Molecular Mechanisms for Regulating Postnatal Ductus Arteriosus Closure


1
Graduate Institute of Medicine, College of Medicine, Kaohsiung Medical University, Kaohsiung 807, Taiwan
2
Department of Pediatrics, St. Joseph Hospital, Kaohsiung 807, Taiwan
3
Department of Pharmacology, College of Medicine, Kaohsiung Medical University, Kaohsiung 807, Taiwan
4
Department of Medical Research, Kaohsiung Medical University Hospital, Kaohsiung 807, Taiwan
5
Department of Marine Biotechnology and Resources, National Sun Yat-sen University, Kaohsiung 804, Taiwan
6
Department of Pediatrics, Kaohsiung Medical University Hospital, Kaohsiung Medical University, Kaohsiung 807, Taiwan
7
Department of Pediatrics, Faculty of Medicine, College of Medicine, Kaohsiung Medical University, Kaohsiung 807, Taiwan
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2018, 19(7), 1861; https://doi.org/10.3390/ijms19071861
Submission received: 4 May 2018
/
Revised: 30 May 2018
/
Accepted: 20 June 2018
/
Published: 25 June 2018
(This article belongs to the Section Molecular Pathology, Diagnostics, and Therapeutics)
Abstract
:The ductus arteriosus (DA) connects the main pulmonary artery and the aorta in fetal circulation and closes spontaneously within days after birth in normal infants. Abnormal patent DA (PDA) causes morbidities and mortality, especially in preterm infants. Closure of the DA is a complex interactive process involving two events: functional and anatomic closure. Functional closure by smooth muscle contraction was achieved through the regulatory factors of vaso-reactivity. These factors include oxygen sensing system, glutamate, osmolality, prostaglandin E2, nitric oxide, and carbon monoxide. Anatomic closure by vascular remodeling involved several vascular components including endothelium, extracellular matrix, smooth muscle cells, and intraluminal blood cells. Despite advances in understanding of PDA pathogenesis, the molecular mechanism for regulation of DA closure is complex and not fully understood. In this article we review recent evidence regarding the molecular mechanisms of DA closure.
1. Introduction
The ductus arteriosus (DA) is a vital vessel that connects pulmonary circulation and systemic circulation in the fetus. Closure of the DA is mostly completed within three days of life in healthy term newborns. The incidence of an isolated patent DA (PDA) ranges from 3 to 8 per 10,000 live births among term infants [1] and is estimated at up to 30 percent in very low birth weight infants (birth weight below 1500 g) [2]. The PDA is a hemodynamic burden in preterm infants and is also the leading cause of mortality and morbidity among these infants [3]. However, maintaining the patency of the DA is life-saving in infants with ductus-dependent congenital heart diseases. Therefore, proper manipulation of DA patency is essential in neonatal intensive care and investigation of its molecular mechanisms is an important field in vascular biology and pediatrics.
Generally, DA closure involves two phases: functional and anatomical closure. Functional closure occurring within hours after birth is caused by DA constriction and the following anatomical closure is mediated mainly by vascular remodeling. After birth, increased oxygen tension and declined prostaglandin E2 (PGE2) are two major factors for DA constriction [4]. Subsequent DA remodeling is associated with several histological changes: internal elastic lamina (IEL) disruption, lifting and ingrowth of endothelial cells (ECs), subendothelial edema due to deposition of extracellular matrix (ECM), migration and proliferation of the SMCs into the subendothelial space [5,6,7]. These histological changes result in intimal cushion for permanent closure of the DA. In this article, we review both mechanisms of functional and anatomical closure of the DA.
2. Functional Closure
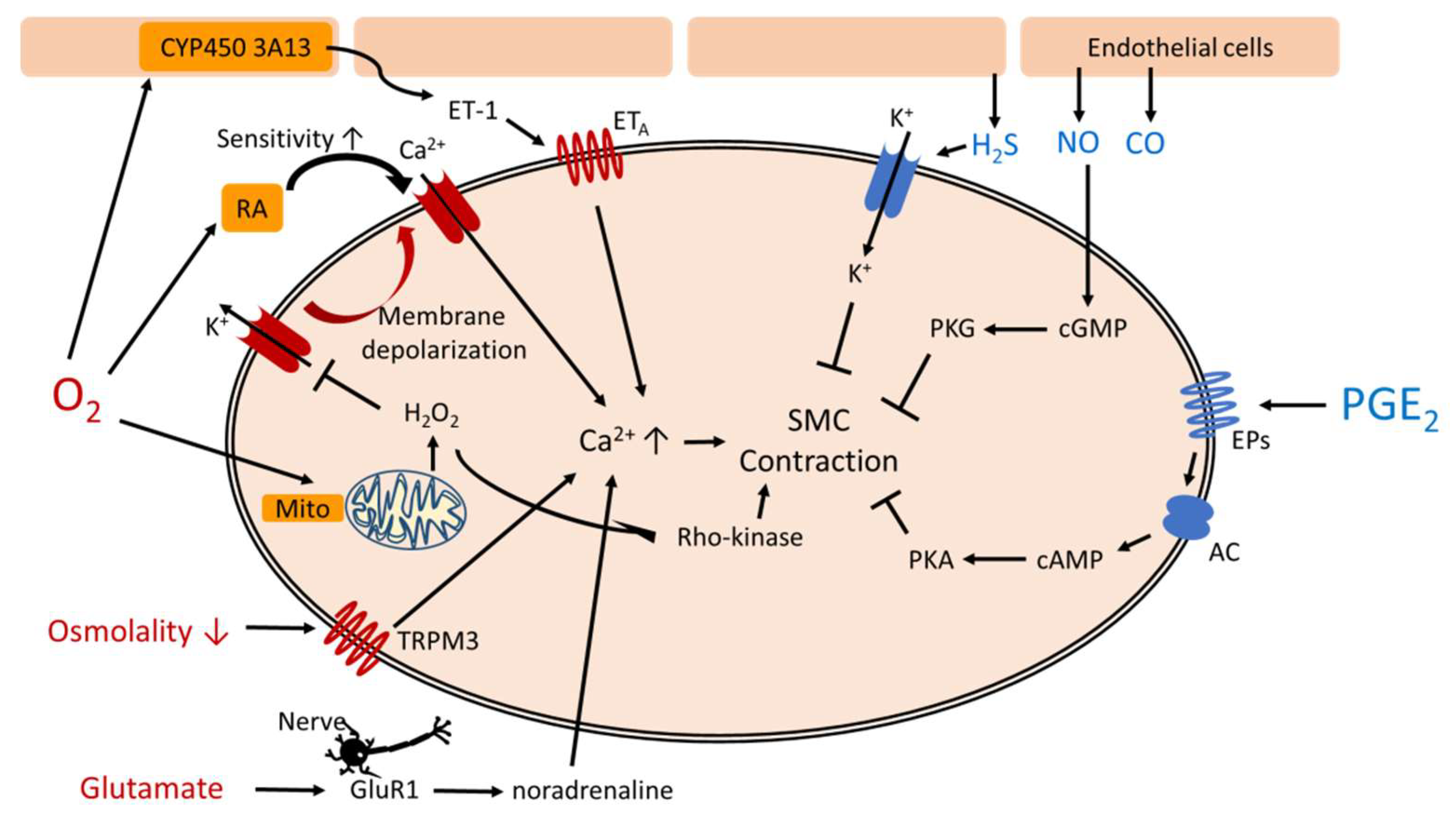
During fetal life, intrauterine hypoxia works synergistically with high circulating PGE2 to maintain DA patency. After birth, the DA constricts in response to elevated oxygen tension and declined PGE2 level [8,9]. However, the preterm infants often have hypoxic events such as respiratory distress syndrome or bronchopulmonary dysplasia, resulting higher incidence of PDA. There are many factors controlling the DA vascular tone (Table 1). Figure 1 shows complex pathways mediating functional closure of the DA.
2.1. Vasoconstriction
2.1.1. Oxygen Pathways
Several mechanisms were found recently to underlie the vasoconstrictive response of high oxygen tension in DA. Archer et al. demonstrated that DA smooth muscle cells (DASMCs) can sense oxygen via dynamic mitochondrial network [35]. They showed O2-induced DA constriction was initiated by inhibition of a voltage-gated potassium channel, which caused membrane depolarization, activation of L-type calcium channels and increment in intracellular calcium (Ca2+) [14]. H2O2 produced by mitochondrial electron transport chain complex served as an oxygen mediator to inhibit potassium channels [15]. Through mitochondrial fission, elevated oxygen tension increased reactive oxygen species (ROS) levels and mitochondrial complex I activity [16]. In brief, oxygen-induced increment of ROS (e.g., H2O2) inhibits potassium channel and subsequent membrane depolarization causes Ca2+ influx due to opening of calcium, inducing DASMCs contraction.
