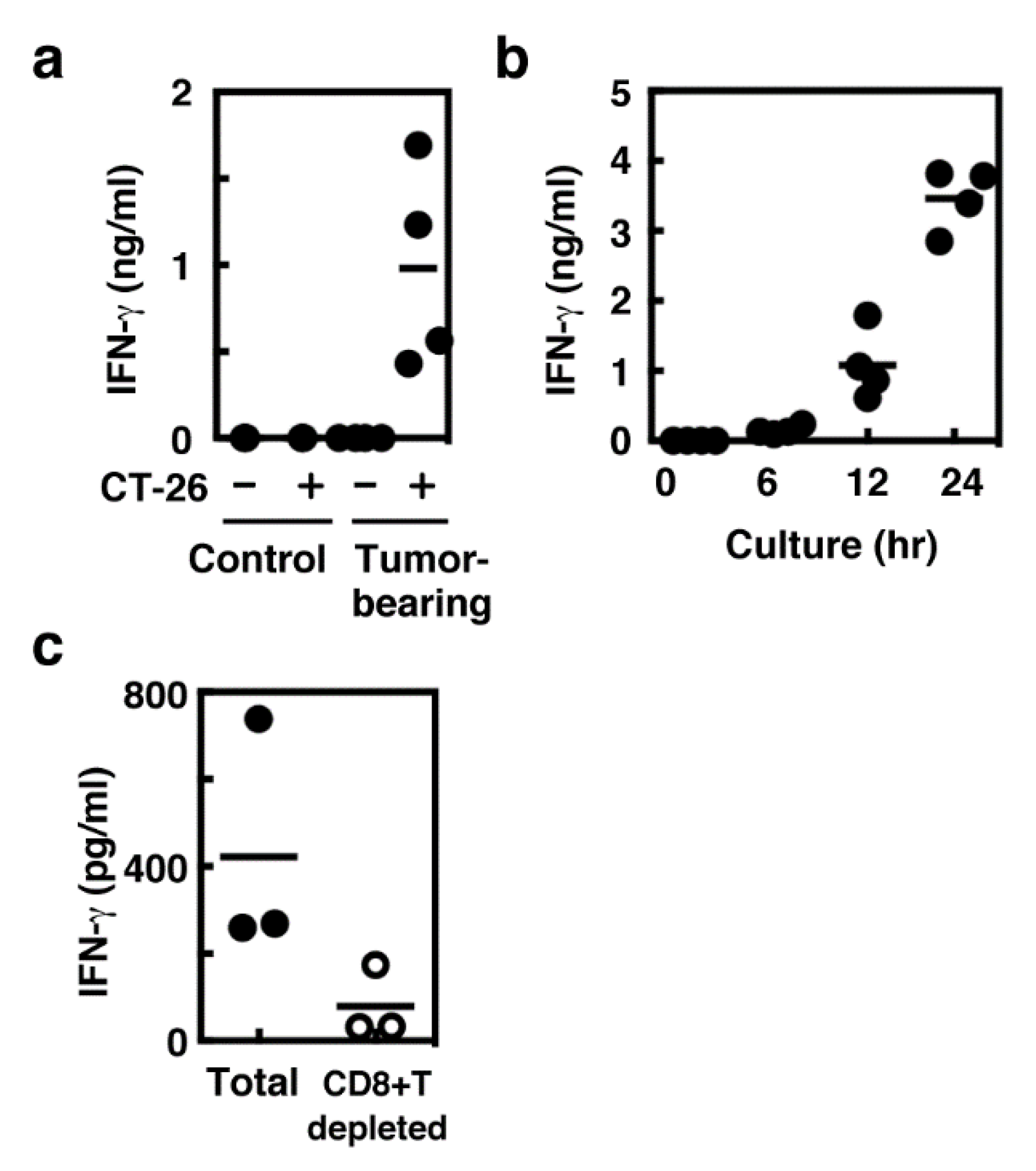
2.2. IFN-γ Production Mainly in Spleen CD8+ T Cells
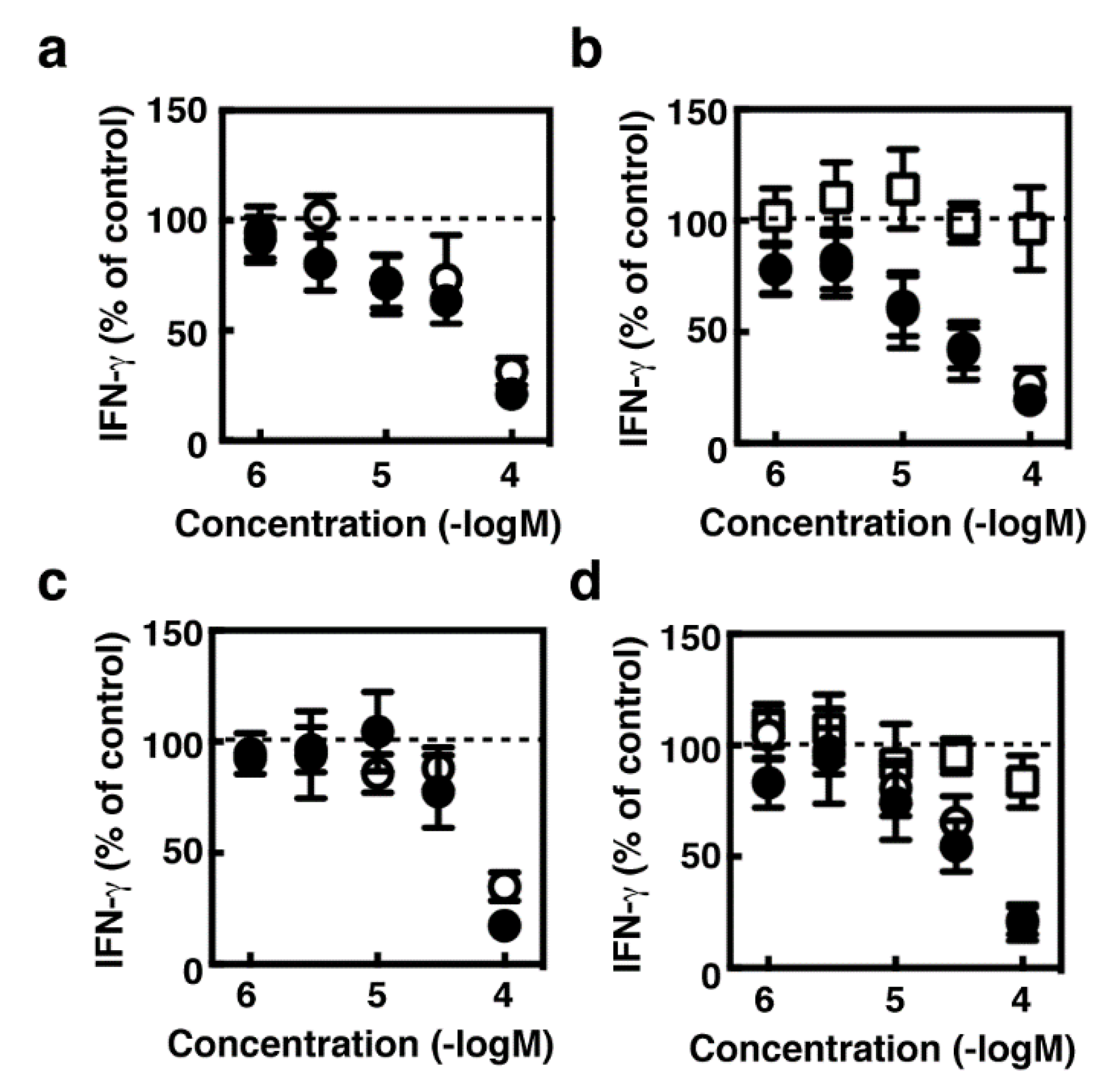
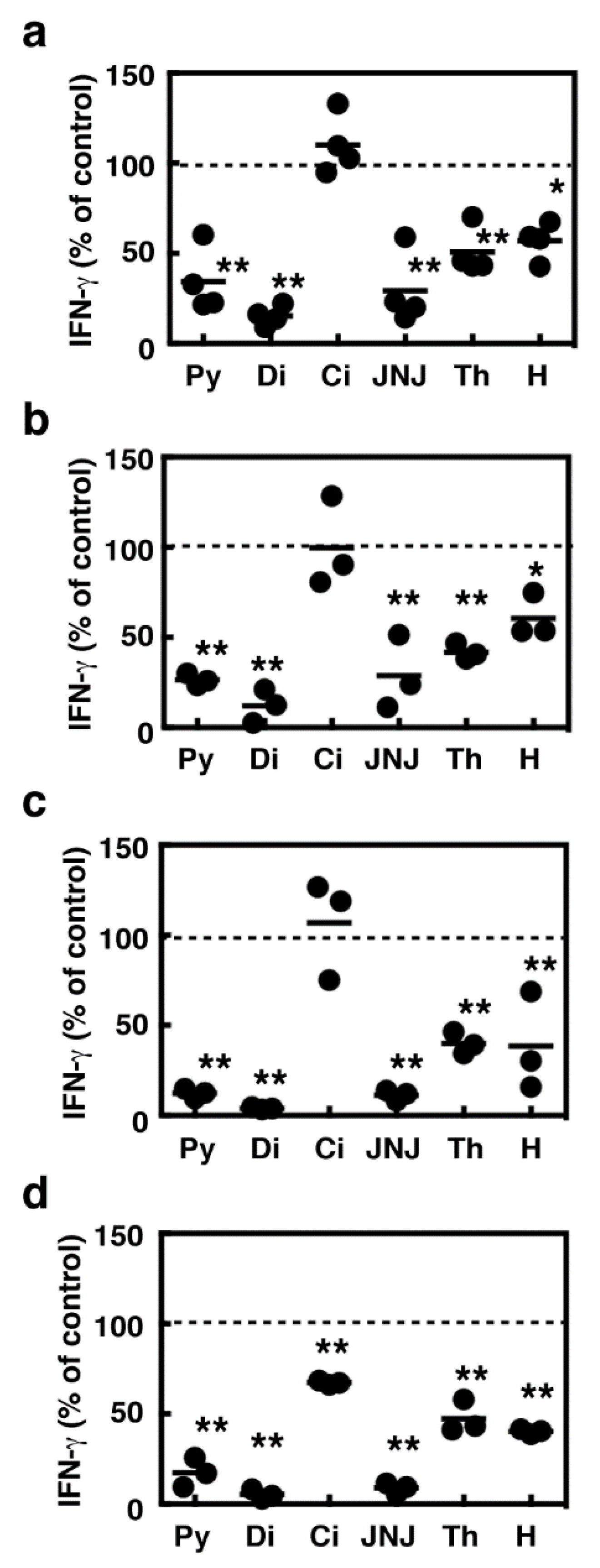
2.3. Suppression of IFN-γ Production During the Co-Culture Period by Several Histamine Receptor Antagonists
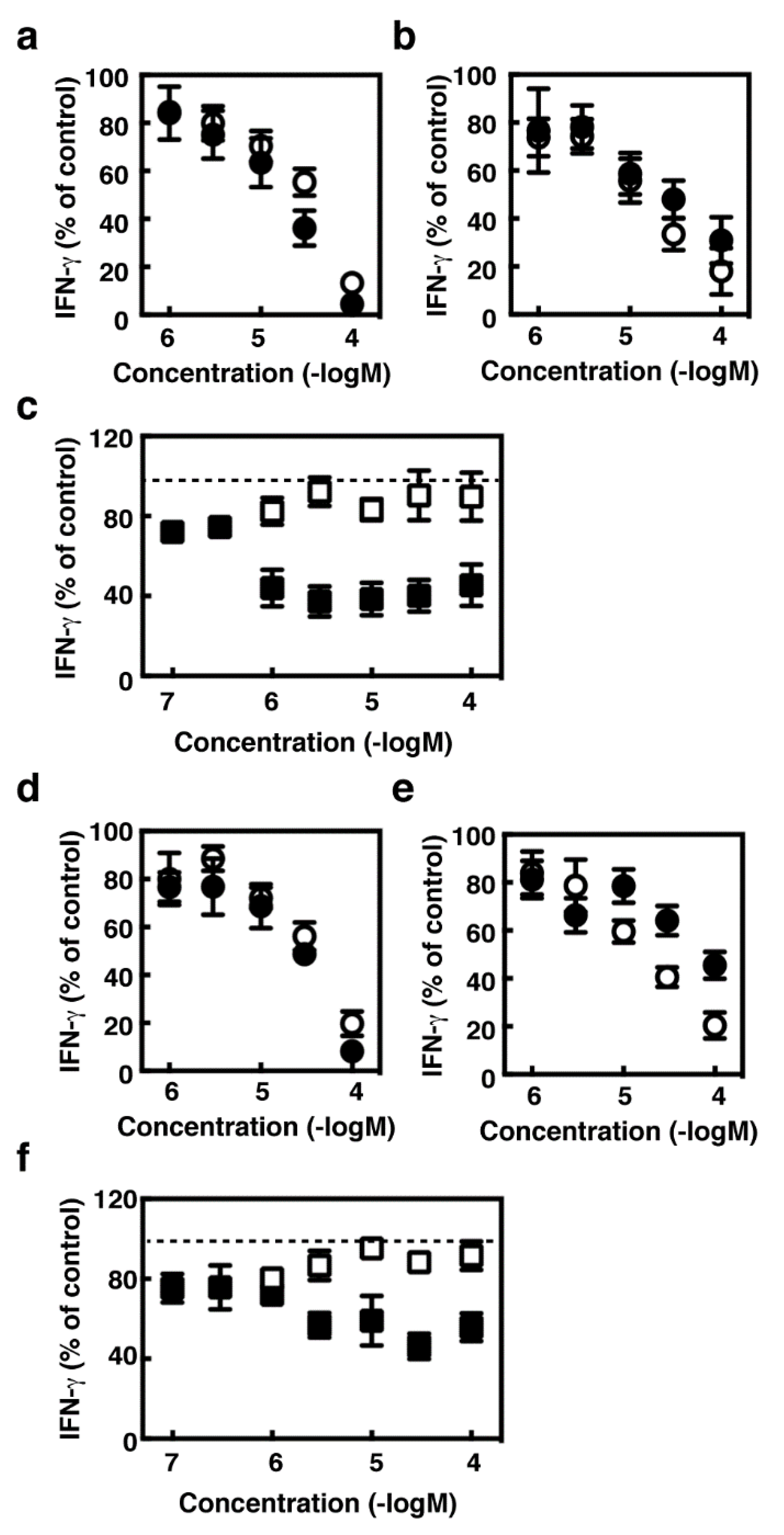
2.4. Effects of Histamine Receptor Ligands on IFN-γ Production in the Activated Splenocytes
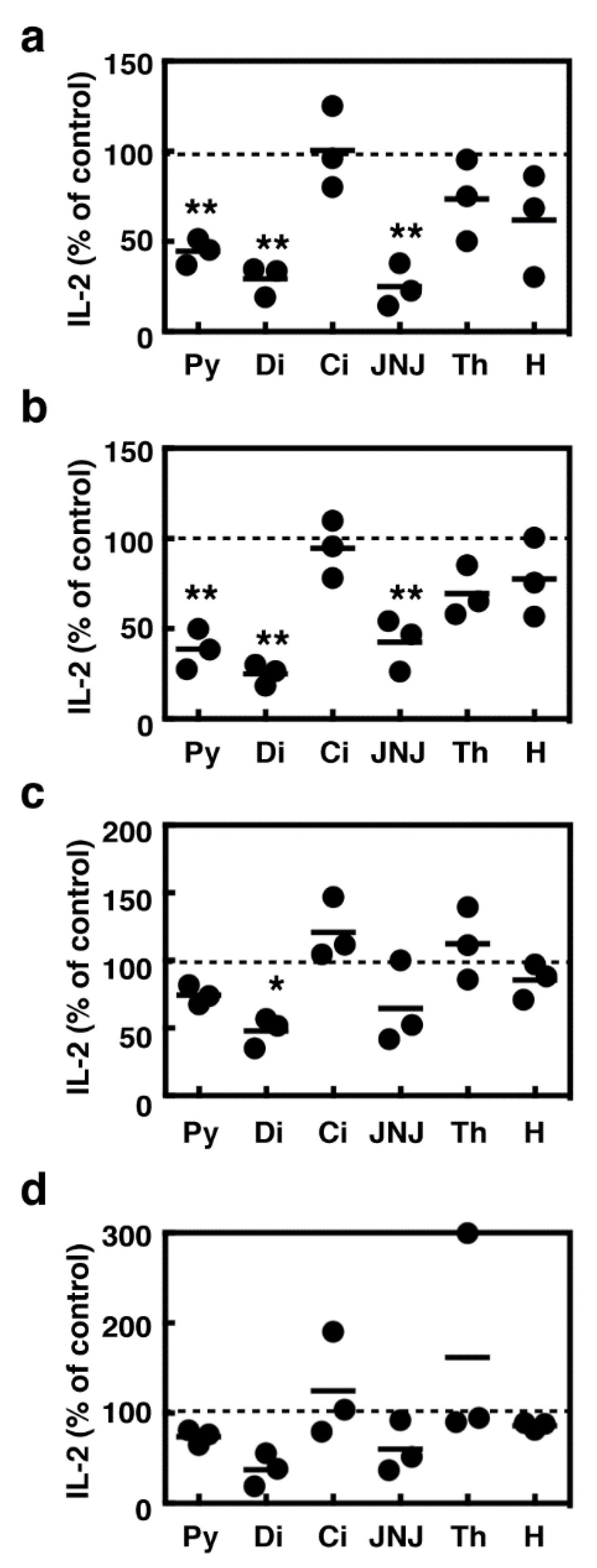
2.5. Effects of Histamine Receptor Ligands on IL-2 Production in the Activated Splenocytes
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Animals
4.3. Murine Syngeneic Tumor Model
4.4. Co-Culture of Splenocytes and CT-26 Cells
4.5. Measurement of Histamine
4.6. Flow Cytometry
4.7. Measurement of Cytokines
4.8. Depletion of CD8+ T cells
4.9. Stimulation of T Cell Receptors
4.10. Measurement of Histamine Receptor mRNAs
4.11. Statistics
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ANOVA | Analysis of variance |
| Con A | Concanavalin A |
| FBS | Fetal bovine serum |
| GM-CSF | Granulocyte–macrophage colony-stimulating factor |
| HDC | l-Histidine decarboxylase |
| IFN | Interferon |
| MDSC | Myeloid-derived suppressor cells |
| PBS | Phosphate buffered saline |
| PCR | Polymerase chain reaction |
| T-bet | T box-containing protein expressed in T cells |
References
