Obesity and Aging in the Drosophila Model


1
Department of Zoology, Stockholm University, Svante Arrhenius väg 18B, S-106 91 Stockholm, Sweden
2
Institute of Zoology, Slovak Academy of Sciences, Dúbravská cesta 9, 845 06 Bratislava, Slovakia
*
Authors to whom correspondence should be addressed.
Int. J. Mol. Sci. 2018, 19(7), 1896; https://doi.org/10.3390/ijms19071896
Submission received: 19 May 2018
/
Revised: 19 June 2018
/
Accepted: 25 June 2018
/
Published: 27 June 2018
(This article belongs to the Special Issue Drosophila Model and Human Disease)
Abstract
:Being overweight increases the risk of many metabolic disorders, but how it affects lifespan is not completely clear. Not all obese people become ill, and the exact mechanism that turns excessive fat storage into a health-threatening state remains unknown. Drosophila melanogaster has served as an excellent model for many diseases, including obesity, diabetes, and hyperglycemia-associated disorders, such as cardiomyopathy or nephropathy. Here, we review the connections between fat storage and aging in different types of fly obesity. Whereas obesity induced by high-fat or high-sugar diet is associated with hyperglycemia, cardiomyopathy, and in some cases, shortening of lifespan, there are also examples in which obesity correlates with longevity. Transgenic lines with downregulations of the insulin/insulin-like growth factor (IIS) and target of rapamycin (TOR) signaling pathways, flies reared under dietary restriction, and even certain longevity selection lines are obese, yet long-lived. The mechanisms that underlie the differential lifespans in distinct types of obesity remain to be elucidated, but fat turnover, inflammatory pathways, and dysregulations of glucose metabolism may play key roles. Altogether, Drosophila is an excellent model to study the physiology of adiposity in both health and disease.
Keywords:
Drosophila; obesity; energy metabolism; lifespan; high-sugar diet; high-fat diet; dietary restriction; obesity paradox; AKH; IIS1. Introduction
Over the last several decades, obesity has become a global epidemic. According to the statement of the World Health Organization from 2017 [1], the prevalence of obesity has nearly tripled since 1975. More than 1.9 billion adults suffer from being overweight, and of these, over 650 million are obese. Obesity increases the risk of many health problems, including diabetes, metabolic syndrome, cardiovascular diseases and cancer, and hence leads to a higher mortality [2,3,4]. However, some studies questioned the causality between adiposity and mortality (e.g., [5,6]), arguing that the increased risk of death is associated with cardiorespiratory fitness, not with body mass index (BMI) [6]. In addition, a certain proportion of obese individuals do not suffer from any health complications nor increased mortality; this condition is known as metabolically healthy obesity (MHO) [7,8,9]. The prevalence of MHO ranges between 6–75% of all obesity cases [10]. The identification of mechanisms that underlie the adiposity-related complications in some, but not all individuals, represents an important challenge in the field of obesity research.
The core metabolic pathways that regulate energy homeostasis are highly evolutionarily conserved, and the fruit fly Drosophila melanogaster has served as an excellent model for metabolic and diet-associated diseases (reviewed in [11,12,13,14,15,16,17,18]). For example, flies have been used to investigate the metabolic control of tissue growth, and the link between the energy metabolism and cancer [18]. Drosophila has also been used to model obesity induced by high-sugar (HSD) [19,20,21,22,23,24,25] and high-fat diets (HFD) [26,27,28,29]. Moreover, like humans, flies fed a sugar-rich diet suffer from hyperglycemia, insulin resistance [20,22,24], and cardiomyopathy [19]. Fly obesity can also be induced by genetic manipulations, which allows decoupling of adiposity from changes in glycemia and carbohydrate metabolism [30]. Altogether, the Drosophila model provides excellent tools to disentangle the effects of excessive fat storage from other obesity-related factors that might be responsible for the metabolic dysregulations and lifespan shortening in some types of obesity.
This review aims to summarize the current knowledge on the lifespan and general physiology of obese flies. In the first part, we compare the regulation of energy storage in Drosophila and humans; in the second part, we discuss the lifespan consequences of different types of fly obesity. Whereas some obesity-inducing diets reduce lifespan expectancy [19,28], the excessive lipid storage can be uncoupled from the lifespan shortening [23,28,31]. Moreover, many fly models of longevity are actually obese (e.g., [32,33,34,35,36]). In the last part, we therefore discuss the potential adaptive roles of obesity and insulin resistance, and the possible mechanisms whereby excessive adiposity leads to metabolic complications and lifespan shortening in some, but not all types of obesity.
2. Energy Homeostasis in Drosophila and Humans
2.1. Circulating and Stored Sources of Energy
As in humans, the main circulating energy sources in Drosophila are sugars. However, the predominant sugar is trehalose, even though glucose is found in the fly hemolymph as well [37]. In contrast to the relatively low, but very stable glycemia in humans (0.1% glucose in the blood), insects have higher and more variable concentration of circulating sugars, with trehalose concentration in the hemolymph between 1–2% [38]. Trehalose is a non-reducing disaccharide, and therefore can be accumulated in the hemolymph at high levels without any detrimental effects [38]. Conditions that increase trehalose levels—such as a high-sugar diet—also increase circulating glucose [20]. Glucose can at least partially compensate for the lack of trehalose, as mutants deficient in trehalose production survive until the late pupal stage [39]. Before being utilized by cells, trehalose needs to be converted into glucose by trehalase [40,41,42]. Many studies therefore report just the total glycemia (e.g., [26,37,43,44]), without differentiating between the circulating sugars.
In contrast to the tight regulation of glycemia in humans, flies tolerate considerable fluctuations in the circulating sugars, from 50% reduction (e.g., [37,43]) to more than 50% increase (e.g., [33]). In humans, hyperglycemia leads to a damage of vascular endothelial cells, causing impairment of blood vessels, cardiovascular diseases, kidney failure, and blindness (reviewed e.g., in [45]). As Drosophila has an open circulatory system, damage to the vascular cells is not an issue. Altogether, hyperglycemia does not seem to be detrimental in this model system. Moreover, several manipulations that extend lifespan are, in the fruit flies, accompanied by increased levels of circulating sugars [21,33].
Like mammals, Drosophila stores excess chemical energy in the form of glycogen and lipids. Although glycogen accumulates also in muscles [46], the main storage organ for both carbohydrates and lipids is the fat body [46,47]—an organ analogous to the human adipose tissue, but also performing the functions of the liver [47] (Figure 1). The fat body consists of polyploid, sometimes multinucleate cells, which store lipids in specialized organelles called lipid droplets (Figure 2). Lipid droplets are conserved from yeast to humans, and their size and numbers per cell are highly variable [48,49]. However, apart from the increased number and volume of lipid droplets, very little is known about the obesity-related changes of the fat body.
2.2. Regulation of the Energy Balance by the Insulin-Like and Glucagon-Like Pathways
In mammals, the balance between circulating and stored energy sources is regulated by the antagonistic action of glucagon and insulin pathways. Whereas insulin triggers conversion of circulating sugars into the energy reserves, glucagon increases glycemia by promoting catabolism of fat and glycogen [50,51]. Flies have functional homologs of both insulin [32,33,52] and glucagon-like signaling [37,43,53], but not all the functions are analogous. For example, in the fly, glycogen mobilization is independent of the glucagon-like signaling [54].
Drosophila has eight insulin-like peptides (Ilp1–8) and a single insulin-like receptor, InR. As this pathway fulfills functions of both insulin and IGF (insulin-like growth factor) signaling, it is often abbreviated as IIS (insulin/insulin-like growth factor signaling) [33,55,56]. Ilp2, 3, and 5 are of special importance from the perspective of glycemia control. These hormones are produced in the median neurosecretory cells of the brain (Figure 1), and act similarly to human insulin produced in the pancreatic beta cells [32,52]. Mechanisms that regulate activity of these cells during development differ from those that act during adulthood [57]. For example, median neurosecretory cells of the larvae are not able to sense circulating sugars, and their activity is, therefore, regulated indirectly via the glucagon-like adipokinetic hormone (AKH) [58], and several other hormones [59,60]. By contrast, median neurosecretory cells of adults sense circulating glucose directly, via a mechanism similar to mammalian pancreatic β cells [61].
Fat body-specific manipulations of the InR indicate that analogously to human insulin, the fly IIS is a positive regulator of fat storage. Overexpression of a constitutively active isoform of InR increases fat content, while dominant negative isoform of InR reduces it [62]. On the other hand, several studies revealed that a global decrease of the IIS leads to obesity, as seen, for example, in the Ilp2-3,5 triple mutants [33], and in the flies with ablation of the insulin-producing median neurosecretory cells [32]. Similarly, genetic manipulations of the regulators of IIS from the IGF-binding family of proteins suggest that IIS is a negative regulator of fat reserves [63,64,65].
The role of the IIS in glycogen storage of flies seems to be analogous to the function of mammalian insulin. A recent study by Yamada et al. [47] showed that similarly to insulin in mammals [50], the fly IIS positively regulates glycogen synthesis. Nonetheless, ablation of the insulin-producing cells in the brain [32] or Ilp2-3,5 deficiency [33] lead to increased glycogen storage, suggesting that Drosophila has an additional IIS-independent factor that promotes glycogen synthesis.
In the fly, hyperglycemic functions are governed by the glucagon-like adipokinetic hormone AKH, which is produced in the endocrine organ called corpora cardiaca (Figure 1) [37,43,53]. Larval corpora cardiaca are localized in the ring gland [37,53]. The ring gland dissociates during metamorphosis, and the corpora cardiaca migrate towards the thorax, where it attaches to the esophagus, and sends axon-like projections toward the brain and crop [53]. AKH regulates catabolism of lipids analogously to mammalian glucagon. Thus, Akh mutants are obese [43,54], whereas Akh overexpression results in a lean phenotype [53,54]. However, in contrast to glucagon, AKH does not induce catabolism of glycogen [43,54] and glycogen levels rise upon Akh overexpression [54]. This is particularly intriguing, as AKH is a hyperglycemic hormone [37,43,53], and glycogen has been considered as the main source of trehalose [38]. Thus, the regulation of circulating and stored energy sources is more complicated, and we are still far from a complete understanding of the pathways that govern energy homeostasis in the Drosophila model.
3. Lifespan in Different Types of Fly Obesity
There are several methods to measure the fat content of a fly, but the most common approach is based on a colorimetric measurement of glycerides [66]. The fat levels are either expressed as absolute raw values, as fat values normalized to body weight, or as fat values normalized to the protein content. In contrast to the clear BMI-based definition of overweight and obesity in humans, obesity in flies is not exactly defined, and the term is used as an equivalent for increased fat storage.
3.1. High Sugar Diet (HSD)-Induced Obesity
Sugar-rich substrates represent a natural food source of fruit flies [67]. However, a high sugar diet (HSD) produces obesity and hyperglycemia [20]. HSD typically refers to a fly diet containing around 30% sugar, which accounts for an approximate increase in the fat content by 50–150% [19,20,21,23,68,69]. The HSD diet is prepared by adding excessive sucrose [19,20,22,23,31], glucose [21,70], or fructose [70] to the standard fly medium. HSD decreases several aspects of fly fitness; for example, delays larval development [20,23], reduces fecundity [71,72,73], and increases the age-independent mortality [23,72]. However, the overall effect on aging is not clear. Whereas some studies reported a lifespan-shortening effect of HSD [69,72], Galenza and colleagues [21] revealed that despite causing obesity and hyperglycemia, HSD leads to a remarkable lifespan extension by 31%. Interestingly, their study suggests an increase in the early mortality of flies reared on HSD, despite an overall lifespan extension in the majority of the flies [21]. The same phenomenon was described in the work of Tânia Reis [23], who showed that the mortality of the HSD flies has a bimodal distribution, although the mean lifespan upon HSD treatment is mildly increased.
HSD leads to an impairment of the IIS [20,21,22], although the mechanism is not entirely clear, and may depend on the developmental stage. A study by Pasco et al. [22] showed that in larvae, HSD increases expression of Ilp2, 3, and 5. However, Ilps cannot activate the InR receptor due to the increased levels of the Ilp-binding protein, lipocalin neural Lazarillo. Thus, HSD-fed larvae are insulin resistant, and their peripheral IIS is attenuated [22]. Musselman et al. [20] showed that the HSD-fed larvae are resistant even to recombinant insulin. The situation is likely different in adults, where HSD decreases the IIS independently of insulin resistance, causing decreased expression of Ilp2, 3, and 5 [24], whilst the periphery remains insulin sensitive [21].
Altogether, HSD reduces the peripheral IIS, either via decreased production of insulin, or via insulin resistance. Interestingly, the survival curves of the Ilp2-3,5 triple mutants [33] are reminiscent of the survival curves of flies fed HSD, that were described by Tânia Reis [23]. Moreover, the Ilp2-3,5 triple mutants [33] also recapitulate other HSD-associated impairments of development and reproduction [20,22,33,72], suggesting that the reduced IIS is responsible for many disorders linked to HSD. Nevertheless, the reduced IIS cannot account for the HSD-induced lifespan shortening detected in some studies [69,72], as attenuation of IIS is typically coupled with lifespan extension [32,33].