Recent evidence demonstrates that the role of Rho-kinase pathway to sustain DA constriction via the mitochondrial system. The oxygen-induced increment of mitochondrial ROS activates the Rho-kinase pathway and induces RhoB and Rho-associated protein kinase-1 expression in human and rabbit DA [17]. The Rho-kinase pathway promotes phosphorylation of myosin phosphatase targeting protein and this phosphorylation inhibits myosin light chain phosphatase, thereby increasing the phosphorylation and activity of the myosin light chain, which leads to DASMC contraction. The activation of the Rho-kinase pathway thus induces calcium sensitization, which sustains DA constriction through a positive feedback mechanism.
There is some evidence suggesting that cytochrome P450 (CYP450) and endothelin-1 (ET-1) also jointly participate in the mechanisms underlying oxygen-induced DA constriction. The level of ET-1 increased in response to oxygen and acted as DA constrictor via ETA receptor [36,37,38]. The CYP450-based mechanism mediates the constrictive response of the DA to oxygen, possibly by stimulating the release and synthesis of ET-1 [21,22].
Another possible oxygen sensing factor is retinoic acid, a metabolite of vitamin A. Wu et al. found that fetal rats born from maternally vitamin A-treated group had better DA contraction induced by oxygen but not by KCl [24]. Yokoyama et al. showed that maternally administered vitamin A significantly upregulated the expression levels of α1G subunit of voltage-dependent calcium channel, which is activated by oxygen-induced inhibition of potassium channel [25].
2.1.2. Pathways Unrelated to Oxygen
Glutamate, an amino acid, has been recently found to promote DA contraction through glutamate inotropic receptor subunit 1 (GluR1)-mediated noradrenaline production. Fujita et al. showed that glutamate increased noradrenaline production in the rat DA and subsequent glutamate-induced DA contraction was attenuated by the GluR receptor antagonist or the adrenergic receptor α1 blocker [28]. This evidence suggests that nutritional adjustment with supply amino acid may have therapeutic implications in newborn infants with PDA.
Recent evidence suggests that hypo-osmolality has a role in mediating DA constriction. This mechanism is mediated by regulating Ca2+, potentially through the transient receptor potential melastatin 3 (TRPM3) pathway. Aoki et al. found that in rats that hypo-osmotic sensor TRPM3 was more upregulated in the DA than in the aorta [31]. They also demonstrated that rats experienced transient hypo-osmolality after birth, which contributed to rat DA constriction.
There are other agents circulating in the blood conveying vasoconstrictive effects on DA. For example, bradykinin shows biphasic effect at rabbit DA through two different receptors, BK-1 and BK-2 receptors. As bradykinin concentration increases, DA has predominantly constrictive responses through BK-1 receptor [32]. Corticosteroids also induce DA constriction, in combination with indomethacin, probably through attenuating the sensitivity of the DA to PGE2 [33,34].
2.2. Vasodilation
2.2.1. PGE2
In current clinical settings, PGE1 administration is the only medical treatment for maintaining DA patency in neonates with DA-dependent congenital heart diseases [39]. PGE2 is produced in both the placenta and the DA in fetal circulation. It maintains DA patency through various PGE receptors (EP2, EP3, and EP4) [10,11,12]. Activation of PGE2 receptors increases intracellular cyclic AMP (cAMP) via adenylyl cyclases and the increased cAMP level inhibits myosin light chain kinase, subsequently dilating DA [11,13]. After birth, the PGE2 level declines due to pulmonary catabolism of PGE2 and the removal of the placenta [9,10]. PGE2 receptors (EP3 and EP4) also decrease in numbers after birth [11]. Postnatal decline of PGE2 signaling has been postulated to be the fundamental mechanism for DA closure [40]. Moreover, elevated oxygen tension can downregulate DA sensitivity to PGE2, thus attenuating the postnatal vasodilating response [41,42].
2.2.2. Vasodilating Factors Unrelated to PGE
Nitric oxide (NO) has been shown to be a vasodilator in DA. NO is produced by endothelial nitric oxide synthase (eNOS) in ECs and then diffuses into adjacent SMCs to bind with soluble guanylyl cyclase (sGC). The activated sGC causes production of cyclic guanosine monophosphate (cGMP), which decreases Ca2+. The lowering Ca2+ relaxes the SMCs and promotes vasodilation [43]. In DA, NO can be synthesized by eNOS in the endothelium of DA lumen and vasa vasorum [18]. Indeed, combined use of indomethacin and NOS inhibitor was shown to have more potent constricting efficacy in DA than indomethacin alone in premature baboons [19]. Intriguingly, indomethacin was also found to promote vasodilatory function of NO in mouse DA [20]. These paradoxical effects could explain the failure of indomethacin therapy in about 30% of premature neonates in clinical practice [44].
Similar with NO, another vasodilating pathway mediated through cGMP is the family of natriuretic peptides. They are cardiac-producing peptides that can dilate vessels through the particulate GC-cGMP pathway [45]. Atrial natriuretic peptide has been shown to dilate rat DA in vivo [23]. Our recent study demonstrated that higher B-type natriuretic peptides (BNP) convey anti-remodeling effects in the pulmonary artery SMCs [46]. Furthermore, in the setting of neonatal intensive care unit, plasma levels of BNP are associated with poor response to indomethacin treatment in preterm infants with PDA [44]. Taken together, this evidence suggests the role of BNP in DA control, but this warrants further investigation.
In addition to NO, other gases including carbon monoxide (CO) and hydrogen sulfide (H2S) can also dilate DA. The CO-forming enzyme, heme oxygenase-1 and -2, identified in DA tissue, was shown to produce CO in the DASMCs [26]. CO dilates DA due to inhibition of a CYP450-based monooxygenase reaction conditioning the formation of the ET-1 [27]. Recently, Baragatti et al. demonstrated H2S synthetic enzyme in the mice DA and confirmed the H2S-induced vasodilatory effects of DA [29]. Interestingly, H2S was found to have biphasic effects, inducing vasoconstriction at lower concentrations while causing vasodilation at higher concentrations [30]. However, in the chicken DA, the vasodilatory effect was not shown [47]. The inter-species differences and dose-specific vasoreactive mechanisms of H2S in DA are not fully understood and warrant further investigations.
