Cynoglossus semilaevis Rspo3 Regulates Embryo Development by Inhibiting the Wnt/β-Catenin Signaling Pathway


Abstract
:1. Introduction
2. Results
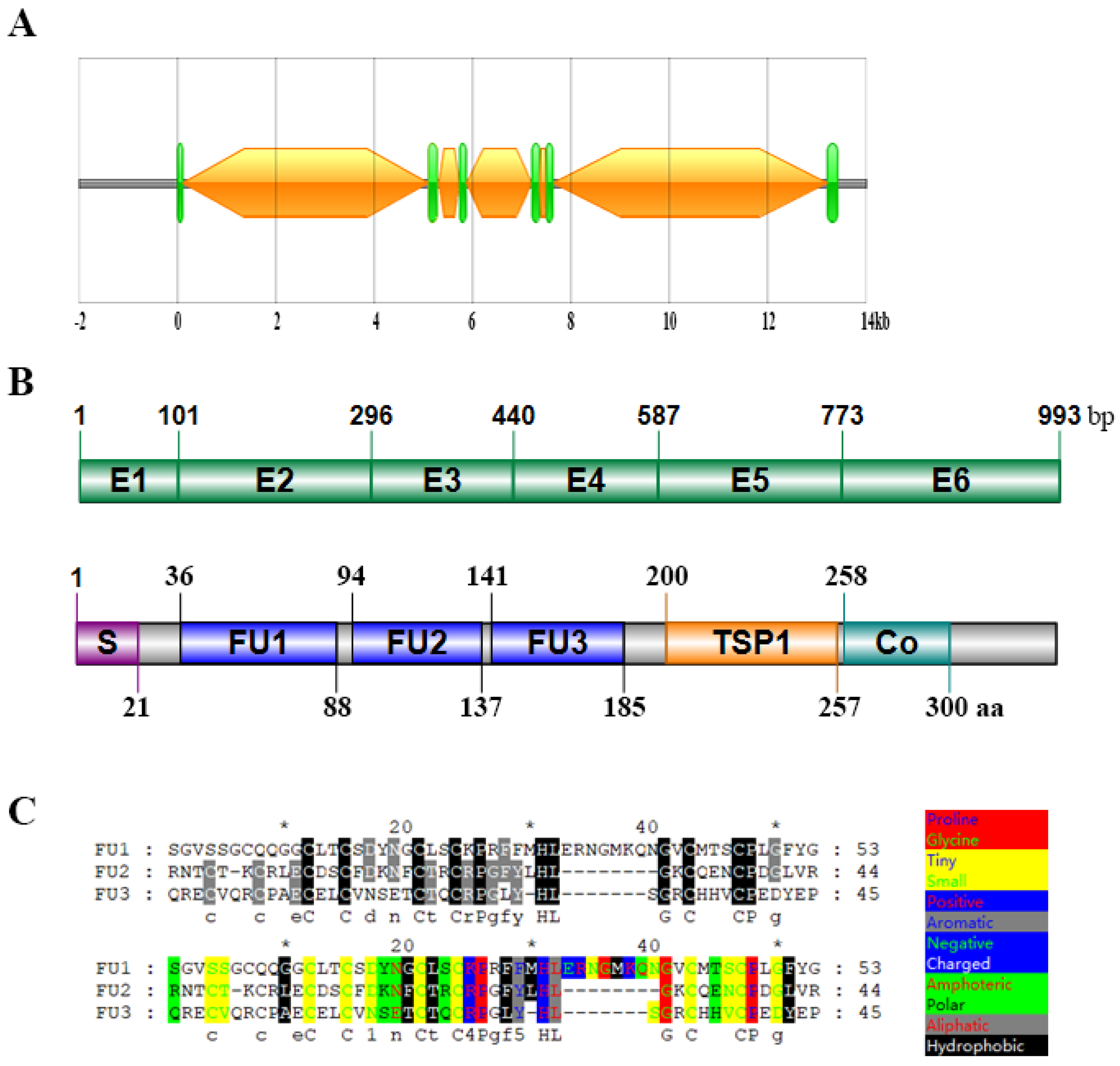
2.1. Cloning and Sequence Structure Analysis
2.2. Phylogenetic Analysis
2.3. Embryonic Expression Pattern
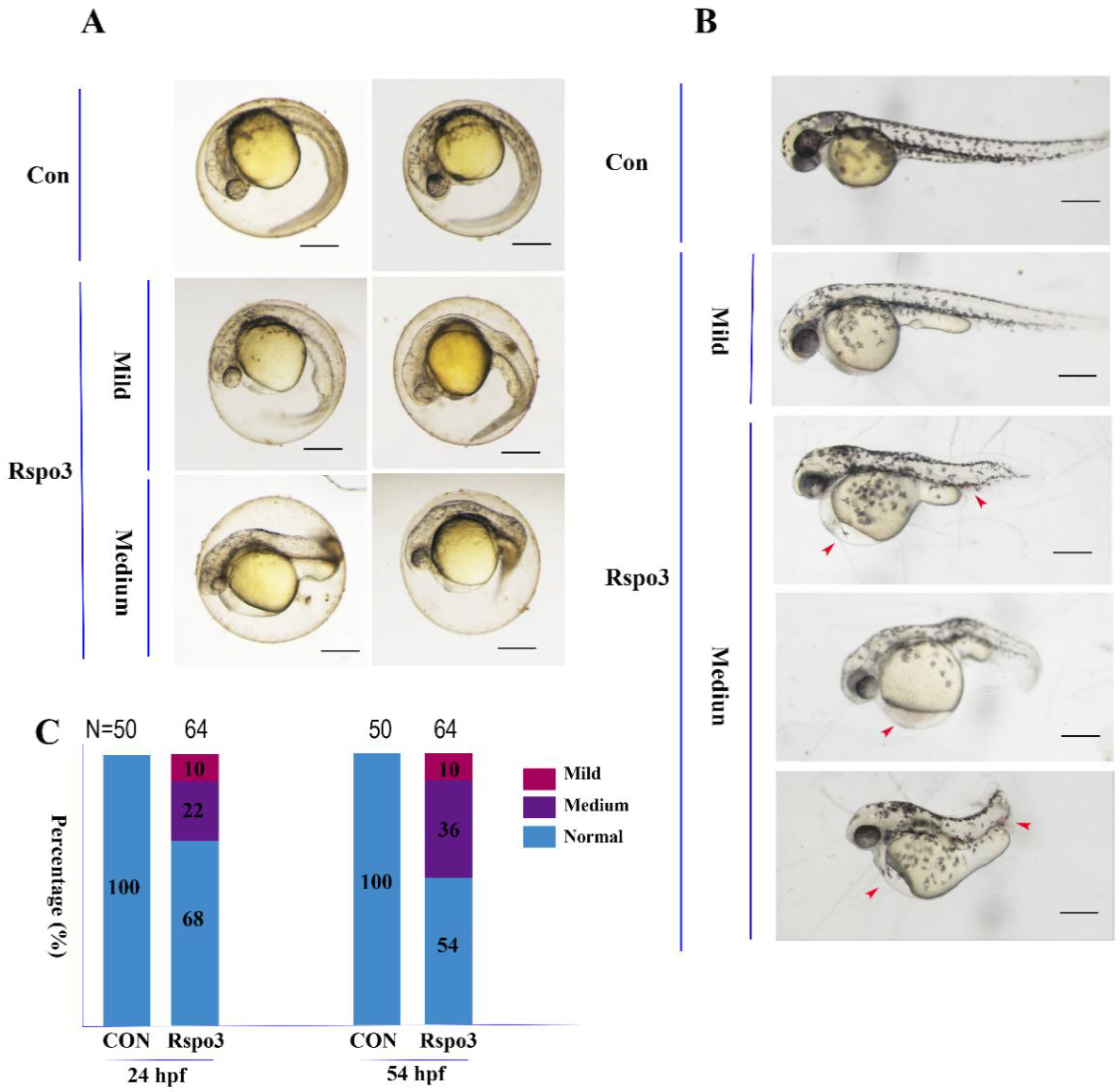
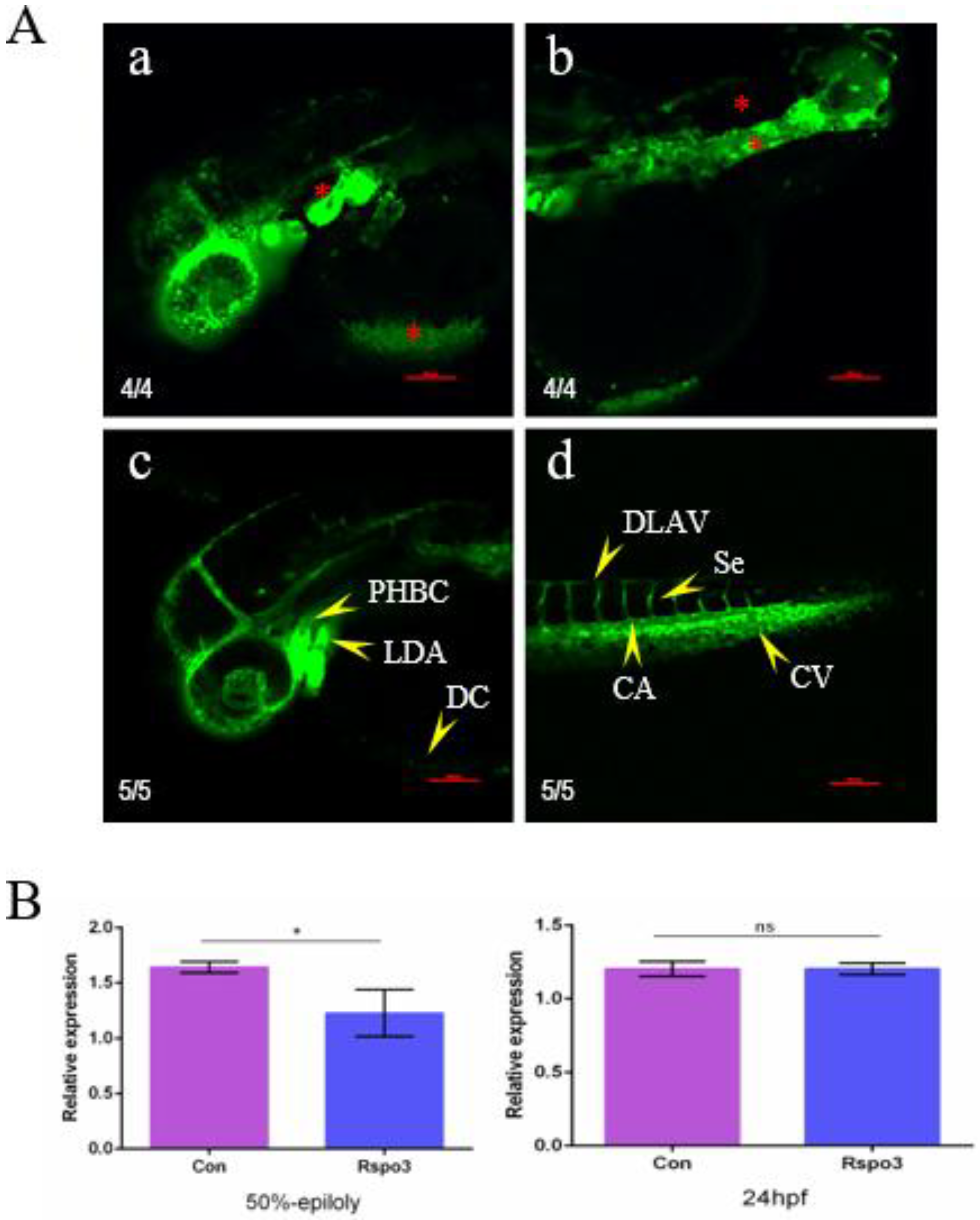
2.4. The Impact of C. semilaevis Rspo3 Overexpression on the Development of D. rerio Embryos
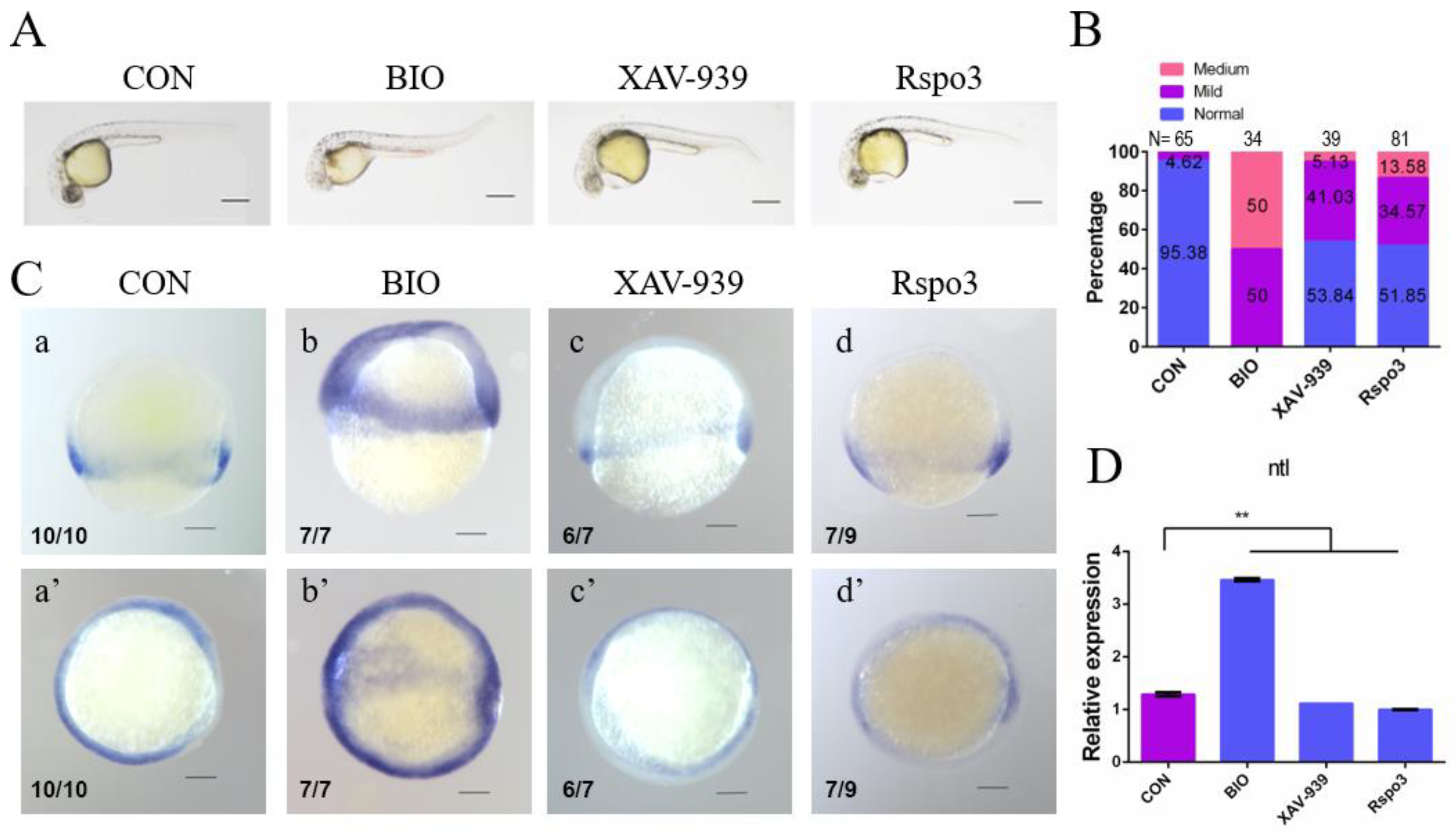
2.5. C. semilaevis Rspo3 Plays a Role in Gastrulation by Regulating the Wnt/β-Catenin Signaling Pathway
2.6. C. semilaevis Rspo3 Suppresses the Wnt/β-Catenin Signaling Pathway in D. rerio Embryos
3. Discussion
4. Methods
4.1. Samples
4.2. Total RNA Extraction and cDNA Synthesis
4.3. Molecular Cloning
4.4. Phylogenetic Analysis
4.5. RT-PCR Analysis and WISH
4.6. mRNA Synthesis and Microinjection
4.7. Luciferase Report Assay
4.8. Drug Treatment
4.9. Mutant Plasmids Construction
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wang, W.; Yi, Q.; Ma, L.; Zhou, X.; Zhao, H.; Wang, X.; Qi, J.; Yu, H.; Wang, Z.; Zhang, Q. Sequencing and characterization of the transcriptome of half-smooth tongue sole (Cynoglossus semilaevis). BMC Genom. 2014, 15, 470. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.L.; Zhang, G.J.; Shao, C.W.; Huang, Q.F.; Liu, G.; Zhang, P.; Song, W.T.; An, N.; Chalopin, D.; Volff, J.N.; et al. Whole-genome sequence of a flatfish provides insights into ZW sex chromosome evolution and adaptation to a benthic lifestyle. Nat. Genet. 2014, 46, 253–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Lau, W.B.; Snel, B.; Clevers, H.C. The R-spondin protein family. Genome Biol. 2012, 13, 242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.A.; Zhao, J.; Andarmani, S.; Kakitani, M.; Oshima, T.; Binnerts, M.E.; Abo, A.; Tomizuka, K.; Funk, W.D. R-Spondin proteins: A novel link to beta-catenin activation. Cell Cycle 2006, 5, 23–26. [Google Scholar] [CrossRef] [PubMed]