- Panula, P.; Chazot, P.L.; Cowart, M.; Gutzmer, R.; Leurs, R.; Liu, W.L.; Stark, H.; Thurmond, R.L.; Haas, H.L. International Union of Basic and Clinical Pharmacology. XCVIII. Histamine Receptors. Pharmacol. Rev. 2015, 67, 601–655. [Google Scholar] [CrossRef] [PubMed]
- Taguchi, Y.; Tsuyama, K.; Watanabe, T.; Wada, H.; Kitamura, Y. Increase in histidine decarboxylase activity in skin of genetically mast-cell-deficient W/Wv mice after application of phorbol 12-myristate 13-acetate: Evidence for the presence of histamine-producing cells without basophilic granules. Proc. Natl. Acad. Sci. USA 1982, 79, 6837–6841. [Google Scholar] [CrossRef]
- Schneider, E.; Ploemacher, R.E.; Nabarra, B.; Brons, N.H.; Dy, M. Mast cells and their committed precursors are not required for interleukin-3-induced histamine synthesis in murine bone marrow: Characteristics of histamine-producing cells. Blood 1993, 81, 1161–1169. [Google Scholar]
- Shiraishi, M.; Hirasawa, N.; Oikawa, S.; Kobayashi, Y.; Ohuchi, K. Analysis of histamine-producing cells at the late phase of allergic inflammation in rats. Immunology 2000, 99, 600–606. [Google Scholar] [CrossRef]
- Kawaguchi-Nagata, K.; Watanabe, T.; Maeyama, K.; Yamatodani, A.; Okamura, H.; Tamura, T.; Shoji, K.; Kitamura, Y. Increase of histidine decarboxylase activity in murine myelomonocytic leukemia cells (WEHI-3B) in parallel to their differentiation into macrophages. Biochim. Biophys. Acta. 1988, 972, 249–256. [Google Scholar] [PubMed]
