Effects of Nitric Oxide on Voltage-Gated K+ Currents in Human Cardiac Fibroblasts through the Protein Kinase G and Protein Kinase A Pathways but Not through S-Nitrosylation


Abstract
:
1. Introduction
2. Results
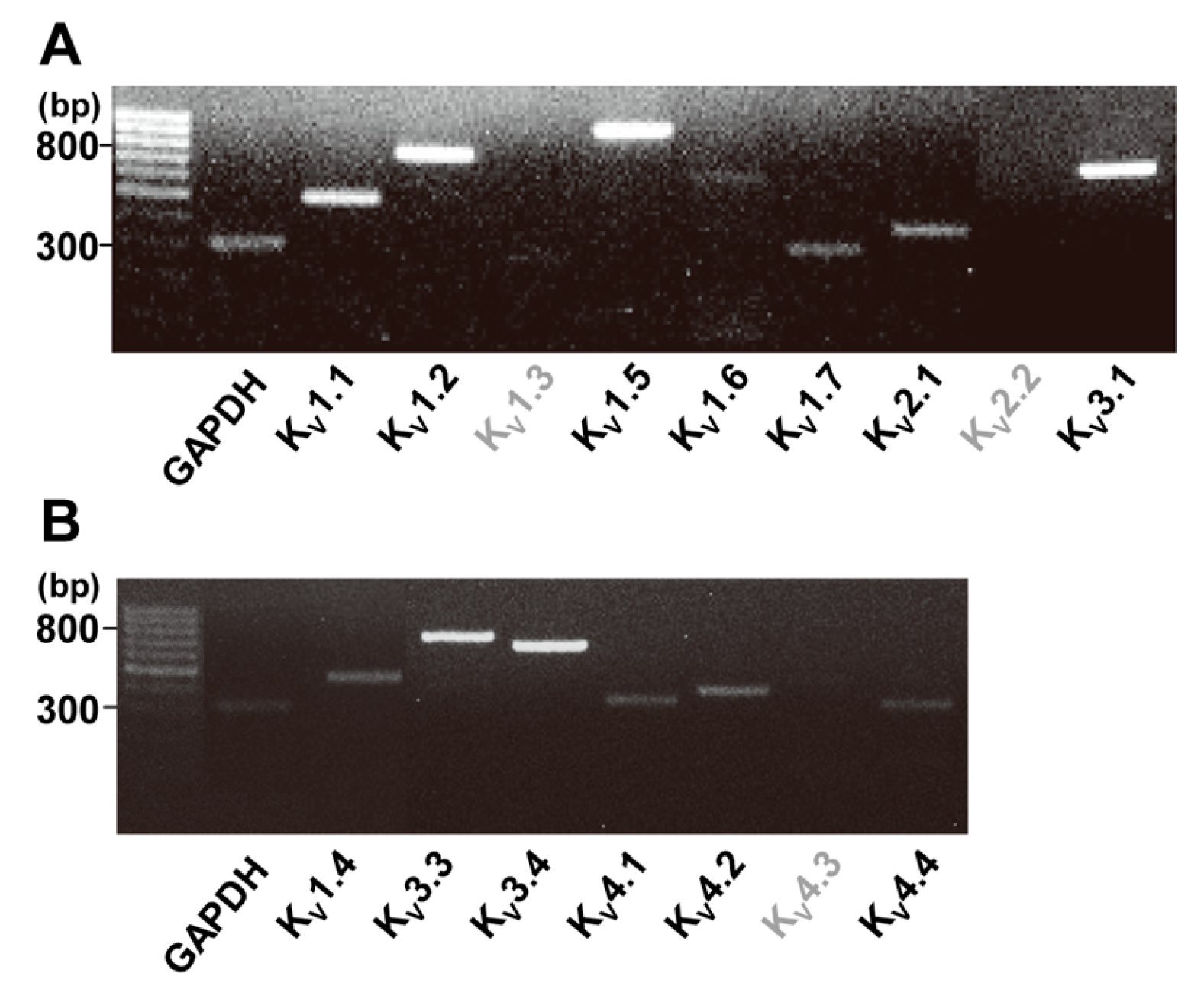
2.1. Identification of Two Types of Voltage-Gated K+ Channels in Human Cardiac Fibroblasts by RT-PCR
2.2. Identification of Two Types of Voltage-Gated K+ Channels using Electrophysiological Methods
2.3. Effect of NO on Two Types of Voltage-Gated K+ Currents
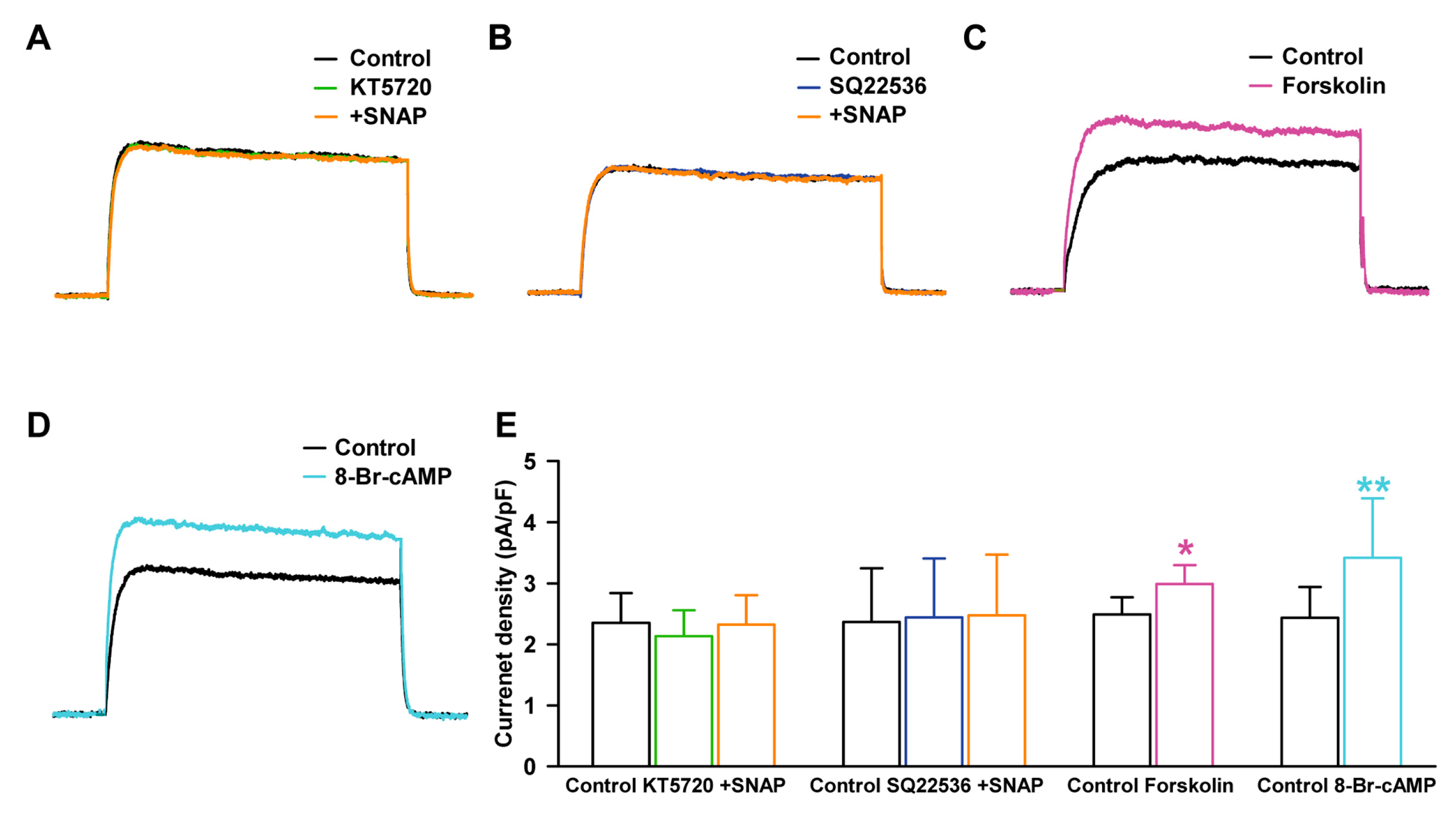
2.4. Effect of NO on Delayed Rectifier K+ Current through PKG Signaling Pathway
2.5. Effect of NO on Delayed Rectifier K+ Current through PKA Signaling Pathway
2.6. Effect of NO on Delayed Rectifier K+ Current through the S-Nitrosylation Pathway
3. Discussion
3.1. Identification of Two Types of Kv Channels in Human Cardiac Fibroblasts by Molecular Methods
3.2. Identification of Voltage-Gated K+ Channels in Human Cardiac Fibroblasts via Electrophysiological Methods
3.3. Effect of NO on Two Types of Voltage-Gated K+ Currents
3.4. Effect of NO on Delayed Rectifier K+ Current through Protein Kinases Signaling Pathways and S-Nitrosylation
4. Materials and Methods
4.1. Cell Preparation and Culture
4.2. Reverse Transcription Polymerase Chain Reaction (RT-PCR)
4.3. Electrophysiological Recordings
4.4. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Camelliti, P.; Borg, T.K.; Kohl, P. Structural and functional characterisation of cardiac fibroblasts. Cardiovasc. Res. 2005, 65, 40–51. [Google Scholar] [CrossRef] [PubMed]
- Kohl, P. Heterogeneous cell coupling in the heart: An electrophysiological role for fibroblasts. Circ. Res. 2003, 93, 381–383. [Google Scholar] [CrossRef] [PubMed]
- Li, G.R.; Sun, H.Y.; Chen, J.B.; Zhou, Y.; Tse, H.F.; Lau, C.P. Characterization of multiple ion channels in cultured human cardiac fibroblasts. PLoS ONE 2009, 4, e7307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaudesius, G.; Miragoli, M.; Thomas, S.P.; Rohr, S. Coupling of cardiac electrical activity over extended distances by fibroblasts of cardiac origin. Circ. Res. 2003, 93, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Kohl, P.; Camelliti, P.; Burton, F.L.; Smith, G.L. Electrical coupling of fibroblasts and myocytes: Relevance for cardiac propagation. J. Electrocardiol. 