The lifespan extension reported for some obese and hyperglycemic HSD-fed flies [21,23] suggests that the adiposity itself is probably not the sole cause of the reduced lifespan in other HSD-feeding experiments [19,72]. Work of Na and colleagues [31] documented that HSD leads to dysfunction of pericardial nephrocytes, cells that filtrate fly hemolymph and act analogously to human podocytes in the kidney (Figure 1). This damage is associated with increased hexosamine flux and the Polycomb gene complex activity. Interestingly, pharmacological inhibition of the hexosamine pathway extends the lifespan on HSD—seemingly beyond the lifespan of controls kept on the standard food—while reducing fat levels only partially [31]. Thus, the HSD-induced damage of nephrocytes—rather than the adiposity itself—is responsible for the lifespan-shortening effect of this diet. In addition to causing dysfunctions of nephrocytes [31], HSD affects other processes that might contribute to the reduction of fly fitness. For example, a short-term exposure to HSD decreases lifespan via a transient inhibition of the IIS-repressed transcriptional factor Foxo (Forkhead box, sub-group O), which causes a long-lasting reprogramming of the signaling in the fat body [74]. Sugar overload also directly affects numerous fat body-unrelated processes; for example, it increases endoplasmic reticulum stress [68], decreases immunity [68], disrupts gut homeostasis, and reduces commensal bacteria [75]. Like in humans, HSD causes in flies heart disorders such as fibrillations, asystolic periods, and arrhythmias, leading to progressive heart failure [19]. The detrimental effect of HSD on the heart is mediated solely by the hexosamine flux, and the cardiac-specific reduction of this pathway fully protects the heart from the HSD-induced pathologies [19]. In addition, the recent RNA-seq analyses of the HSD-induced transcriptional changes may provide further useful hints on the metabolic changes elicited by the sugar overfeeding [76].
Flies fed on HSD (e.g., [19,20,21,23,68,69,70]) do not have access to any additional water source to compensate for the increased osmolarity of their diet. Part of the pathologies associated with HSD may, therefore, result from the hyperosmolarity or hypovolemia. A study by Rovenko et al. [70], showed that the HSD-fed flies have indeed reduced body water content, further supporting this hypothesis.
In conclusion, HSD may affect the fly physiology independently of the obese phenotype, for example, via direct damage of the heart [19], nephrocytes [31], or gut [75], via reprogramming of signaling pathways in the fat body [74], or via reduction of the body fluids [70]. The mechanism that circumvents these complications in the glucose-based HSD [21] remains to be elucidated, nevertheless, it might contribute to a better understanding of the MHO in humans as well.
3.2. Dietary Restriction and the Paradox of a Carbohydrate-Rich Diet
Lifespan extension by dietary restriction (DR) is a phenomenon conserved from yeast to humans. There are several methods to apply DR: intermittent fasting, restriction of certain nutrients, or food dilution [77,78,79]. Diet dilution is a frequently used method in various animal models, yet several studies showed that this treatment leads to increased, compensatory feeding. This was explained by the protein leverage hypothesis [80], which states that the reduction of proteins increases appetite. Thus, when the DR food is provided ad libitum, animals consume relatively higher proportion of carbohydrates and calories, which leads to obesity [80,81]. This mechanism seems to be evolutionarily conserved, with evidence for it found from fruit flies [82] to humans [80,83,84]. In the Drosophila field, the DR regime typically refers to the protein restriction. Restriction of dietary yeast, the main source of proteins in the fly diet, increases both lifespan and fat reserves [35,36,82,85]. Geometric framework studies in Drosophila showed that ad libitum feeding on high carbohydrate/low protein diet extends lifespan, whereas the caloric restriction itself does not [71,81,86,87]. Similar data were obtained in mice [88]. The question then arises: what is the mechanism whereby the high carbohydrate/low protein diet extends lifespan? DR reduces IIS activity [89], but the lifespan extension in flies is mediated by the target of rapamycin (TOR) signaling [35]. Interestingly, pharmacological inhibition of the TOR pathway by rapamycin not only extends lifespan [34], but also increases fat reserves [34,35].
Thus, both DR and genetic inhibition of the nutrient-sensing pathways extend lifespan and increase fat storage [32,34,35,36,85]. How exactly the obese phenotype contributes to the longevity is unknown. However, two studies [90,91] revealed that the DR-mediated lifespan extension is associated with a beneficial effect of increased fat turnover. This process is mediated by the lipolytic AKH hormone [91] and requires the peripheral circadian clock in the fat body [90]. It remains to be investigated whether a similar increase in the lipid metabolism accompanies obesity in other long-lived models, and whether an experimental increase in the fat turnover would rescue longevity of the short-lived obese flies.
3.3. The HFD-Induced Obesity
Natural food sources, as well as the standard culture media for Drosophila, are relatively poor in fat content. Nevertheless, flies can feed and survive on artificial experimental diets with added fat. The HFD for Drosophila typically contains 20% [92] to 30% [26,29,93,94] coconut oil, or 15% lard [28,95]. HFD leads to obesity [26,28,29,92], hyperglycemia [20,26,28], reduced cardiac contractility, ectopic accumulation of fat in the heart, and to other pathologies reminiscent of the diabetic cardiomyopathy [26]. HFD is also associated with lifespan reduction [28,29].
HFD increases TGF-β signaling, which is responsible for the development of insulin resistance [92]. Experimental reduction of TGF-β is sufficient to ameliorate both obese and hyperglycemic phenotype [92], but whether this treatment restores lifespan is not known. Interestingly, genetically enhanced lipolysis targeted to the heart is sufficient to prevent the HFD-triggered cardiac dysfunctions [26,93], suggesting that HSD causes cardiomyopathy in a tissue-autonomous manner via ectopic lipid storage. Ectopic fat accumulation appears to be a common mechanism behind the HFD-induced cardiac dysfunction also in mammals—a heart-specific increase in lipolysis improves the heart functions in mice as well [96,97,98].
Similarly to human obesity [99,100], HFD leads, in flies, to an overactivation of immune responses [28]. A macrophage-specific knockdown of the immune response restores insulin sensitivity and ameliorates the lifespan shortening, but nevertheless, does not ameliorate the HFD-induced obesity [28]. Whether and how the inflammatory pathways affect heart functions is, nevertheless, not known. In conclusion, it seems that—at least in the HFD model—lifespan is not shortened by the adiposity, but by the inflammatory pathways.
3.4. Obesity and Longevity in the Flies with Abrogated Reproduction
Reduced reproduction is associated with increased fat accumulation and longevity in multiple organisms (reviewed in [101]). Conversely, numerous treatments that result in longevity and excessive fat storage decrease fertility. In Drosophila, these examples include DR and inhibition of the nutrient-sensing pathways [32,33,34,102]. However, there are also cases where reduced reproduction correlates with obesity, but not with lifespan extension. Drosophila females homozygotic in the naturally-occurring allele female sterile (2) adipose are sterile and obese, yet not long-lived [103]. In addition, lifespan-extension associated with inhibition of breeding sometimes correlates with reduced fat reserves. For example, virgin Drosophila female flies have lowered starvation resistance, indicating that they are leaner than their reproducing siblings [104].
The interactions between the fat storage and lifespan might be partially explained by the trade-offs between energy allocation to egg production and fat storage (reviewed in [105]). Importantly, in addition to the direct costs of reproduction, fly gonads may regulate metabolism and lifespan by modulating the main signaling pathways, such as the IIS and steroid signaling. As a case in point, elimination of the germline cells in the ovaries or testis extends lifespan, and this effect is coupled with attenuated IIS [106].
Fly gonads also produce ecdysteroids [107,108], the only steroid hormones in Drosophila, which could mediate the trade-offs between reproduction and lifespan [109]. Although the experimental evidence in females is equivocal, reduced ecdysteroid levels or mild RNAi against the Ecdysone receptor (EcR) extends lifespan in males [110,111]. Interestingly, the functions of ecdysteroids in adult flies are reminiscent of the roles of human sex steroid hormones estrogen and testosterone. Like sex steroids, ecdysteroids regulate maturation (metamorphosis) [112] and reproduction [113,114,115]. In larvae, EcR acts as a negative regulator of fat accumulation [116], and it is possible that the same role is conserved in adults as well. In that case, Drosophila could be an appropriate model to study the obesity associated with reduced levels of sex steroids, a condition which occurs in humans after gonadectomy, menopause, or during aging [117,118,119].
3.5. Genetic Links between Fat Storage and Lifespan
The typical genetic interventions that result in a lifespan extension—such as attenuation of the IIS and TOR signaling—also result in obesity [32,33,34,35,36]. Similarly, the long-lived Methuselah (Mth) mutants have increased starvation resistance [120], indicating that their fat stores are increased. In addition, several selection experiments revealed existence of a genetic basis for the positive correlation between obesity and lifespan. For example, some Drosophila lines selected for longevity have increased resistance to starvation, suggesting higher energy reserves [121,122]. Conversely, selection for increased starvation resistance resulted in increased fat storage [123,124], suggesting that longevity and obesity are determined by the same genetic variants. Nevertheless, there are also longevity selection lines without increased resistance to starvation [125], as well as fly lines with reduced body fat [126]. Thus, the longevity can evolve in various ways, which may or may not involve an increase in the body fat.
Altogether, several selection experiments revealed the existence of a genetically based variation in the fat metabolism and lifespan, suggesting that the fat storage and longevity share a common genetic architecture [121,122,123,124]. The recent advancement of sequencing technologies, together with the genome-wide association studies on the inbred lines from the Drosophila Genetic Reference Panel, provide additional tools for identification of genes underlying the variation in fat storage and lifespan in wild-type flies [127,128,129,130]. Further studies of these adiposity-related alleles thus hold new promises for a better understanding of the fitness and lifespan in naturally obese flies.
3.6. Lifespan in Other Genetic Models of Obesity
Fly genetics provides numerous tools to induce obesity, such as manipulations of genes associated with adiposity in mice or humans, as well as mis-expressions of metabolic genes identified by the genome-wide RNAi screens in Drosophila [30,131,132]. These genetic manipulations allow for the analysis of the consequences of obesity without altering diet or the nutrient-sensing pathways. For example, severe obesity can be triggered via inhibition of either lipolytic pathway, one acting via the lipase Brummer, the other via the AKH hormone signaling [133]. Mutation of brummer (bmm) results in doubling of the fat, but only in a mild reduction of lifespan [134]. Similarly, the fat content of the Akh mutants is increased by around 75–100% [43,54], however, the lifespan is reduced only moderately (MG and PK; unpublished data [135]). On the other hand, increased lipolysis caused by Akh overexpression or by activation of the AKH secretion is associated with longevity [91,136]. Reduction of fat storage is, nevertheless, not sufficient for lifespan extension, as seen, for example, in the lean but short-lived mutants deficient for the α-Esterase-7 [137].
In contrast to HSD- or HFD-induced obesity, genetic interventions enable distinguishing the effects of adiposity from the consequences of hyperglycemia. For example, obesity associated with euglycemia can be triggered by manipulations of calcium signaling via Stim RNAi [30], whereas obesity coupled with hypoglycemia can be induced by the Akh mutations [43].
Altogether, as summarized in Table 1, some types of obesity are associated with lifespan shortening, whereas others with longevity.
4. Pathways and Tissues Linking Lipid Metabolism and Lifespan
Energy homeostasis of Drosophila is controlled by a complex neuroendocrine system with extensive interorgan communications (reviewed e.g., in [139,140,141,142,143]). Here, we focus only on the most prominent tissues and pathways that could link obesity and aging.
4.1. The IIS and AKH Endocrine Systems
Human obesity is often associated with profound changes in the production of adipose tissue-derived factors, such as leptin, adiponectin, and tumor-necrosis factor alpha (TNF-α). These dysregulations lead to reduced glucose uptake, increased glycemia, and development of insulin resistance (reviewed in [144,145])., Hyperglycemia-associated glucotoxicity damages, among other tissues, also pancreatic β cells, which further exacerbates the diabetic phenotype [145]. Similarly, many types of Drosophila obesity correlate with changes in signaling via fat body-produced hormones. For example, both HSD and HFD increase expression of the fly homolog of leptin, unpaired2 (upd2) [59]. Interestingly, the typical models of fly obesity (flies reared on DR, HSD, or HFD) also have reduced insulin-like signaling [21,89,92,146]. The mechanism of the IIS attenuation seems to be diet-specific. Whereas DR selectively decreases production of Ilp5 [89,146], HSD in adult flies reduces expression of all three brain-produced Ilps [24], while the periphery remains insulin-sensitive [21]. By contrast, the HFD-induced changes in the brain-produced Ilps are less clear [26,92]. Nevertheless, HFD leads to reduced peripheral IIS, and to insulin resistance [92]. Altogether, the diet-induced obesity of flies appears to be accompanied by reduced IIS, reminiscent of the diabetic phenotype of many obese patients. However, in contrast to humans, the attenuation of insulin signaling does not endanger health of the flies, but—on the contrary—extends their lifespan [32].
Diabetes and hyperglycemia in humans are associated with increased glucagon production, which further worsens the diabetic phenotype [145]. Interestingly, HSD in flies leads to increased signaling via the analogous AKH hormone [147]. Nevertheless—in contrast to humans—overactivation of the AKH signaling extends lifespan [91,136].
In summary, the diet-induced dysregulation of the IIS and AKH signaling in obese flies resembles the dysregulations of the corresponding pathways in obese humans. However, the health- and lifespan-associated consequences of these dysregulations differ tremendously between flies and humans.
4.2. The Fat Body
Similar to the human adipose tissue [144], the insect fat body is not only a deposit of neutral fat [148], but also an important endocrine and immune organ [82,139,141,148]. In addition, the fat body has a key role in the sensing of proteins and carbohydrates, and in the regulation of the hypothalamus-like centers in the brain according to these dietary cues [82]. These endocrine functions are mediated by several hormones secreted from the fat body, including the leptin analog Upd2, which signals the nutritional state by acting on the GABAeric neurons that regulate secretion of Ilp2 [59]. There are several other factors derived from the fat body that regulate the insulin-producing cells in the brain, including the peptide CCHamide-2 (CHHa2) with important developmental roles [60,149], and the satiety signal encoded by the female-specific independent of transformer (fit) [150].