- Tanaka, S.; Deai, K.; Konomi, A.; Takahashi, K.; Yamane, H.; Sugimoto, Y.; Ichikawa, A. Expression of L-histidine decarboxylase in granules of elicited mouse polymorphonuclear leukocytes. Eur. J. Immunol. 2004, 34, 1472–1482. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Zhang, D.; Zhang, H.; Wolters, P.J.; Killeen, N.P.; Sullivan, B.M.; Locksley, R.M.; Lowell, C.A.; Caughey, G.H. Neutrophil histamine contributes to inflammation in mycoplasma pneumonia. J. Exp. Med. 2006, 203, 2907–2917. [Google Scholar] [CrossRef] [PubMed]
- Jutel, M.; Watanabe, T.; Klunker, S.; Akdis, M.; Thomet, O.A.; Malolepszy, J.; Zak-Nejmark, T.; Kog, R.; Kobayashi, T.; Blaser, K.; et al. Histamine regulated T-cell and antibody responses by differential expression of H1 and H2 receptors. Nature 2001, 413, 420–425. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Tanaka, S.; Ichikawa, A. Effect of cimetidine on intratumoral cytokine expression in an experimental tumor. Biochem. Biophys. Res. Commun. 2001, 281, 1113–1119. [Google Scholar] [CrossRef]
- Takahashi, K.; Tanaka, S.; Furuta, K.; Ichikawa, A. Histamine H2 receptor-mediated modulation of local cytokine expression in a mouse experimental tumor model. Biochem. Biophys. Res. Commun. 2002, 297, 1205–1210. [Google Scholar] [CrossRef]