3. Anatomical Closure
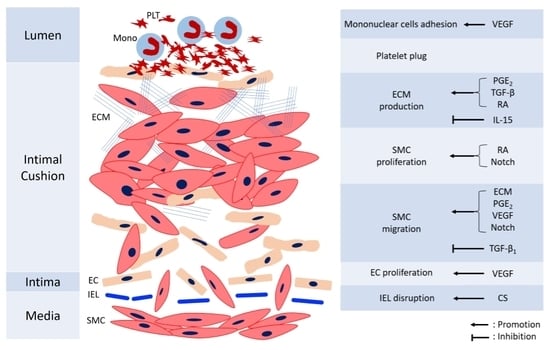
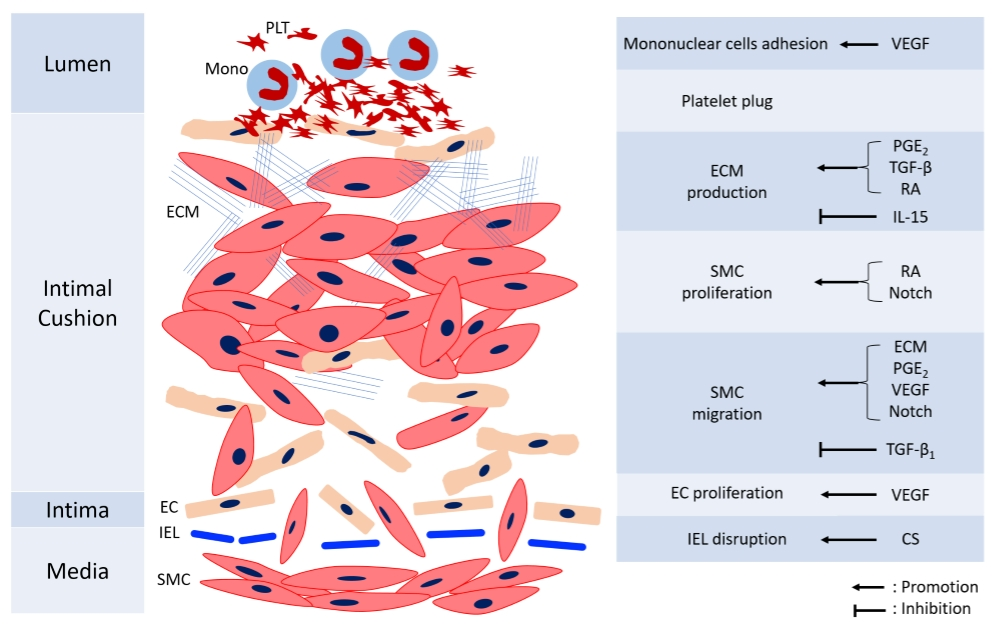
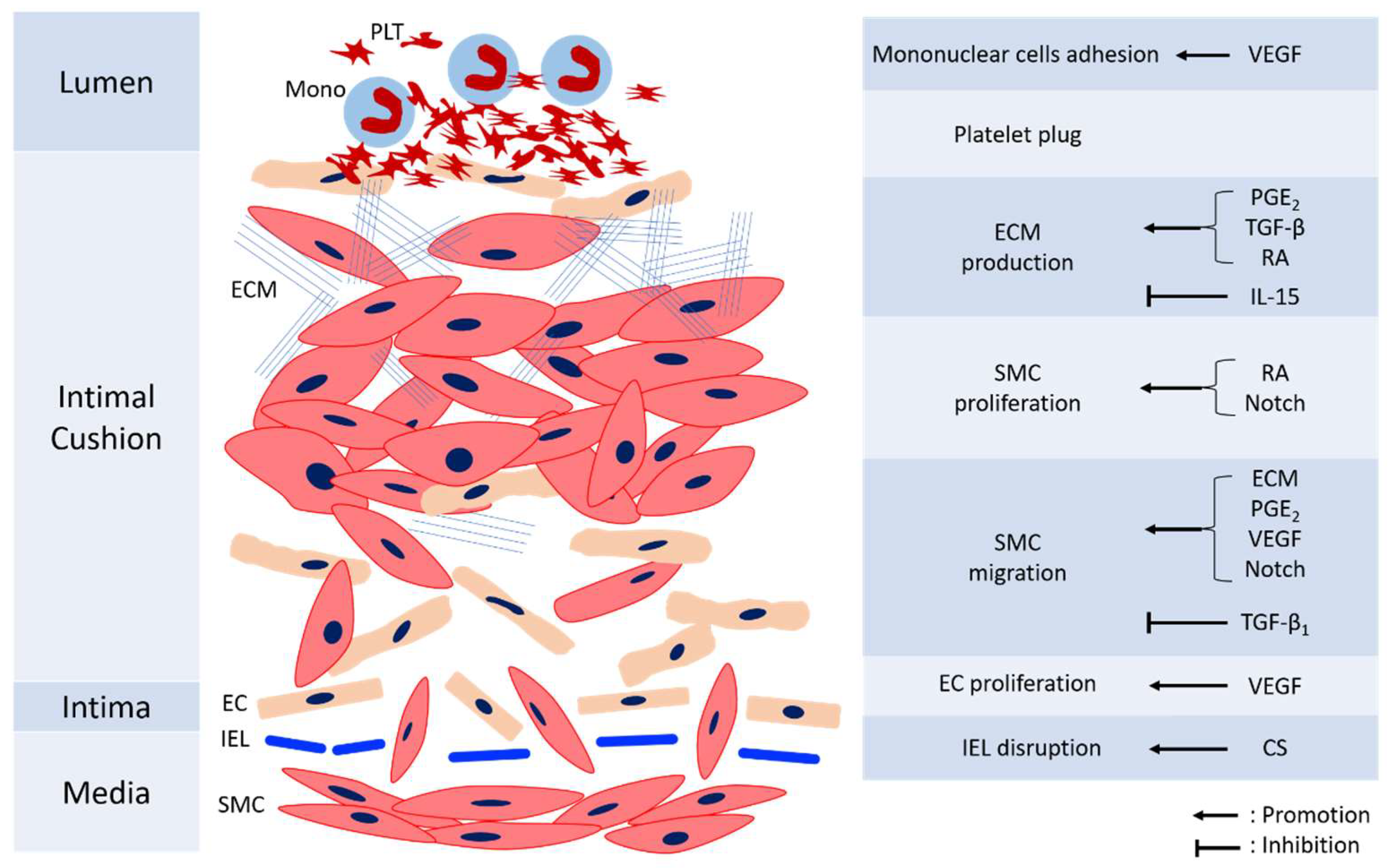
Remodeling of DA is essential to permanent anatomical closure to prevent re-opening. The process is complex and not fully understood, with several mechanisms including intimal cushion formation, SMC migration and proliferation, ECM production, EC proliferation, and blood cell interaction. These steps interact with each other and construct an orchestrated process. Figure 2 shows the mechanisms involved in the anatomic closure of the DA. The detailed references of mechanisms mediating various cells for anatomical closure are shown in Table 2.
3.1. Factors Regulating SMC Proliferation and Migration
Similar to the vascular remodeling of other diseases such as atherosclerosis and pulmonary hypertension, SMC migration and proliferation play important roles in DA remodeling. DA remodeling starts with separation of the EC from the IEL resulting in creation of subendothelial space for migration and proliferation of undifferentiated SMCs [64]. These factors include PGE2, retinoic acid, transforming growth factor-β1 (TGF-β1), and Notch signaling.
3.1.1. PGE2
PGE2 induces DASMC migration through exchange protein activated by cAMP (Epac) pathway. Epac signaling is also regulated by cAMP but is distinctly different from the PKA pathway [65]. Serial activations of the PGE2-EP4-cAMP-Epac signaling pathway induce DASMC migration without changing SMC proliferation and hyaluronan production [48]. PGE2 has paradoxical effects on the functional and anatomical aspects of DA closure, that is, vasodilatation and remodeling.
3.1.2. Retinoic Acid
Retinoic acid also participates in vascular remodeling via promoting SMC and ECM proliferation. Wu et al. showed that retinoic acid stimulated the growth of DASMCs by the stimulation of proliferating cell nuclear antigen expression and decreased apoptosis [54]. Yokoyama et al. demonstrated that maternally administrated vitamin A increased the production of fibronectin and hyaluronic acid, promoting intimal thickening in the DA at preterm rats [55]. Taken together, retinoic acid mediates both vasoconstriction and vascular remodeling.
3.1.3. TGF-β1
TGF-β1 anchors the SMC’s cytoskeleton to the ECM, making SMCs more adherent to ECM and less migrative. TGF-β1 increases focal plaque formation in DASMCs by increasing adhesion of the integrin with the cytoskeleton, possibly maintaining the tension necessary to sustain DA contracture during remodeling [49,66].
3.1.4. Notch Signaling
The Notch system is highly expressed in human vasculature and regulates cell behavior, including proliferation, migration, and angiogenesis [67]. Recent reports suggest that it has a role in DA remodeling. Baeten et al. showed that the loss of Notch receptors in DASMCs is associated with downregulated contractile SMC gene expression, contributing to the formation of PDA [68]. Krebs et al. demonstrated that Notch signaling is required for contractile smooth muscle cell differentiation and DA closure in mice [69]. Our recent study suggests a role of Notch signaling in the proliferation and migration of DASMCs [50]. Specifically, we found that γ-secretase inhibitor DAPT, a Notch signaling inhibitor, could prevent the angiotensin II-induced proliferation and migration of DASMCs. These effects are potentially mediated by attenuated calcium overload, reduced ROS production, and deactivations of ERK1/2, JNK, and Akt signal transduction through the Notch3-HES1/2/5 pathway.
3.2. Extracellular Matrix (ECM)
It has been found in many vascular proliferative diseases that the ECM can promote SMC migration and proliferation [70]. ECM consists of hyaluronan, fibronectin, chondroitin sulfate, and elastin, and each of them has a different role in DA remodeling.
