Blooming of Unusual Cytochrome P450s by Tandem Duplication in the Pathogenic Fungus Conidiobolus coronatus

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
2.1. C. coronatus Has the Highest Number of P450s among Entomopathogenic Fungi
2.2. Large Number of New P450s Found in C. coronatus
2.3. P450 Signature Motifs EXXR and CXG Are Conserved in C. coronatus P450s
2.4. P450 Blooming in C. coronatus
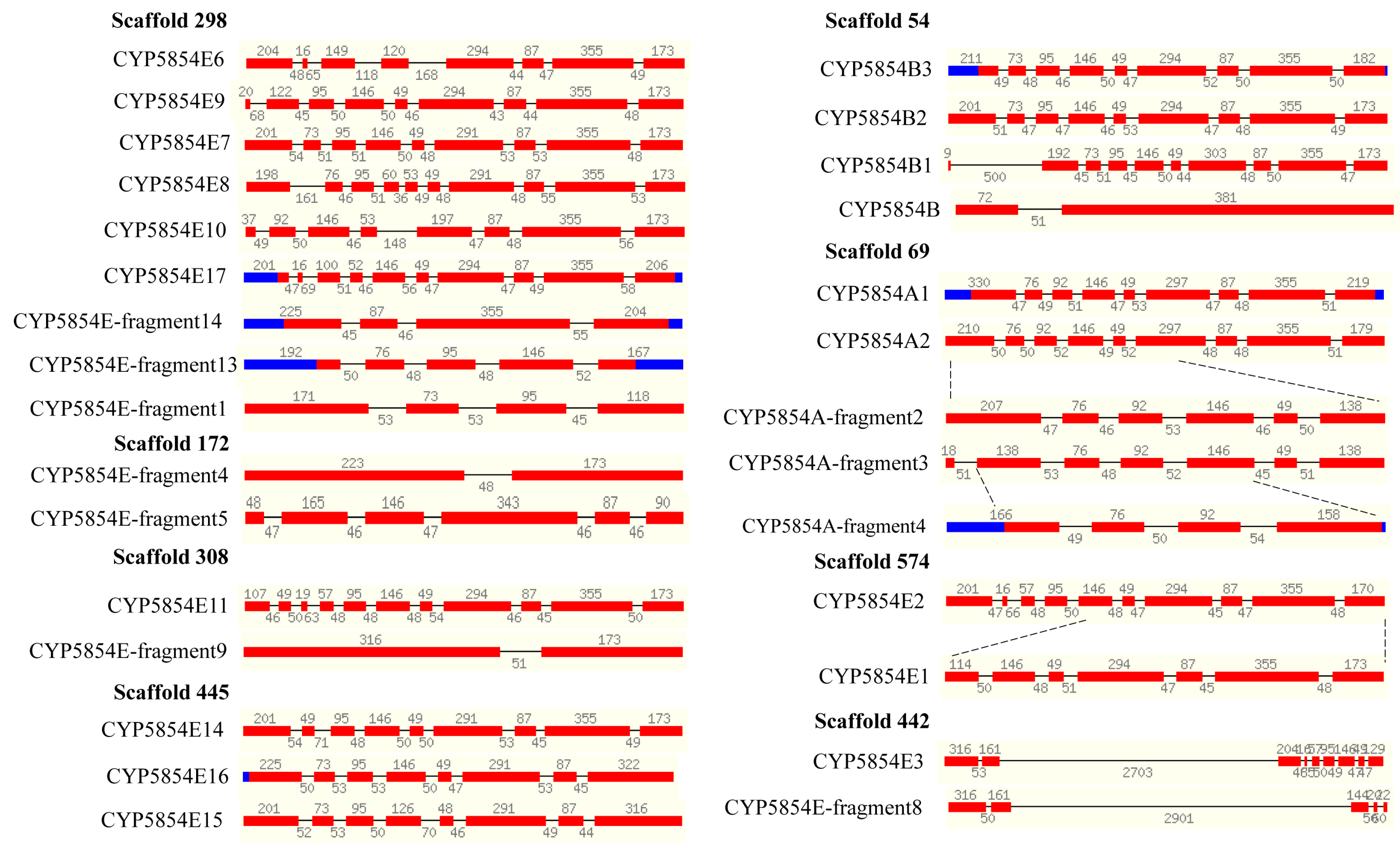
2.5. Extensive Tandem Duplications Led to P450 Blooming in C. coronatus
2.6. C. coronatus P450s Have Same Gene Structure
2.7. C. coronatus Has the Lowest P450 Diversity among Entomopathogenic Fungi
2.8. Functional Prediction of C. coronatus P450s
3. Materials and Methods
3.1. Genome Data Mining for P450s
3.2. Annotation of P450s
3.3. Phylogenetic Analysis of P450s
3.4. Analysis of EXXR and CXG Motifs
3.5. Identification of Tandemly Duplicated P450s
3.6. Gene Structure Analysis
3.7. P450 Diversity Percentage Analysis
3.8. Comparative Analysis of P450s
4. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Nelson, D.R. Cytochrome P450 diversity in tree of life. Biochim. Biophys. Acta 2018, 1866, 141–154. [Google Scholar] [CrossRef] [PubMed]
- Bernhardt, R. Cytochromes P450 as versatile biocatalysts. J. Biotechnol. 2006, 124, 128–145. [Google Scholar] [CrossRef] [PubMed]
- Montellano, P.R.O. Cytochrome P450: Structure, Mechanism, and Biochemistry, 4th ed.; Springer: Basel, Switzerland, 2015; pp. 1–912. ISBN 978-3-319-12108-6. [Google Scholar]
- Syed, K.; Shale, K.; Pagadala, N.S.; Tuszynski, J. Systematic identification and evolutionary analysis of catalytically versatile cytochrome P450 monooxygenase families enriched in model basidiomycete fungi. PLoS ONE 2014, 9, e86683. [Google Scholar] [CrossRef] [PubMed]
- Kgosiemang, I.K.R.; Mashele, S.S.; Syed, K. Comparative genomics and evolutionary analysis of cytochrome P450 monooxygenases in fungal subphylum Saccharomycotina. J. Pure Appl. Microbiol. 2014, 8, 291–302. [Google Scholar]
- Qhanya, L.B.; Matowane, G.; Chen, W.; Sun, Y.; Letsimo, E.M.; Parvez, M.; Yu, J.H.; Mashele, S.S.; Syed, K. Genome-wide annotation and comparative analysis of cytochrome P450 monooxygenases in Basidiomycete biotrophic plant pathogens. PLoS ONE 2015, 10, e0142100. [Google Scholar] [CrossRef] [PubMed]
- Matowane, R.G.; Wieteska, L.; Bamal, H.D.; Kgosiemang, I.K.R.; Van Wyk, M.; Manume, N.A.; Abdalla, S.M.H.; Mashele, S.S.; Gront, D.; Syed, K. In silico analysis of cytochrome P450 monooxygenases in chronic granulomatous infectious fungus Sporothrix schenckii: Special focus on CYP51. Biochim. Biophys. Acta 2018, 1866, 166–177. [Google Scholar] [CrossRef] [PubMed]
- Sakaki, T. Practical application of cytochrome P450. Biol. Pharm. Bull. 2012, 35, 844–849. [Google Scholar] [CrossRef] [PubMed]