- Rong, X.; Chen, C.; Zhou, P.; Zhou, Y.; Li, Y.; Lu, L.; Liu, Y.; Zhou, J.; Duan, C. R-spondin 3 regulates dorsoventral and anteroposterior patterning by antagonizing Wnt/beta-catenin signaling in zebrafish embryos. PLoS ONE 2014, 9, e99514. [Google Scholar] [CrossRef] [PubMed]
- Kazanskaya, O.; Glinka, A.; Barrantes, I.D.; Stannek, P.; Niehrs, C.; Wu, W. R-spondin2 is a secreted activator of Wnt/beta-catenin signaling and is required for Xenopus myogenesis. Dev. Cell 2004, 7, 525–534. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.A.; Kakitani, M.; Zhao, J.; Oshima, T.; Tang, T.; Binnerts, M.; Liu, Y.; Boyle, B.; Park, E.; Emtage, P.; et al. Mitogenic influence of human R-spondin1 on the intestinal epithelium. Science 2005, 309, 1256–1259. [Google Scholar] [CrossRef] [PubMed]
- Lowther, W.; Wiley, K.; Smith, G.H.; Callahan, R. A new common integration site, Int7, for the mouse mammary tumor virus in mouse mammary tumors identifies a gene whose product has furin-like and thrombospondin-like sequences. J. Virol. 2005, 79, 10093–10096. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.A.; Wagle, M.; Tran, K.; Zhan, X.M.; Dixon, M.A.; Liu, S.C.; Gros, D.; Korver, W.; Yonkovich, S.; Tomasevic, N.; et al. R-Spondin family members regulate the Wnt pathway by a common mechanism. Mol. Biol. Cell 2008, 19, 2588–2596. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.Z.; Wang, S.; Tang, R.; Yang, Q.S.; Zhao, E.P.; Chao, Y.Q.; Ying, K.; Xie, Y.; Mao, Y.M. Cloning and identification of a cDNA that encodes a novel human protein with thrombospondin type I repeat domain, hPWTSR. Mol. Biol. Rep. 2002, 29, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Aoki, M.; Mieda, M.; Ikeda, T.; Hamada, Y.; Nakamura, H.; Okamoto, H. R-spondin3 is required for mouse placental development. Dev. Biol. 2007, 301, 218–226. [Google Scholar] [CrossRef] [PubMed]
- Nam, J.S.; Turcotte, T.J.; Smith, P.F.; Choi, S.; Yoon, J.K. Mouse cristin/R-spondin family proteins are novel ligands for the frizzled 8 and LRP6 receptors and activate beta-catenin-dependent gene expression. J. Biol. Chem. 2006, 281, 13247–13257. [Google Scholar] [CrossRef] [PubMed]
