BAS2 Is Required for Conidiation and Pathogenicity of Colletotrichum gloeosporioides from Hevea brasiliensis


Abstract
:1. Introduction
2. Results
2.1. Generation of the ∆BAS2 Mutant
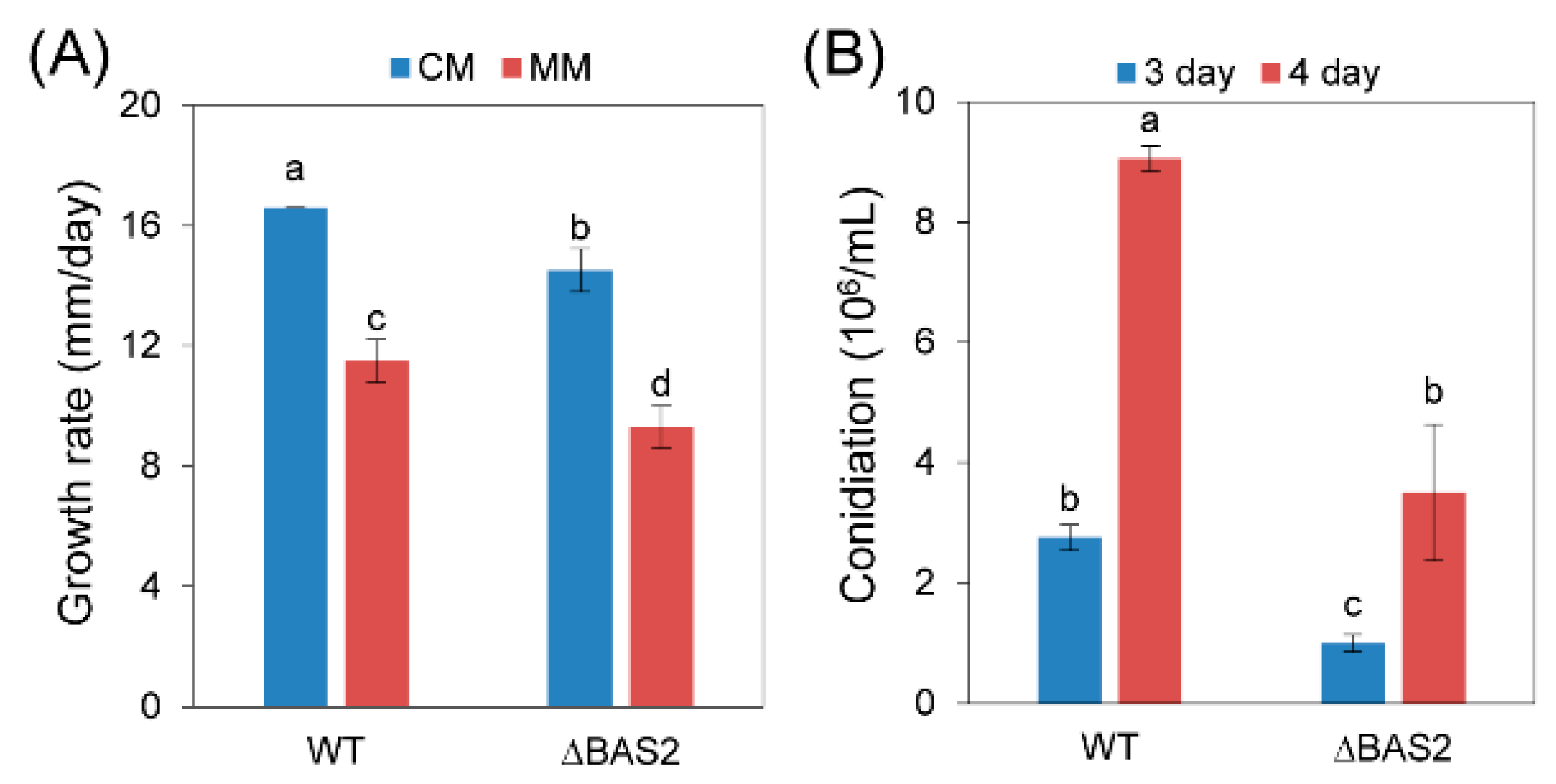
2.2. BAS2 Is Involved in Vegetative Growth and Conidiation
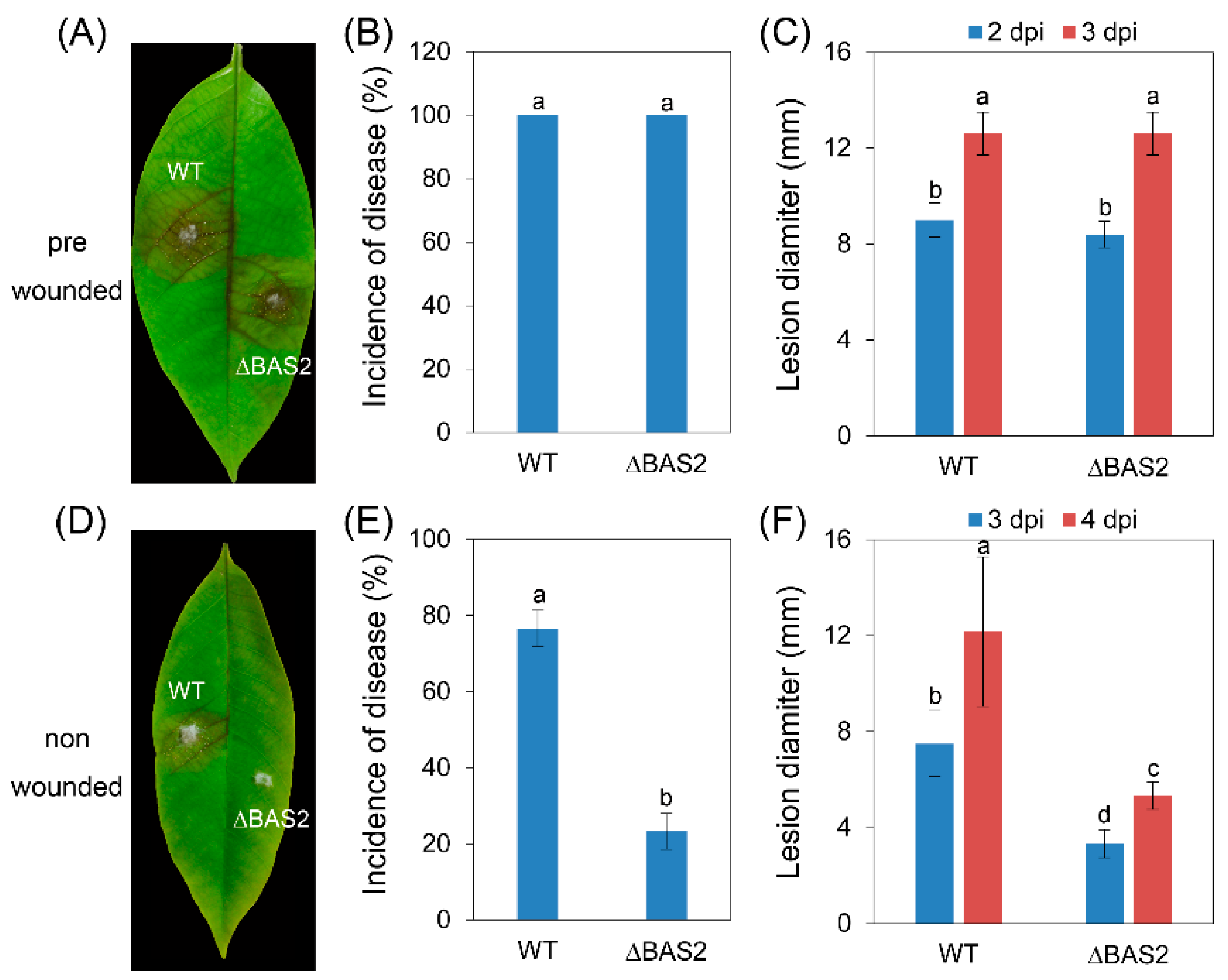
2.3. BAS2 Are Required for Pathogenicity
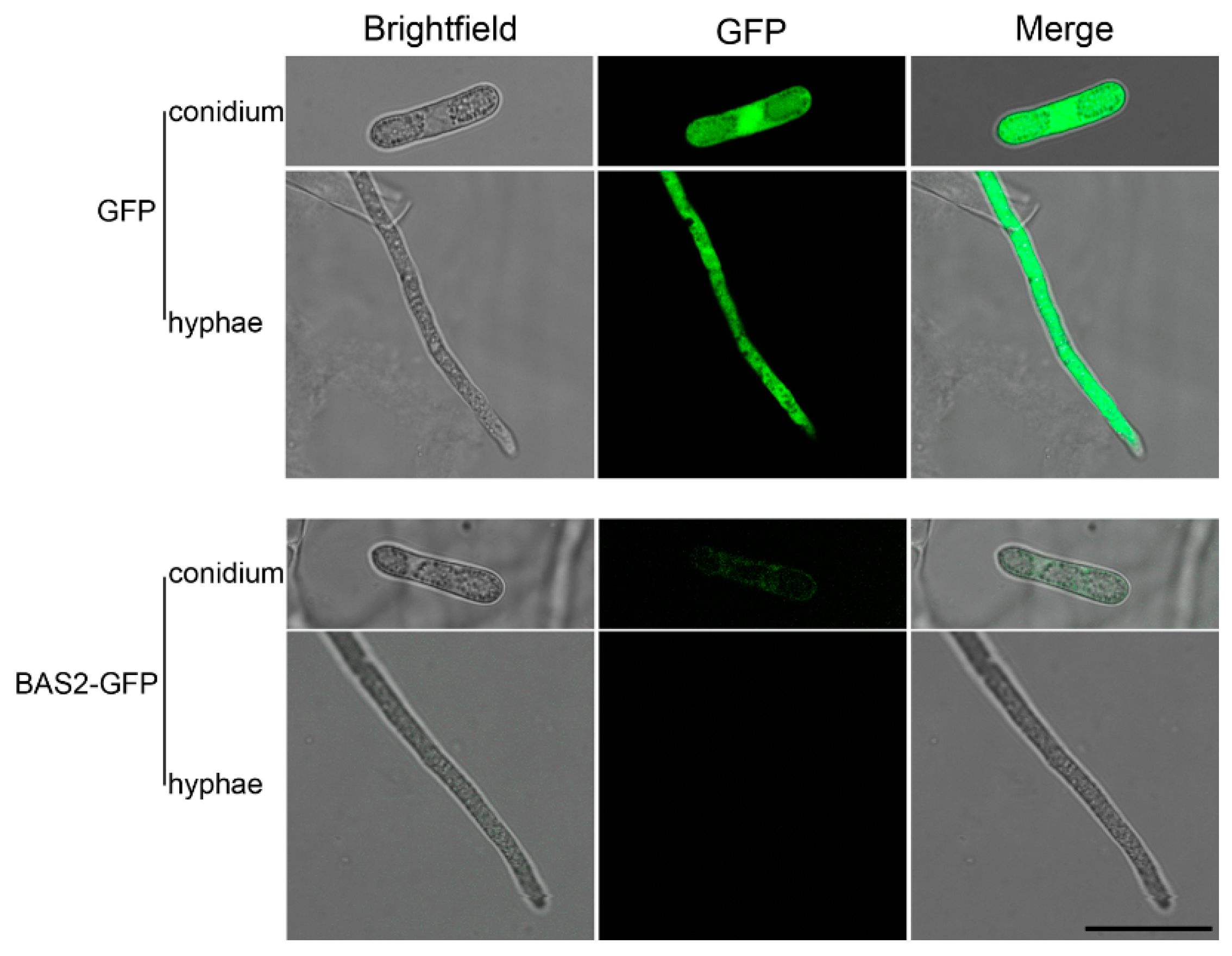
2.4. Subcellular Localization of BAS2-GFP in C. gloeosporioides
2.5. Subcellular Localization of BAS2-GFP in Mesophyll Cell Protoplasts of H. brasiliensis
2.6. BAS2 Is Involved in Extracellular Protein Secretion
3. Discussion
4. Materials and Methods
4.1. Fungal Strains and Culture Conditions
4.2. Construction of the Knockout Vector of BAS2
4.3. Transformation of C. gloeosporioides, PCR Diagnosis, and Single Conidia Purification
4.4. Growth and Conidiation Assay
4.5. Pathogenicity Assay
4.6. Generation of BAS2-GFP Fusion Expressing Mutant
4.7. Construction of BAS2-GFP Fusion Expressing Vector and Transient Expression in Mesophyll Cell Protoplasts of H. brasiliensis
4.8. Extracellular Proteomic Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Koeck, M.; Hardham, A.R.; Dodds, P.N. The role of effectors of biotrophic and hemibiotrophic fungi in infection. Cell. Microbiol. 2011, 13, 1849–1857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Göhre, V.; Robatzek, S. Breaking the barriers: Microbial effector molecules subvert plant immunity. Annu. Rev. Phytopathol. 2008, 46, 189–215. [Google Scholar] [CrossRef] [PubMed]
- Rafiqi, M.; Ellis, J.G.; Ludowici, V.A.; Hardham, A.R.; Dodds, P.N. Challenges and progress towards understanding the role of effectors in plant-fungal interactions. Curr. Opin. Plant Biol. 2012, 15, 477–482. [Google Scholar] [CrossRef] [PubMed]