- Girhard, M.; Bakkes, P.J.; Mahmoud, O.; Urlacher, V.B. P450 Biotechnology. In Cytochrome P450: Structure, Mechanism, and Biochemistry, 4th ed.; Montellano, P.R.O., Ed.; Springer: Basel, Switzerland, 2015; pp. 451–520. ISBN 978-3-319-12108-6. [Google Scholar]
- Durairaj, P.; Hur, J.S.; Yun, H. Versatile biocatalysis of fungal cytochrome P450 monooxygenases. Microb. Cell Fact. 2016, 15, 125. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.; Kim, J.E.; Lee, Y.W.; Son, H. Fungal cytochrome P450s and the P450 complement (CYPome) of Fusarium graminearum. Toxins 2018, 10, 112. [Google Scholar] [CrossRef] [PubMed]
- Kelly, S.L.; Kelly, D.E. Microbial cytochromes P450: Biodiversity and biotechnology. Where do cytochromes P450 come from, what do they do and what can they do for us? Phil. Trans. R. Soc. B 2013, 368. [Google Scholar] [CrossRef] [PubMed]
- Jawallapersand, P.; Mashele, S.S.; Kovačič, L.; Stojan, J.; Komel, R.; Pakala, S.B.; Kraševec, N.; Syed, K. Cytochrome P450 monooxygenase CYP53 family in fungi: Comparative structural and evolutionary analysis and its role as a common alternative anti-fungal drug target. PLoS ONE 2014, 9, e107209. [Google Scholar] [CrossRef] [PubMed]
- Sheehan, D.J.; Hitchcock, C.A.; Sibley, C.M. Current and emerging azole antifungal agents. Clin. Microbiol. Rev. 1999, 12, 40–79. [Google Scholar] [PubMed]
- Shapiro, R.S.; Robbins, N.; Cowen, L.E. Regulatory circuitry governing fungal development, drug resistance, and disease. Microbiol. Mol. Biol. Rev. 2011, 75, 213–267. [Google Scholar] [CrossRef] [PubMed]
- Hargrove, T.Y.; Wawrzak, Z.; Lamb, D.C.; Guengerich, F.P.; Lepesheva, G.I. Structure-fuctional characterization of cytochrome P450 sterol 14α-demethylase (CYP51B) from Aspergillus fumigatus and molecular basis for the development of antifungal drugs. J. Biol. Chem. 2015, 290, 23916–23934. [Google Scholar] [CrossRef] [PubMed]
- Sanglard, D. Emerging threats in antifungal-resistant fungal pathogens. Front. Med. 2016, 3, 11. [Google Scholar] [CrossRef] [PubMed]
- Ghannoum, M.A.; Rice, L.B. Antifungal agents: Mode of action, mechanisms of resistance, and correlation of these mechanisms with bacterial resistance. Clin. Microbiol. Rev. 1999, 12, 501–517. [Google Scholar] [PubMed]
- Shaikh, N.; Hussain, K.A.; Petraitiene, R.; Schuetz, A.N.; Walsh, T.J. Entomophthoramycosis: A neglected tropical mycosis. Clin. Microbiol. Infect. 2016, 22, 688–694. [Google Scholar] [CrossRef] [PubMed]
- Page, R.M.; Humber, R.A. Phototropism in Conidiobolus coronatus. Mycologia 1973, 65, 335–354. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.F.; Callaghan, A.A. Quantitative survey of Conidiobolus and Basidiobolus in soils and litter. Trans. Br. Mycol. Soc. 1987, 89, 179–185. [Google Scholar] [CrossRef]
- Subramanian, C.; Sobel, J.D. A case of Conidiobolus coronatus in the vagina. Med. Mycol. 2011, 49, 427–429. [Google Scholar] [CrossRef] [PubMed]