2005, 38, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Quinn, T.A.; Camelliti, P.; Rog-Zielinska, E.A.; Siedlecka, U.; Poggioli, T.; O’Toole, E.T.; Knopfel, T.; Kohl, P. Electrotonic coupling of excitable and nonexcitable cells in the heart revealed by optogenetics. Proc. Natl. Acad. Sci. USA 2016, 113, 14852–14857. [Google Scholar] [CrossRef] [PubMed]
- Pellman, J.; Zhang, J.; Sheikh, F. Myocyte-fibroblast communication in cardiac fibrosis and arrhythmias: Mechanisms and model systems. J. Mol. Cell. Cardiol. 2016, 94, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Burstein, B.; Oi, X.Y.; Yeh, Y.H.; Calderone, A.; Nattel, S. Atrial cardiomyocyte tachycardia alters cardiac fibroblast function: A novel consideration in atrial remodeling. Circulation 2007, 116, 442–452. [Google Scholar] [CrossRef] [PubMed]
- Pedrotty, D.M.; Klinger, R.Y.; Kirkton, R.D.; Bursac, N. Cardiac fibroblast paracrine factors alter impulse conduction and ion channel expression of neonatal rat cardiomyocytes. Cardiovasc. Res. 2009, 83, 688–697. [Google Scholar] [CrossRef] [PubMed]
- Vasquez, C.; Mohandas, P.; Louie, K.L.; Benamer, N.; Bapat, A.C.; Morley, G.E. Enhanced fibroblast-myocyte interactions in response to cardiac injury. Circ. Res. 2010, 107, 1011–1120. [Google Scholar] [CrossRef] [PubMed]
- Chilton, L.; Ohya, S.; Freed, D.; George, E.; Drobic, V.; Shibukawa, Y.; Maccannell, K.A.; Imaizumi, Y.; Clark, R.B.; Dixon, I.M.; et al. K+ currents regulate the resting membrane potential, proliferation, and contractile responses in ventricular fibroblasts and myofibroblasts. Am. J. Physiol. Heart Circ. Physiol. 2005, 288, H2931–H2939. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.J.; Sung, R.J.; Lin, M.W.; Wu, S.N. Contribution of bk(ca)-channel activity in human cardiac fibroblasts to electrical coupling of cardiomyocytes-fibroblasts. J. Membr. Biol. 2006, 213, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Bae, H.; Lim, I. Effects of nitric oxide on large-conductance Ca2+ -activated K+ currents in human cardiac fibroblasts through pka and pkg-related pathways. Clin. Exp. Pharmacol. Physiol. 2017, 44, 1116–1124. [Google Scholar] [CrossRef] [PubMed]
