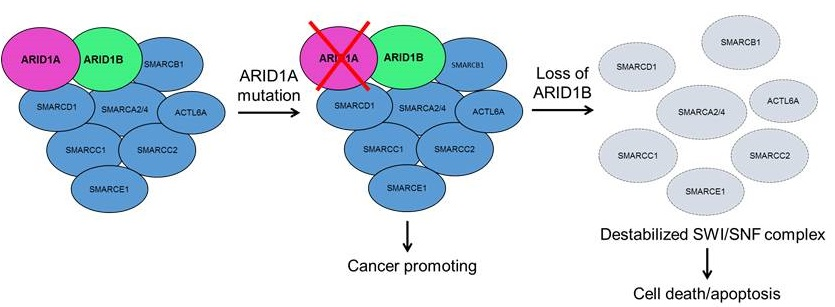
ARID1B as a Potential Therapeutic Target for ARID1A-Mutant Ovarian Clear Cell Carcinoma

Abstract
:
1. Introduction
2. Results
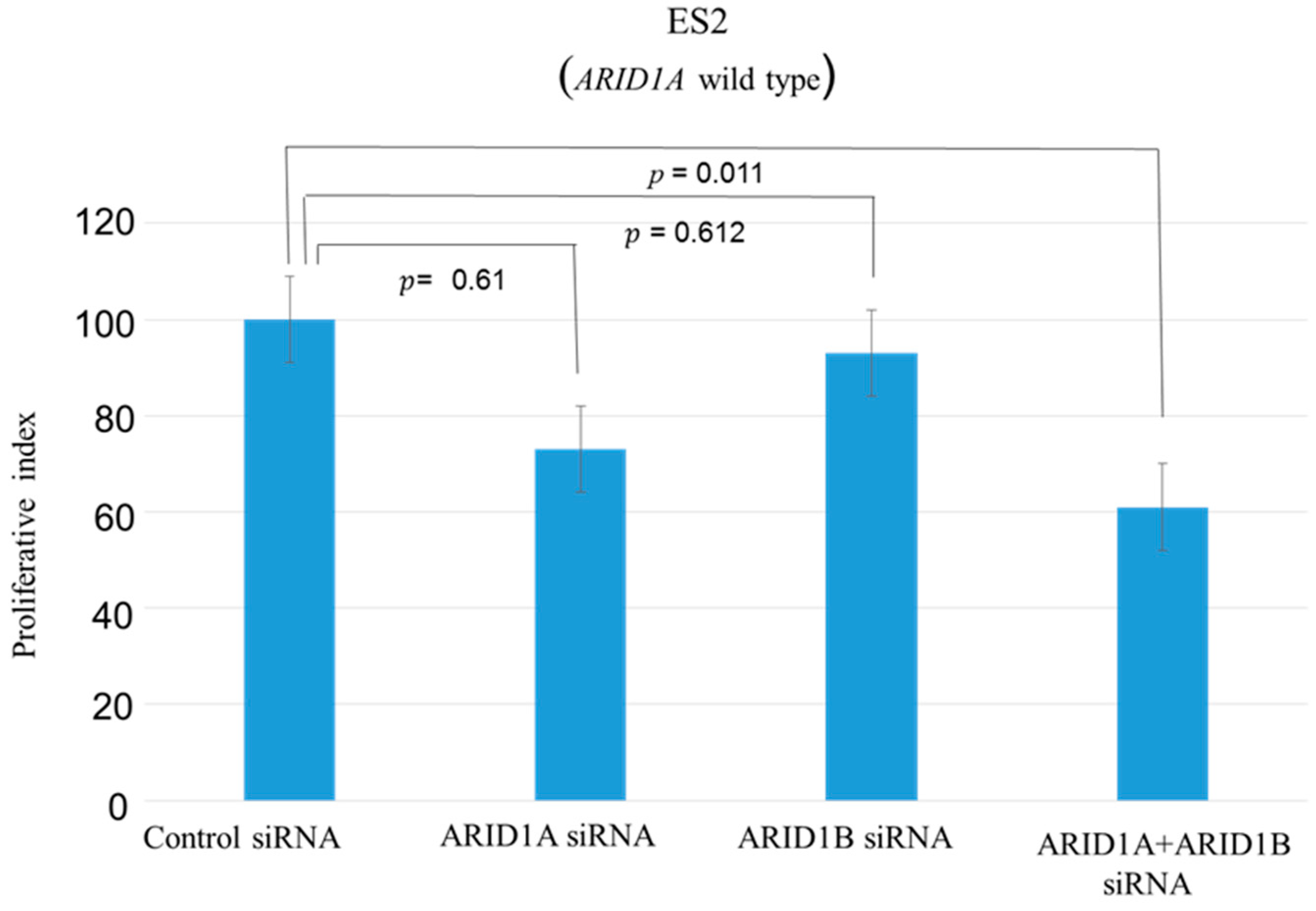
2.1. Effects of ARID1A and ARID1B Knockdown on OCCC In Vitro
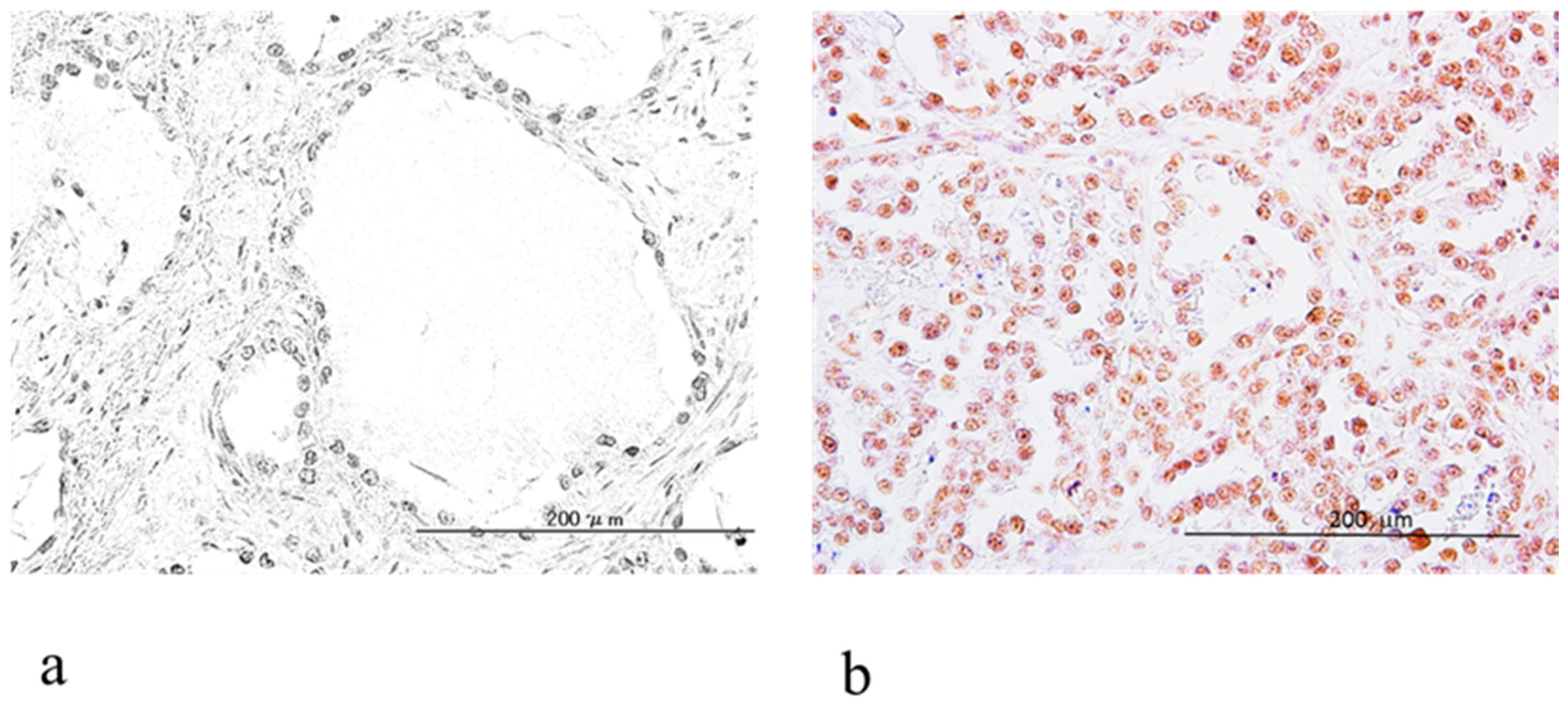
2.2. Relationship between ARID1A and ARID1B Protein Expression in OCCC
2.3. Relationship between ARID1B Expression and Clinicopathologic Factors
2.4. Relationship between ARID1B Protein Expression and Progression-Free Survival
2.5. Relationship between ARID1B Protein Expression and Overall Survival
3. Discussion
4. Materials and Methods
4.1. Tissue Samples
4.2. Immunohistochemistry
4.3. Cell Lines and Culture
4.4. Silencing of ARID1A and ARID1B Gene Expression
4.5. Western Blot Analysis
4.6. Cell Proliferation Assay
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Conflicts of Interest
References
- Wingo, P.A.; Tong, T.; Bolden, S. Cancer statistics 1995. CA Cancer J. Clin. 1995, 45, 8–30. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, T.; Kamura, T.; Kigawa, J.; Terakawa, N.; Kikuchi, Y.; Kita, T.; Suzuki, M.; Sato, I.; Taguchi, K. Clinical characteristics of clear cell carcinoma of the ovary: A distinct histologic type with poor prognosis and resistance to platinum-based chemotherapy. Cancer 2000, 88, 2584–2589. [Google Scholar] [CrossRef]
- Takano, M.; Kikuchi, Y.; Yaegashi, N.; Kuzuya, K.; Ueki, M.; Tsuda, H.; Suzuki, M.; Kigawa, J.; Takeuchi, S.; Tsuda, H.; et al. Clear cell carcinoma of the ovary: A retrospective multicenter experience of 254 patients with complete surgical staging. Br. J. Cancer 2006, 94, 1369–1374. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, H.; Yamada, Y.; Kanayama, S.; Furukawa, N.; Noguchi, T.; Haruta, S.; Yoshida, S.; Sakata, M.; Sado, T.; Oi, H. The role of hepatocyte nuclear factor-1β in the pathogenesis of clear cell carcinoma of the ovary. Int. J. Gynecol. Cancer 2009, 19, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Wiegand, K.C.; Shah, S.P.; Al-Agha, O.M.; Zhao, Y.; Tse, K.; Zeng, T.; Senz, J.; McConechy, M.K.; Anglesio, M.S.; Kalloger, S.E.; et al. ARID1A mutations in endometriosis-associated ovarian carcinomas. N. Engl. J. Med. 2010, 363, 1532–1543. [Google Scholar] [CrossRef] [PubMed]
- Reisman, D.; Glaros, S.; Thompson, E.A. The SWI/SNF complex and cancer. Oncogene 2009, 28, 1653–1668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helming, K.C.; Wang, X.; Wilson, B.G.; Vazquez, F.; Haswell, J.R.; Manchester, H.E.; Kim, Y.; Kryukov, G.V.; Ghandi, M.; Aguirre, A.J.; et al. ARID1B is a specific vulnerability in ARID1A-mutant cancers. Nat. Med. 2014, 20, 251–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.N.; Roberts, C.W. ARID1A mutations in cancer: Another epigenetic tumor suppressor? Cancer Discov. 2013, 3, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Flores-Alcantar, A.; Gonzalez-Sandoval, A.; Escalante-Alcalde, D.; Lomeli, H. Dynamics of expression of ARID1A and ARID1B subunits in mouse embryos and in cells during the cell cycle. Cell Tissue Res. 2011, 345, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Nagl, N.G., Jr.; Wang, X.; Patsialou, A.; Van Scoy, M.; Moran, E. Distinct mammalian SWI/SNF chromatin remodeling complexes with opposing roles in cell-cycle control. EMBO J. 2007, 26, 752–763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, S.; Wang, T.L.; Shih, I.M.; Mao, T.L.; Nakayama, K.; Roden, R.; Glas, R.; Slamon, D.; Diaz, L.A., Jr.; Vogelstein, B.; et al. Frequent mutations of chromatin remodeling gene ARID1A in ovarian clear cell carcinoma. Science 2010, 330, 228–231. [Google Scholar] [CrossRef] [PubMed]