The fat body negatively regulates the production of Ilps in the mid-brain via Ilp6 [138]. The fat body-specific overexpression of Ilp6 extends lifespan and fat storage [138], thus mimicking the ablation of the insulin-producing cells in the brain [32]. In addition to the regulation of insulin production, the fat body inhibits the IIS in the periphery by secreting Impl2, an Ilp2-binding protein from the IGF-binding protein family [64]. The fat body remotely regulates additional pathways and processes; for example, slows down the digestion via secreting the Activin-like ligand Dawdle, a repressor of digestive enzymes [151].
During larval development, the fat body has an important role in coupling nutrient availability to growth. The nutrient sensing occurs via several pathways, including the amino-acid transporter Slimfast, the TOR pathway [141], and the pathway activated by the systemically circulating lipoprotein-associated form of the signaling protein Hedgehog (Hh) [152]. The circulating Hh is produced by the midgut. The expression of hh increases upon starvation, when the Hh pathway promotes mobilization of lipid reserves in the fat body [152]. Interestingly, the Hh signaling was identified as the top-scoring anti-obesity pathway in a genome-wide obesity screen in adult Drosophila [132]. Hh plays a crucial role in fat metabolism also in mammals [132], and is considered an antagonist of aging and aging-associated diseases [153].
Many lifespan-extending manipulations, including those concerning the IIS and TOR pathways, act via the fat body, and they are frequently coupled with obesity. For example, the fat body-directed overexpression of foxo, a transcriptional factor repressed by the IIS, extends lifespan and increases lipid accumulation [154]. The nutrient-sensing TOR pathway regulates lifespan via the fat body as well [155]. Similarly, the fat body-specific inhibition of steroid signaling via EcR RNAi is sufficient for the lifespan extension [111].
4.3. The Heart
Several studies have shown the importance of the heart in a remote regulation of fat reserves. For example, a heart-specific signaling via Skuld—a subunit of the mediator transcriptional complex—remotely regulates the fat storage in flies [156]. Similarly, the mammalian homolog of Skuld, MED13, remotely regulates fat storage in mice [157]. The fly heart produces a substantial proportion of the apoB-lipoproteins—the essential lipid carriers [158]. Under HFD, the apolipoproteins derived from cardiomyocytes, and not those produced by the fat body, are the predominate regulators of lipid metabolism [158].
As described in the part on the diet-induced obesity (Section 3.1 and Section 3.3), the HSD- or HFD-induced cardiomyopathies [19,26,93] may contribute to the lowered fitness in these types of obesity. The heart damage seems to occur in a tissue-autonomous manner, either via the increased hexosamine flux (in the case of HSD) [19], or via the overactivation of the TOR pathway and ectopic fat accumulation (in the case of HFD) [93]. However, it is unknown whether the heart accumulates ectopic fat only under the HFD [93] and HSD [19] regimes, or also in other types of obesity. Similarly enigmatic is the mechanism that putatively prevents heart dysfunctions in the long-lived obese flies.
4.4. The Immune Cells
Human obesity is coupled with a chronic low-grade inflammation and activation of the immune response, which leads to insulin resistance and metabolic syndrome [99,100]. The precise mechanism behind the obesity-associated inflammation is not entirely clear. The immune response involves an increase in macrophage numbers, infiltration of the adipose tissue and other organs with macrophages, and enhanced production of proinflammatory cytokines TNF-α, interleukin 6 (IL-6), leptin, visfatin, resistin, and others [100,159]. These cytokines trigger chronic inflammation, which subsequently causes insulin resistance, hyperglycemia, and other metabolic complications [100]. In Drosophila, feeding on HFD leads to an immune response as well, namely, to the enhanced production of the macrophage-derived cytokine Upd3, which is responsible for the subsequent development of hyperglycemia and insulin resistance [28]. The elegant study of Woodcock and colleagues [28] showed that HFD shortens lifespan solely via the Upd3-dependent inflammatory response, and not via the increase in fat storage. It remains to be investigated whether HFD activates, in flies, production of other cytokines, and whether all types of fly obesity cause activation of the immune response. Altogether, it is possible that the activation of an immune response is a common feature triggered by adiposity. However, how is this process initiated under HFD, and avoided in the long-lived obese flies, remains to be investigated.
4.5. The Pericardial Nephrocytes
In humans, inflammation plays a central role in the development of diabetic nephropathy—end-stage kidney disease associated with diabetes [160,161]. Although accompanied by changes in several cell types in the kidney, diabetic nephropathy is caused mainly by the injury of podocytes, specialized cells that encircle capillaries of the glomerulus [162,163]. Nephrocytes—specialized cells that filter hemolymph—are the functional homologs of mammalian podocytes in the fly [164,165]. Analogous to the diabetic conditions in humans, HSD in flies leads to damage of the pericardial nephrocytes, which is a condition reminiscent of the diabetic nephropathy [31]. The podocyte injury is induced by increased hexosamine flux and enhanced activity of the Polycomb gene complex. Interestingly, inhibition of this pathway is sufficient to protect from the lifespan-shortening effect of HSD [31].
4.6. The Gut
The gut plays a central role in the digestion and absorption of nutrients. In addition, the gut regulates energy homeostasis via its endocrine and neuroendocrine functions [166]. The epithelial cells of the midgut produce at least 12 different peptide hormones [166]. These peptides include, for example, regulators of feeding, such as neuropeptide F (NPF) [167] and CCHa2 [60,168], and regulators of lipid metabolism, such as Tachykinin (Tk) [169]. The digestive tract also express starvation-inducible gene takeout (to) [166], which codes for a juvenile hormone-binding protein that promotes feeding [170,171] and extends lifespan [172]. Interestingly, to is activated under several lifespan-extending manipulations, including dietary restriction, downregulation of the IIS pathway, mth mutation, and repression of ecdysteroid signaling [172,173],
The adult gut of Drosophila is a plastic organ capable of regeneration; epithelium of the midgut renews within two weeks [166]. Maintaining intestinal homeostasis is important for healthy aging [174]. Senescence of the gut is associated with leakage of the epithelial barrier, increase in the proliferation of the stem cells and accumulation of the undifferentiated progenitor cells [166,174]. The loss of the tissue homeostasis leads to inflammation-like state, which activates the stress-induced Jun-N-terminal kinase (JNK) pathway [175]. The JNK pathway promotes further overproliferation of intestinal stem cells, which is directly responsible for lifespan shortening [176]. Overactivation of the JNK pathway also leads to the chronic activation of the IIS-repressible factor Foxo, which inhibits expression of the lipase Magro (Mag) [175], the key lipase necessary for digestion of triacylgylcerol [177]. Reduced Mag activity thus leads to lowered body fat content in old flies [175]. Experimental inhibition of the JNK pathway, or overexpression of Mag in the intestine increases fat reserves in old flies [175]. Moderate inhibition of the JNK pathway also extends lifespan [176], but whether this effect is mediated by improved lipid metabolism remains unclear. Mild increase in the intestinal Foxo signaling in aging flies might actually be an adaptive response, as lifespan is extended by overexpression of Foxo by GeneSwitch drivers with strong intestinal expression patterns, such as S106- and S32-GAL4 [154,178,179].
The microbiome of the fly gut depends on the diet [130], and contributes to the regulation of energy metabolism [180]. Elimination of the microbiota leads to obesity, despite lower food intake of axenic flies [180]. The effect of the gut bacteria on lifespan is less clear, as both lifespan shortening effect [181], as well as longevity where reported for axenic flies [182]. The effect of the antibiotic treatment on lifespan likely depends on the sex and age of the flies when antibiotics are applied [183].
Altogether, maintenance of the gut homeostasis is required for proper digestion and uptake of lipids, as well as for longevity. In addition, the gut may link energy metabolism and longevity via numerous endocrine and neuroendocrine factors, and via the gut microbiota.
5. The Adaptive Roles of Hyperglycemia and Obesity
5.1. The Thrifty Genotype
Diabetes mellitus type 2 is a clearly harmful, nevertheless, surprisingly widespread disease. In 1962, James Neel proposed the “thrifty genotype hypothesis” [184], arguing that in the course of evolution, “thrifty” genetic variants that promoted accumulation of fat were positively selected, because they enabled survival during famine periods. However, these genotypes became maladaptive in modern societies where nutrition is not restricted, being now responsible for obesity, diabetes type 2, and other metabolic complications. The thrifty genotype hypothesis predicted the existence of a genetic predisposition for obesity and diabetes, and recent genome-wide association studies indeed identified numerous obesity-associated loci [185]. Like human societies in the past, natural populations of Drosophila are exposed to fluctuations in the food availability. Interestingly, there is also a genetic variation in the fly genes associated with fat accumulation [127,128,129,130], suggesting that the thrifty genotype hypothesis might explain the evolution of some types of Drosophila obesity as well.
5.2. Lipogenesis as a Protective Mechanism against Glucose Toxicity
Lipogenesis can be considered as a protective mechanism of sugar detoxification [25]. The evolution of fat storage not only enables survival during periods of food shortage, but also provides a protection against hyperglycemia. However, when lipogenesis exceeds the storage capacity of the adipose tissues, fat starts to accumulate in other organs. The ectopic fat storage elicits lipotoxicity, which, in humans, leads to damage of the pancreatic beta cells, cardiomyocytes, hepatocytes, renal parenchymal cells, and endothelial cells [186]. It has been suggested that many medical complications of obesity result from the lipotoxic response in non-adipose tissues, rather than from accumulation of fat in the adipocytes [186]. Thus, an increase in the lipid storage capacity of adipocytes might actually be beneficial. This notion has been confirmed in the Drosophila models of the HSD-induced obesity, in which an experimental increase in the fat storage improved tolerance to dietary sugars, whereas genetic interventions leading to a lean phenotype exacerbated the consequences of HSD [25]. These experiments highlight the importance of understanding the physiology of different types of obesity, as the reduction of body fat may actually worsen health under certain conditions.
5.3. The “Obesity Paradox” and the Beneficial Roles of Body Fat
The fat reserves clearly provide an advantage under conditions of nutrient scarcity. Nevertheless, numerous studies showed that the fat might have a protective role under ad libitum feeding as well. For example, increased BMI correlates with an improved prognosis and lowered mortality in several medical conditions. These include, for example, chronic heart failure and coronary diseases [187,188], end-stage kidney disease [189], and cancer [190]. This phenomenon—a better prognosis of obese individuals—is known also as the “obesity paradox”.
Obesity typically correlates with increased all-cause mortality [3,4]. Nevertheless, a meta-analysis study by Flegal et al. [2] showed that whereas this is true for obesity (BMI ≥ 30), overweight (BMI of 25 < 30) is actually associated with a significantly lower all-cause mortality, suggesting that there are instances in which increased body fat provides an advantage. A positive role of stored lipids has been shown in several others systems, including the budding yeast Saccharomyces cerevisiae, in which excessive fat increases lifespan [191]. A recent review on the protective roles of triacylglycerides argued that neutral fat prolongs lifespan in an energy-independent fashion, by protecting against various stressors [192]. Interestingly, lipid droplets have antioxidant roles and protect stem cells also in flies [193], suggesting that this beneficial function may involve other fly tissues as well. Longevity models of Drosophila—such as the flies with downregulated TOR [34] and IIS [32] signaling, and the Methuselah mutants [194]—have increased resistance to paraquat, a commonly used agent to elicit oxidative stress in insects. Thus, it is tempting to speculate on a potential antioxidant role of the lipid droplets in these long-lived obese flies.
5.4. Adaptive Roles of the Stress-Induced Hyperglycemia
Chronic diabetes is dangerous for human health, yet there are instances when hyperglycemia provides an advantage. In contrast to the chronic hyperglycemia, stress hyperglycemia is an important, evolutionarily conserved mechanism to cope with injury and infection (reviewed in [195,196]). In humans, this type of hyperglycemia is hormonally regulated by the hypothalamus–pituitary–adrenal axis, sympathoadrenal system, and proinflammatory cytokines, which collectively induce gluconeogenesis, glycogenolysis, and insulin resistance [195]. Stress-induced hyperglycemia was reported in organisms ranging from worms to humans [196]. In Drosophila, an increase in circulating sugars seems to be a common response to various forms of infections, including bacterial [197], nematode [198], and wasp parasites [199]. This mechanism is highly adaptive, documented by the fact that inhibition of the rise in circulating sugars, either by inhibition of the adenosine transport or AKH signaling, increases sensitivity to the infections [198,199]. The infection-induced hyperglycemia is likely mediated by the attenuation of the IIS pathway, as shown, for example, for the infection by Mycobacterium marinum [197]. An experimental decrease of the IIS (by overexpression of foxo or by mutation in the InR substrate Chico) is sufficient to increase the transcription of several immune genes [200]. Thus, it seems that the increase in circulating sugars driven by the low IIS is sufficient to enhance Drosophila immunity. As a case in point, the glucose-based HSD increases glycemia and resistance to infection by the enteric pathogen Vibrio cholerae [21]. However, the sucrose-based HSD increases susceptibility to infection by Pseudomonas aeruginosa [68]. Thus, it remains to be determined whether these inconsistencies result from differential effects of dietary sugars (glucose [21] vs sucrose [68]), or rather reflect pathogen-specific effects of HSD.
6. Limitations of the Comparisons among Drosophila Studies
Comparisons of individual studies on obese flies is limited by the variability in the methods used to analyze and express the fat content (fat per fly, or normalized per body weight or per protein content), as well as by the differences in the diets used across laboratories. There is no clear definition of fly obesity yet; nevertheless, the normalization of fat content to the protein levels could fulfill a similar function as the BMI index in humans. The field of Drosophila research would also undoubtedly benefit from standardized diets, and from reporting the raw lipid and protein data in all obesity studies, which would allow meaningful comparisons and meta-analyses of individual papers.