- Nerbonne, J.M. Molecular basis of functional voltage-gated K+ channel diversity in the mammalian myocardium. J. Physiol. (Lond.) 2000, 525, 285–298. [Google Scholar] [CrossRef]
- Nerbonne, J.M. Molecular basis of functional myocardial potassium channel diversity. Card. Electrophysiol. Clin. 2016, 8, 257–273. [Google Scholar] [CrossRef] [PubMed]
- Seddon, M.; Shah, A.M.; Casadei, B. Cardiomyocytes as effectors of nitric oxide signalling. Cardiovasc. Res. 2007, 75, 315–326. [Google Scholar] [CrossRef] [PubMed]
- Ziolo, M.T.; Kohr, M.J.; Wang, H. Nitric oxide signaling and the regulation of myocardial function. J. Mol. Cell. Cardiol. 2008, 45, 625–632. [Google Scholar] [CrossRef] [PubMed]
- Massion, P.B.; Pelat, M.; Belge, C.; Balligand, J.L. Regulation of the mammalian heart function by nitric oxide. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2005, 142, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Kanno, S.; Kim, P.K.; Sallam, K.; Lei, J.; Billiar, T.R.; Shears, L.L., 2nd. Nitric oxide facilitates cardiomyogenesis in mouse embryonic stem cells. Proc. Natl. Acad. Sci. USA 2004, 101, 12277–12281. [Google Scholar] [CrossRef] [PubMed]
- Takimoto, E.; Champion, H.C.; Li, M.; Ren, S.; Rodriguez, E.R.; Tavazzi, B.; Lazzarino, G.; Paolocci, N.; Gabrielson, K.L.; Wang, Y.; et al. Oxidant stress from nitric oxide synthase–3 uncoupling stimulates cardiac pathologic remodeling from chronic pressure load. J. Clin. Investig. 2005, 115, 1221–1231. [Google Scholar] [CrossRef] [PubMed]
- Andreka, P.N.Z.; Muzes, G.; Szantho, G.; Vandor, L.; Konya, L.; Turner, M.S.; Tulassay, Z.; Bishopric, N.H. Possible therapeutic targets in cardiac myocyte apoptosis. Curr. Pharm. Des. 2004, 10, 2445–2461. [Google Scholar] [CrossRef] [PubMed]
- Tamargo, J.; Caballero, R.; Gomez, R.; Delpon, E. Cardiac electrophysiological effects of nitric oxide. Cardiovasc. Res. 2010, 87, 593–600. [Google Scholar] [CrossRef] [PubMed]
- Nunez, L.; Vaquero, M.; Gomez, R.; Caballero, R.; Mateos-Caceres, P.; Macaya, C.; Iriepa, I.; Galvez, E.; Lopez-Farre, A.; Tamargo, J.; et al. Nitric oxide blocks hKv1.5 channels by s-nitrosylation and by a cyclic gmp-dependent mechanism. Cardiovasc. Res. 2006, 72, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Bai, C.X.; Takahashi, K.; Masumiya, H.; Sawanobori, T.; Furukawa, T. Nitric oxide-dependent modulation of the delayed rectifier k+ current and the l-type ca2+ current by ginsenoside re, an ingredient of panax ginseng, in guinea-pig cardiomyocytes. Br. J. Pharmacol. 2004, 142, 567–575. [Google Scholar] [CrossRef] [PubMed]