- Nam, J.S.; Turcotte, T.J.; Yoon, J.K. Dynamic expression of R-spondin family genes in mouse development. Gene Expr. Patterns 2007, 7, 306–312. [Google Scholar] [CrossRef] [PubMed]
- Kazanskaya, O.; Ohkawara, B.; Heroult, M.; Wu, W.; Maltry, N.; Augustin, H.G.; Niehrs, C. The Wnt signaling regulator R-spondin 3 promotes angioblast and vascular development. Development 2008, 135, 3655–3664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohkawara, B.; Glinka, A.; Niehrs, C. Rspo3 binds Syndecan 4 and induces Wnt/PCP signaling via clathrin-mediated endocytosis to promote morphogenesis. Dev. Cell 2011, 20, 303–314. [Google Scholar] [CrossRef] [PubMed]
- Aoki, M.; Kiyonari, H.; Nakamura, H.; Okamoto, H. R-spondin2 expression in the apical ectodermal ridge is essential for outgrowth and patterning in mouse limb development. Dev. Growth Differ. 2008, 50, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, F.; Rocha, A.S.; Motamedi, F.J.; Massa, F.; Basboga, C.; Morrison, H.; Wagner, K.D.; Schedl, A. Coronary artery formation is driven by localized expression of R-spondin3. Cell Rep. 2017, 20, 1745–1754. [Google Scholar] [CrossRef] [PubMed]
- Kannan, L.; Kis-Toth, K.; Yoshiya, K.; Thai, T.H.; Sehrawat, S.; Mayadas, T.N.; Lucca, J.J.D.; Tsokos, G.C. R-spondin3 prevents mesenteric ischemia/reperfusion-induced tissue damage by tightening endothelium and preventing vascular leakage. Proc. Natl. Acad. Sci. USA 2013, 110, 14348–14353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cambier, L.; Plate, M.; Sucov, H.M.; Pashmforoush, M. Nkx2-5 regulates cardiac growth through modulation of Wnt signaling by R-spondin3. Development 2014, 141, 2959–2971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neufeld, S.; Rosin, J.M.; Ambasta, A.; Hui, K.; Shaneman, V.; Crowder, R.; Vickerman, L.; Cobb, J. A conditional allele of Rspo3 reveals redundant function of R-spondins during mouse limb development. Genesis 2012, 50, 741–749. [Google Scholar] [CrossRef] [PubMed]