- Katagiri, A.; Nakayama, K.; Rahman, M.T.; Rahman, M.; Katagiri, H.; Nakayama, N.; Ishikawa, M.; Ishibashi, T.; Iida, K.; Kobayashi, H.; et al. Loss of ARID1A expression is related to shorter progression-free survival and chemoresistance in ovarian clear cell carcinoma. Mod. Pathol. 2012, 25, 282–288. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.T.; Nakayama, K.; Rahman, M.; Nakayama, N.; Ishikawa, M.; Katagiri, A.; Iida, K.; Nakayama, S.; Otsuki, Y.; Shih, I.M.; et al. Prognostic and therapeutic impact of the chromosome 20q13.2 ZNF217 locus amplification in ovarian clear cell carcinoma. Cancer 2012, 118, 2846–2857. [Google Scholar] [CrossRef] [PubMed]
- Sausen, M.; Leary, R.J.; Jones, S.; Wu, J.; Reynolds, C.P.; Liu, X.; Blackford, A.; Parmigiani, G.; Diaz, L.A., Jr.; Papadopoulos, N.; et al. Integrated genomic analyses identify ARID1A and ARID1B alterations in the childhood cancer neuroblastoma. Nat. Genet. 2013, 45, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, A.; Totoki, Y.; Abe, T.; Boroevich, K.A.; Hosoda, F.; Nguyen, H.H.; Aoki, M.; Hosono, N.; Kubo, M.; Miya, F.; et al. Whole-genome sequencing of liver cancers identifies etiological influences on mutation patterns and recurrent mutations in chromatin regulators. Nat. Genet. 2012, 44, 760–764. [Google Scholar] [CrossRef] [PubMed]
- Coatham, M.; Li, X.; Karnezis, A.N.; Hoang, L.N.; Tessier-Cloutier, B.; Meng, B.; Soslow, R.A.; Blake Gilks, C.; Huntsman, D.G.; Stewart, C.J.; et al. Concurrent ARID1A and ARID1B inactivation in endometrial and ovarian dedifferentiated carcinomas. Mod. Pathol. 2016, 29, 1586–1593. [Google Scholar] [CrossRef] [PubMed]