- Gomez, R.; Nunez, L.; Vaquero, M.; Amoros, I.; Barana, A.; de Prada, T.; Macaya, C.; Maroto, L.; Rodriguez, E.; Caballero, R.; et al. Nitric oxide inhibits kv4.3 and human cardiac transient outward potassium current (ito1). Cardiovasc. Res. 2008, 80, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Hammond, J.; Balligand, J.L. Nitric oxide synthase and cyclic gmp signaling in cardiac myocytes: From contractility to remodeling. J. Mol. Cell. Cardiol. 2012, 52, 330–340. [Google Scholar] [CrossRef] [PubMed]
- Vila-Petroff, M.G.; Younes, A.; Egan, J.; Lakatta, E.G.; Sollott, S.J. Activation of distinct camp-dependent and cgmp-dependent pathways by nitric oxide in cardiac myocytes. Circ. Res. 1999, 84, 1020–1031. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, D.R.; Treuer, A.; Sun, Q.A.; Stamler, J.S.; Hare, J.M. S-nitrosylation of cardiac ion channels. J. Cardiovasc. Pharmacol. 2009, 54, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Lima, B.; Forrester, M.T.; Hess, D.T.; Stamler, J.S. S-nitrosylation in cardiovascular signaling. Circ. Res. 2010, 106, 633–646. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Ruiz, A.; Lamas, S. S-nitrosylation: A potential new paradigm in signal transduction. Cardiovasc. Res. 2004, 62, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Ordog, B.; Brutyo, E.; Puskas, L.G.; Papp, J.G.; Varro, A.; Szabad, J.; Boldogkoi, Z. Gene expression profiling of human cardiac potassium and sodium channels. Int. J. Cardiol. 2006, 111, 386–393. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.L.; Wible, B.; Li, G.R.; Wang, Z.G.; Nattel, S. Antisense oligodeoxynucleotides directed against kv1.5 mRNA specifically inhibit ultrarapid delayed rectifier k+ current in cultured adult human atrial myocytes. Circ. Res. 1997, 80, 572–579. [Google Scholar] [CrossRef] [PubMed]