- O’Connell, R.J.; Thon, M.R.; Hacquard, S.; Amyotte, S.G.; Kleemann, J.; Torres, M.F.; Damm, U.; Buiate, E.A.; Epstein, L.; Alkan, N.; et al. Lifestyle transitions in plant pathogenic Colletotrichum fungi deciphered by genome and transcriptome analyses. Nat. Genet. 2012, 44, 1060–1065. [Google Scholar] [CrossRef] [PubMed]
- Stephenson, S.A.; Hatfield, J.; Rusu, A.G.; Maclean, D.J.; Manners, J.M. CgDN3: An essential pathogenicity gene of Colletotrichum gloeosporioides necessary to avert a hypersensitive-like response in the host Stylosanthes guianensis. Mol. Plant Microbe Interact. 2000, 13, 929–941. [Google Scholar] [CrossRef] [PubMed]
- Takahara, H.; Hacquard, S.; Kombrink, A.; Hughes, H.B.; Halder, V.; Robin, G.P.; Hiruma, K.; Neumann, U.; Shinya, T.; Kombrink, E.; et al. Colletotrichum higginsianum extracellular lysm proteins play dual roles in appressorial function and suppression of chitin-triggered plant immunity. New Phytol. 2016, 211, 1323–1337. [Google Scholar] [CrossRef] [PubMed]
- Sanzmartín, J.M.; Pachecoarjona, J.R.; Bellorico, V.; Vargas, W.A.; Monod, M.; Díazmínguez, J.M.; Thon, M.R.; Sukno, S.A. A highly conserved metalloprotease effector enhances virulence in the maize anthracnose fungus Colletotrichum graminicola. Mol. Plant Pathol. 2015, 17, 1048–1062. [Google Scholar] [CrossRef] [PubMed]
- Bhadauria, V.; Banniza, S.; Vandenberg, A.; Selvaraj, G.; Wei, Y. Overexpression of a novel biotrophy-specific Colletotrichum truncatum effector, CtNUDIX, in hemibiotrophic fungal phytopathogens causes incompatibility with their host plants. Eukaryot. Cell 2013, 12, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Kleemann, J.; Rincon-Rivera, L.J.; Takahara, H.; Neumann, U.; Ver Loren van Themaat, E.H.; van der Does, C.; Hacquard, S.; Stüber, K.; Will, I.; Schmalenbach, W.; et al. Sequential delivery of host-Induced virulence effectors by appressoria and intracellular hyphae of the phytopathogen Colletotrichum higginsianum. PLoS Pathog. 2012, 8, e1002643. [Google Scholar] [CrossRef]
