Comparison of the Effect of Melatonin Treatment before and after Brain Ischemic Injury in the Inflammatory and Apoptotic Response in Aged Rats

 , ,
, ,
Abstract
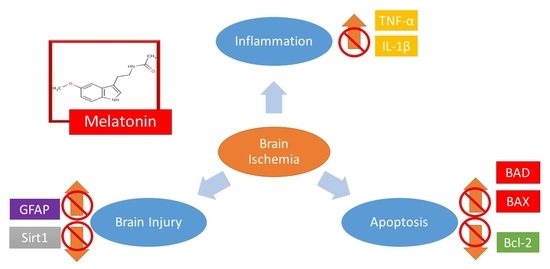
:
1. Introduction
2. Results
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Surgical Procedure
4.3. Administration of Melatonin
4.4. RNA Isolation and RT-PCR
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lo, E.H.; Dalkara, T.; Moskowitz, M.A. Mechanisms, challenges and opportunities in stroke. Nat. Rev. Neurosci. 2003, 4, 399–415. [Google Scholar] [CrossRef] [PubMed]
- Donnan, G.A.; Fisher, M.; Macleod, M.; Davis, S.M. Stroke. Lancet 2008, 371, 1612–1623. [Google Scholar] [CrossRef]
- Muir, K.W.; Tyrrell, P.; Sattar, N.; Warburton, E. Inflammation and ischaemic stroke. Curr. Opin. Neurol. 2007, 20, 334–342. [Google Scholar] [CrossRef] [PubMed]
- Dirnagl, U.; Iadecola, C.; Moskowitz, M.A. Pathobiology of ischaemic stroke: An integrated view. Trends Neurosci. 1999, 22, 391–397. [Google Scholar] [CrossRef]
- Ferrarese, C.; Mascarucci, P.; Zoia, C.; Cavarretta, R.; Frigo, M.; Begni, B.; Sarinella, F.; Frattola, L.; De Simoni, M.G. Increased cytokine release from peripheral blood cells after acute stroke. J. Cereb. Blood Flow Metab. 1999, 19, 1004–1009. [Google Scholar] [CrossRef] [PubMed]
- Caso, J.R.; Moro, M.A.; Lorenzo, P.; Lizasoain, I.; Leza, J.C. Involvement of IL-1beta in acute stress induced worsening of cerebral ischemia in rats. Eur. Neuropsychopharmacol. 2007, 17, 600–607. [Google Scholar] [CrossRef] [PubMed]
- Paredes, S.D.; Rancan, L.; Kireev, R.; González, A.; Louzao, P.; González, P.; Rodríguez-Bobada, C.; García, C.; Vara, E.; Tresguerres, J.A. Melatonin Counteracts at a Transcriptional Level the Inflammatory and Apoptotic Response Secondary to Ischemic Brain Injury Induced by Middle Cerebral Artery Blockade in Aging Rats. Biores. Open Access 2015, 4, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Yang, G.Y.; Ahlemeyer, B.; Pang, L.; Che, X.M.; Culmsee, C.; Klumpp, S.; Krieglstein, J. Transforming growth factor-beta 1 increases bad phosphorylation and protects neurons against damage. J. Neurosci. 2002, 22, 3898–3909. [Google Scholar] [CrossRef] [PubMed]
- Spera, P.A.; Ellison, J.A.; Feuerstein, G.Z.; Barone, F.C. IL-10 reduces rat brain injury following focal stroke. Neurosci. Lett. 1998, 251, 189–192. [Google Scholar] [CrossRef]
- Puig, Á.; Rancan, L.; Paredes, S.D.; Carrasco, A.; Escames, G.; Vara, E.; Tresguerres, J.A. Melatonin decreases the expression of inflammation and apoptosis markers in the lung of a senescence-accelerated mice model. Exp. Gerontol. 2016, 75, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Paredes, S.D.; Forman, K.A.; García, C.; Vara, E.; Escames, G.; Tresguerres, J.A. Protective actions of melatonin and growth hormone on the aged cardiovascular system. Horm. Mol. Biol. Clin. Investig. 2014, 18, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Tan, D.X.; Fuentes-Broto, L.; Paredes, S.D.; Sanchez-Barcelo, E.; Mediavilla, M.D. Melatonin salvages neural tissue from ischemia/reperfusion injury. Open Neurendocrinol. J. 2010, 3, 112–120. [Google Scholar]
- Jones, T.H.; Morawetz, R.B.; Crowell, R.M.; Marcoux, F.W.; FitzGibbon, S.J.; DeGirolami, U.; Ojemann, R.G. Thresholds of focal ischemia in awake monkeys. J. Neurosurg. 1981, 54, 773–782. [Google Scholar] [CrossRef] [PubMed]
- Astrup, J.; Seisjo, B.K.; Symon, L. Thresholds in cerebral ischemia—The ischemic penumbra. Stroke 1981, 12, 723–725. [Google Scholar] [CrossRef] [PubMed]
- Wise, R.J.; Bernardi, S.; Frackowiak, R.S.; Legg, N.J.; Jones, T. Serial observations on the pathophysiology of acute stroke: The transition from ischaemia to infarction as reflected in regional oxygen extraction. Brain 1983, 106, 197–222. [Google Scholar] [CrossRef] [PubMed]