7. Potential Roles of the Fat Body Dynamics in Longevity
Even though fat accumulation in Drosophila often correlates with longevity (e.g., [21,32,33,34,35,36,85]), experimental increase in the fat content via inhibition of the lipolytic pathways, for example, by mutations of bmm or Akh, moderately decreases lifespan ([134] and M.G. and P.K unpublished data [135]). Conversely, an increase in lipolytic AKH signaling extends lifespan [91,136], despite causing a lean phenotype [53,54]. Studies by Katewa and colleagues [90,91] argued that lifespan is extended by an increase in the fat turnover, i.e., by increase in both lipogenesis and lipolysis, irrespective of whether it is associated with a lean phenotype (as in the case of Akh overexpression [91]), or with obesity (as in the case of DR [90]). This hypothesis also explains the decrease in the lifespans of the bmm [134] and Akh mutants (M.G. and P.K., unpublished data [135]), as their defects in lipid mobilization probably reduce the global fat turnover rates.
However, how the DR-induced turnover is facilitated at the cellular level remains an open question. Is it specific for the triacylglycerides, or does it reflect a general increase in the turnover of all macromolecules, or even organelles? The longevity effects of the IIS and TOR downregulations are associated with increased autophagy-mediated turnover of organelles [201,202,203]. Importantly, as shown in mice, autophagy has an important role in lipid metabolism, promoting lipolysis and breakdown of lipid droplets [204]. Thus, increased fat turnover might be linked to longevity via the well-known lifespan-extending effects of autophagy [203].
The relation between obesity, aging, and the dynamics of the fat body at the cellular level remains enigmatic as well. In contrast to the larval fat body that consists of post-mitotic cells that grow solely via endocycling [205], the cellular biology of the adult fat body is not well understood. The number of fat body cells remains rather constant throughout life, while their size increases several fold within the first days after eclosion [206]. The fat body cells are susceptible to stress-induced apoptosis [207], can adapt to the increased adiposity by increasing the cellular proportion of lipid droplets [48,49], and possibly also by growth and division [206]. Nevertheless, how these multinucleate and polyploid cells proliferate, is not known, and the existence of fat body stem cells has not been reported to date. The mechanism whereby the lifespan-extending manipulations affect the physiology of this tissue remains to be investigated. Nevertheless, differential contribution of fat turnover, autophagy, cell proliferation and growth in different types of obesity might explain why certain types of obesity are coupled with lifespan shortening, whereas others are associated with longevity. In conclusion, the so far unknown cellular biology of the Drosophila fat body may hold the key to a better understanding of the interactions between fat storage, inflammatory responses, and lifespan.
Author Contributions
M.G. and P.K. wrote the review.
Funding
This research was funded by the Slovak granting agency [APVV-14-0556] and the Carl Tryggers Foundation.
Acknowledgments
We thank to Heinrich Dircksen and David Wildridge for helpful suggestions and English corrections.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| Akh | Adipokinetic hormone |
| BMI | body mass index |
| bmm | brummer |
| CCHa2 | CCHamide-2 |
| DR | dietary restriction |
| EcR | Ecdysone receptor |
| fit | female-specific independent of transformer |
| foxo | Forkhead Box, Sub-group O |
| HFD | high-fat diet |
| hh | hedgehog |
| HSD | high-sugar diet |
| IGF | Insulin-like growth factor |
| Ilp1–8 | Insulin-like peptide 1–8 |
| IIS | insulin/insulin-like growth factor signaling |
| IL-6 | interleukin 6 |
| ImpL2 | Ecdysone-inducible L2 |
| InR | Insulin-like receptor |
| JNK | Jun-N-terminal kinase pathway |
| mag | magro |
| MHO | metabolically healthy obesity |
| mth | methuselah |
| NPF | neuropeptide F |
| Tk | Tachykinin |
| TNF-α | tumor necrosis factor-α |
| to | takeout |
| TOR | target of rapamycin |
| upd2 | unpaired 2 |
| upd3 | unpaired 3 |
References
- World Health Organisation. Available online: http://www.who.int/mediacentre/factsheets/fs311/en/ (accessed on 9 May 2018).
- Flegal, K.M.; Kit, B.K.; Orpana, H.; Graubard, B.I. Association of All-Cause Mortality with Overweight and Obesity Using Standard Body Mass Index Categories a Systematic Review and Meta-analysis. JAMA 2013, 309, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Aune, D.; Sen, A.; Prasad, M.; Norat, T.; Janszky, I.; Tonstad, S.; Romundstad, P.; Vatten, L.J. BMI and all cause mortality: Systematic review and non-linear dose-response meta-analysis of 230 cohort studies with 3.74 million deaths among 30.3 million participants. BMJ 2016, 353, i2156. [Google Scholar] [CrossRef] [PubMed]
- Di Angelantonio, E.; Bhupathiraju, S.N.; Wormser, D.; Gao, P.; Kaptoge, S.; de Gonzalez, A.B.; Cairns, B.J.; Huxley, R.; Jackson, C.L.; Joshy, G.; et al. Body-mass index and all-cause mortality: Individual-participant-data meta-analysis of 239 prospective studies in four continents. Lancet 2016, 388, 776–786. [Google Scholar] [CrossRef]
- Angadi, S.S.; Gaesser, G.A. Body-mass index and all-cause mortality. Lancet 2017, 389, 2285. [Google Scholar] [CrossRef] [Green Version]
- Barry, V.W.; Baruth, M.; Beets, M.W.; Durstine, J.L.; Liu, J.H.; Blair, S.N. Fitness vs. Fatness on All-Cause Mortality: A Meta-Analysis. Prog. Cardiovasc. Dis. 2014, 56, 382–390. [Google Scholar] [CrossRef] [PubMed]
- Bluher, S.; Schwarz, P. Metabolically healthy obesity from childhood to adulthood—Does weight status alone matter? Metabolism 2014, 63, 1084–1092. [Google Scholar] [CrossRef] [PubMed]
- Kaur, A.; Johnston, D.G.; Godsland, I.F. Does metabolic health in overweight and obesity persist?—Individual variation and cardiovascular mortality over two decades. Eur. J. Endocrinol. 2016, 175, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Van Vliet-Ostaptchouk, J.V.; Nuotio, M.L.; Slagter, S.N.; Doiron, D.; Fischer, K.; Foco, L.; Gaye, A.; Gogele, M.; Heier, M.; Hiekkalinna, T.; et al. The prevalence of metabolic syndrome and metabolically healthy obesity in Europe: A collaborative analysis of ten large cohort studies. BMC Endocr. Disord. 2014, 14, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rey-Lopez, J.P.; de Rezende, L.F.; Pastor-Valero, M.; Tess, B.H. The prevalence of metabolically healthy obesity: A systematic review and critical evaluation of the definitions used. Obes. Rev. 2014, 15, 781–790. [Google Scholar] [CrossRef] [PubMed]
- Owusu-Ansah, E.; Perrimon, N. Modeling metabolic homeostasis and nutrient sensing in Drosophila: Implications for aging and metabolic diseases. Dis. Models Mech. 2014, 7, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Padmanabha, D.; Baker, K.D. Drosophila gains traction as a repurposed tool to investigate metabolism. Trends Endocrinol. Metab. 2014, 25, 518–527. [Google Scholar] [CrossRef] [PubMed]
- Musselman, L.P.; Kühnlein, R.P. Drosophila as a model to study obesity and metabolic disease. J. Exp. Biol. 2018, 221, jeb163881. [Google Scholar] [CrossRef] [PubMed]
- Bharucha, K.N. The Epicurean Fly: Using Drosophila Melanogaster to Study Metabolism. Pediatr. Res. 2009, 65, 132–137. [Google Scholar] [CrossRef] [PubMed]
- Diop, S.B.; Bodmer, R. Drosophila as a model to study the genetic mechanisms of obesity-associated heart dysfunction. J. Cell. Mol. Med. 2012, 16, 966–971. [Google Scholar] [CrossRef] [PubMed]
- Graham, P.; Pick, L. Drosophila as a Model for Diabetes and Diseases of Insulin Resistance. Curr. Top. Dev. Biol. 2017, 121, 397–419. [Google Scholar] [CrossRef] [PubMed]
- Murillo-Maldonado, J.M.; Riesgo-Escovar, J.R. Development and diabetes on the fly. Mech. Dev. 2017, 144, 150–155. [Google Scholar] [CrossRef] [PubMed]
- Herranz, H.; Cohen, S.M. Drosophila as a Model to Study the Link between Metabolism and Cancer. J. Dev. Biol. 2017, 5, 15. [Google Scholar] [CrossRef] [PubMed]
- Na, J.; Musselman, L.P.; Pendse, J.; Baranski, T.J.; Bodmer, R.; Ocorr, K.; Cagan, R. A Drosophila model of high sugar diet-induced cardiomyopathy. PLoS Genet. 2013, 9, e1003175. [Google Scholar] [CrossRef] [PubMed]
- Musselman, L.P.; Fink, J.L.; Narzinski, K.; Ramachandran, P.V.; Hathiramani, S.S.; Cagan, R.L.; Baranski, T.J. A high-sugar diet produces obesity and insulin resistance in wild-type Drosophila. Dis. Models Mech. 2011, 4, 842–849. [Google Scholar] [CrossRef] [PubMed]
- Galenza, A.; Hutchinson, J.; Campbell, S.D.; Hazes, B.; Foley, E. Glucose modulates Drosophila longevity and immunity independent of the microbiota. Biol. Open 2016, 5, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Pasco, M.Y.; Leopold, P. High sugar-induced insulin resistance in Drosophila relies on the lipocalin Neural Lazarillo. PLoS ONE 2012, 7, e36583. [Google Scholar] [CrossRef] [PubMed]
- Reis, T. Effects of Synthetic Diets Enriched in Specific Nutrients on Drosophila Development, Body Fat, and Lifespan. PLoS ONE 2016, 11, e0146758. [Google Scholar] [CrossRef] [PubMed]
- Morris, S.N.; Coogan, C.; Chamseddin, K.; Fernandez-Kim, S.O.; Kolli, S.; Keller, J.N.; Bauer, J.H. Development of diet-induced insulin resistance in adult Drosophila melanogaster. Biochim. Biophys. Acta 2012, 1822, 1230–1237. [Google Scholar] [CrossRef] [PubMed]
- Musselman, L.P.; Fink, J.L.; Ramachandran, P.V.; Patterson, B.W.; Okunade, A.L.; Maier, E.; Brent, M.R.; Turk, J.; Baranski, T.J. Role of fat body lipogenesis in protection against the effects of caloric overload in Drosophila. J. Biol. Chem. 2013, 288, 8028–8042. [Google Scholar] [CrossRef] [PubMed]
- Birse, R.T.; Choi, J.; Reardon, K.; Rodriguez, J.; Graham, S.; Diop, S.; Ocorr, K.; Bodmer, R.; Oldham, S. High-fat-diet-induced obesity and heart dysfunction are regulated by the TOR pathway in Drosophila. Cell Metab. 2010, 12, 533–544. [Google Scholar] [CrossRef] [PubMed]
- Heinrichsen, E.T.; Haddad, G.G. Role of high-fat diet in stress response of Drosophila. PLoS ONE 2012, 7, e42587. [Google Scholar] [CrossRef] [PubMed]
- Woodcock, K.J.; Kierdorf, K.; Pouchelon, C.A.; Vivancos, V.; Dionne, M.S.; Geissmann, F. Macrophage-derived upd3 cytokine causes impaired glucose homeostasis and reduced lifespan in Drosophila fed a lipid-rich diet. Immunity 2015, 42, 133–144. [Google Scholar] [CrossRef] [PubMed]
- Wen, D.T.; Zheng, L.; Yang, F.; Li, H.Z.; Chen, J. Endurance exercise prevents high-fat-diet induced locomotor impairment, cardiac dysfunction, lifespan shortening, and dSir2 expression decline in aging Drosophila. Exp. Gerontol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Baumbach, J.; Hummel, P.; Bickmeyer, I.; Kowalczyk, K.M.; Frank, M.; Knorr, K.; Hildebrandt, A.; Riedel, D.; Jäckle, H.; Kühnlein, R.P. A Drosophila in vivo screen identifies store-operated calcium entry as a key regulator of adiposity. Cell Metab. 2014, 19, 331–343. [Google Scholar] [CrossRef] [PubMed]
- Na, J.; Sweetwyne, M.T.; Park, A.S.; Susztak, K.; Cagan, R.L. Diet-Induced Podocyte Dysfunction in Drosophila and Mammals. Cell Rep. 2015, 12, 636–647. [Google Scholar] [CrossRef] [PubMed]