- Chowdhary, A.; Randhawa, H.S.; Khan, Z.U.; Ahmad, S.; Khanna, G.; Gupta, R.; Chakravarti, A.; Roy, P. Rhinoentomophthoromycosis due to Conidiobolus coronatus. A case report and an overview of the disease in India. Med. Mycol. 2010, 48, 870–879. [Google Scholar] [CrossRef] [PubMed]
- Dutta, S.; Sarkar, S.; Linka, U.; Dora, S. Conidiobolomycosis: A case report of rare fungal infection from the eastern India. Indian Dermatol. Online J. 2015, 6, 393–395. [Google Scholar] [CrossRef] [PubMed]
- Hoogendijk, C.F.; Pretorius, E.; Marx, J.; van Heerden, W.E.P.; Imhof, A.; Schneemann, M. Detection of villous conidia of Conidiobolus coronatus in a blood sample by scanning electron microscopy investigation. Ultrastruct. Pathol. 2006, 30, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Gugnani, H.C. A review of zygomycosis due to Basidiobolus ranarum. Eur. J. Epidemiol. 1999, 15, 923–929. [Google Scholar] [CrossRef] [PubMed]
- Rippon, J.W. Medical Mycology: The Pathogenic Fungi and the Pathogenic Actinomycetes, 3rd ed.; W.B. Saunders Co.: Philadelphia, PA, USA, 1988; pp. 506–531. ISBN 0-7216-2444-8. [Google Scholar]
- Kwon-Chung, K.J.; Bennett, J.E. Medical Mycology; Lea and Febiger: Philadelphia, PA, USA, 1992; p. 866. ISBN 0-8121-1463-9. [Google Scholar]
- Ribes, J.A.; Vanover-Sams, C.L.; Baker, D.J. Zygomycetes in human disease. Clin. Microbiol. Rev. 2000, 13, 236–301. [Google Scholar] [CrossRef] [PubMed]
- Ellis, D.H. Systemic zygomycosis. In Topley and Wilson’ Microbiology and Microbial Infections. Medical Mycology, 10th ed.; Merz, W.G., Hay, R.J., Eds.; Edward Arnold: London, UK, 2005; pp. 659–686. [Google Scholar]
- Paszkiewicz, M.; Tyma, M.; Ligęza-Żuber, M.; Włóka, E.; Boguś, M.I.; Stepnowski, P. Mycotoxin production by entomopathogenic fungus Conidiobolus coronatus. Int. J. Environ. Agric. Res. 2017, 3, 33–40. [Google Scholar]
- Paszkiewicz, M.; Tyma, M.; Ligęza-Żuber, M.; Włóka, E.; Boguś, M.I.; Stepnowski, P. Trichothecenes production by entomopathogenic fungus Conidiobolus Coronatus. Adv. Toxicol. Toxic Eff. 2016, 1, 7–14. [Google Scholar] [CrossRef]
- Bogus, M.I.; Maria, S. Histopathology of Conidiobolus coronatus (entomophthorales) infection in Galleria mellonella (lepidoptera) larvae. Acta Parasitol. 2000, 45, 48–54. [Google Scholar]
- Malinowski, H. Entomopathogenic fungi as insecticides in protecting the forest. Prog. Plant Prot. 2009, 49, 865–873. [Google Scholar]
- Chang, Y.; Wang, S.; Sekimoto, S.; Aerts, A.L.; Choi, C.; Clum, A.; LaButti, K.M.; Lindquist, E.A.; Yee Ngan, C.; Ohm, R.A.; et al. Phylogenomic analyses indicate that early fungi evolved digesting cell walls of algal ancestors of land plants. Genome Biol. Evol. 2015, 7, 1590–1601. [Google Scholar] [CrossRef] [PubMed]