- Heros, R. Stroke: Early pathophysiology and treatment. Summary of the Fifth Annual Decade of the Brain Symposium. Stroke 1994, 25, 1877–1881. [Google Scholar] [CrossRef] [PubMed]
- Garcia, J.H.; Yoshida, Y.; Chen, H.; Li, Y.; Zhang, Z.G.; Lian, J.; Chen, S.; Chopp, M. Progression from ischemic injury to infarct following middle cerebral artery occlusion in the rat. Am. J. Pathol. 1993, 142, 623–635. [Google Scholar] [PubMed]
- Hossmann, K.A. Viability thresholds and the penumbra of focal ischemia. Ann. Neurol. 1994, 36, 557–565. [Google Scholar] [CrossRef] [PubMed]
- Kalogeris, T.; Bao, Y.; Korthuis, R.J. Mitochondrial reactive oxygen species: A double edged sword in ischemia/reperfusion vs preconditioning. Redox Biol. 2014, 2, 702–714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doll, D.N.; Rellick, S.L.; Barr, T.L.; Ren, X.; Simpkins, J.W. Rapid mitochondrial dysfunction mediates TNF-alpha-induced neurotoxicity. J. Neurochem. 2015, 132, 443–451. [Google Scholar] [CrossRef] [PubMed]
- Siu, A.W.; Maldonado, M.; Sanchez-Hidalgo, M.; Tan, D.X.; Reiter, R.J. Protective effects of melatonin in experimental free radical-related ocular diseases. J. Pineal Res. 2006, 40, 101–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, D.X.; Manchester, L.C.; Terron, M.P.; Flores, L.J.; Reiter, R.J. One molecule, many derivatives: A never-ending interaction of melatonin with reactive oxygen and nitrogen species? J. Pineal Res. 2007, 42, 28–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montilla, P.L.; Túnez, I.F.; Muñoz de Agueda, C.; Gascón, F.L.; Soria, J.V. Protective role of melatonin and retinol palmitate in oxidative stress and hyperlipidemic nephropathy induced by adriamycin in rats. J. Pineal Res. 1998, 25, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Okatani, Y.; Wakatsuki, A.; Kaneda, C. Melatonin increases activities of glutathione peroxidase and superoxide dismutase in fetal rat brain. J. Pineal Res. 2000, 28, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Tan, D.X.; Terron, M.P.; Flores, L.J.; Czarnocki, Z. Melatonin and its metabolites: New findings regarding their production and their radical scavenging actions. Acta Biochim. Pol. 2007, 54, 1–9. [Google Scholar] [PubMed]
- Reiter, R.J.; Tan, D.X.; Cabrera, J.; D’Arpa, D. Melatonin and tryptophan derivatives as free radical scavengers and antioxidants. Adv. Exp. Med. Biol. 1999, 467, 379–387. [Google Scholar] [PubMed]
- Mésenge, C.; Margaill, I.; Verrecchia, C.; Allix, M.; Boulu, R.G.; Plotkine, M. Protective effect of melatonin in a model of traumatic brain injury in mice. J. Pineal Res. 1998, 25, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Wakatsuki, A.; Okatani, Y.; Shinohara, K.; Ikenoue, N.; Fukaya, T. Melatonin protects against ischemia/reperfusion-induced oxidative damage to mitochondria in fetal rat brain. J. Pineal Res. 2001, 31, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Cuesta, J.; Tajes, M.; Jiménez, A.; Coto-Montes, A.; Camins, A.; Pallàs, M. Evaluation of potential pro-survival pathways regulated by melatonin in a murine senescence model. J. Pineal Res. 2008, 45, 497–505. [Google Scholar] [CrossRef] [PubMed]
- Mayo, J.C.; Sainz, R.M.; González Meléndez, P.; Cepas, V.; Tan, D.X.; Reiter, R.J. Melatonin and sirtuins: A ‘not-so unexpected’ relationship. J. Pineal Res. 2017, 62, e12391. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Liu, H.; Yue, L.; Zhang, J.; Li, X.; Wang, B.; Lin, Y.; Qu, Y. Melatonin attenuates early brain injury via the melatonin receptor/Sirt1/NF-KB signaling pathway following subarachnoid hemorrhage in mice. Mol. Neurobiol. 2017, 54, 1612–1621. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.S.; Wu, Q.; Wu, L.Y.; Ye, Z.N.; Jiang, T.W.; Li, W.; Zhuang, Z.; Zhou, M.L.; Zhang, X.; Hang, C.H. Sirtuin 1 activation protects against early brain injury after experimental subarachnoid hemorrhage in rats. Cell Death Dis. 2016, 7, e2416. [Google Scholar] [CrossRef] [PubMed]