- McIntyre, J.B.; Wu, J.S.; Craighead, P.S.; Phan, T.; Köbel, M.; Lees-Miller, S.P.; Ghatage, P.; Magliocco, A.M.; Doll, C.M. PIK3CA mutational status and overall survival in patients with cervical cancer treated with radical chemoradiotherapy. Gynecol. Oncol. 2013, 128, 409–414. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Intensity of Expression | Patients Number | ARID1A Loss | ARID1A Positive |
|---|---|---|---|
| ARID1B Low | 8 | 4 (50%) | 4 (50%) |
| ARID1B High | 45 | 3 (6%) | 42 (94%) |
| Factors | Patients | ARID1B Immunostaining | p-Value | |
|---|---|---|---|---|
| Low | High | |||
| FIGO stage | ||||
| I, II | 40 | 5 | 35 | 0.3547 |
| III, IV | 13 | 3 | 10 | |
| CA125 U/mL | ||||
| <90 | 27 | 4 | 23 | 0.9538 |
| ≥90 | 26 | 4 | 22 | |
| Age (years) | ||||
| <54 | 27 | 4 | 23 | 0.9538 |
| ≥54 | 26 | 4 | 22 | |
| Endometriosis | ||||
| Without | 29 | 5 | 24 | 0.6313 |
| With | 24 | 3 | 21 | |
| Ki-67 | ||||
| Low | 25 | 3 | 22 | 0.5521 |
| High | 28 | 5 | 23 | |
| Residual tumor | ||||
| <2 cm | 41 | 6 | 35 | 0.8627 |
| ≥2 cm | 12 | 2 | 10 | |
| Factors | Patients | Hazard Ratio | 95% CI | p Value |
|---|---|---|---|---|
| FIGO stage | ||||
| I, II | 40 | 6.3 | 2.7–14.7 | <0.0001 |
| III, IV | 13 | |||
| CA125 U/mL | ||||
| <90 | 27 | 3.5 | 1.4–9.0 | 0.0088 |
| ≥90 | 26 | |||
| Age (years) | ||||
| <54 | 27 | 1.7 | 0.7–4.1 | 0.2278 |
| ≥54 | 26 | |||
| Endometriosis | ||||
| Without | 29 | 0.5 | 0.2–1.3 | 0.1851 |
| With | 24 | |||
| Ki-67 | ||||
| Low | 25 | 1.7 | 0.7–3.9 | 0.2323 |
| High | 28 | |||
| Residual tumor | ||||
| <2 cm | 41 | 6.1 | 2.6–14.2 | <0.0001 |
| ≥2 cm | 12 | |||
| ARID1B immunostaining | ||||
| Low | 8 | 3.9 | 1.4–8.9 | 0.04 |