- Mosquera, G.; Giraldo, M.C.; Khang, C.H.; Coughlan, S.; Valent, B. Interaction transcriptome analysis identifies Magnaporthe oryzae BAS1-4 as biotrophy-associated secreted proteins in rice blast disease. Plant Cell 2009, 21, 1273–1290. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; He, C.; Luo, H. Pathogenicity analysis and construction of CgBASP2 knockout mutant of Colletotrichum gloeosporioides infecting Hevea brasiliensis. J. Trop. Biol. 2015, 6, 41–46. (In Chinese) [Google Scholar]
- Schumacher, J. Tools for Botrytis cinerea: New expression vectors make the gray mold fungus more accessible to cell biology approaches. Fungal Genet. Biol. 2012, 49, 483–497. [Google Scholar] [CrossRef] [PubMed]
- Khang, C.H.; Berruyer, R.; Giraldo, M.C.; Kankanala, P.; Park, S.Y.; Czymmek, K.; Kang, S.; Valent, B. Translocation of Magnaporthe oryzae effectors into rice cells and their subsequent cell-to-cell movement. Plant Cell 2010, 22, 1388–1403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xin, X.F.; Nomura, K.; Ding, X.; Chen, X.; Wang, K.; Aung, K.; Uribe, F.; Rosa, B.; Yao, J.; Chen, J.; et al. Pseudomonas syringae effector avirulence protein E localizes to the host plasma membrane and down-regulates the expression of the nonrace-specific disease resistance1/harpin-induced1-like13 gene required for antibacterial immunity in Arabidopsis. Plant Physiol. 2015, 169, 793–802. [Google Scholar] [CrossRef] [PubMed]
- Hewezi, T.; Juvale, P.S.; Piya, S.; Maier, T.R.; Rambani, A.; Rice, J.H.; Mitchum, M.G.; Davis, E.L.; Hussey, R.S.; Baum, T.J. The cyst nematode effector protein 10a07 targets and recruits host posttranslational machinery to mediate its nuclear trafficking and to promote parasitism in Arabidopsis. Plant Cell 2015, 27, 891–907. [Google Scholar] [CrossRef] [PubMed]