- Valle, A.C.; Wanke, B.; Lazéra, M.S.; Monteiro, P.C.; Viegas, M.L. Entomophthoramycosis by Conidiobolus coronatus. Report of a case successfully treated with the combination of itraconazole and fluconazole. Rev. Inst. Med. Trop. Sao Paulo 2001, 43, 233–236. [Google Scholar] [CrossRef] [PubMed]
- Choon, S.E.; Kang, J.; Neafie, R.C.; Ragsdale, B.; Klassen-Fischer, M.; Carlson, J.A. Conidiobolomycosis in a young Malaysian woman showing chronic localized fibrosing leukocytoclastic vasculitis: A case report and meta-analysis focusing on clinicopathologic and therapeutic correlations with outcome. Am. J. Dermatopathol. 2012, 34, 511–522. [Google Scholar] [CrossRef] [PubMed]
- Guarro, J.; Aguilar, C.; Pujol, I. In-vitro antifungal susceptibilities of Basidiobolus and Conidiobolus spp. strains. J. Antimicrob. Chemother. 1999, 44, 557–560. [Google Scholar] [CrossRef] [PubMed]
- Xiao, G.; Ying, S.H.; Zheng, P.; Wang, Z.L.; Zhang, S.; Xie, X.Q.; Shang, Y.; Leger, R.J.S.; Zhao, G.P.; Wang, C.; et al. Genomic perspectives on the evolution of fungal entomopathogenicity in Beauveria bassiana. Sci. Rep. 2012, 2, 483. [Google Scholar] [CrossRef] [PubMed]
- Zheng, P.; Xia, Y.; Xiao, G.; Xiong, C.; Hu, X.; Zhang, S.; Zheng, H.; Huang, Y.; Zhou, Y.; Wang, S.; et al. Genome sequence of the insect pathogenic fungus Cordyceps militaris, a valued traditional Chinese medicine. Genome Biol. 2011, 12, R116. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Jin, K.; Ying, S.H.; Zhang, Y.; Xiao, G.; Shang, Y.; Duan, Z.; Hu, X.; Xie, X.Q.; Zhou, G.; et al. Genome sequencing and comparative transcriptomics of the model entomopathogenic fungi Metarhizium anisopliae and M. acridum. PLoS Genet. 2011, 7, e1001264. [Google Scholar] [CrossRef] [PubMed]
- Cuomo, C.A.; Rodriguez-Del Valle, N.; Perez-Sanchez, L.; Abouelleil, A.; Goldberg, J.; Young, S.; Zeng, Q.; Birren, B.W. Genome sequence of the pathogenic fungus Sporothrix schenckii (ATCC 58251). Genome Announc. 2014, 2. [Google Scholar] [CrossRef] [PubMed]
- Lepesheva, G.I.; Waterman, M.R. Sterol 14α-demethylase cytochrome P450 (CYP51), a P450 in all biological kingdoms. Biochim. Biophys. Acta 2007, 1770, 467–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelly, S.L.; Lamb, D.C.; Baldwin, B.C.; Corran, A.J.; Kelly, D.E. Characterization of Saccharomyces cerevisiae CYP61, Sterol Δ22-desaturase, and inhibition by azole antifungal agents. J. Biol. Chem. 1997, 272, 9986–9988. [Google Scholar] [CrossRef] [PubMed]
- Gotoh, O. Substrate recognition sites in cytochrome P450 family 2 (CYP2) proteins inferred from comparative analyses of amino acid and coding nucleotide sequences. J. Biol. Chem. 1992, 267, 83–90. [Google Scholar] [PubMed]
- Sirim, D.; Widmann, M.; Wagner, F.; Pleiss, J. Prediction and analysis of the modular structure of cytochrome P450 monooxygenases. BMC Struct. Biol. 2010, 10, 34. [Google Scholar] [CrossRef] [PubMed]