- Rocha, A.S.; Vidal, V.; Mertz, M.; Kendall, T.J.; Charlet, A.; Okamoto, H.; Schedl, A. The angiocrine factor Rspondin3 is a key determinant of liver zonation. Cell Rep. 2015, 13, 1757–1764. [Google Scholar] [CrossRef] [PubMed]
- Theodorou, V.; Kimm, M.A.; Boer, M.; Wessels, L.; Theelen, W.; Jonkers, J.; Hilkens, J. MMTV insertional mutagenesis identifies genes, gene families and pathways involved in mammary cancer. Nat. Genet. 2007, 39, 759–769. [Google Scholar] [CrossRef] [PubMed]
- Galetzka, D.; Hansmann, T.; El Hajj, N.; Weis, E.; Irmscher, B.; Ludwig, M.; Schneider-Ratzke, B.; Kohlschmidt, N.; Beyer, V.; Bartsch, O.; et al. Monozygotic twins discordant for constitutive BRCA1 promoter methylation, childhood cancer and secondary cancer. Epigenetics 2012, 7, 47–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seshagiri, S.; Stawiski, E.W.; Durinck, S.; Modrusan, Z.; Storm, E.E.; Conboy, C.B.; Chaudhuri, S.; Guan, Y.H.; Janakiraman, V.; Jaiswal, B.S.; et al. Recurrent R-spondin fusions in colon cancer. Nature 2012, 488, 660–664. [Google Scholar] [CrossRef] [PubMed]
- Han, T.; Schatoff, E.M.; Murphy, C.; Zafra, M.P.; Wilkinson, J.E.; Elemento, O.; Dow, L.E. R-Spondin chromosome rearrangements drive Wnt-dependent tumour initiation and maintenance in the intestine. Nat. Commun. 2017, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Duncan, E.L.; Danoy, P.; Kemp, J.P.; Leo, P.J.; McCloskey, E.; Nicholson, G.C.; Eastell, R.; Prince, R.L.; Eisman, J.A.; Jones, G.; et al. Genome-wide association study using extreme truncate selection identifies novel genes affecting bone mineral density and fracture risk. PLoS Genet. 2011, 7, e1001372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, Y.H.; Kiel, D.P. Genome-wide association studies of skeletal phenotypes: What we have learned and where we are headed. J. Clin. Endocr. Metab. 2012, 97, E1958–E1977. [Google Scholar] [CrossRef] [PubMed]