3.2.1. Hyaluronan
Hyaluronan is important during DA remodeling due to its effects on promoting DASMC migration. It is regulated by other factors, including TGF-β, PGE2, and interleukin-15 (IL-15). TGF-β is produced in ECs and can promote synthesis of hyaluronan and chondroitin sulfate in DA [56]. PGE2 regulates hyaluronan production via the EP4-cAMP-PKA signaling pathway [71]. PGE2-mediated activation of EP4 leads to increased cAMP production and PKA signaling, leading to increased hyaluronan synthase activity in DASMCs. Chronic EP4 can also stimulate hyaluronan production [51]. IL-15 is predominantly expressed in the IEL in rat DA and inhibits hyaluronan production [57].
3.2.2. Fibronectin
Fibronectin is secreted by DASMCs and can promote SMC migration into the subendothelium in the process of intimal cushion formation. DA patency can be maintained by inhibiting fibronectin-dependent intimal cushion formation [52]. In addition, maternally administrated vitamin A increased fibronectin production in the DA of neonatal rats [55].
3.2.3. Chondroitin Sulfate
Chondroitin sulfate promotes DA remodeling through supporting the stability of hyaluronan and impairing the assembly of elastin fibers. Chondroitin sulfate causes 67-kD elastin binding protein to be released from the SMC surface, impairing elastin assembly [62]. Chondroitin sulfate promotes SMC migration indirectly by promoting detachment of SMCs from elastin and upregulates synthesis of fibronectin, which facilitates migration of SMCs through IEL [72].
3.2.4. Elastin
Elastin confers elasticity of blood vessels and contributes to maintaining PA patency. The loss of elastin-binding protein and the production of elastin peptides can enhance DASMC migration [73]. Intriguingly, the production of elastin is regulated by PGE2 and oxygen. A recent study indicates that PGE2 can inhibit elastogenesis via the EP4 receptor and such attenuated elastin formation promotes vascular collapse and subsequent DA closure after birth [58]. In addition, oxygenation reduces elastin secretion in DASMCs [59]. Thus, from the perspective of elastogenesis, both PGE2 and oxygen play a role in anti-remodeling.
3.3. Factors Affecting Endothelial Cells (ECs)
In the process of DA remodeling, ECs separates from IEL to create a subendothelial space for the further migration of SMCs and ECs. The migration of ECs is influenced by integrins and VEGF.
Integrins are transmembrane receptors that create traction with surrounding ECM and provide signals for cytoskeleton rearrangement and initiation of cytoplasmic flow. Both ECs and SMCs experience an increase in their integrin supply during intimal cushion formation. Indeed, preterm infants with PDA were found to have downregulation of integrin expression [74]. Thus, integrin may participate in the interaction between ECM and ECs during DA remodeling, as it does between ECM and SMCs [75].
Vascular endothelial cell growth factor (VEGF) is a hypoxia-induced growth factor and can stimulate EC proliferation and migration. VEGF regulates DA remodeling by stimulating EC proliferation and SMC migration and is induced by tissue hypoxia [53]. Clyman et al. proposed an important role of VEGF in DA remodeling [60]. They demonstrated that initial functional vasoconstriction causes a loss of luminal blood flow, producing a hypoxic zone in the DA muscle media layer. They also found that distribution of VEGF is closely associated with the area of hypoxia in the constricting DA. In addition, anti-VEGF antibody was found to inhibit mononuclear cells from adhering to the DA lumen and decreases intimal cushion expansion [61].
3.4. Blood Cells’ Interaction
Circulating blood cells adhering to DA lumen have essential roles during DA remodeling. Mononuclear cells activated by inflammatory responses-induced vascular wall ischemia have recently been postulated as necessary for DA remodeling. After DA constriction, VLA4+ mononuclear cells (monocytes and macrophages) adhere to the ductus lumen via vascular cell adhesion molecule-1 expressed in the luminal cells [76]. The degree of mononuclear cells adhesion is correlated with the extent of intimal cushion formation [61].
Platelets also have a central role in permanent DA closure. Echtler et al. demonstrated that, during DA constriction, ECs become detached and trigger the recruitment of platelets passing through the constricted DA [77]. The formation of a platelet plug seals the residual lumen of the constricted DA and facilitates luminal remodeling. Engur et al. reported that platelet-derived growth factor levels were lower in infants who had persistent PDA after birth [63]. Emerging evidence shows the relationships between thrombocytopenia and the failure of spontaneous closure of DA [78,79].
4. Pharmacological Agents for Management of DA Patency
Current medications for the management of DA patency mainly convey physiological effects on vascular tone by vasodilation or vasoconstriction, rather than a remodeling effect. Table 3 summarizes the currently used or experimental agents to close or open DA.
4.1. Agents for Closing the DA
Drugs for closing the DA involve inhibition of prostaglandin (PG) production. In current clinical settings, indomethacin or ibuprofen is administrated for closing the DA in preterm newborn with DA-induced heart failure. Indomethacin and ibuprofen inhibit cyclooxygenase-1 and cyclooxygenase-2, which convert arachidonic acid to PGG2 for further production of various PGs. Among the PGs, PGE2 is the most potent vasodilator to open DA [91]. Indomethacin and ibuprofen both inhibit PGE2 production and are effective in closing the PDA in preterm infants. Oral ibuprofen may be the preferred agent due to feasibility and fewer side effects [80]. Recently, acetaminophen has been found to achieve DA closure in preterm infants [82]. Acetaminophen reduces PG production, probably through affecting peroxidase segment of cyclooxygenase [83]. Although some studies showed that acetaminophen was as effective as ibuprofen in closing the PDA [84,85], conflicting results preclude the routine use of acetaminophen for closing the PDA so far [86]. Further clinical studies are needed to reveal the efficacy of acetaminophen in closing the PDA.
4.2. Agents for Opening the DA
Several agents are found to maintain DA patency through conveying vasodilatory effect in clinical practice or animal studies. Clinically, PGE1 (Alprostadil) is administered in infants with ductus-dependent congenital heart diseases to maintain DA patency. PGE1 binds to the EP4 receptor and then increases intracellular cAMP levels, which inhibit myosin light-chain kinase, resulting in the relaxation of the DA [81]. Milrinone, a phosphodiesterase 3 inhibitor, can dilate the rat DA through increasing cAMP levels [87]. Enalapril, an angiotensin-converting enzyme inhibitor, can also delay DA closure when given during caesarean delivery and can reopen the closed DA temporarily when given at 180 min of life in newborn rats [88]. A nonselective endothelin receptor antagonist (ERA), TAK-044, was found to inhibit DA construction in rats [89]. NO regulates the patency of the DA through the NO-cGMP pathway [90]. We have recently found that inhibition of the Notch pathway may convey anti-remodeling effects on DASMCs, suggesting its potential role in DA control [50]. Our ongoing study shows that BNP, an activator of PKG-cGMP, can prevent postnatal DA closure. Therefore, the cGMP pathway may be a potential research target in regulating DA patency.
5. Conclusions
DA closure consists of complex interactive processes involving vascular tone and vascular remodeling. The current clinical pharmacological strategy of regulating DA is based on the PG pathway and has some adverse effects and limitations. However, many other agents with vasodilatory or anti-remodeling effects through non-PG pathways have been shown with potential roles in maintaining DA patency, such as the NO, BNP, or Notch pathways. However, these agents require more animal or clinical studies to confirm their efficacy. Therefore, future research targeting a new pharmacological strategy of DA is essential in the fields of vascular biology and pediatrics.
Author Contributions
Y.-C.H. contributed to manuscript writing. J.-L.Y. contributed to editing of the tables, figures, and references. J.-H.H. contributed to study design and manuscript writing.