| High | 45 |
| Factors | Patients | Hazard Ratio | 95% CI | p Value |
|---|---|---|---|---|
| FIGO stage | ||||
| I, II | 40 | 1.6 | 0.3–8.0 | 0.5624 |
| III, IV | 13 | |||
| CA125 U/mL | ||||
| <90 | 27 | 3.2 | 1.0–9.8 | 0.0446 |
| ≥90 | 25 | |||
| Residual tumor | ||||
| <2 cm | 41 | 3.4 | 0.7–16.1 | 0.0692 |
| ≥2 cm | 12 | |||
| ARID1B immunostaining | ||||
| Low | 8 | 3.1 | 1.1–8.7 | 0.036 |
| High | 45 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sato, E.; Nakayama, K.; Razia, S.; Nakamura, K.; Ishikawa, M.; Minamoto, T.; Ishibashi, T.; Yamashita, H.; Iida, K.; Kyo, S. ARID1B as a Potential Therapeutic Target for ARID1A-Mutant Ovarian Clear Cell Carcinoma. Int. J. Mol. Sci. 2018, 19, 1710. https://doi.org/10.3390/ijms19061710
Sato E, Nakayama K, Razia S, Nakamura K, Ishikawa M, Minamoto T, Ishibashi T, Yamashita H, Iida K, Kyo S. ARID1B as a Potential Therapeutic Target for ARID1A-Mutant Ovarian Clear Cell Carcinoma. International Journal of Molecular Sciences. 2018; 19(6):1710. https://doi.org/10.3390/ijms19061710
Chicago/Turabian StyleSato, Emi, Kentaro Nakayama, Sultana Razia, Kohei Nakamura, Masako Ishikawa, Toshiko Minamoto, Tomoka Ishibashi, Hitomi Yamashita, Kouji Iida, and Satoru Kyo. 2018. "ARID1B as a Potential Therapeutic Target for ARID1A-Mutant Ovarian Clear Cell Carcinoma" International Journal of Molecular Sciences 19, no. 6: 1710. https://doi.org/10.3390/ijms19061710
APA StyleSato, E., Nakayama, K., Razia, S., Nakamura, K., Ishikawa, M., Minamoto, T., Ishibashi, T., Yamashita, H., Iida, K., & Kyo, S. (2018). ARID1B as a Potential Therapeutic Target for ARID1A-Mutant Ovarian Clear Cell Carcinoma. International Journal of Molecular Sciences, 19(6), 1710. https://doi.org/10.3390/ijms19061710