- Chen, X.; Takemoto, Y.; Deng, H.; Middelhoff, M.; Friedman, R.A.; Chu, T.H.; Churchill, M.J.; Ma, Y.; Nagar, K.K.; Tailor, Y.H.; et al. Histidine decarboxylase (HDC)-expressing granulocytic myeloid cells induce and recruit Foxp3+ regulatory T cells in murine colon cancer. Oncoimmunology 2017, 6, e1290034. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.D.; Ai, W.; Asfaha, S.; Bhagat, G.; Friedman, R.A.; Jin, G.; Park, H.; Shykind, B.; Diacovo, T.G.; Falus, A.; Wang, T.C. Histamine deficiency promotes inflammation-associated carcinogenesis through reduced myeloid maturation and accumulation of CD11b+Ly6G+ immature myeloid cells. Nat. Med. 2011, 17, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Ahn, B.; Kohanbash, G.; Ohkuri, T.; Kosaka, A.; Chen, X.; Ikeura, M.; Wang, T.C.; Okada, H. Histamine deficiency promotes accumulation of immunosuppressive immature myeloid cells and growth of murine gliomas. Oncoimmunology 2015, 4, e1047581. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, S.; Ichikawa, A. Recent Advances in Molecular Pharmacology of the Histamine Systems: Immune Regulatory Roles of Histamine Produced by Leukocytes. J. Pharmacol. Sci. 2006, 101, 19–23. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.Z.; Gao, J.; Meeker, N.D.; Fillmore, P.D.; Tung, K.S.; Watanabe, T.; Zachary, J.F.; Offner, H.; Blankenhorn, E.P.; Teuscher, C. Identification of Bphs, an autoimmune disease locus, as histamine receptor H1. Science 2002, 297, 620–623. [Google Scholar] [CrossRef]