- Brahmajothi, M.V.; Campbell, D.L.; Rasmusson, R.L.; Morales, M.J.; Trimmer, J.S.; Nerbonne, J.M.; Strauss, H.C. Distinct transient outward potassium current (ito) phenotypes and distribution of fast-inactivating potassium channel alpha subunits in ferret left ventricular myocytes. J. Gen. Physiol. 1999, 113, 581–600. [Google Scholar] [CrossRef] [PubMed]
- Isbrandt, D.; Leicher, T.; Waldschutz, R.; Zhu, X.; Luhmann, U.; Michel, U.; Sauter, K.; Pongs, O. Gene structures and expression profiles of three human kcnd (kv4) potassium channels mediating a-type currents i(to) and i(sa). Genomics 2000, 64, 144–154. [Google Scholar] [CrossRef] [PubMed]
- Bertaso, F.; Sharpe, C.C.; Hendry, B.M.; James, A.F. Expression of voltage-gated k+ channels in human atrium. Basic Res. Cardiol. 2002, 97, 424–433. [Google Scholar] [CrossRef] [PubMed]
- Tamargo, J.; Caballero, R.; Gomez, R.; Valenzuela, C.; Delpon, E. Pharmacology of cardiac potassium channels. Cardiovasc. Res. 2004, 62, 9–33. [Google Scholar] [CrossRef] [PubMed]
- Mahoney, V.M.; Mezzano, V.; Morley, G.E. A review of the literature on cardiac electrical activity between fibroblasts and myocytes. Prog. Biophys. Mol. Biol. 2016, 120, 128–133. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.J.; Wu, S.N.; Lin, M.W.; Sung, R.J. Characterization of Ca2+-activated K+ current in human cardiac fibroblasts: Its contribution to electrical coupling of cardiomyocyte-fibroblast. FASEB J. 2007, 21. [Google Scholar] [CrossRef]
- Zlochiver, S.; Munoz, V.; Vikstrom, K.L.; Taffet, S.M.; Berenfeld, O.; Jalife, J. Electrotonic myofibroblast-to-myocyte coupling increases propensity to reentrant arrhythmias in two-dimensional cardiac monolayers. Biophys. J. 2008, 95, 4469–4480. [Google Scholar] [CrossRef] [PubMed]
- Bae, H.; Lee, D.; Kim, Y.W.; Choi, J.; Lee, H.J.; Kim, S.W.; Kim, T.; Noh, Y.H.; Ko, J.H.; Bang, H.; et al. Effects of hydrogen peroxide on voltage-dependent K(+) currents in human cardiac fibroblasts through protein kinase pathways. Korean J. Physiol. Pharmacol. 2016, 20, 315–324. [Google Scholar] [CrossRef] [PubMed]
- He, M.L.; Liu, W.J.; Sun, H.Y.; Wu, W.; Liu, J.; Tse, H.F.; Lau, C.P.; Li, G.R. Effects of ion channels on proliferation in cultured human cardiac fibroblasts. J. Mol. Cell. Cardiol. 2011, 51, 198–206. [Google Scholar] [CrossRef] [PubMed]