- Gricman, Ł.; Vogel, C.; Pleiss, J. Conservation analysis of class-specific positions in cytochrome P450 monooxygenases: Functional and structural relevance. Proteins 2014, 82, 491–504. [Google Scholar] [CrossRef] [PubMed]
- Syed, K.; Mashele, S.S. Comparative analysis of P450 signature motifs EXXR and CXG in the large and diverse kingdom of fungi: Identification of evolutionarily conserved amino acid patterns characteristic of P450 family. PLoS ONE 2014, 9, e95616. [Google Scholar] [CrossRef] [PubMed]
- Sello, M.M.; Jafta, N.; Nelson, D.R.; Chen, W.; Yu, J.H.; Parvez, M.; Kgosiemang, I.K.R.; Monyaki, R.; Raselemane, S.C.; Qhanya, L.B.; et al. Diversity and evolution of cytochrome P450 monooxygenases in Oomycetes. Sci. Rep. 2015, 5, 11572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feyereisen, R. Arthropod CYPomes illustrate the tempo and mode in P450 evolution. Biochim. Biophys. Acta 2011, 1814, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Parvez, M.; Qhanya, L.B.; Mthakathi, N.T.; Kgosiemang, I.K.R.; Bamal, H.D.; Pagadala, N.S.; Xie, T.; Yang, H.; Chen, H.; Theron, C.W.; et al. Molecular evolutionary dynamics of cytochrome P450 monooxygenases across kingdoms: Special focus on mycobacterial P450s. Sci. Rep. 2016, 6, 33099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Syed, K.; Shale, K.; Nazir, K.N.H.; Krasevec, N.; Mashele, S.S.; Pagadala, N.S. Genome-wide identification, annotation and characterization of novel thermostable cytochrome P450 monooxygenases from the thermophilic biomass-degrading fungi Thielavia terrestris and Myceliophthora thermophila. Genes Genom. 2014, 36, 321–333. [Google Scholar] [CrossRef]
- Marchler-Bauer, A.; Bo, Y.; Han, L.; He, J.; Lanczycki, C.J.; Lu, S.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; et al. CDD/SPARCLE: Functional classification of proteins via subfamily domain architectures. Nucleic Acids Res. 2016, 45, D200–D203. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.R. The cytochrome p450 homepage. Hum. Genom. 2009, 4, 59–65. [Google Scholar]
- Nelson, D.R.; Kamataki, T.; Waxman, D.J.; Guengerich, F.P.; Estabrook, R.W.; Feyereisen, R.; Gonzalez, F.J.; Coon, M.J.; Gunsalus, I.C.; Gotoh, O.; et al. The P450 superfamily: Update on new sequences, gene mapping, accession numbers, early trivial names of enzymes, and nomenclature. DNA Cell Biol. 1993, 12, 1–51. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.R. Cytochrome P450 nomenclature. Methods Mol. Biol. 1998, 107, 15–24. [Google Scholar] [PubMed]
- Nelson, D.R. Cytochrome P450 Nomenclature, 2004. Methods Mol. Biol. 2006, 320, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Trifinopoulos, J.; Nguyen, L.T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive tree of life (iTOL) v3: An online tool for the display and annotation of phylogenetic and other trees. Nucleic Acids Res. 2016, 44, W242–W245. [Google Scholar] [CrossRef] [PubMed]