- Inoue, I.; Yanai, K.; Kitamura, D.; Taniuchi, I.; Kobayashi, T.; Niimura, K.; Watanabe, T.; Watanabe, T. Impaired locomotor activity and exploratory behavior in mice lacking histamine H1 receptors. Proc. Natl. Acad. Sci. USA 1996, 93, 13316–13320. [Google Scholar] [CrossRef] [PubMed]
- Endo, Y. Induction of histidine and ornithine decarboxylase activities in mouse tissues by recombinant interleukin-1 and tumor necrosis factor. Biochem. Pharmacol. 1989, 38, 1287–1292. [Google Scholar] [CrossRef]
- Schneider, E.; Pollard, H.; Lepault, F.; Guy-Grand, D.; Minkowski, M.; Dy, M. Histamine-producing cell-stimulating activity. Interleukin 3 and granulocyte-macrophage colony-stimulating factor induce de novo synthesis of histidine decarboxylase in hemopoietic progenitor cells. J. Immunol. 1987, 139, 3710–3717. [Google Scholar]
- Oda, T.; Morikawa, N.; Saito, Y.; Masuho, Y.; Matsumoto, S. Molecular cloning and characterization of a novel type of histamine receptor preferentially expressed in leukocytes. J. Biol. Chem. 2000, 275, 36781–36786. [Google Scholar] [CrossRef]
- Chen, M.L.; Tsai, T.C.; Wang, L.K.; Lin, Y.Y.; Tsai, Y.M.; Lee, M.C.; Tsai, F.M. Clozapine inhibits Th1 cell differentiation and causes the suppression of IFN-γ production in peripheral blood mononuclear cells. Immunopharmacol. Immunotoxicol. 2012, 34, 686–694. [Google Scholar] [CrossRef]