- Cole, W.C.; Clement-Chomienne, O.; Aiello, E.A. Regulation of 4-aminopyridine-sensitive, delayed rectifier k+ channels in vascular smooth muscle by phosphorylation. Biochem. Cell Biol. 1996, 74, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Rogers, P.A.; Chilian, W.M.; Bratz, I.N.; Bryan, R.M., Jr.; Dick, G.M. H2O2 activates redox- and 4-aminopyridine-sensitive kv channels in coronary vascular smooth muscle. Am. J. Physiol. Heart Circ. Physiol. 2007, 292, H1404–H1411. [Google Scholar] [CrossRef] [PubMed]
- Snyders, D.J.; Tamkun, M.M.; Bennett, P.B. A rapidly activating and slowly inactivating potassium channel cloned from human heart. Functional analysis after stable mammalian cell culture expression. J. Gen. Physiol. 1993, 101, 513–543. [Google Scholar] [CrossRef] [PubMed]
- Beech, D.J.; Bolton, T.B. Two components of potassium current activated by depolarization of single smooth muscle cells from the rabbit portal vein. J. Physiol. 1989, 418, 293–309. [Google Scholar] [CrossRef] [PubMed]
- Krick, S.; Platoshyn, O.; Sweeney, M.; McDaniel, S.S.; Zhang, S.; Rubin, L.J.; Yuan, J.X.J. Nitric oxide induces apoptosis by activating K+ channels in pulmonary vascular smooth muscle cells. Am. J. Physiol. Heart Circ. Physiol. 2002, 282, H184–H193. [Google Scholar] [CrossRef] [PubMed]
- Nelson, M.T.; Quayle, J.M. Physiological roles and properties of potassium channels in arterial smooth muscle. Am. J. Physiol. 1995, 268, C799–C822. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, K.; Shintani, Y.; Ding, W.G.; Matsuura, H.; Bamba, T. Potentiation of slow component of delayed rectifier k(+) current by cgmp via two distinct mechanisms: Inhibition of phosphodiesterase 3 and activation of protein kinase g. Br. J. Pharmacol. 2002, 137, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Brady, A.J.B.; Warren, J.B.; Poolewilson, P.A.; Williams, T.J.; Harding, S.E. Nitric-oxide attenuates cardiac myocyte contraction. Am. J. Physiol. 1993, 265, H176–H182. [Google Scholar] [CrossRef] [PubMed]
- Hare, J.M.; Loh, E.; Creager, M.A.; Colucci, W.S. Nitric-oxide inhibits the positive inotropic response to beta-adrenergic stimulation in humans with left-ventricular dysfunction. Circulation 1995, 92, 2198–2203. [Google Scholar] [CrossRef] [PubMed]