- Moayyeri, A.; Hsu, Y.H.; Karasik, D.; Estrada, K.; Xiao, S.M.; Nielson, C.; Srikanth, P.; Giroux, S.; Wilson, S.G.; Zheng, H.F.; et al. Genetic determinants of heel bone properties: Genome-wide association meta-analysis and replication in the GEFOS/GENOMOS consortium. Hum. Mol. Genet. 2014, 23, 3054–3068. [Google Scholar] [CrossRef] [PubMed]
- Heid, I.M.; Jackson, A.U.; Randall, J.C.; Winkler, T.W.; Qi, L.; Steinthorsdottir, V.; Thorleifsson, G.; Zillikens, M.C.; Speliotes, E.K.; Magi, R.; et al. Magic, Meta-analysis identifies 13 new loci associated with waist-hip ratio and reveals sexual dimorphism in the genetic basis of fat distribution. Nat. Genet. 2010, 42, 949–960. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Go, M.J.; Hu, C.; Hong, C.B.; Kim, Y.K.; Lee, J.Y.; Hwang, J.Y.; Oh, J.H.; Kim, D.J.; Kim, N.H.; et al. Large-scale genome-wide association studies in east Asians identify new genetic loci influencing metabolic traits. Nat. Genet. 2011, 43, 990–995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.T.; Monda, K.L.; Taylor, K.C.; Lange, L.; Demerath, E.W.; Palmas, W.; Wojczynski, M.K.; Ellis, J.C.; Vitolins, M.Z.; Liu, S.M.; et al. Genome-wide association of body fat distribution in African ancestry populations suggests new loci. PLoS Genet. 2013, 9, e1003681. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Ryu, J.; Lee, C. Genome-wide identification of expression quantitative trait loci for human telomerase. Medicine 2016, 95, e5209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schier, A.F.; Talbot, W.S. Molecular genetics of axis formation in zebrafish. Annu. Rev. Genet. 2005, 39, 561–613. [Google Scholar] [CrossRef] [PubMed]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic-development of the zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef] [PubMed]
- Huber, T.L.; Kouskoff, V.; Fehling, H.J.; Palis, J.; Keller, G. Haemangioblast commitment is initiated in the primitive streak of the mouse embryo. Nature 2004, 432, 625–630. [Google Scholar] [CrossRef] [PubMed]