- Oh, Y.J.; Kim, D.; Oh, S.; Jang, E.J.; Won, H.Y.; Jeong, H.; Jeong, M.G.; Choo, H.P.; Hwang, E.S. Novel benzoxazole derivatives DCPAB and HPAB attenuate Th1 cell-mediated inflammation through T-bet suppression. Sci. Rep. 2017, 7, 42144. [Google Scholar] [CrossRef]
- Szabo, S.J.; Sullivan, B.M.; Stemmann, C.; Satoskar, A.R.; Sleckman, B.P.; Glimcher, L.H. Distinct effects of T-bet in Th1 lineage commitment and IFN-γ production in CD4 and CD8 T cells. Science 2002, 295, 338–342. [Google Scholar] [CrossRef] [PubMed]
- Pearce, E.L.; Mullen, A.C.; Martins, G.A.; Krawczyk, C.M.; Hutchins, A.S.; Zediak, V.P.; Banica, M.; DiCioccio, C.B.; Gross, D.A.; Mao, C.A.; et al. Control of effector CD8+ T cell function by the transcription factor Eomesodermin. Science 2003, 302, 1041–1043. [Google Scholar] [CrossRef] [PubMed]
- Way, S.S.; Wilson, C.B. Cutting edge: Immunity and IFN-γ production during Listeria monocytogenes infection in the absence of T-bet. J. Immunol. 2004, 173, 5918–5922. [Google Scholar] [CrossRef]
- Bakker, R.A.; Wieland, K.; Timmerman, H.; Leurs, R. Constitutive activity of the histamine H1 receptor reveals inverse agonism of histamine H1 receptor antagonists. Eur. J. Pharmacol. 2000, 387, R5–R7. [Google Scholar] [CrossRef]
- Fitzsimons, C.P.; Monczor, F.; Fernández, N.; Shayo, C.; Davio, C. Mepyramine, a histamine H1 receptor inverse agonist, binds preferentially to a G protein-coupled form of the receptor and sequesters G protein. J. Biol. Chem. 2004, 279, 34431–34439. [Google Scholar] [CrossRef] [PubMed]