Acknowledgments
This study was supported in part by research grants from the Ministry of Science and Technology (MOST 106-2314-B-037-081-MY2 and MOST 106-2320-B-037-010-MY3).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Reller, M.D.; Strickland, M.J.; Riehle-Colarusso, T.; Mahle, W.T.; Correa, A. Prevalence of congenital heart defects in metropolitan Atlanta, 1998–2005. J. Pediatr. 2008, 153, 807–813. [Google Scholar] [CrossRef] [PubMed]
- Lemons, J.A.; Bauer, C.R.; Oh, W.; Korones, S.B.; Papile, L.A.; Stoll, B.J.; Verter, J.; Temprosa, M.; Wright, L.L.; Ehrenkranz, R.A.; et al. Very low birth weight outcomes of the National Institute of Child Health and Human Development neonatal research network, January 1995 through December 1996. NICHD Neonatal Research Network. Pediatrics 2001, 107, E1. [Google Scholar] [CrossRef] [PubMed]
- Noori, S.; McCoy, M.; Friedlich, P.; Bright, B.; Gottipati, V.; Seri, I.; Sekar, K. Failure of ductus arteriosus closure is associated with increased mortality in preterm infants. Pediatrics 2009, 123, e138–e144. [Google Scholar] [CrossRef] [PubMed]
- Heymann, M.A.; Rudolph, A.M. Control of the ductus arteriosus. Physiol. Rev. 1975, 55, 62–78. [Google Scholar] [CrossRef] [PubMed]
- Gittenberger-de Groot, A.C. Persistent ductus arteriosus: Most probably a primary congenital malformation. Br. Heart J. 1977, 39, 610–618. [Google Scholar] [CrossRef] [PubMed]
- Gittenberger-de Groot, A.C.; van Ertbruggen, I.; Moulaert, A.J.; Harinck, E. The ductus arteriosus in the preterm infant: Histologic and clinical observations. J. Pediatr. 1980, 96, 88–93. [Google Scholar] [CrossRef] [Green Version]
- De Reeder, E.; Girard, N.; Poelmann, R.; Van Munsteren, J.; Patterson, D.; Gittenberger-De Groot, A. Hyaluronic acid accumulation and endothelial cell detachment in intimal thickening of the vessel wall. The normal and genetically defective ductus arteriosus. Am. J. Pathol. 1988, 132, 574–585. [Google Scholar] [PubMed]
- Kovalčík, V. The response of the isolated ductus arteriosus to oxygen and anoxia. J. Physiol. 1963, 169, 185–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, M.Y.; Brown, D.M. Effect of dexamethasone or fetal lung 15-hydroxy-prostaglandin dehydrogenase: Possible mechanism for the prevention of patent ductus arteriosus by maternal dexamethasone therapy. Prostaglandins Leuko. Med. 1987, 27, 237–245. [Google Scholar] [CrossRef]
- Thorburn, G.D. The placenta, PGE2 and parturition. Early Hum. Dev. 1992, 29, 63–73. [Google Scholar] [CrossRef]
- Bouayad, A.; Kajino, H.; Waleh, N.; Fouron, J.C.; Andelfinger, G.; Varma, D.R.; Skoll, A.; Vazquez, A.; Gobeil, F., Jr.; Clyman, R.I.; et al. Characterization of PGE2 receptors in fetal and newborn lamb ductus arteriosus. Am. J. Physiol. Heart Circ. Physiol. 2001, 280, H2342–H2349. [Google Scholar] [CrossRef] [PubMed]
- Leonhardt, A.; Glaser, A.; Wegmann, M.; Schranz, D.; Seyberth, H.; Nusing, R. Expression of prostanoid receptors in human ductus arteriosus. Br. J. Pharmacol. 2003, 138, 655–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokoyama, U.; Minamisawa, S.; Katayama, A.; Tang, T.; Suzuki, S.; Iwatsubo, K.; Iwasaki, S.; Kurotani, R.; Okumura, S.; Sato, M.; et al. Differential regulation of vascular tone and remodeling via stimulation of type 2 and type 6 adenylyl cyclases in the ductus arteriosus. Circ. Res. 2010, 106, 1882–1892. [Google Scholar] [CrossRef] [PubMed]
- Tristani-Firouzi, M.; Reeve, H.L.; Tolarova, S.; Weir, E.K.; Archer, S.L. Oxygen-induced constriction of rabbit ductus arteriosus occurs via inhibition of a 4-aminopyridine-, voltage-sensitive potassium channel. J. Clin. Investig. 1996, 98, 1959–1965. [Google Scholar] [CrossRef] [PubMed]
- Michelakis, E.D.; Rebeyka, I.; Wu, X.; Nsair, A.; Thebaud, B.; Hashimoto, K.; Dyck, J.R.; Haromy, A.; Harry, G.; Barr, A.; et al. O2 sensing in the human ductus arteriosus: Regulation of voltage-gated K+ channels in smooth muscle cells by a mitochondrial redox sensor. Circ. Res. 2002, 91, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Hong, Z.; Kutty, S.; Toth, P.T.; Marsboom, G.; Hammel, J.M.; Chamberlain, C.; Ryan, J.J.; Zhang, H.J.; Sharp, W.W.; Morrow, E.; et al. Role of dynamin-related protein 1 (Drp1)-mediated mitochondrial fission in oxygen sensing and constriction of the ductus arteriosus. Circ. Res. 2013, 112, 802–815. [Google Scholar] [CrossRef] [PubMed]
- Kajimoto, H.; Hashimoto, K.; Bonnet, S.N.; Haromy, A.; Harry, G.; Moudgil, R.; Nakanishi, T.; Rebeyka, I.; Thébaud, B.; Michelakis, E.D.; et al. Oxygen activates the Rho/Rho-kinase pathway and induces RhoB and ROCK-1 expression in human and rabbit ductus arteriosus by increasing mitochondria-derived reactive oxygen species: A newly recognized mechanism for sustaining ductal constriction. Circulation 2007, 115, 1777–1788. [Google Scholar] [CrossRef] [PubMed]
- Clyman, R.I.; Waleh, N.; Black, S.M.; Riemer, R.K.; Mauray, F.; Chen, Y.Q. Regulation of ductus arteriosus patency by nitric oxide in fetal lambs: The role of gestation, oxygen tension, and vasa vasorum. Pediatr. Res. 1998, 43, 633–644. [Google Scholar] [CrossRef] [PubMed]
- Seidner, S.R.; Chen, Y.Q.; Oprysko, P.R.; Mauray, F.; Mary, M.T.; Lin, E.; Koch, C.; Clyman, R.I. Combined prostaglandin and nitric oxide inhibition produces anatomic remodeling and closure of the ductus arteriosus in the premature newborn baboon. Pediatr. Res. 2001, 50, 365–373. [Google Scholar] [CrossRef] [PubMed]
- Sodini, D.; Baragatti, B.; Barogi, S.; Laubach, V.E.; Coceani, F. Indomethacin promotes nitric oxide function in the ductus arteriosus in the mouse. Br. J. Pharmacol. 2008, 153, 1631–1640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coceani, F.; Kelsey, L.; Seidlitz, E.; Korzekwa, K. Inhibition of the contraction of the ductus arteriosus to oxygen by 1-aminobenzotriazole, a mechanism-based inactivator of cytochrome P450. Br. J. Pharmacol. 1996, 117, 1586–1592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coceani, F.; Breen, C.A.; Leesm, J.G.; Falck, J.R.; Olley, P.M. Further evidence implicating a cytochrome P-450-mediated reaction in the contractile tension of the lamb ductus arteriosus. Circ. Res. 1988, 62, 471–477. [Google Scholar] [CrossRef] [PubMed]
- Toyoshima, K.; Momma, K.; Imamura, S.; Nakanishi, T. In vivo dilatation of the postnatal ductus arteriosus by atrial natriuretic peptide in the rat. Neonatology 2007, 92, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.R.; Jing, S.; Momma, K.; Nakanishi, T. The effect of vitamin A on contraction of the ductus arteriosus in fetal rat. Pediatr. Res. 2001, 49, 747–754. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, U.; Minamisawa, S.; Adachi-Akahane, S.; Akaike, T.; Naguro, I.; Funakoshi, K.; Iwamoto, M.; Nakagome, M.; Uemura, N.; Hori, H.; et al. Multiple transcripts of Ca2+ channel alpha1-subunits and a novel spliced variant of the alpha1C-subunit in rat ductus arteriosus. Am. J. Physiol. Heart Circ. Physiol. 2006, 290, H1660–H1670. [Google Scholar] [CrossRef] [PubMed]