- Sievers, F.; Higgins, D.G. Clustal Omega, accurate alignment of very large numbers of sequences. Methods Mol. Biol. 2014, 1079, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Crooks, G.E.; Hon, G.; Chandonia, J.M.; Brenner, S.E. WebLogo: A sequence logo generator. Genome Res. 2004, 14, 1188–1190. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| P450 Family | P450 Subfamily | No. of P450s in a Family | |
|---|---|---|---|
| Subfamily Name | No. of P450s | ||
| CYP51 | F | 1 | 1 |
| CYP5854 | A | 8 | 48 |
| B | 4 | ||
| C | 1 | ||
| D | 1 | ||
| E | 34 | ||
| CYP5855 * | A | 5 | 36 |
| B | 2 | ||
| C | 12 | ||
| D | 15 | ||
| E | 1 | ||
| 1 | |||
| CYP5856 | A | 5 | 20 |
| B | 15 | ||
| CYP5857 | A | 10 | 10 |
| CYP5858 | A | 11 | 11 |
| CYP5859 | A | 5 | 5 |
| CYP5860 | A | 3 | 3 |
| CYP5861 | A | 3 | 3 |
| CYP5862 | A | 2 | 2 |
| CYP5863 | A | 1 | 1 |
| CYP5864 | A | 1 | 1 |
| CYP-fragment # | - | 1 | 1 |
| 12 Families | 21 subfamilies | 142 | 142 |
| Fungus | Host | P450 Count | No. of P450 Families | Reference |
|---|---|---|---|---|
| Entomopathogenic fungus | ||||
| Beauveria bassiana | Arthropods (termites, thrips, whiteflies, aphids, and beetles) | 83 | 49 | [39] |
| Cordyceps militaris | Butterflies and caterpillars | 57 | 37 | [40] |
| Metarhizium acridum | ≥200 insects | 100 | 67 | [41] |
| Metarhizium anisopliae (formerly Metarhizium robertsii) | Locusts | 123 | 87 | [41] |
| Animal including human pathogen | ||||
| Sporothrix schenckii | Humans and other animals (cats, dogs, rodents, squirrels, horses, and birds) | 40 | 32 | [7,42] |
| Animals (including humans) and entomopathogenic fungus | ||||
| Conidiobolus coronatus | Insects, humans, and other animals (horses, sheep and dogs) | 142 | 12 | This work |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ngwenya, M.L.; Chen, W.; Basson, A.K.; Shandu, J.S.; Yu, J.-H.; Nelson, D.R.; Syed, K. Blooming of Unusual Cytochrome P450s by Tandem Duplication in the Pathogenic Fungus Conidiobolus coronatus. Int. J. Mol. Sci. 2018, 19, 1711. https://doi.org/10.3390/ijms19061711
Ngwenya ML, Chen W, Basson AK, Shandu JS, Yu J-H, Nelson DR, Syed K. Blooming of Unusual Cytochrome P450s by Tandem Duplication in the Pathogenic Fungus Conidiobolus coronatus. International Journal of Molecular Sciences. 2018; 19(6):1711. https://doi.org/10.3390/ijms19061711
Chicago/Turabian StyleNgwenya, Mathula Lancelot, Wanping Chen, Albert Kotze Basson, Jabulani Siyabonga Shandu, Jae-Hyuk Yu, David R. Nelson, and Khajamohiddin Syed. 2018. "Blooming of Unusual Cytochrome P450s by Tandem Duplication in the Pathogenic Fungus Conidiobolus coronatus" International Journal of Molecular Sciences 19, no. 6: 1711. https://doi.org/10.3390/ijms19061711