- Broughton, S.J.; Piper, M.D.; Ikeya, T.; Bass, T.M.; Jacobson, J.; Driege, Y.; Martinez, P.; Hafen, E.; Withers, D.J.; Leevers, S.J.; et al. Longer lifespan, altered metabolism, and stress resistance in Drosophila from ablation of cells making insulin-like ligands. Proc. Natl. Acad. Sci. USA 2005, 102, 3105–3110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grönke, S.; Clarke, D.F.; Broughton, S.; Andrews, T.D.; Partridge, L. Molecular evolution and functional characterization of Drosophila insulin-like peptides. PLoS Genet. 2010, 6, e1000857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bjedov, I.; Toivonen, J.M.; Kerr, F.; Slack, C.; Jacobson, J.; Foley, A.; Partridge, L. Mechanisms of life span extension by rapamycin in the fruit fly Drosophila melanogaster. Cell Metab. 2010, 11, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Emran, S.; Yang, M.; He, X.; Zandveld, J.; Piper, M.D. Target of rapamycin signalling mediates the lifespan-extending effects of dietary restriction by essential amino acid alteration. Aging 2014, 6, 390–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simmons, F.H.; Bradley, T.J. An analysis of resource allocation in response to dietary yeast in Drosophila melanogaster. J. Insect Physiol. 1997, 43, 779–788. [Google Scholar] [CrossRef]
- Kim, S.K.; Rulifson, E.J. Conserved mechanisms of glucose sensing and regulation by Drosophila corpora cardiaca cells. Nature 2004, 431, 316–320. [Google Scholar] [CrossRef] [PubMed]
- Becker, A.; Schloder, P.; Steele, J.E.; Wegener, G. The regulation of trehalose metabolism in insects. Experientia 1996, 52, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, H.; Yamada, T.; Yoshida, M.; Nishimura, T. Flies without Trehalose. J. Biol. Chem. 2015, 290, 1244–1255. [Google Scholar] [CrossRef] [PubMed]
- Shukla, E.; Thorat, L.J.; Nath, B.B.; Gaikwad, S.M. Insect trehalase: Physiological significance and potential applications. Glycobiology 2015, 25, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Oliver, M.J.; Huber, R.E.; Williamson, J.H. Genetic and Biochemical Aspects of Trehalase from Drosophila-Melanogaster. Biochem. Genet. 1978, 16, 927–940. [Google Scholar] [CrossRef] [PubMed]
- Yasugi, T.; Yamada, T.; Nishimura, T. Adaptation to dietary conditions by trehalose metabolism in Drosophila. Sci. Rep. 2017, 7, 1619. [Google Scholar] [CrossRef] [PubMed]
- Gáliková, M.; Diesner, M.; Klepsatel, P.; Hehlert, P.; Xu, Y.J.; Bickmeyer, I.; Predel, R.; Kühnlein, R.P. Energy Homeostasis Control in Drosophila Adipokinetic Hormone Mutants. Genetics 2015, 201, 665–683. [Google Scholar] [CrossRef] [PubMed]
- Maniere, G.; Ziegler, A.B.; Geillon, F.; Featherstone, D.E.; Grosjean, Y. Direct Sensing of Nutrients via a LAT1-like Transporter in Drosophila Insulin-Producing Cells. Cell Rep. 2016, 17, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Campos, C. Chronic Hyperglycemia and Glucose Toxicity: Pathology and Clinical Sequelae. Postgrad. Med. 2012, 124, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Wigglesworth, V.B. The Utilization of Reserve Substances in Drosophila during Flight. J. Exp. Biol. 1949, 26, 150–163. [Google Scholar] [PubMed]
- Yamada, T.; Habara, O.; Kubo, H.; Nishimura, T. Fat body glycogen serves as a metabolic safeguard for the maintenance of sugar levels in Drosophila. Development 2018, 145. [Google Scholar] [CrossRef] [PubMed]
- Beller, M.; Thiel, K.; Thul, P.J.; Jäckle, H. Lipid droplets: A dynamic organelle moves into focus. FEBS Lett. 2010, 584, 2176–2182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kühnlein, R.P. Thematic Review Series: Lipid Droplet Synthesis and Metabolism: From Yeast to Man Lipid droplet-based storage fat metabolism in Drosophila. J. Lipid Res. 2012, 53, 1430–1436. [Google Scholar] [CrossRef] [PubMed]
- Saltiel, A.R. Insulin Signaling in the Control of Glucose and Lipid Homeostasis. Handb. Exp. Pharmacol. 2016, 233, 51–71. [Google Scholar] [CrossRef] [PubMed]
- Rui, L. Energy metabolism in the liver. Compr. Physiol. 2014, 4, 177–197. [Google Scholar] [CrossRef] [PubMed]
- Rulifson, E.J.; Kim, S.K.; Nusse, R. Ablation of insulin-producing neurons in flies: Growth and diabetic phenotypes. Science 2002, 296, 1118–1120. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.; Park, J.H. Hemolymph sugar homeostasis and starvation-induced hyperactivity affected by genetic manipulations of the adipokinetic hormone-encoding gene in Drosophila melanogaster. Genetics 2004, 167, 311–323. [Google Scholar] [CrossRef] [PubMed]
- Gáliková, M.; Klepsatel, P.; Xu, Y.J.; Kühnlein, R.P. The obesity-related Adipokinetic hormone controls feeding and expression of neuropeptide regulators of Drosophila metabolism. Eur. J. Lipid Sci. Technol. 2017, 119. [Google Scholar] [CrossRef]
- Brogiolo, W.; Stocker, H.; Ikeya, T.; Rintelen, F.; Fernandez, R.; Hafen, E. An evolutionarily conserved function of the Drosophila insulin receptor and insulin-like peptides in growth control. Curr. Biol. 2001, 11, 213–221. [Google Scholar] [CrossRef]
- Nassel, D.R.; Liu, Y.; Luo, J. Insulin/IGF signaling and its regulation in Drosophila. Gen. Comp. Endocrinol. 2015, 221, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Mattila, J.; Hietakangas, V. Regulation of Carbohydrate Energy Metabolism in Drosophila melanogaster. Genetics 2017, 207, 1231–1253. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Neufeld, T.P. Dietary sugar promotes systemic TOR activation in Drosophila through AKH-dependent selective secretion of Dilp3. Nat. Commun. 2015, 6, 6846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajan, A.; Perrimon, N. Drosophila Cytokine Unpaired 2 Regulates Physiological Homeostasis by Remotely Controlling Insulin Secretion. Cell 2012, 151, 123–137. [Google Scholar] [CrossRef] [PubMed]
- Sano, H.; Nakamura, A.; Texada, M.J.; Truman, J.W.; Ishimoto, H.; Kamikouchi, A.; Nibu, Y.; Kume, K.; Ida, T.; Kojima, M. The Nutrient-Responsive Hormone CCHamide-2 Controls Growth by Regulating Insulin-like Peptides in the Brain of Drosophila melanogaster. PLoS Genet. 2015, 11, e1005209. [Google Scholar] [CrossRef]
- Kreneisz, O.; Chen, X.; Fridell, Y.W.; Mulkey, D.K. Glucose increases activity and Ca2+ in insulin-producing cells of adult Drosophila. Neuroreport 2010, 21, 1116–1120. [Google Scholar] [CrossRef] [PubMed]
- DiAngelo, J.R.; Birnbaum, M.J. Regulation of fat cell mass by insulin in Drosophila melanogaster. Mol. Cell. Biol. 2009, 29, 6341–6352. [Google Scholar] [CrossRef] [PubMed]
- Arquier, N.; Geminard, C.; Bourouis, M.; Jarretou, G.; Honegger, B.; Paix, A.; Leopold, P. Drosophila ALS regulates growth and metabolism through functional interaction with insulin-like peptides. Cell Metab. 2008, 7, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Honegger, B.; Galic, M.; Kohler, K.; Wittwer, F.; Brogiolo, W.; Hafen, E.; Stocker, H. Imp-L2, a putative homolog of vertebrate IGF-binding protein 7, counteracts insulin signaling in Drosophila and is essential for starvation resistance. J. Biol. 2008, 7, 10. [Google Scholar] [CrossRef] [PubMed]
- Alic, N.; Hoddinott, M.P.; Vinti, G.; Partridge, L. Lifespan extension by increased expression of the Drosophila homologue of the IGFBP7 tumour suppressor. Aging Cell 2011, 10, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Tennessen, J.M.; Barry, W.E.; Cox, J.; Thummel, C.S. Methods for studying metabolism in Drosophila. Methods 2014, 68, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Chng, W.A.; Hietakangas, V.; Lemaitre, B. Physiological Adaptations to Sugar Intake: New Paradigms from Drosophila melanogaster. Trends Endocrinol. Metab. 2017, 28, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Musselman, L.P.; Fink, J.L.; Grant, A.R.; Gatto, J.A.; Tuthill, B.F.; Baranski, T.J. A Complex Relationship between Immunity and Metabolism in Drosophila Diet-Induced Insulin Resistance. Mol. Cell. Biol. 2018, 38. [Google Scholar] [CrossRef] [PubMed]
- Na, J.B.; Cagan, R. The Drosophila Nephrocyte: Back on Stage. J. Am. Soc. Nephrol. 2013, 24, 161–163. [Google Scholar] [CrossRef] [PubMed]
- Rovenko, B.M.; Perkhulyn, N.V.; Gospodaryov, D.V.; Sanz, A.; Lushchak, O.V.; Lushchak, V.I. High consumption of fructose rather than glucose promotes a diet-induced obese phenotype in Drosophila melanogaster. Comp. Biochem. Physiol. A 2015, 180, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Skorupa, D.A.; Dervisefendic, A.; Zwiener, J.; Pletcher, S.D. Dietary composition specifies consumption, obesity, and lifespan in Drosophila melanogaster. Aging Cell 2008, 7, 478–490. [Google Scholar] [CrossRef] [PubMed]
- Lushchak, O.V.; Gospodaryov, D.V.; Rovenko, B.M.; Yurkevych, I.S.; Perkhulyn, N.V.; Lushchak, V.I. Specific dietary carbohydrates differentially influence the life span and fecundity of Drosophila melanogaster. J. Gerontol. A Biol. Sci. Med. Sci. 2014, 69, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Brookheart, R.T.; Swearingen, A.R.; Collins, C.A.; Cline, L.M.; Duncan, J.G. High-sucrose-induced maternal obesity disrupts ovarian function and decreases fertility in Drosophila melanogaster. Biochim. Biophys. Acta 2017, 1863, 1255–1263. [Google Scholar] [CrossRef] [PubMed]
- Dobson, A.J.; Ezcurra, M.; Flanagan, C.E.; Summerfield, A.C.; Piper, M.D.; Gems, D.; Alic, N. Nutritional Programming of Lifespan by FOXO Inhibition on Sugar-Rich Diets. Cell Rep. 2017, 18, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Jin, Q.; Jin, L.H. High sugar diet disrupts gut homeostasis though JNK and STAT pathways in Drosophila. Biochem. Biophys. Res. Commun. 2017, 487, 910–916. [Google Scholar] [CrossRef] [PubMed]
- Hemphill, W.; Rivera, O.; Talbert, M. RNA-Sequencing of Drosophila melanogaster Head Tissue on High-Sugar and High-Fat Diets. G3 (Bethesda) 2018, 8, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Kapahi, P.; Kaeberlein, M.; Hansen, M. Dietary restriction and lifespan: Lessons from invertebrate models. Ageing Res. Rev. 2017, 39, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Fontana, L.; Partridge, L. Promoting Health and Longevity through Diet: From Model Organisms to Humans. Cell 2015, 161, 106–118. [Google Scholar] [CrossRef] [PubMed]
- Fontana, L.; Partridge, L.; Longo, V.D. Extending Healthy Life Span-From Yeast to Humans. Science 2010, 328, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Simpson, S.J.; Raubenheimer, D. Obesity: The protein leverage hypothesis. Obes. Rev. 2005, 6, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Le Couteur, D.G.; Solon-Biet, S.; Cogger, V.C.; Mitchell, S.J.; Senior, A.; de Cabo, R.; Raubenheimer, D.; Simpson, S.J. The impact of low-protein high-carbohydrate diets on aging and lifespan. Cell. Mol. Life Sci. 2016, 73, 1237–1252. [Google Scholar] [CrossRef] [PubMed]
- Tatar, M.; Post, S.; Yu, K. Nutrient control of Drosophila longevity. Trends Endocrinol. Metab. 2014, 25, 509–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bekelman, T.A.; Santamaria-Ulloa, C.; Dufour, D.L.; Marin-Arias, L.; Dengo, A.L. Using the protein leverage hypothesis to understand socioeconomic variation in obesity. Am. J. Hum. Biol. 2017, 29. [Google Scholar] [CrossRef] [PubMed]