- Coceani, F.; Kelsey, L.; Seidlitz, E.; Marks, G.S.; McLaughlin, B.E.; Vreman, H.J.; Stevenson, D.K.; Rabinovitch, M.; Ackerley, C. Carbon monoxide formation in the ductus arteriosus in the lamb: Implications for the regulation of muscle tone. Br. J. Pharmacol. 1997, 120, 599–608. [Google Scholar] [CrossRef] [PubMed]
- Coceani, F.; Kelsey, L.; Seidlitz, E. Carbon monoxide-induced relaxation of the ductus arteriosus in the lamb: Evidence against the prime role of guanylyl cyclase. Br. J. Pharmacol. 1996, 118, 1689–1696. [Google Scholar] [CrossRef] [PubMed]
- Fujita, S.; Yokoyama, U.; Ishiwata, R.; Aoki, R.; Nagao, K.; Masukawa, D.; Umemura, M.; Fujita, T.; Iwasaki, S.; Nishimaki, S.; et al. Glutamate promotes contraction of the rat ductus arteriosus. Circ. J. 2016, 80, 2388–2396. [Google Scholar] [CrossRef] [PubMed]
- Baragatti, B.; Ciofini, E.; Sodini, D.; Luin, S.; Scebba, F.; Coceani, F. Hydrogen sulfide in the mouse ductus arteriosus: A naturally occurring relaxant with potential EDHF function. Am. J. Physiol. Heart Circ. Physiol. 2013, 304, H927–H934. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.H.; Yan, C.D.; Bian, J.S. Hydrogen sulfide: A novel signaling molecule in the vascular system. J. Cardiovasc. Pharmacol. 2011, 58, 560–569. [Google Scholar] [CrossRef] [PubMed]
- Aoki, R.; Yokoyama, U.; Ichikawa, Y.; Taguri, M.; Kumagaya, S.; Ishiwata, R.; Yanai, C.; Fujita, S.; Umemura, M.; Fujita, T.; et al. Decreased serum osmolality promotes ductus arteriosus constriction. Cardiovasc. Res. 2014, 104, 326–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bateson, E.A.; Schulz, R.; Olley, P.M. Response of fetal rabbit ductus arteriosus to bradykinin: Role of nitric oxide, prostaglandins, and bradykinin receptors. Pediatr. Res. 1999, 45, 568–574. [Google Scholar] [CrossRef] [PubMed]
- Clyman, R.I.; Mauray, F.; Roman, C.; Rudolph, A.M.; Heymann, M.A. Glucocorticoids alter the sensitivity of the lamb ductus arteriosus to prostaglandin E2. J. Pediatr. 1981, 98, 126–128. [Google Scholar] [CrossRef]
- Momma, K.; Takao, A. Increased constriction of the ductus arteriosus with combined administration of indomethacin and betamethasone in fetal rats. Pediatr. Res. 1989, 25, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Dunham-Snary, K.J.; Hong, Z.G.; Xiong, P.Y.; Del Paggio, J.C.; Herr, J.E.; Johri, A.M.; Archer, S.L. A mitochondrial redox oxygen sensor in the pulmonary vasculature and ductus arteriosus. Pflugers Arch. 2016, 468, 43–58. [Google Scholar] [CrossRef] [PubMed]
- Coceani, F.; Armstrong, C.; Kelsey, L. Endothelin is a potent constrictor of the lamb ductus arteriosus. Can. J. Physiol. Pharmacol. 1989, 67, 902–904. [Google Scholar] [CrossRef] [PubMed]
- Coceani, F.; Liu, Y.-A.; Seidlitz, E.; Kelsey, L.; Kuwaki, T.; Ackerley, C.; Yanagisawa, M. Endothelin A receptor is necessary for O2 constriction but not closure of ductus arteriosus. Am. J. Physiol. 1999, 277, H1521–H1531. [Google Scholar] [CrossRef] [PubMed]
- Coceani, F.; Kelsey, L.; Seidlitz, E. Evidence for an effector role of endothelin in closure of the ductus arteriosus at birth. Can. J. Physiol. Pharmacol. 1992, 70, 1061–1064. [Google Scholar] [CrossRef] [PubMed]
- Freed, M.D.; Heymann, M.A.; Lewis, A.B.; Roehl, S.L.; Kensey, R.C. Prostaglandin E1 infants with ductus arteriosus-dependent congenital heart disease. Circulation 1981, 64, 899–905. [Google Scholar] [CrossRef] [PubMed]
- Clyman, R.I. Mechanisms regulating the ductus arteriosus. Biol. Neonate 2006, 89, 330–335. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.C.; McGrath, J.C. Prostaglandin E2 and fetal oxygen tension synergistically inhibit response of isolated fetal rabbit ductus arteriosus to norepinephrine. J. Cardiovasc. Pharmacol. 1991, 17, 861–866. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.C.; McGrath, J.C. Characterisation of the effect of oxygen tension on response of fetal rabbit ductus arteriosus to vasodilators. Cardiovasc. Res. 1993, 27, 2205–2211. [Google Scholar] [CrossRef] [PubMed]
- Walford, G.; Loscalzo, J. Nitric oxide in vascular biology. J. Thromb. Haemost. 2003, 1, 2112–2118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, J.H.; Yang, S.N.; Chen, H.L.; Tseng, H.I.; Dai, Z.K.; Wu, J.R. B-type natriuretic peptide predicts responses to indomethacin in premature neonates with patent ductus arteriosus. J. Pediatr. 2010, 157, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Levin, E.R.; Gardner, D.G.; Samson, W.K. Natriuretic peptides. N. Engl. J. Med. 1998, 339, 321–328. [Google Scholar] [PubMed]
- Hsu, J.H.; Liou, S.F.; Yang, S.N.; Wu, B.N.; Dai, Z.K.; Chen, I.J.; Yeh, J.L.; Wu, J.R. B-type natriuretic peptide inhibits angiotensin II-induced proliferation and migration of pulmonary arterial smooth muscle cells. Pediatr. Pulmonol. 2014, 49, 734–744. [Google Scholar] [CrossRef] [PubMed]
- Van der Sterren, S.; Kleikers, P.; Zimmermann, L.J.; Villamor, E. Vasoactivity of the gasotransmitters hydrogen sulfide and carbon monoxide in the chicken ductus arteriosus. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 301, R1186–R1198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokoyama, U.; Minamisawa, S.; Quan, H.; Akaike, T.; Suzuki, S.; Jin, M.; Jiao, Q.; Watanabe, M.; Otsu, K.; Iwasaki, S.; et al. Prostaglandin E2-activated Epac promotes neointimal formation of the rat ductus arteriosus by a process distinct from that of cAMP-dependent protein kinase A. J. Biol. Chem. 2008, 283, 28702–28709. [Google Scholar] [CrossRef] [PubMed]
- Tannenbaum, J.E.; Waleh, N.S.; Mauray, F.; Breuss, J.; Pytela, R.; Kramer, R.H.; Clyman, R.I. Transforming growth factor beta 1 inhibits fetal lamb ductus arteriosus smooth muscle cell migration. Pediatr. Res. 1995, 37, 561–570. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.R.; Yeh, J.L.; Liou, S.F.; Dai, Z.K.; Wu, B.N.; Hsu, J.H. Gamma-secretase inhibitor prevents proliferation and migration of ductus arteriosus smooth muscle cells through the Notch3-HES1/2/5 pathway. Int. J. Biol. Sci. 2016, 12, 1063–1073. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, U.; Minamisawa, S.; Quan, H.; Ghatak, S.; Akaike, T.; Segi-Nishida, E.; Iwasaki, S.; Iwamoto, M.; Misra, S.; Tamura, K.; et al. Chronic activation of the prostaglandin receptor EP4 promotes hyaluronan-mediated neointimal formation in the ductus arteriosus. J. Clin. Investig. 2006, 116, 3026–3034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mason, C.A.; Bigras, J.L.; O’Blenes, S.B.; Zhou, B.; McIntyre, B.; Nakamura, N.; Kaneda, Y.; Rabinovitch, M. Gene transfer in utero biologically engineers a patent ductus arteriosus in lambs by arresting fibronectin-dependent neointimal formation. Nat. Med. 1999, 5, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Clyman, R.I.; Seidner, S.R.; Kajino, H.; Roman, C.; Koch, C.J.; Ferrara, N.; Waleh, N.; Mauray, F.; Chen, Y.Q.; Perkett, E.A.; et al. VEGF regulates remodeling during permanent anatomic closure of the ductus arteriosus. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2002, 282, R199–R206. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.H.; Xu, S.J.; Teng, J.Y.; Wu, W.; Ye, D.Y.; Wu, X.Z. Differential response of human fetal smooth muscle cells from arterial duct to retinoid acid. Acta Pharmacol. Sin. 2008, 29, 413–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokoyama, U.; Sato, Y.; Akaike, T.; Ishida, S.; Sawada, J.; Nagao, T.; Quan, H.; Jin, M.; Iwamoto, M.; Yokota, S.; et al. Maternal vitamin A alters gene profiles and structural maturation of the rat ductus arteriosus. Physiol. Genom. 2007, 31, 139–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boudreau, N.; Clausell, N.; Boyle, J.; Rabinovitch, M. Transforming growth factor-beta regulates increased ductus arteriosus endothelial glycosaminoglycan synthesis and a post-transcriptional mechanism controls increased smooth muscle fibronectin, features associated with intimal proliferation. Lab. Investig. 1992, 67, 350–359. [Google Scholar] [PubMed]