- Binnerts, M.E.; Kim, K.A.; Bright, J.M.; Patel, S.M.; Tran, K.; Zhou, M.; Leung, J.M.; Liu, Y.; Lomas, W.E.; Dixon, M.; et al. R-Spondin1 regulates Wnt signaling by inhibiting internalization of LRP6. Proc. Natl. Acad. Sci. USA 2007, 104, 14700–14705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glinka, A.; Dolde, C.; Kirsch, N.; Huang, Y.L.; Kazanskaya, O.; Ingelfinger, D.; Boutros, M.; Cruciat, C.M.; Niehrs, C. LGR4 and LGR5 are R-spondin receptors mediating Wnt/beta-catenin and Wnt/PCP signalling. Embo Rep. 2011, 12, 1055–1061. [Google Scholar] [CrossRef] [PubMed]
- Langdon, Y.G.; Mullins, M.C. Maternal and zygotic control of zebrafish dorsoventral axial patterning. Annu. Rev. Genet. 2011, 45, 357–377. [Google Scholar] [CrossRef] [PubMed]
- Carmon, K.S.; Gong, X.; Lin, Q.S.; Thomas, A.; Liu, Q.Y. R-spondins function as ligands of the orphan receptors LGR4 and LGR5 to regulate Wnt/beta-catenin signaling. Proc. Natl. Acad. Sci. USA 2011, 108, 11452–11457. [Google Scholar] [CrossRef] [PubMed]
- De Lau, W.; Barker, N.; Low, T.Y.; Koo, B.K.; Li, V.S.W.; Teunissen, H.; Kujala, P.; Haegebarth, A.; Peters, P.J.; van de Wetering, M.; et al. Lgr5 homologues associate with Wnt receptors and mediate R-spondin signalling. Nature 2011, 476, 293–297. [Google Scholar] [CrossRef] [PubMed]
- Hao, H.X.; Xie, Y.; Zhang, Y.; Charlat, O.; Oster, E.; Avello, M.; Lei, H.; Mickanin, C.; Liu, D.; Ruffner, H.; et al. ZNRF3 promotes Wnt receptor turnover in an R-spondin-sensitive manner. Nature 2012, 485, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Koo, B.K.; Spit, M.; Jordens, I.; Low, T.Y.; Stange, D.E.; van de Wetering, M.; van Es, J.H.; Mohammed, S.; Heck, A.J.R.; Maurice, M.M.; et al. Tumour suppressor RNF43 is a stem-cell E3 ligase that induces endocytosis of Wnt receptors. Nature 2012, 488, 665–669. [Google Scholar] [CrossRef] [PubMed]
- Carmon, K.S.; Lin, Q.S.; Gong, X.; Thomas, A.; Liu, Q.Y. LGR5 Interacts and cointernalizes with Wnt receptors to modulate Wnt/beta-catenin signaling. Mol. Cell. Biol. 2012, 32, 2054–2064. [Google Scholar] [CrossRef] [PubMed]