- Gosby, A.K.; Conigrave, A.D.; Raubenheimer, D.; Simpson, S.J. Protein leverage and energy intake. Obes. Rev. 2014, 15, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Mair, W.; Piper, M.D.; Partridge, L. Calories do not explain extension of life span by dietary restriction in Drosophila. PLoS Biol. 2005, 3, e223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.P. Dietary protein:carbohydrate balance is a critical modulator of lifespan and reproduction in Drosophila melanogaster: A test using a chemically defined diet. J. Insect Physiol. 2015, 75, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.P.; Simpson, S.J.; Clissold, F.J.; Brooks, R.; Ballard, J.W.; Taylor, P.W.; Soran, N.; Raubenheimer, D. Lifespan and reproduction in Drosophila: New insights from nutritional geometry. Proc. Natl. Acad. Sci. USA 2008, 105, 2498–2503. [Google Scholar] [CrossRef] [PubMed]
- Solon-Biet, S.M.; McMahon, A.C.; Ballard, J.W.; Ruohonen, K.; Wu, L.E.; Cogger, V.C.; Warren, A.; Huang, X.; Pichaud, N.; Melvin, R.G.; et al. The ratio of macronutrients, not caloric intake, dictates cardiometabolic health, aging, and longevity in ad libitum-fed mice. Cell Metab. 2014, 19, 418–430. [Google Scholar] [CrossRef] [PubMed]
- Min, K.J.; Yamamoto, R.; Buch, S.; Pankratz, M.; Tatar, M. Drosophila lifespan control by dietary restriction independent of insulin-like signaling. Aging Cell 2008, 7, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Katewa, S.D.; Akagi, K.; Bose, N.; Rakshit, K.; Camarella, T.; Zheng, X.Z.; Hall, D.; Davis, S.; Nelson, C.S.; Brem, R.B.; et al. Peripheral Circadian Clocks Mediate Dietary Restriction-Dependent Changes in Lifespan and Fat Metabolism in Drosophila. Cell Metab. 2016, 23, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Katewa, S.D.; Demontis, F.; Kolipinski, M.; Hubbard, A.; Gill, M.S.; Perrimon, N.; Melov, S.; Kapahi, P. Intramyocellular fatty-acid metabolism plays a critical role in mediating responses to dietary restriction in Drosophila melanogaster. Cell Metab. 2012, 16, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.H.; Kang, M.; Lee, K.S.; Yu, K. High fat diet-induced TGF-beta/Gbb signaling provokes insulin resistance through the tribbles expression. Sci. Rep. 2016, 6, 30265. [Google Scholar] [CrossRef] [PubMed]
- Diop, S.B.; Bisharat-Kernizan, J.; Birse, R.T.; Oldham, S.; Ocorr, K.; Bodmer, R. PGC-1/Spargel Counteracts High-Fat-Diet-Induced Obesity and Cardiac Lipotoxicity Downstream of TOR and Brummer ATGL Lipase. Cell Rep. 2015, 10, 1572–1584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schultzhaus, J.N.; Bennett, C.J.; Iftikhar, H.; Yew, J.Y.; Mallett, J.; Carney, G.E. High fat diet alters Drosophila melanogaster sexual behavior and traits: Decreased attractiveness and changes in pheromone profiles. Sci. Rep. 2018, 8, 5387. [Google Scholar] [CrossRef] [PubMed]
- Kayashima, Y.; Murata, S.; Sato, M.; Matsuura, K.; Asanuma, T.; Chimoto, J.; Ishii, T.; Mochizuki, K.; Kumazawa, S.; Nakayama, T.; et al. Tea polyphenols ameliorate fat storage induced by high-fat diet in Drosophila melanogaster. Biochem. Biophys. Rep. 2015, 4, 417–424. [Google Scholar] [CrossRef] [PubMed]
- Pulinilkunnil, T.; Kienesberger, P.C.; Nagendran, J.; Sharma, N.; Young, M.E.; Dyck, J.R. Cardiac-specific adipose triglyceride lipase overexpression protects from cardiac steatosis and dilated cardiomyopathy following diet-induced obesity. Int. J. Obes. (Lond.) 2014, 38, 205–215. [Google Scholar] [CrossRef] [PubMed]
- Pulinilkunnil, T.; Kienesberger, P.C.; Nagendran, J.; Waller, T.J.; Young, M.E.; Kershaw, E.E.; Korbutt, G.; Haemmerle, G.; Zechner, R.; Dyck, J.R. Myocardial adipose triglyceride lipase overexpression protects diabetic mice from the development of lipotoxic cardiomyopathy. Diabetes 2013, 62, 1464–1477. [Google Scholar] [CrossRef] [PubMed]
- Kienesberger, P.C.; Pulinilkunnil, T.; Sung, M.M.; Nagendran, J.; Haemmerle, G.; Kershaw, E.E.; Young, M.E.; Light, P.E.; Oudit, G.Y.; Zechner, R.; et al. Myocardial ATGL overexpression decreases the reliance on fatty acid oxidation and protects against pressure overload-induced cardiac dysfunction. Mol. Cell. Biol. 2012, 32, 740–750. [Google Scholar] [CrossRef] [PubMed]
- Hotamisligil, G.S. Inflammation and metabolic disorders. Nature 2006, 444, 860–867. [Google Scholar] [CrossRef] [PubMed]
- Saltiel, A.R.; Olefsky, J.M. Inflammatory mechanisms linking obesity and metabolic disease. J. Clin. Investig. 2017, 127, 1–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, M.; Flatt, T.; Aguilaniu, H. Reproduction, Fat Metabolism, and Life Span: What Is the Connection? Cell Metab. 2013, 17, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Salmon, A.B.; Marx, D.B.; Harshman, L.G. A cost of reproduction in Drosophila melanogaster: Stress susceptibility. Evolution 2001, 55, 1600–1608. [Google Scholar] [CrossRef] [PubMed]
- Doane, W.W. Developmental Physiology of the Mutant Female Sterile(2)Adipose of Drosophila-Melanogaster. 1. Adult Morphology, Longevity, Egg Production, and Egg Lethality. J. Exp. Zool. 1960, 145, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Rush, B.; Sandver, S.; Bruer, J.; Roche, R.; Wells, M.; Giebultowicz, J. Mating increases starvation resistance and decreases oxidative stress resistance in Drosophila melanogaster females. Aging Cell 2007, 6, 723–726. [Google Scholar] [CrossRef] [PubMed]
- Flatt, T. Survival costs of reproduction in Drosophila. Exp. Gerontol. 2011, 46, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Flatt, T.; Min, K.J.; D’Alterio, C.; Villa-Cuesta, E.; Cumbers, J.; Lehmann, R.; Jones, D.L.; Tatar, M. Drosophila germ-line modulation of insulin signaling and lifespan. Proc. Natl. Acad. Sci. USA 2008, 105, 6368–6373. [Google Scholar] [CrossRef] [PubMed]
- Rubenstein, E.C.; Kelly, T.J.; Schwartz, M.B.; Woods, C.W. In vitro synthesis and secretion of ecdysteroids by Drosophila melanogaster ovaries. J. Exp. Zool. 1982, 223, 305–308. [Google Scholar] [CrossRef]
- Hentze, J.L.; Moeller, M.E.; Jorgensen, A.F.; Bengtsson, M.S.; Bordoy, A.M.; Warren, J.T.; Gilbert, L.I.; Andersen, O.; Rewitz, K.F. Accessory Gland as a Site for Prothoracicotropic Hormone Controlled Ecdysone Synthesis in Adult Male Insects. PLoS ONE 2013, 8, e55131. [Google Scholar] [CrossRef] [PubMed]
- Gáliková, M.; Klepsatel, P.; Senti, G.; Flatt, T. Steroid hormone regulation of C. elegans and Drosophila aging and life history. Exp. Gerontol. 2011, 46, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Simon, A.F.; Shih, C.; Mack, A.; Benzer, S. Steroid control of longevity in Drosophila melanogaster. Science 2003, 299, 1407–1410. [Google Scholar] [CrossRef] [PubMed]
- Tricoire, H.; Battisti, V.; Trannoy, S.; Lasbleiz, C.; Pret, A.M.; Monnier, V. The steroid hormone receptor EcR finely modulates Drosophila lifespan during adulthood in a sex-specific manner. Mech. Ageing Dev. 2009, 130, 547–552. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, N.; Rewitz, K.F.; O’Connor, M.B. Ecdysone Control of Developmental Transitions: Lessons from Drosophila Research. Annu. Rev. Entomol. 2013, 58, 497–516. [Google Scholar] [CrossRef] [PubMed]
- Ables, E.T.; Drummond-Barbosa, D. The Steroid Hormone Ecdysone Functions with Intrinsic Chromatin Remodeling Factors to Control Female Germline Stem Cells in Drosophila. Cell Stem Cell 2010, 7, 581–592. [Google Scholar] [CrossRef] [PubMed]
- Buszczak, M.; Freeman, M.R.; Carlson, J.R.; Bender, M.; Cooley, L.; Segraves, W.A. Ecdysone response genes govern egg chamber development during mid-oogenesis in Drosophila. Development 1999, 126, 4581–4589. [Google Scholar] [PubMed]
- Hackney, J.F.; Pucci, C.; Naes, E.; Dobens, L. Ras signaling modulates activity of the ecdysone receptor EcR during cell migration in the Drosophila ovary. Dev. Dyn. 2007, 236, 1213–1226. [Google Scholar] [CrossRef] [PubMed]
- Kamoshida, Y.; Fujiyama-Nakamura, S.; Kimura, S.; Suzuki, E.; Lim, J.; Shiozaki-Sato, Y.; Kato, S.; Takeyama, K. Ecdysone receptor (EcR) suppresses lipid accumulation in the Drosophila fat body via transcription control. Biochem. Biophys. Res. Commun. 2012, 421, 203–207. [Google Scholar] [CrossRef] [PubMed]
- Mauvais-Jarvis, F. Estrogen and androgen receptors: Regulators of fuel homeostasis and emerging targets for diabetes and obesity. Trends Endocrinol. Metab. 2011, 22, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, E.J.; Gianatti, E.; Strauss, B.J.; Wentworth, J.; Lim-Joon, D.; Bolton, D.; Zajac, J.D.; Grossmann, M. Increase in visceral and subcutaneous abdominal fat in men with prostate cancer treated with androgen deprivation therapy. Clin. Endocrinol. 2011, 74, 377–383. [Google Scholar] [CrossRef] [PubMed]
- Lizcano, F.; Guzman, G. Estrogen Deficiency and the Origin of Obesity during Menopause. BioMed Res. Int. 2014, 2014, 757461. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.J.; Seroude, L.; Benzer, S. Extended life-span and stress resistance in the Drosophila mutant methuselah. Science 1998, 282, 943–946. [Google Scholar] [CrossRef] [PubMed]
- Djawdan, M.; Sugiyama, T.T.; Schlaeger, L.K.; Bradley, T.J.; Rose, M.R. Metabolic aspects of the trade-off between fecundity and longevity in Drosophila melanogaster. Physiol. Zool. 1996, 69, 1176–1195. [Google Scholar] [CrossRef]
- Chippindale, A.K.; Leroi, A.M.; Kim, S.B.; Rose, M.R. Phenotypic Plasticity and Selection in Drosophila Life-History Evolution. 1. Nutrition and the Cost of Reproduction. J. Evol. Biol. 1993, 6, 171–193. [Google Scholar] [CrossRef]
- Baldal, E.A.; Brakefield, P.M.; Zwaan, B.J. Multitrait evolution in lines of Drosophila melanogaster selected for increased starvation resistance: The role of metabolic rate and implications for the evolution of longevity. Evolution 2006, 60, 1435–1444. [Google Scholar] [CrossRef] [PubMed]
- Harshman, L.G.; Hoffmann, A.A.; Clark, A.G. Selection for starvation resistance in Drosophila melanogaster: Physiological correlates, enzyme activities and multiple stress responses. J. Evol. Biol. 1999, 12, 370–379. [Google Scholar] [CrossRef]
- Zwaan, B.; Bijlsma, R.; Hoekstra, R.E. Direct Selection on Life-Span in Drosophila Melanogaster. Evolution 1995, 49, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Moghadam, N.N.; Holmstrup, M.; Manenti, T.; Mouridsen, M.B.; Pertoldi, C.; Loeschcke, V. The Role of Storage Lipids in the Relation between Fecundity, Locomotor Activity, and Lifespan of Drosophila melanogaster Longevity-Selected and Control Lines. PLoS ONE 2015, 10, e0130334. [Google Scholar] [CrossRef]
- Jumbo-Lucioni, P.; Ayroles, J.F.; Chambers, M.M.; Jordan, K.W.; Leips, J.; Mackay, T.F.; De Luca, M. Systems genetics analysis of body weight and energy metabolism traits in Drosophila melanogaster. BMC Genom. 2010, 11, 297. [Google Scholar] [CrossRef] [PubMed]
- Nelson, C.S.; Beck, J.N.; Wilson, K.A.; Pilcher, E.R.; Kapahi, P.; Brem, R.B. Cross-phenotype association tests uncover genes mediating nutrient response in Drosophila. BMC Genom. 2016, 17, 867. [Google Scholar] [CrossRef] [PubMed]
- Unckless, R.L.; Rottschaefer, S.M.; Lazzaro, B.P. A genome-wide association study for nutritional indices in Drosophila. G3 (Bethesda) 2015, 5, 417–425. [Google Scholar] [CrossRef] [PubMed]