- Marshall, R.; Kombrink, A.; Motteram, J.; Lozareyes, E.; Lucas, J.; Hammond-Kosack, K.E.; Thomma, B.P.; Rudd, J.J. Analysis of two in planta expressed LysM effector homologs from the fungus Mycosphaerella graminicola reveals novel functional properties and varying contributions to virulence on wheat. Plant Physiol. 2011, 156, 756–769. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; An, B.; Hou, X.; Guo, Y.; Luo, H.; He, C. Dicer-like proteins regulate the growth, conidiation, and pathogenicity of Colletotrichum gloeosporioides from Hevea brasiliensis. Front. Microbiol. 2018, 8, 2621. [Google Scholar] [CrossRef] [PubMed]
- Reineke, G.; Heinze, B.; Schirawski, J.; Buettner, H.; Kahmann, R.; Basse, C.W. Indole-3-acetic acid (IAA) biosynthesis in the smut fungus Ustilago maydis, and its relevance for increased IAA levels in infected tissue and host tumour formation. Mol. Plant Pathol. 2008, 9, 339–355. [Google Scholar] [CrossRef] [PubMed]
- Yin, C.; Park, J.J.; Gang, D.R.; Hulbert, S.H. Characterization of a tryptophan 2-monooxygenase gene from Puccinia graminis f. sp. tritici involved in auxin biosynthesis and rust pathogenicity. Mol. Plant Microbe Interact. 2014, 27, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Chumley, F.G.; Valent, B. Genetic analysis of melanin-deficient, nonpathogenic mutants of Magnaporthe grisea. Mol. Plant Microbe Interact. 1990, 3, 135–143. [Google Scholar] [CrossRef]
- Kawamura, C.; Moriwaki, J.; Kimura, N.; Fujita, Y.; Fuji, S.; Hirano, T.; Koizumi, S.; Tsuge, T. The melanin biosynthesis genes of Alternaria alternata can restore pathogenicity of the melanin-deficient mutants of Magnaporthe grisea. Mol. Plant Microbe Interact. 1997, 10, 446–453. [Google Scholar] [CrossRef] [PubMed]
- Veneault-Fourrey, C.; Barooah, M.; Egan, M.; Wakley, G.; Talbot, N.J. Autophagic fungal cell death is necessary for infection by the rice blast fungus. Science 2006, 312, 580–583. [Google Scholar] [CrossRef] [PubMed]