- Iwasaki, S.; Minamisawa, S.; Yokoyama, U.; Akaike, T.; Quan, H.; Nagashima, Y.; Nishimaki, S.; Ishikawa, Y.; Yokota, S. Interleukin-15 inhibits smooth muscle cell proliferation and hyaluronan production in rat ductus arteriosus. Pediatr. Res. 2007, 62, 392–398. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, U.; Minamisawa, S.; Shioda, A.; Ishiwata, R.; Jin, M.H.; Masuda, M.; Asou, T.; Sugimoto, Y.; Aoki, H.; Nakamura, T.; et al. Prostaglandin E2 inhibits elastogenesis in the ductus arteriosus via EP4 signaling. Circulation 2014, 129, 487–496. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, S.; Minamisawa, S. Oxygenation decreases elastin secretion from rat ductus arteriosus smooth muscle cells. Pediatr. Int. 2015, 57, 541–545. [Google Scholar] [CrossRef] [PubMed]
- Clyman, R.I.; Chan, C.Y.; Mauray, F.; Chen, Y.Q.; Cox, W.; Seidner, S.R.; Lord, E.M.; Weiss, H.; Waleh, N.; Evans, S.M.; et al. Permanent anatomic closure of the ductus arteriosus in newborn baboons: The roles of postnatal constriction, hypoxia, and gestation. Pediatr. Res. 1999, 45, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Waleh, N.; Seidner, S.; McCurnin, D.; Giavedoni, L.; Hodara, V.; Goelz, S.; Liu, B.M.; Roman, C.; Clyman, R.I. Anatomic closure of the premature patent ductus arteriosus: The role of CD14+/CD163+ mononuclear cells and VEGF in neointimal mound formation. Pediatr. Res. 2011, 70, 332–338. [Google Scholar] [CrossRef] [PubMed]
- Hinek, A.; Mecham, R.P.; Keeley, F.; Rabinovitch, M. Impaired elastin fiber assembly related to reduced 67-kD elastin-binding protein in fetal lamb ductus arteriosus and in cultured aortic smooth muscle cells treated with chondroitin sulfate. J. Clin. Investig. 1991, 88, 2083–2094. [Google Scholar] [CrossRef] [PubMed]
- Engur, D.; Kaynak-Turkmen, M.; Deveci, M.; Yenisey, C. Platelets and platelet-derived growth factor in closure of the ductus arteriosus. Turk. J. Pediatr. 2015, 57, 242–247. [Google Scholar] [PubMed]
- Gittenberger-de Groot, A.C.; Strengers, J.L.; Mentink, M.; Poelmann, R.E.; Patterson, D.F. Histologic studies on normal and persistent ductus arteriosus in the dog. J. Am. Coll. Cardiol. 1985, 6, 394–404. [Google Scholar] [CrossRef] [Green Version]
- Lezoualc’h, F.; Fazal, L.; Laudette, M.; Conte, C. Cyclic AMP sensor EPAC proteins and their role in cardiovascular function and disease. Circ. Res. 2016, 118, 881–897. [Google Scholar] [CrossRef] [PubMed]
- Tannenbaum, J.E.; Waleh, N.S.; Mauray, F.; Gold, L.; Perkett, E.A.; Clyman, R.I. Transforming growth factor-beta protein and messenger RNA expression is increased in the closing ductus arteriosus. Pediatr. Res. 1996, 39, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Boucher, J.; Gridley, T.; Liaw, L. Molecular pathways of notch signaling in vascular smooth muscle cells. Front. Physiol. 2012, 3, 81. [Google Scholar] [CrossRef] [PubMed]
- Baeten, J.T.; Jackson, A.R.; McHugh, K.M.; Lilly, B. Loss of Notch2 and Notch3 in vascular smooth muscle causes patent ductus arteriosus. Genesis 2015, 53, 738–748. [Google Scholar] [CrossRef] [PubMed]
- Krebs, L.T.; Norton, C.R.; Gridley, T. Notch signal reception is required in vascular smooth muscle cells for ductus arteriosus closure. Genesis 2016, 54, 86–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexander, M.R.; Owens, G.K. Epigenetic control of smooth muscle cell differentiation and phenotypic switching in vascular development and disease. Annu. Rev. Physiol. 2012, 74, 13–40. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, U.; Minamisawa, S.; Ishikawa, Y. Regulation of vascular tone and remodeling of the ductus arteriosus. J. Smooth Muscle Res. 2010, 46, 77–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinek, A.; Boyle, J.; Rabinovitch, M. Vascular smooth muscle cell detachment from elastin and migration through elastic laminae is promoted by chondroitin sulfate-induced “shedding” of the 67-kDa cell surface elastin binding protein. Exp. Cell Res. 1992, 203, 344–353. [Google Scholar] [CrossRef]
- Rabinovitch, M. Cell-extracellular matrix interactions in the ductus arteriosus and perinatal pulmonary circulation. Semin. Perinatol. 1996, 20, 531–541. [Google Scholar] [CrossRef]
- Clyman, R.I.; Goetzman, B.W.; Chen, Y.Q.; Mauray, F.; Kramer, R.H.; Pytela, R.; Schnapp, L.M. Changes in endothelial cell and smooth muscle cell integrin expression during closure of the ductus arteriosus: An immunohistochemical comparison of the fetal, preterm newborn, and full-term newborn rhesus monkey ductus. Pediatr. Res. 1996, 40, 198–208. [Google Scholar] [CrossRef] [PubMed]
- Clyman, R.I.; Mauray, F.; Kramer, R.H. Beta 1 and beta 3 integrins have different roles in the adhesion and migration of vascular smooth muscle cells on extracellular matrix. Exp. Cell Res. 1992, 200, 272–284. [Google Scholar] [CrossRef]
- Waleh, N.; Seidner, S.; McCurnin, D.; Yoder, B.; Liu, B.M.; Roman, C.; Mauray, F.; Clyman, R.I. The role of monocyte-derived cells and inflammation in baboon ductus arteriosus remodeling. Pediatr. Res. 2005, 57, 254–262. [Google Scholar] [CrossRef] [PubMed]
- Echtler, K.; Stark, K.; Lorenz, M.; Kerstan, S.; Walch, A.; Jennen, L.; Rudelius, M.; Seidl, S.; Kremmer, E.; Emambokus, N.R.; et al. Platelets contribute to postnatal occlusion of the ductus arteriosus. Nat. Med. 2010, 16, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Dani, C.; Poggi, C.; Fontanelli, G. Relationship between platelet count and volume and spontaneous and pharmacological closure of ductus arteriosus in preterm infants. Am. J. Perinatol. 2013, 30, 359–364. [Google Scholar] [PubMed]
- Mitra, S.; Chan, A.K.; Paes, B.A. The association of platelets with failed patent ductus arteriosus closure after a primary course of indomethacin or ibuprof1en: A systematic review and meta-analysis. J. Matern. Fetal Neonatal Med. 2017, 30, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Ohlsson, A.; Walia, R.; Shah, S.S. Ibuprofen for the treatment of patent ductus arteriosus in preterm or low birth weight (or both) infants. Cochrane Database Syst. Rev. 2015, CD003481. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, U.; Iwatsubo, K.; Umemura, M.; Fujita, T.; Ishikawa, Y. The prostanoid EP4 receptor and its signaling pathway. Pharmacol. Rev. 2013, 65, 1010–1052. [Google Scholar] [CrossRef] [PubMed]