- Kamga Pride, C.; Mo, L.; Quesnelle, K.; Dagda, R.K.; Murillo, D.; Geary, L.; Corey, C.; Portella, R.; Zharikov, S.; St Croix, C.; et al. Nitrite activates protein kinase a in normoxia to mediate mitochondrial fusion and tolerance to ischaemia/reperfusion. Cardiovasc. Res. 2014, 101, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Flammer, J.; Haefliger, I.O. Isoproterenol, forskolin, and camp-induced nitric oxide production in pig ciliary processes. Investig. Ophthalmol. Vis. Sci. 1999, 40, 1833–1837. [Google Scholar]
- Ahmmed, G.U.; Xu, Y.; Hong Dong, P.; Zhang, Z.; Eiserich, J.; Chiamvimonvat, N. Nitric oxide modulates cardiac na(+) channel via protein kinase a and protein kinase g. Circ. Res. 2001, 89, 1005–1013. [Google Scholar] [CrossRef] [PubMed]
- Gomez, R.; Caballero, R.; Barana, A.; Amoros, I.; Calvo, E.; Lopez, J.A.; Klein, H.; Vaquero, M.; Osuna, L.; Atienza, F.; et al. Nitric oxide increases cardiac ik1 by nitrosylation of cysteine 76 of kir2.1 channels. Circ. Res. 2009, 105, 383–392. [Google Scholar] [CrossRef] [PubMed]
- Bai, C.X.; Namekata, I.; Kurokawa, J.; Tanaka, H.; Shigenobu, K.; Furukawa, T. Role of nitric oxide in Ca2+ sensitivity of the slowly activating delayed rectifier K+ current in cardiac myocytes. Circ. Res. 2005, 96, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Bunda, S.; Liu, P.; Wang, Y.; Liu, K.; Hinek, A. Aldosterone induces elastin production in cardiac fibroblasts through activation of insulin-like growth factor-i receptors in a mineralocorticoid receptor-independent manner. Am. J. Pathol. 2007, 171, 809–819. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.B.; Tao, R.; Sun, H.Y.; Tse, H.F.; Lau, C.P.; Li, G.R. Multiple Ca2+ signaling pathways regulate intracellular Ca2+ activity in human cardiac fibroblasts. J. Cell. Physiol. 2010, 223, 68–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meredith, A.; Boroomand, S.; Carthy, J.; Luo, Z.S.; McManus, B. 1,25 dihydroxyvitamin d-3 inhibits tgf beta(1)-mediated primary human cardiac myofibroblast activation. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer | Size |
|---|---|---|---|
| GAPDH | 5′-AGCCACATCGCTCAGACACC-3′ | 5′-GTACTCAGCGGCCAGCATCG-3 | 302 |
| KV1.1 | 5′-CCATCATTCCTTATTTCATCAC-3 | 5′-CTCTTCCCCCTCAGTTTCTC-3′ | 488 |
| KV1.2 | 5′-TCCGGGATGAGAATGAAGAC-3′ | 5′-TTGGACAGCTTGTCACTTGC-3′ | 747 |
| KV1.3 | 5′-TCTGCCTATGCCCTTGTTTT-3′ | 5′-TTCCTCCCAGGATGTACTGC-3′ | 259 |