- Mizuguchi, H.; Ono, S.; Hattori, M.; Fukui, H. Inverse agonistic activity of antihistamines and suppression of histamine H1 receptor gene expression. J. Pharmacol. Sci. 2012, 118, 117–121. [Google Scholar] [CrossRef]
- Monczor, F.; Fernandez, N.; Fitzsimons, C.P.; Shayo, C.; Davio, C. Antihistaminergics and inverse agonism: Potential therapeutic applications. Eur. J. Pharmacol. 2013, 715, 26–32. [Google Scholar] [CrossRef]
- Liu, C.; Wilson, S.J.; Kuei, C.; Lovenberg, T.W. Comparison of human, mouse, rat, and guinea pig histamine H4 receptors reveals substantial pharmacological species variation. J. Pharmacol. Exp. Ther. 2001, 299, 121–130. [Google Scholar]
- Green, D.S.; Young, H.A.; Valencia, J.C. Current prospects of type II interferon g signaling and autoimmunity. J. Biol. Chem. 2017, 292, 13925–13933. [Google Scholar] [CrossRef]
- Giuffrida, P.; Corazza, G.R.; Di Sabatino, A. Old and New Lymphocyte Players in Inflammatory Bowel Disease. Dig. Dis. Sci. 2018, 63, 277–288. [Google Scholar] [CrossRef] [PubMed]
- Ohtsu, H.; Tanaka, S.; Terui, T.; Hori, Y.; Makabe-Kobayashi, Y.; Pejler, G.; Tchougounova, E.; Hellman, L.; Gertsenstein, M.; Hirasawa, N.; et al. Mice lacking histidine decarboxylase exhibit abnormal mast cells. FEBS Lett. 2001, 502, 53–56. [Google Scholar] [CrossRef]