- Jehrke, L.; Stewart, F.A.; Droste, A.; Beller, M. The impact of genome variation and diet on the metabolic phenotype and microbiome composition of Drosophila melanogaster. Sci. Rep. 2018, 8, 6215. [Google Scholar] [CrossRef] [PubMed]
- Reis, T.; Van Gilst, M.R.; Hariharan, I.K. A buoyancy-based screen of Drosophila larvae for fat-storage mutants reveals a role for Sir2 in coupling fat storage to nutrient availability. PLoS Genet. 2010, 6, e1001206. [Google Scholar] [CrossRef] [PubMed]
- Pospisilik, J.A.; Schramek, D.; Schnidar, H.; Cronin, S.J.F.; Nehme, N.T.; Zhang, X.Y.; Knauf, C.; Cani, P.D.; Aumayr, K.; Todoric, J.; et al. Drosophila Genome-wide Obesity Screen Reveals Hedgehog as a Determinant of Brown versus White Adipose Cell Fate. Cell 2010, 140, 148–160. [Google Scholar] [CrossRef] [PubMed]
- Grönke, S.; Muller, G.; Hirsch, J.; Fellert, S.; Andreou, A.; Haase, T.; Jäckle, H.; Kühnlein, R.P. Dual lipolytic control of body fat storage and mobilization in Drosophila. PLoS Biol. 2007, 5, e137. [Google Scholar] [CrossRef] [PubMed]
- Grönke, S.; Mildner, A.; Fellert, S.; Tennagels, N.; Petry, S.; Muller, G.; Jäckle, H.; Kühnlein, R.P. Brummer lipase is an evolutionary conserved fat storage regulator in Drosophila. Cell Metab. 2005, 1, 323–330. [Google Scholar] [CrossRef] [PubMed]
- Gáliková, M.; (Stockholm University, Stockholm, Sweden); Klepsatel, P.; (Institute of Zoology, Bratislava, Slovakia). Regulation of lifespan by Adipokinetic hormone. 2018. [Google Scholar]
- Waterson, M.J.; Chung, B.Y.; Harvanek, Z.M.; Ostojic, I.; Alcedo, J.; Pletcher, S.D. Water sensor ppk28 modulates Drosophila lifespan and physiology through AKH signaling. Proc. Natl. Acad. Sci. USA 2014, 111, 8137–8142. [Google Scholar] [CrossRef] [PubMed]
- Birner-Gruenberger, R.; Bickmeyer, I.; Lange, J.; Hehlert, P.; Hermetter, A.; Kollroser, M.; Rechberger, G.N.; Kühnlein, R.P. Functional fat body proteomics and gene targeting reveal in vivo functions of Drosophila melanogaster alpha-Esterase-7. Insect Biochem. Mol. Biol. 2012, 42, 220–229. [Google Scholar] [CrossRef] [PubMed]
- Bai, H.; Kang, P.; Tatar, M. Drosophila insulin-like peptide-6 (dilp6) expression from fat body extends lifespan and represses secretion of Drosophila insulin-like peptide-2 from the brain. Aging Cell 2012, 11, 978–985. [Google Scholar] [CrossRef] [PubMed]
- Droujinine, I.A.; Perrimon, N. Interorgan Communication Pathways in Physiology: Focus on Drosophila. Annu. Rev. Genet. 2016, 50, 539–570. [Google Scholar] [CrossRef] [PubMed]
- Nassel, D.R.; Vanden Broeck, J. Insulin/IGF signaling in Drosophila and other insects: Factors that regulate production, release and post-release action of the insulin-like peptides. Cell. Mol. Life Sci. 2016, 73, 271–290. [Google Scholar] [CrossRef] [PubMed]
- Andersen, D.S.; Colombani, J.; Leopold, P. Coordination of organ growth: Principles and outstanding questions from the world of insects. Trends Cell Biol. 2013, 23, 336–344. [Google Scholar] [CrossRef] [PubMed]
- Rajan, A.; Perrimon, N. Drosophila as a Model for Interorgan Communication: Lessons from Studies on Energy Homeostasis. Dev. Cell 2011, 21, 29–31. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Jin, L.H. Organ-to-Organ Communication: A Drosophila Gastrointestinal Tract Perspective. Front. Cell Dev. Biol. 2017, 5, 29. [Google Scholar] [CrossRef] [PubMed]
- Galic, S.; Oakhill, J.S.; Steinberg, G.R. Adipose tissue as an endocrine organ. Mol. Cell. Endocrinol. 2010, 316, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Kahn, S.E.; Cooper, M.E.; Del Prato, S. Pathophysiology and treatment of type 2 diabetes: Perspectives on the past, present, and future. Lancet 2014, 383, 1068–1083. [Google Scholar] [CrossRef]
- Broughton, S.J.; Slack, C.; Alic, N.; Metaxakis, A.; Bass, T.M.; Driege, Y.; Partridge, L. DILP-producing median neurosecretory cells in the Drosophila brain mediate the response of lifespan to nutrition. Aging Cell 2010, 9, 336–346. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Cheng, D.; Hong, S.; Sappe, B.; Hu, Y.; Wei, N.; Zhu, C.; O’Connor, M.B.; Pissios, P.; Perrimon, N. Midgut-Derived Activin Regulates Glucagon-like Action in the Fat Body and Glycemic Control. Cell Metab. 2017, 25, 386–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arrese, E.L.; Soulages, J.L. Insect Fat Body: Energy, Metabolism, and Regulation. Annu. Rev. Entomol. 2010, 55, 207–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sano, H. Coupling of growth to nutritional status: The role of novel periphery-to-brain signaling by the CCHa2 peptide in Drosophila melanogaster. Fly 2015, 9, 183–187. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.H.; Liu, C.; Bai, X.B.; Li, X.T.; Li, J.Y.; Zhang, Z.P.; Zhang, Y.P.; Guo, J.; Li, Y. Drosophila FIT is a protein-specific satiety hormone essential for feeding control. Nat. Commun. 2017, 8, 14161. [Google Scholar] [CrossRef] [PubMed]
- Chng, W.A.; Sleiman, M.S.B.; Schupfer, F.; Lemaitre, B. Transforming growth factor beta/activin signaling functions as a sugar-sensing feedback loop to regulate digestive enzyme expression. Cell Rep. 2014, 9, 336–348. [Google Scholar] [CrossRef] [PubMed]
- Rodenfels, J.; Lavrynenko, O.; Ayciriex, S.; Sampaio, J.L.; Carvalho, M.; Shevchenko, A.; Eaton, S. Production of systemically circulating Hedgehog by the intestine couples nutrition to growth and development. Genes Dev. 2014, 28, 2636–2651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dashti, M.; Peppelenbosch, M.P.; Rezaee, F. Hedgehog signalling as an antagonist of ageing and its associated diseases. BioEssays 2012, 34, 849–856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwangbo, D.S.; Gershman, B.; Tu, M.P.; Palmer, M.; Tatar, M. Drosophila dFOXO controls lifespan and regulates insulin signalling in brain and fat body. Nature 2004, 429, 562–566. [Google Scholar] [CrossRef] [PubMed]
- Kapahi, P.; Zid, B.M.; Harper, T.; Koslover, D.; Sapin, V.; Benzer, S. Regulation of lifespan in Drosophila by modulation of genes in the TOR signaling pathway. Curr. Biol. 2004, 14, 885–890. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Bassel-Duby, R.; Olson, E.N. Heart- and muscle-derived signaling system dependent on MED13 and Wingless controls obesity in Drosophila. Proc. Natl. Acad. Sci. USA 2014, 111, 9491–9496. [Google Scholar] [CrossRef] [PubMed]
- Grueter, C.E.; van Rooij, E.; Johnson, B.A.; DeLeon, S.M.; Sutherland, L.B.; Qi, X.; Gautron, L.; Elmquist, J.K.; Bassel-Duby, R.; Olson, E.N. A cardiac microRNA governs systemic energy homeostasis by regulation of MED13. Cell 2012, 149, 671–683. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Bao, H.; Ishikawa, Z.; Wang, W.; Lim, H.Y. Cardiomyocyte Regulation of Systemic Lipid Metabolism by the Apolipoprotein B-Containing Lipoproteins in Drosophila. PLoS Genet. 2017, 13, e1006555. [Google Scholar] [CrossRef] [PubMed]
- Makki, K.; Froguel, P.; Wolowczuk, I. Adipose tissue in obesity-related inflammation and insulin resistance: Cells, cytokines, and chemokines. ISRN Inflamm. 2013, 2013, 139239. [Google Scholar] [CrossRef] [PubMed]
- Lim, A.K.H.; Tesch, G.H. Inflammation in Diabetic Nephropathy. Mediat. Inflamm. 2012. [Google Scholar] [CrossRef] [PubMed]
- Wada, J.; Makino, H. Inflammation and the pathogenesis of diabetic nephropathy. Clin. Sci. 2013, 124, 139–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maezawa, Y.; Takemoto, M.; Yokote, K. Cell biology of diabetic nephropathy: Roles of endothelial cells, tubulointerstitial cells and podocytes. J. Diabetes Investig. 2015, 6, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.S.; Susztak, K. Podocytes: The Weakest Link in Diabetic Kidney Disease? Curr. Diabetes Rep. 2016, 16, 45. [Google Scholar] [CrossRef] [PubMed]
- Weavers, H.; Prieto-Sanchez, S.; Grawe, F.; Garcia-Lopez, A.; Artero, R.; Wilsch-Brauninger, M.; Ruiz-Gomez, M.; Skaer, H.; Denholm, B. The insect nephrocyte is a podocyte-like cell with a filtration slit diaphragm. Nature 2009, 457, 322–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helmstadter, M.; Huber, T.B.; Hermle, T. Using the Drosophila Nephrocyte to Model Podocyte Function and Disease. Front. Pediatr. 2017, 5, 262. [Google Scholar] [CrossRef] [PubMed]
- Lemaitre, B.; Miguel-Aliaga, I. The digestive tract of Drosophila melanogaster. Annu. Rev. Genet. 2013, 47, 377–404. [Google Scholar] [CrossRef] [PubMed]
- Chung, B.Y.; Ro, J.; Hutter, S.A.; Miller, K.M.; Guduguntla, L.S.; Kondo, S.; Pletcher, S.D. Drosophila Neuropeptide F Signaling Independently Regulates Feeding and Sleep-Wake Behavior. Cell Rep. 2017, 19, 2441–2450. [Google Scholar] [CrossRef] [PubMed]
- Ren, G.R.; Hauser, F.; Rewitz, K.F.; Kondo, S.; Engelbrecht, A.F.; Didriksen, A.K.; Schjott, S.R.; Sembach, F.E.; Li, S.; Sogaard, K.C.; et al. CCHamide-2 Is an Orexigenic Brain-Gut Peptide in Drosophila. PLoS ONE 2015, 10, e0133017. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Veenstra, J.A.; Perrimon, N. Control of lipid metabolism by tachykinin in Drosophila. Cell Rep. 2014, 9, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Meunier, N.; Belgacem, Y.H.; Martin, J.-R. Regulation of feeding behaviour and locomotor activity by takeout in Drosophila. J. Exp. Biol. 2007, 210, 1424–1434. [Google Scholar] [CrossRef] [PubMed]
- Sarov-Blat, L.; So, W.V.; Liu, L.; Rosbash, M. The Drosophila takeout gene is a novel molecular link between circadian rhythms and feeding behavior. Cell 2000, 101, 647–656. [Google Scholar] [CrossRef]
- Bauer, J.; Antosh, M.; Chang, C.; Schorl, C.; Kolli, S.; Neretti, N.; Helfand, S.L. Comparative transcriptional profiling identifies takeout as a gene that regulates life span. Aging 2010, 2, 298–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gáliková, M.; Flatt, T. Dietary restriction and other lifespan extending pathways converge at the activation of the downstream effector takeout. Aging 2010, 2, 387–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rera, M.; Azizi, M.J.; Walker, D.W. Organ-specific mediation of lifespan extension: More than a gut feeling? Ageing Res. Rev. 2013, 12, 436–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karpac, J.; Biteau, B.; Jasper, H. Misregulation of an adaptive metabolic response contributes to the age-related disruption of lipid homeostasis in Drosophila. Cell Rep. 2013, 4, 1250–1261. [Google Scholar] [CrossRef] [PubMed]
- Biteau, B.; Karpac, J.; Supoyo, S.; Degennaro, M.; Lehmann, R.; Jasper, H. Lifespan extension by preserving proliferative homeostasis in Drosophila. PLoS Genet. 2010, 6, e1001159. [Google Scholar] [CrossRef] [PubMed]
- Sieber, M.H.; Thummel, C.S. Coordination of triacylglycerol and cholesterol homeostasis by DHR96 and the Drosophila LipA homolog magro. Cell Metab. 2012, 15, 122–127. [Google Scholar] [CrossRef] [PubMed]
- Poirier, L.; Shane, A.; Zheng, J.; Seroude, L. Characterization of the Drosophila gene-switch system in aging studies: A cautionary tale. Aging Cell 2008, 7, 758–770. [Google Scholar] [CrossRef] [PubMed]
- Giannakou, M.E.; Goss, M.; Junger, M.A.; Hafen, E.; Leevers, S.J.; Partridge, L. Long-lived Drosophila with overexpressed dFOXO in adult fat body. Science 2004, 305, 361. [Google Scholar] [CrossRef] [PubMed]