- Talbot, N.J.; Ebbole, D.J.; Hamer, J.E. Identification and characterization of MPG1, a gene involved in pathogenicity from the rice blast fungus Magnaporthe grisea. Plant Cell 1993, 5, 1575–1590. [Google Scholar] [CrossRef] [PubMed]
- Lorang, J.M.; Tuori, R.P.; Martinez, J.P.; Sawyer, T.L.; Redman, R.S.; Rollins, J.A.; Wolpert, T.J.; Johnson, K.B.; Rodriguez, R.J.; Dickman, M.B.; et al. Green fluorescent protein is lighting up fungal biology. Appl. Environ. Microbiol. 2001, 67, 1987–1994. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, L.; He, C.; Luo, H. An efficient transient mesophyll protoplast system for investigation of the innate immunity responses in the rubber tree (Hevea brasiliensis). Plant Cell Tiss. Org. 2016, 126, 281–290. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spot | Protein Function | Accession Number | NP | SC (%) | WT vs. ∆BAS2 | Biological Function |
|---|---|---|---|---|---|---|
| Cell structure | ||||||
| D1 | Actin | ELA34037.1 | 2 | 6 | ∞ | Cytoskeleton |
| Redox | ||||||
| D6 | Peroxidase | ELA30823.1 | 14 | 37 | 38.56 | Redox |
| D8 | Peroxidase | ELA30823.1 | 11 | 30 | ∞ | Redox |
| D9 | Peroxidase | ELA30823.1 | 12 | 34 | ∞ | Redox |
| Metabolism | ||||||
| D7 | Endonuclease/exonuclease/phosphatase family protein | ELA30823.1 | 13 | 35 | ∞ | Metabolism |
| D11 | Choline dehydrogenase | ELA24295.1 | 10 | 17 | ∞ | Choline metabolism |
| D12 | Choline dehydrogenase | ELA24295.1 | 9 | 16 | ∞ | Choline metabolism |
| D13 | Flavine adenine dinucleotide-dependent oxygenase | ELA25647.1 | 10 | 22 | 33.04 | Metabolism |
| D15 | Flavine adenine dinucleotide-binding dehydrogenase | ELA24979.1 | 3 | 5 | 13.20 | Metabolism |
| D21 | Enolase | ELA34631.1 | 15 | 30 | 14.04 | Glycolytic pathways |
| Secondary metabolism | ||||||
| D10 | Indoleacetamide hydrolase (IaaH) | ELA36057.1 | 8 | 14 | ∞ | IAA synthesis |