- Ruffner, H.; Sprunger, J.; Charlat, O.; Leighton-Davies, J.; Grosshans, B.; Salathe, A.; Zietzling, S.; Beck, V.; Therier, M.; Isken, A.; et al. R-Spondin potentiates Wnt/beta-catenin signaling through orphan receptors LGR4 and LGR5. PLoS ONE 2012, 7, e40976. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.H.; Chen, X.Y.; Lin, Z.H.; Fang, D.Y.; He, X.L. The structural basis of R-spondin recognition by LGR5 and RNF43. Gene Dev. 2013, 27, 1345–1350. [Google Scholar] [CrossRef] [PubMed]
- Lebensohn, A.M.; Rohatgi, R. R-spondins can potentiate WNT signaling without LGRs. eLife 2018, 7, e33126. [Google Scholar] [CrossRef] [PubMed]
- Zebisch, M.; Jones, E.Y. Crystal structure of R-spondin 2 in complex with the ectodomains of its receptors LGR5 and ZNRF3. J. Struct. Biol. 2015, 191, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Zebisch, M.; Xu, Y.; Krastev, C.; MacDonald, B.T.; Chen, M.R.; Gilbert, R.J.C.; He, X.; Jones, E.Y. Structural and molecular basis of ZNRF3/RNF43 transmembrane ubiquitin ligase inhibition by the Wnt agonist R-spondin. Nat. Commun. 2013, 4, 2787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Hohna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.X.; Liu, T.T.; Niu, J.J.; Wu, X.L.; Zhai, J.M.; Zhang, Q.Q.; Qi, J. Expression pattern and functional analysis of R-spondin1 in tongue sole Cynoglossus semilaevis. Gene 2018, 642, 453–460. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′to 3′) |
|---|---|
| CS-Rspo3-ORF-Fw | TGGGCTACTATGCAATTACAACTG |
| CS-Rspo3-ORF-Rv | AGAGGGAGCTCACTGTACAA |
| CS-Rspo3-WISH-Fw | ATTTAGGTGACACTATAGAAGNGGGCACCGGCAACATAAAC |
| CS-Rspo3-WISH-Rv | TAATACGACTCACTATAGGGAGACTCCACTCACCCACTTCAC |
| CS-Rspo3-RT-PCR-Fw | GGGTCCTGTTGTGTTTAGGA |
| CS-Rspo3-RT-PCR-Rv | TGCTGACCACCAGTGTAATC |
| CS-Rspo3-mRNA-Fw | TAATACGACTCACTATAGGGAGAATGCAATTACAACTGATCTCCTTTG |
| CS-Rspo3-mRNA-Rv | AGAGGGAGCTCACTGTACAA |
| CS-Rspo3-ORF-EC-Fw | CGCGGATCCGCCACCATGCAATTACAACTGATCTCC |
| CS-Rspo3-ORF-EC-Rv | CCGGAATTCGCTGTACAAGGTCATCATC |
| CS-Rspo3-delFU1-Fw | CGGCAACATAAACAGATCATGCGCTCTCACGAAAGA |