- Qian, C.; Jin, J.; Chen, J.; Li, J.; Yu, X.; Mo, H.; Chen, G. SIRT1 activation by resveratrol reduces brain edema and neuronal apoptosis in an experimental rat subarachnoid hemorrhage model. Mol. Med. Rep. 2017, 16, 9627–9635. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Jiang, S.; Dong, Y.; Fan, C.; Zhao, L.; Yang, X.; Li, J.; Di, S.; Yue, L.; Liang, G.; et al. Melatonin prevents cell death and mitochondrial dysfunction via a SIRT1-dependent mechanism during ischemic-stroke in mice. J. Pineal Res. 2015, 58, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Ramos, E.; Patiño, P.; Reiter, R.J.; Gil-Martín, E.; Marco-Contelles, J.; Parada, E.; de Los Rios, C.; Romero, A.; Egea, J. Ischemic brain injury: New insights on the protective role of melatonin. Free Radic. Biol. Med. 2017, 104, 32–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cipolla, M.J.; Lessov, N.; Hammer, E.S.; Curry, A.B. Threshold duration of ischemia for myogenic tone in middle cerebral arteries: Effect on vascular smooth muscle actin. Stroke 2001, 32, 1658–1664. [Google Scholar] [CrossRef] [PubMed]
- Reagan-Shaw, S.; Nihal, M.; Ahmad, N. Dose translation from animal to human studies revisited. FASEB J. 2008, 22, 659–661. [Google Scholar] [CrossRef] [PubMed]
- Weishaupt, J.H.; Bartels, C.; Pölking, E.; Dietrich, J.; Rohde, G.; Poeggeler, B.; Mertens, N.; Sperling, S.; Bohn, M.; Hüther, G.; et al. Reduced oxidative damage in ALS by high-dose enteral melatonin treatment. J. Pineal Res. 2006, 41, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Buscemi, N.; Vandermeer, B.; Pandya, R.; Hooton, N.; Tjosvold, L.; Hartling, L.; Baker, G.; Vohra, S.; Klassen, T. Melatonin for treatment of sleep disorders. Evid. Rep. Technol. Assess 2004, 108, 1–7. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Marker | Primers | Sequence (5′–3′) |
|---|---|---|
| 18S | Forward | GGTGCATGGCCGTTCTTA |
| Reverse | TCGTTCGTTATCGGAATTAACC | |
| IL-1β | Forward | TGTGATGAAAGACGGCACAC |
| Reverse | CTTCTTCTTTGGGTATTGTTTGG | |
| TNF-α | Forward | ATGAGAAGTTCCCAAATGGC |
| Reverse | CTCCACTTGGTGGTTTGCTA | |
| BAD | Forward | GCCCTAGGCTTGAGGAAGTC |
| Reverse | CAAACTCTGGGATCTGGAACA | |
| BAX | Forward | GTGAGCGGCTGCTTGTCT |
| Reverse | GGTCCCGAAGTAGGAGAGGA | |
| GFAP | Forward | ACAGACTTTCTCCAACCTCCAG |
| Reverse | CCTTCTGACACGGATTTGGT | |
| Bcl-2 | Forward | CAGGTATGCACCCAGAGTGA |
| Reverse | GTCTCTGAAGACGCTGCTCA | |
| SIRT1 | Forward | TCGTGGAGACATTTTTAATCAGG |
| Reverse | GCTTCATGATGGCAAGTGG |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rancan, L.; Paredes, S.D.; García, C.; González, P.; Rodríguez-Bobada, C.; Calvo-Soto, M.; Hyacinthe, B.; Vara, E.; Tresguerres, J.A.F. Comparison of the Effect of Melatonin Treatment before and after Brain Ischemic Injury in the Inflammatory and Apoptotic Response in Aged Rats. Int. J. Mol. Sci. 2018, 19, 2097. https://doi.org/10.3390/ijms19072097
Rancan L, Paredes SD, García C, González P, Rodríguez-Bobada C, Calvo-Soto M, Hyacinthe B, Vara E, Tresguerres JAF. Comparison of the Effect of Melatonin Treatment before and after Brain Ischemic Injury in the Inflammatory and Apoptotic Response in Aged Rats. International Journal of Molecular Sciences. 2018; 19(7):2097. https://doi.org/10.3390/ijms19072097
Chicago/Turabian StyleRancan, Lisa, Sergio D. Paredes, Cruz García, Pablo González, Cruz Rodríguez-Bobada, Mario Calvo-Soto, Bryan Hyacinthe, Elena Vara, and Jesús A. F. Tresguerres. 2018. "Comparison of the Effect of Melatonin Treatment before and after Brain Ischemic Injury in the Inflammatory and Apoptotic Response in Aged Rats" International Journal of Molecular Sciences 19, no. 7: 2097. https://doi.org/10.3390/ijms19072097