| D14 | Versicolorin b synthase | ELA30672.1 | 13 | 19 | 26.33 | Aflatoxin synthesis |
| D16 | Versicolorin b synthase | ELA30672.1 | 16 | 25 | 45.11 | Aflatoxin synthesis |
| D19 | Tyrosinase central domain containing protein | ELA33276.1 | 4 | 8 | ∞ | Melanin synthesis |
| Cell wall degradation | ||||||
| D17 | Endo-β-1,6-glucanase | ELA27535.1 | 11 | 21 | 1.70 | Degradation of chitin |
| D18 | Endo-β-1,6-glucanase | ELA27535.1 | 8 | 19 | ∞ | Degradation of chitin |
| D20 | Exopolygalacturonase | ELA28364.1 | 4 | 9 | 2.29 | Degradation of pectin |
| Unknown | ||||||
| D2 | Not identified | – | – | – | ∞ | – |
| D3 | Not identified | – | – | – | 14.69 | – |
| D4 | Not identified | – | – | – | ∞ | – |
| D5 | Not identified | – | – | – | ∞ | – |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
An, B.; Wang, W.; Guo, Y.; Wang, Q.; Luo, H.; He, C. BAS2 Is Required for Conidiation and Pathogenicity of Colletotrichum gloeosporioides from Hevea brasiliensis. Int. J. Mol. Sci. 2018, 19, 1860. https://doi.org/10.3390/ijms19071860
An B, Wang W, Guo Y, Wang Q, Luo H, He C. BAS2 Is Required for Conidiation and Pathogenicity of Colletotrichum gloeosporioides from Hevea brasiliensis. International Journal of Molecular Sciences. 2018; 19(7):1860. https://doi.org/10.3390/ijms19071860
Chicago/Turabian StyleAn, Bang, Wenfeng Wang, Yunfeng Guo, Qiannan Wang, Hongli Luo, and Chaozu He. 2018. "BAS2 Is Required for Conidiation and Pathogenicity of Colletotrichum gloeosporioides from Hevea brasiliensis" International Journal of Molecular Sciences 19, no. 7: 1860. https://doi.org/10.3390/ijms19071860
APA StyleAn, B., Wang, W., Guo, Y., Wang, Q., Luo, H., & He, C. (2018). BAS2 Is Required for Conidiation and Pathogenicity of Colletotrichum gloeosporioides from Hevea brasiliensis. International Journal of Molecular Sciences, 19(7), 1860. https://doi.org/10.3390/ijms19071860