| CS-Rspo3-delFU1-Rv | TCTTTCGTGAGAGCGCATGATCTGTTTATGTTGCCG |
| CS-Rspo3-delFU2-Fw | CCTCTGTGCGTCACTTTCGTGAGAGCGCATGC |
| CS-Rspo3-delFU2-Rv | GCATGCGCTCTCACGAAAGTGACGCACCAGAGG |
| CS-Rspo3-delFU3-Fw | GGTCCGCAGTGACGCAAACGACAAACTCATGG |
| CS-Rspo3-delFU3-Rv | CCATGAGTTTGTCGTTTGCGTCACTGCGGACC |
| CS-Rspo3-delTSP-Fw | CCACAAGTGCACTGTGAAGGACGGAGGAAGAATG |
| CS-Rspo3-delTSP-Rv | CATTCTTCCTCCHTCCTTCACAGTGCACTTGTGG |
| B2M-RT-PCR-Fw | TGTTCGTCGTTC TGCCGTGT |
| B2M-RT-PCR-Rv | TCAGGGTGTTGGGCTTGTTGT |
| DR-VEGFa-RT-PCR-Fw | AGTTATTTCTCGCGGCTCTCC |
| DR-VEGFa-RT-PCR-Rv | ACACATCCATGAAGGGAATCAC |
| DR-ntl-RT-PCR-Fw | GGATGAAAGCACCCGTATC |
| DR-ntl-RT-PCR-Rv | GTGTATCCTGGGTTCGTATTT |
| DR-boz-RT-PCR-Fw | TAGAGACAGAGCAAGAGGAG |
| DR-boz-RT-PCR-Rv | GGTGTCTCCTAAGATGTAATCAA |
| DR-ZNRF3-RT-PCR-Fw | TTGGACCCAAGCTGTCTTAC |
| DR-ZNRF3-RT-PCR-Rv | CACGCTGACCCTTGAACTTA |
| DR-sp5l-RT-PCR-Fw | GGAGGTCACGTTGAGGATGG |
| DR-sp5l-RT-PCR-Rv | GCGACAGCGACGAGTAGAGC |
| DR-β-actin-RT-PCR-Fw | CCCAGACATCAGGGAGTGAT |
| DR-β-actin-RT-PCR-Rv | TCTCTGTTGGCTTTGGATT |
| DR-ntl-WISH-Fw | ATTTAGGTGACACTATAGAAGNGATGACTTCTTCCAACCCCG |
| DR-ntl-WISH-Rv | TAATACGACTCACTATAGGGAGATCAGTAGCTCTGAGCCACAGG |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niu, J.; Guan, J.; Li, R.; Li, X.; Zhai, J.; Qi, J.; He, Y. Cynoglossus semilaevis Rspo3 Regulates Embryo Development by Inhibiting the Wnt/β-Catenin Signaling Pathway. Int. J. Mol. Sci. 2018, 19, 1915. https://doi.org/10.3390/ijms19071915
Niu J, Guan J, Li R, Li X, Zhai J, Qi J, He Y. Cynoglossus semilaevis Rspo3 Regulates Embryo Development by Inhibiting the Wnt/β-Catenin Signaling Pathway. International Journal of Molecular Sciences. 2018; 19(7):1915. https://doi.org/10.3390/ijms19071915
Chicago/Turabian StyleNiu, Jingjing, Jian Guan, Rui Li, Xuemei Li, Jieming Zhai, Jie Qi, and Yan He. 2018. "Cynoglossus semilaevis Rspo3 Regulates Embryo Development by Inhibiting the Wnt/β-Catenin Signaling Pathway" International Journal of Molecular Sciences 19, no. 7: 1915. https://doi.org/10.3390/ijms19071915