- Wong, A.C.; Dobson, A.J.; Douglas, A.E. Gut microbiota dictates the metabolic response of Drosophila to diet. J. Exp. Biol. 2014, 217, 1894–1901. [Google Scholar] [CrossRef] [PubMed]
- Brummel, T.; Ching, A.; Seroude, L.; Simon, A.F.; Benzer, S. Drosophila lifespan enhancement by exogenous bacteria. Proc. Natl. Acad. Sci. USA 2004, 101, 12974–12979. [Google Scholar] [CrossRef] [PubMed]
- Clark, R.I.; Salazar, A.; Yamada, R.; Fitz-Gibbon, S.; Morselli, M.; Alcaraz, J.; Rana, A.; Rera, M.; Pellegrini, M.; Ja, W.W.; et al. Distinct Shifts in Microbiota Composition during Drosophila Aging Impair Intestinal Function and Drive Mortality. Cell Rep. 2015, 12, 1656–1667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, R.I.; Walker, D.W. Role of gut microbiota in aging-related health decline: Insights from invertebrate models. Cell. Mol. Life Sci. 2018, 75, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Neel, J.V. Diabetes Mellitus—A Thrifty Genotype Rendered Detrimental by Progress. Am. J. Hum. Genet. 1962, 14, 353–362. [Google Scholar] [PubMed]
- Locke, A.E.; Kahali, B.; Berndt, S.I.; Justice, A.E.; Pers, T.H.; Day, F.R.; Powell, C.; Vedantam, S.; Buchkovich, M.L.; Yang, J.; et al. Genetic studies of body mass index yield new insights for obesity biology. Nature 2015, 518, 197–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brookheart, R.T.; Michel, C.I.; Schaffer, J.E. As a Matter of Fat. Cell Metab. 2009, 10, 9–12. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.P.; Fonarow, G.C.; Horwich, T.B. Obesity and the obesity paradox in heart failure. Can. J. Cardiol. 2015, 31, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Hainer, V.; Aldhoon-Hainerova, I. Obesity Paradox Does Exist. Diabetes Care 2013, 36, S276–S281. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Ahmadi, S.F.; Streja, E.; Molnar, M.Z.; Flegal, K.M.; Gillen, D.; Kovesdy, C.P.; Kalantar-Zadeh, K. Obesity paradox in end-stage kidney disease patients. Prog. Cardiovasc. Dis. 2014, 56, 415–425. [Google Scholar] [CrossRef] [PubMed]
- Lennon, H.; Sperrin, M.; Badrick, E.; Renehan, A.G. The Obesity Paradox in Cancer: A Review. Curr. Oncol. Rep. 2016, 18, 56. [Google Scholar] [CrossRef] [PubMed]
- Handee, W.; Li, X.; Hall, K.W.; Deng, X.; Li, P.; Benning, C.; Williams, B.L.; Kuo, M.H. An Energy-Independent Pro-longevity Function of Triacylglycerol in Yeast. PLoS Genet. 2016, 12, e1005878. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Handee, W.; Kuo, M.H. The slim, the fat, and the obese: Guess who lives the longest? Curr. Genet. 2017, 63, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Bailey, A.P.; Koster, G.; Guillermier, C.; Hirst, E.M.; MacRae, J.I.; Lechene, C.P.; Postle, A.D.; Gould, A.P. Antioxidant Role for Lipid Droplets in a Stem Cell Niche of Drosophila. Cell 2015, 163, 340–353. [Google Scholar] [CrossRef] [PubMed]
- Shukla, A.K.; Pragya, P.; Chaouhan, H.S.; Patel, D.K.; Abdin, M.Z.; Kar Chowdhuri, D. Mutation in Drosophila methuselah resists paraquat induced Parkinson-like phenotypes. Neurobiol. Aging 2014, 35, 2419.e1–2419.e16. [Google Scholar] [CrossRef] [PubMed]
- Marik, P.E.; Bellomo, R. Stress hyperglycemia: An essential survival response! Crit. Care 2013, 17. [Google Scholar] [CrossRef] [PubMed]
- Soeters, M.R.; Soeters, P.B. The evolutionary benefit of insulin resistance. Clin. Nutr. 2012, 31, 1002–1007. [Google Scholar] [CrossRef] [PubMed]
- Dionne, M.S.; Pham, L.N.; Shirasu-Hiza, M.; Schneider, D.S. Akt and FOXO dysregulation contribute to infection-induced wasting in Drosophila. Curr. Biol. 2006, 16, 1977–1985. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, E.; Dobes, P.; Kunc, M.; Hyrsl, P.; Kodrik, D. Adipokinetic hormone and adenosine interfere with nematobacterial infection and locomotion in Drosophila melanogaster. J. Insect Physiol. 2018, 107, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Bajgar, A.; Kucerova, K.; Jonatova, L.; Tomcala, A.; Schneedorferova, I.; Okrouhlik, J.; Dolezal, T. Extracellular Adenosine Mediates a Systemic Metabolic Switch during Immune Response. PLoS Biol. 2015, 13, e1002135. [Google Scholar] [CrossRef] [PubMed]
- Becker, T.; Loch, G.; Beyer, M.; Zinke, I.; Aschenbrenner, A.C.; Carrera, P.; Inhester, T.; Schultze, J.L.; Hoch, M. FOXO-dependent regulation of innate immune homeostasis. Nature 2010, 463, 369–373. [Google Scholar] [CrossRef] [PubMed]
- Martins, R.; Lithgow, G.J.; Link, W. Long live FOXO: Unraveling the role of FOXO proteins in aging and longevity. Aging Cell 2016, 15, 196–207. [Google Scholar] [CrossRef] [PubMed]
- Partridge, L.; Alic, N.; Bjedov, I.; Piper, M.D. Ageing in Drosophila: The role of the insulin/Igf and TOR signalling network. Exp. Gerontol. 2011, 46, 376–381. [Google Scholar] [CrossRef] [PubMed]
- Lionaki, E.; Markaki, M.; Tavernarakis, N. Autophagy and ageing: Insights from invertebrate model organisms. Ageing Res. Rev. 2013, 12, 413–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.; Kaushik, S.; Wang, Y.; Xiang, Y.; Novak, I.; Komatsu, M.; Tanaka, K.; Cuervo, A.M.; Czaja, M.J. Autophagy regulates lipid metabolism. Nature 2009, 458, 1131–1135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Liu, H.H.; Liu, S.M.; Wang, S.; Jiang, R.J.; Li, S. Hormonal and nutritional regulation of insect fat body development and function. Arch. Insect Biochem. 2009, 71, 16–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, M.B.; Butterworth, F.M. Maturation and Aging of Adult Fat-Body and Enocytes in Drosophila as Revealed by Light Microscopic Morphometry. J. Morphol. 1985, 184, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Klepsatel, P.; Gáliková, M.; Xu, Y.; Kühnlein, R.P. Thermal stress depletes energy reserves in Drosophila. Sci. Rep. 2016, 6, 33667. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The organs that regulate energy homeostasis and may link obesity to lifespan. The tissues that share similar functions in Drosophila and humans are depicted in the same colors. The median neurosecretory cells of the fly brain produce Insulin-like peptides 2, 3, and 5 (Ilp2, 3, and 5), which act similarly to human insulin produced in the pancreatic beta cells. The Drosophila analog of human glucagon—adipokinetic hormone—is produced in the corpora cardiaca, whereas human glucagon is produced in the alpha cells of the pancreas. In the fly, fat and glycogen are stored in the fat body, which is an organ fulfilling the functions of the human adipose tissue and liver. The dorsal vessel (fly heart) is a linear tube that pumps hemolymph into the open circulatory system, and is considered as a functional counterpart of the human heart. Pericardial nephrocytes are cells that filter fly hemolymph, and share morphological and functional features with podocytes, cells of the kidney glomerulus. See the text for further details.
Figure 1.
The organs that regulate energy homeostasis and may link obesity to lifespan. The tissues that share similar functions in Drosophila and humans are depicted in the same colors. The median neurosecretory cells of the fly brain produce Insulin-like peptides 2, 3, and 5 (Ilp2, 3, and 5), which act similarly to human insulin produced in the pancreatic beta cells. The Drosophila analog of human glucagon—adipokinetic hormone—is produced in the corpora cardiaca, whereas human glucagon is produced in the alpha cells of the pancreas. In the fly, fat and glycogen are stored in the fat body, which is an organ fulfilling the functions of the human adipose tissue and liver. The dorsal vessel (fly heart) is a linear tube that pumps hemolymph into the open circulatory system, and is considered as a functional counterpart of the human heart. Pericardial nephrocytes are cells that filter fly hemolymph, and share morphological and functional features with podocytes, cells of the kidney glomerulus. See the text for further details.

Figure 2.
The fat body of Drosophila. (A) Bright-field image of a subcuticular fat body (white tissue) attached to the abdominal cuticle. (B) Confocal microscope image of the fat body. Cell membranes in white (CellMask Deep Red), lipid droplets in red (BODIPY 493/503) and DNA in blue (Hoechst 33342).
Figure 2.
The fat body of Drosophila. (A) Bright-field image of a subcuticular fat body (white tissue) attached to the abdominal cuticle. (B) Confocal microscope image of the fat body. Cell membranes in white (CellMask Deep Red), lipid droplets in red (BODIPY 493/503) and DNA in blue (Hoechst 33342).


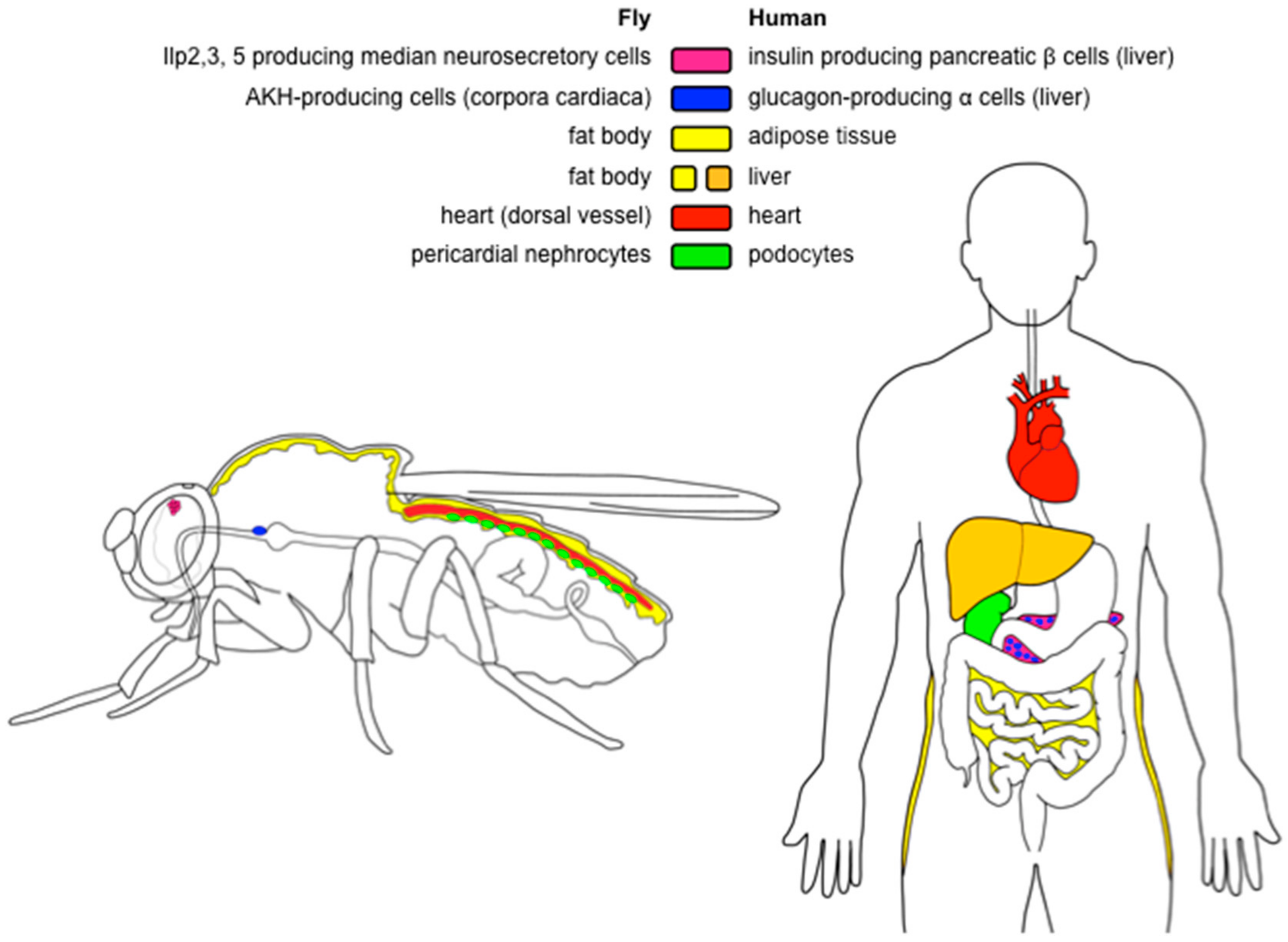{kind=link}
{kind=link}
Table 1.
Examples of obesity-causing manipulations and their effects on the lifespan of Drosophila. Legend: ↑ increase; ↓ decrease; - no change; nd not determined in the study.
Table 1.
Examples of obesity-causing manipulations and their effects on the lifespan of Drosophila. Legend: ↑ increase; ↓ decrease; - no change; nd not determined in the study.
| Obesity Type | Details | Study | Fat | Glycemia | Lifespan | Note | |
|---|---|---|---|---|---|---|---|
| Diet-induced | HSD | sucrose | [23] | ↑ | nd | ↑ | Increase in the mean lifespan, but also increase in the early mortality. |
| glucose | [21] | ↑ | ↑ | ↑ | |||
| sucrose | [31] | ↑ | ↑ | ↓ | Inhibition of the hexosamine pathway rescues lifespan. | ||
| sucrose | [19] | ↑ | ↑ | ↓ | |||
| sucrose | [71] | ↑ | nd | ↓ | Importance of the protein/carbohydrate ratio. | ||
| HFD | lard | [28] | ↑ | ↑ | ↓ | Inhibition of the immune response rescues lifespan, but not obesity. | |
| coconut oil | [29] | ↑ | nd | ↓ | Obesity and decrease in lifespan are ameliorated by endurance exercise. | ||
| coconut oil | [27] | ↑ | ↑ | ↓ | Increased levels of the total body glucose. | ||
| dietary restriction | amino acid restriction | [35] | ↑ | - | ↑ | ||
| yeast restriction | [91] | ↑ | nd | ↑ | |||
| Genetic | IIS related (reduced IIS) | Ilp2-3,5 mutants | [33] | ↑ | nd | - | |
| Ilp2 > rpr | [32] | ↑ | ↑ | ↑ | Ablation of the insulin-producing cells in the brain. | ||
| hsGAL4 > ImpL2 | [65] | ↑ | - | ↑ | Heat shock-inducible overexpression of ImpL2. | ||
| S106 > Ilp6 | [138] | ↑ | ↑ | ↑ | Overexpression of Ilp6 in the fat body. | ||
| Mth mutants | [120] | ↑ | nd | ↑ | Increased lipid reserves inferred from the starvation resistance | ||
| other | female sterile (2) adipose mutants | [103] | ↑ | nd | ↓ | Lifespan decreased in mated females. | |
| bmm mutant | [134] | ↑ | nd | ↓ | |||
| rapamycin | [34] | ↑ | nd | ↑ | Feeding with rapamycin (inhibitor of TOR). | ||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Gáliková, M.; Klepsatel, P. Obesity and Aging in the Drosophila Model. Int. J. Mol. Sci. 2018, 19, 1896. https://doi.org/10.3390/ijms19071896
AMA Style
Gáliková M, Klepsatel P. Obesity and Aging in the Drosophila Model. International Journal of Molecular Sciences. 2018; 19(7):1896. https://doi.org/10.3390/ijms19071896
Chicago/Turabian StyleGáliková, Martina, and Peter Klepsatel. 2018. "Obesity and Aging in the Drosophila Model" International Journal of Molecular Sciences 19, no. 7: 1896. https://doi.org/10.3390/ijms19071896
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.