| KV1.5 | 5′-TGCGTCATCTGGTTCACCTTCG-3′ | 5′-TGTTCAGCAAGCCTCCCATTCC-3′ | 906 |
| KV1.6 | 5′-TCAACAGGATGGAAACCAGCCC-3′ | 5′-CTGCCATCTGCAACACGATTCC-3′ | 608 |
| KV1.7 | 5′-CTTCCAGGGGCATGTTATTT-3′ | 5′-CTCAATGGAACTCAATTCAG-3′ | 300 |
| KV2.1 | 5′-ACAGAGCAAACCAAAGGAAGAAC-3′ | 5′-CACCCTCCATGAAGTTGACTTTA-3′ | 383 |
| KV2.2 | 5′-AACGAACTGAGGCGAGAG -3′ | 5′-ACTCCGCCTAAGGGTGAAAC-3′ | 546 |
| KV3.1 | 5′-AACCCCATCGTGAACAAGACGG-3′ | 5′-TCATGGTGACCACGGCCCA-3′ | 550 |
| Gene | Forward Primer | Reverse Primer | Size |
|---|---|---|---|
| GAPDH | 5′-AGCCACATCGCTCAGACACC-3′ | 5′-ATCATTCAACAACCCACCAT-3′ | 302 |
| KV1.4 | 5′-TGGCGGCTACAGTTCAGTCC-3′ | 5′-TGTTGACAATGACGGGCACAGG-3′ | 571 |
| KV3.3 | 5′-TTCTGCCTGGAAACCCATGAGG-3′ | 5′-TGCCAAATCCCAAGGTCTGAGG-3′ | 694 |
| KV3.4 | 5′-TTCAAGCTCACACGCCACTTCG-3′ | 5′-TTCTTTCGGTCCCGATAC-3′ | 656 |
| KV4.1 | 5′-ATCTCGAGGAGATGAGGTTC-3′ | 5′-GATCCGCACGGCACTGTTTC-3′ | 318 |
| KV4.2 | 5′-ATCTTCCGCCACATCCTGAA-3′ | 5′-GATCCGCACGGCACTGTTTC-3′ | 362 |
| KV4.3 | 5′-GATGAGCAGATGTTTGAGCAG-3′ | 5′-AGCAGGTGGTAGTGAGGCC-3′ | 106 |
| KV4.4 | 5′-AGCCAAGAAGAACAAGCTG-3′ | 5′-AGGAAGTTTAGGACATGCC-3′ | 315 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bae, H.; Choi, J.; Kim, Y.-W.; Lee, D.; Kim, J.-H.; Ko, J.-H.; Bang, H.; Kim, T.; Lim, I. Effects of Nitric Oxide on Voltage-Gated K+ Currents in Human Cardiac Fibroblasts through the Protein Kinase G and Protein Kinase A Pathways but Not through S-Nitrosylation. Int. J. Mol. Sci. 2018, 19, 814. https://doi.org/10.3390/ijms19030814
Bae H, Choi J, Kim Y-W, Lee D, Kim J-H, Ko J-H, Bang H, Kim T, Lim I. Effects of Nitric Oxide on Voltage-Gated K+ Currents in Human Cardiac Fibroblasts through the Protein Kinase G and Protein Kinase A Pathways but Not through S-Nitrosylation. International Journal of Molecular Sciences. 2018; 19(3):814. https://doi.org/10.3390/ijms19030814
Chicago/Turabian StyleBae, Hyemi, Jeongyoon Choi, Young-Won Kim, Donghee Lee, Jung-Ha Kim, Jae-Hong Ko, Hyoweon Bang, Taeho Kim, and Inja Lim. 2018. "Effects of Nitric Oxide on Voltage-Gated K+ Currents in Human Cardiac Fibroblasts through the Protein Kinase G and Protein Kinase A Pathways but Not through S-Nitrosylation" International Journal of Molecular Sciences 19, no. 3: 814. https://doi.org/10.3390/ijms19030814