- Yamatodani, A.; Fukuda, H.; Wada, H.; Iwaeda, T.; Watanabe, T. High-performance liquid chromatographic determination of plasma and brain histamine without previous purification of biological samples: Cation-exchange chromatography coupled with post-column derivatization fluorometry. J. Chromatogr. 1985, 344, 115–123. [Google Scholar] [CrossRef]







© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kamei, M.; Otani, Y.; Hayashi, H.; Nakamura, T.; Yanai, K.; Furuta, K.; Tanaka, S. Suppression of IFN-γ Production in Murine Splenocytes by Histamine Receptor Antagonists. Int. J. Mol. Sci. 2018, 19, 4083. https://doi.org/10.3390/ijms19124083
Kamei M, Otani Y, Hayashi H, Nakamura T, Yanai K, Furuta K, Tanaka S. Suppression of IFN-γ Production in Murine Splenocytes by Histamine Receptor Antagonists. International Journal of Molecular Sciences. 2018; 19(12):4083. https://doi.org/10.3390/ijms19124083
Chicago/Turabian StyleKamei, Miho, Yukie Otani, Hidenori Hayashi, Tadaho Nakamura, Kazuhiko Yanai, Kazuyuki Furuta, and Satoshi Tanaka. 2018. "Suppression of IFN-γ Production in Murine Splenocytes by Histamine Receptor Antagonists" International Journal of Molecular Sciences 19, no. 12: 4083. https://doi.org/10.3390/ijms19124083
APA StyleKamei, M., Otani, Y., Hayashi, H., Nakamura, T., Yanai, K., Furuta, K., & Tanaka, S. (2018). Suppression of IFN-γ Production in Murine Splenocytes by Histamine Receptor Antagonists. International Journal of Molecular Sciences, 19(12), 4083. https://doi.org/10.3390/ijms19124083