- Hammerman, C.; Bin-Nun, A.; Markovitch, E.; Schimmel, M.S.; Kaplan, M.; Fink, D. Ductal closure with paracetamol: A surprising new approach to patent ductus arteriosus treatment. Pediatrics 2011, 128, e1618–e1621. [Google Scholar] [CrossRef] [PubMed]
- Lucas, R.; Warner, T.D.; Vojnovic, I.; Mitchell, J.A. Cellular mechanisms of acetaminophen: Role of cyclo-oxygenase. FASEB J. 2005, 19, 635–637. [Google Scholar] [CrossRef] [PubMed]
- Dang, D.; Wang, D.; Zhang, C.; Zhou, W.; Zhou, Q.; Wu, H. Comparison of oral paracetamol versus ibuprofen in premature infants with patent ductus arteriosus: A randomized controlled trial. PLoS ONE 2013, 8, e77888. [Google Scholar] [CrossRef] [PubMed]
- Bagheri, M.M.; Niknafs, P.; Sabsevari, F.; Torabi, M.H.; Bijari, B.B.; Noroozi, E.; Mossavi, H. Comparison of Oral Acetaminophen Versus Ibuprofen in Premature Infants With Patent Ductus Arteriosus. Iran. J. Pediatr. 2016, 26, e3975. [Google Scholar] [CrossRef] [PubMed]
- Roofthooft, D.W.; van Beynum, I.M.; de Klerk, J.C.; van Dijk, M.; van den Anker, J.N.; Reiss, I.K.; Tibboel, D.; Simons, S.H. Limited effects of intravenous paracetamol on patent ductus arteriosus in very low birth weight infants with contraindications for ibuprofen or after ibuprofen failure. Eur. J. Pediatr. 2015, 174, 1433–1440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ichikawa, Y.; Yokoyama, U.; Iwamoto, M.; Oshikawa, J.; Okumura, S.; Sato, M.; Yokota, S.; Masuda, M.; Asou, T.; Ishikawa, Y. Inhibition of phosphodiesterase type 3 dilates the rat ductus arteriosus without inducing intimal thickening. Circ. J. 2012, 76, 245624–245664. [Google Scholar] [CrossRef]
- Takizawa, T.; Oda, T.; Arishima, K.; Yamamoto, M.; Somiya, H.; Eguchi, Y.; Shiota, K. Inhibitory effect of enalapril on the constriction of the ductus arteriosus in newborn rats. J. Vet. Med. Sci. 1994, 56, 605–606. [Google Scholar] [CrossRef] [PubMed]
- Takizawa, T.; Horikoshi, E.; Shen, M.H.; Masaoka, T.; Takagi, H.; Yamamoto, M.; Kasai, K.; Arishima, K. Effects of TAK-044, a nonselective endothelin receptor antagonist, on the spontaneous and indomethacin- or methylene blue-induced constriction of the ductus arteriosus in rats. J. Vet. Med. Sci. 2000, 62, 505–509. [Google Scholar] [CrossRef] [PubMed]
- Fox, J.J.; Ziegler, J.W.; Ivy, D.D.; Halbower, A.C.; Kinsella, J.P.; Abman, S.H. Role of nitric oxide and cGMP system in regulation of ductus arteriosus tone in ovine fetus. Am. J. Physiol. 1996, 271, H2638–H2645. [Google Scholar] [CrossRef] [PubMed]
- Coceani, F.; Olley, P.M. The response of the ductus arteriosus to prostaglandins. Can. J. Physiol. Pharmacol. 1973, 51, 220–225. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Pathways mediating functional closure of the ductus arteriosus. AC: adenylyl cyclase, cAMP: cyclic adenosine monophosphate, cGMP: cyclic guanosine monophosphate, EPs: PGE2 receptors, ET: endothelin, GluR1: glutamate inotropic receptor subunit 1, Mito: mitochondria, PGE2: prostaglandin E2, PKA: protein kinase A, PKG: protein kinase G, RA: retinoic acid, SMC: smooth muscle cells, TRPM3: transient receptor potential melastatin 3.
Figure 1.
Pathways mediating functional closure of the ductus arteriosus. AC: adenylyl cyclase, cAMP: cyclic adenosine monophosphate, cGMP: cyclic guanosine monophosphate, EPs: PGE2 receptors, ET: endothelin, GluR1: glutamate inotropic receptor subunit 1, Mito: mitochondria, PGE2: prostaglandin E2, PKA: protein kinase A, PKG: protein kinase G, RA: retinoic acid, SMC: smooth muscle cells, TRPM3: transient receptor potential melastatin 3.

Figure 2.
The diagram of anatomical closure of the ductus arteriosus. CS: chondroitin sulfate, EC: endothelial cells, ECM: extracellular matrix, IEL: internal elastic laminae, IL-15: Interleukin-15, Mono: monocyte, PDGF: platelet-derived growth factor, PGE2: prostaglandin E2, PLT: platelet, RA: retinoic acid, SMC: smooth muscle cells, TGF-β: transforming growth factor-β, VEGF: vascular endothelial growth factor.
Figure 2.
The diagram of anatomical closure of the ductus arteriosus. CS: chondroitin sulfate, EC: endothelial cells, ECM: extracellular matrix, IEL: internal elastic laminae, IL-15: Interleukin-15, Mono: monocyte, PDGF: platelet-derived growth factor, PGE2: prostaglandin E2, PLT: platelet, RA: retinoic acid, SMC: smooth muscle cells, TGF-β: transforming growth factor-β, VEGF: vascular endothelial growth factor.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Factors mediating functional closure of the ductus arteriosus.
| Vasoconstrictors | References | Vasodilators | References |
|---|---|---|---|
| Oxygen sensing | Prostaglandin E2 | [10,11,12,13] | |
| Mitochondria | [14,15,16,17] | Nitric oxide | [18,19,20] |
| Cytochrome P450 | [21,22] | Natriuretic peptides | [23] |
| Retinoic acid | [24,25] | Carbon monoxide | [26,27] |
| Glutamate | [28] | Hydrogen sulfide | [29,30] |
| Hypoosmolality | [31] | ||
| Bradykinin | [32] | ||
| Corticosteroid | [33,34] |
Table 2.
Factors mediating anatomical closure of the ductus arteriosus.
| Cells | Mechanisms | Factors | Effects | Reference |
|---|---|---|---|---|
| SMCs | Migration | PGE2 | + | [48] |
| TGF-β1 | − | [49] | ||
| Notch | + | [50] | ||
| Fibronectin & Hyaluronan | + | [51,52] | ||
| VEGF | + | [53] | ||
| Proliferation | Retinoic acid | + | [54] | |
| Notch | + | [50] | ||
| ECM production | Hyaluronan | Retinoic acid | + | [55] |
| TGF-β | + | [56] | ||
| PGE2 | + | [51] | ||
| IL-15 | − | [57] | ||
| Fibronectin | Retinoic acid | + | [55] | |
| Chondroitin sulfate | TGF-β | + | [56] | |
| Elastin | PGE2 | − | [58] | |
| Oxygen | − | [59] | ||
| ECs | Proliferation | VEGF | + | [53,60,61] |
| IEL | Disruption | Chondroitin sulfate | + | [62] |
| Blood cells | Mononuclear cells adhesion | VEGF | + | [61] |
| Platelet plug | PDGF | + | [63] |
ECs: endothelial cells, ECM: extracellular matrix, IEL: internal elastic laminae, IL-15: Interleukin-15, PDGF: platelet-derived growth factor, PGE2: prostaglandin E2, SMCs: smooth muscle cells, TGF-β: transforming growth factor-β, VEGF: vascular endothelial growth factor.
Table 3.
Clinical and experimental agents for management of ductus arteriosus.
| Ductus Closure | References | Ductus Patency | References |
|---|---|---|---|
| Indomethacin * | [80] | Notch inhibitor | [50] |
| Ibuprofen * | [80] | Prostaglandin E1 * | [81] |
| Acetaminophen | [82,83,84,85,86] | Milrinone | [87] |
| Enalapril | [88] | ||
| Endothelin receptor antagonist | [89] | ||
| Nitric oxide | [90] |
* Currently used drugs in patients.
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hung, Y.-C.; Yeh, J.-L.; Hsu, J.-H. Molecular Mechanisms for Regulating Postnatal Ductus Arteriosus Closure. Int. J. Mol. Sci. 2018, 19, 1861. https://doi.org/10.3390/ijms19071861
AMA Style
Hung Y-C, Yeh J-L, Hsu J-H. Molecular Mechanisms for Regulating Postnatal Ductus Arteriosus Closure. International Journal of Molecular Sciences. 2018; 19(7):1861. https://doi.org/10.3390/ijms19071861
Chicago/Turabian StyleHung, Yu-Chi, Jwu-Lai Yeh, and Jong-Hau Hsu. 2018. "Molecular Mechanisms for Regulating Postnatal Ductus Arteriosus Closure" International Journal of Molecular Sciences 19, no. 7: 1861. https://doi.org/10.3390/ijms19071861
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.