Effect of ACTH and hCG on the Expression of Gonadotropin-Inducible Ovarian Transcription Factor 1 (Giot1) Gene in the Rat Adrenal Gland



Abstract
:
1. Introduction
2. Results
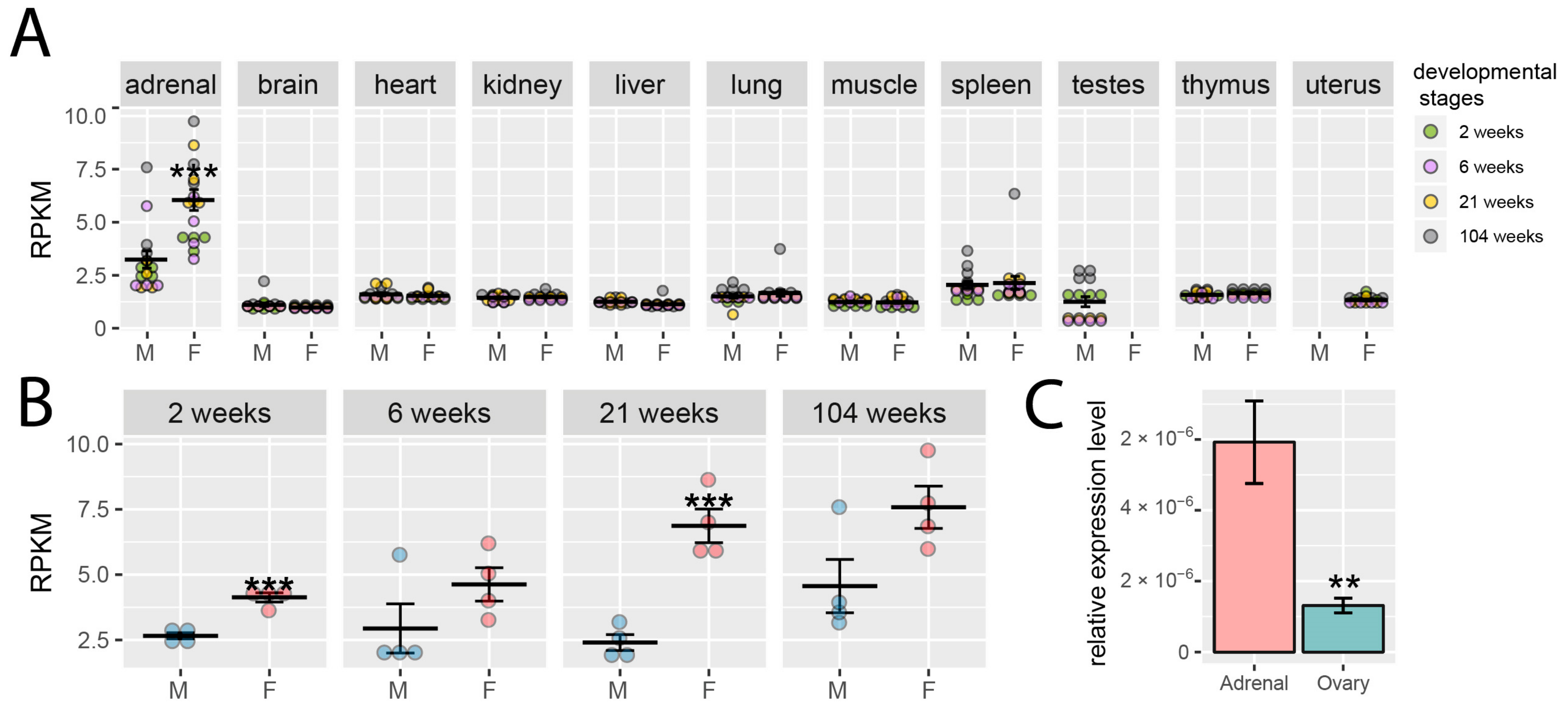
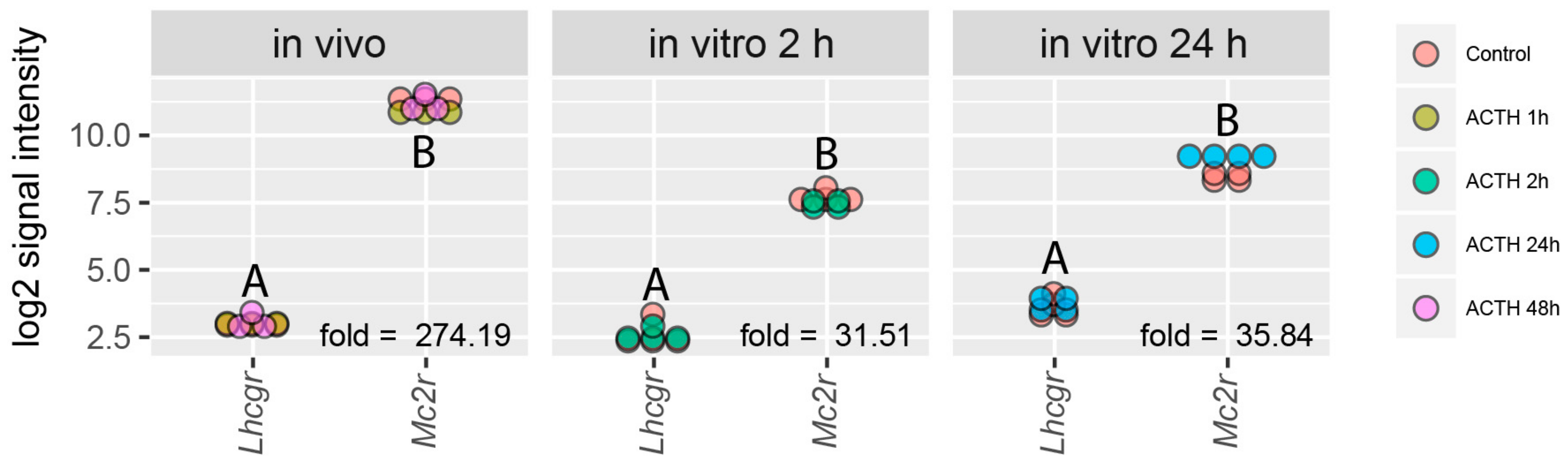
2.1. Analysis of RNA-Seq Dataset Obtained from Adrenals of Male and Female Rats during Postnatal Ontogenesis
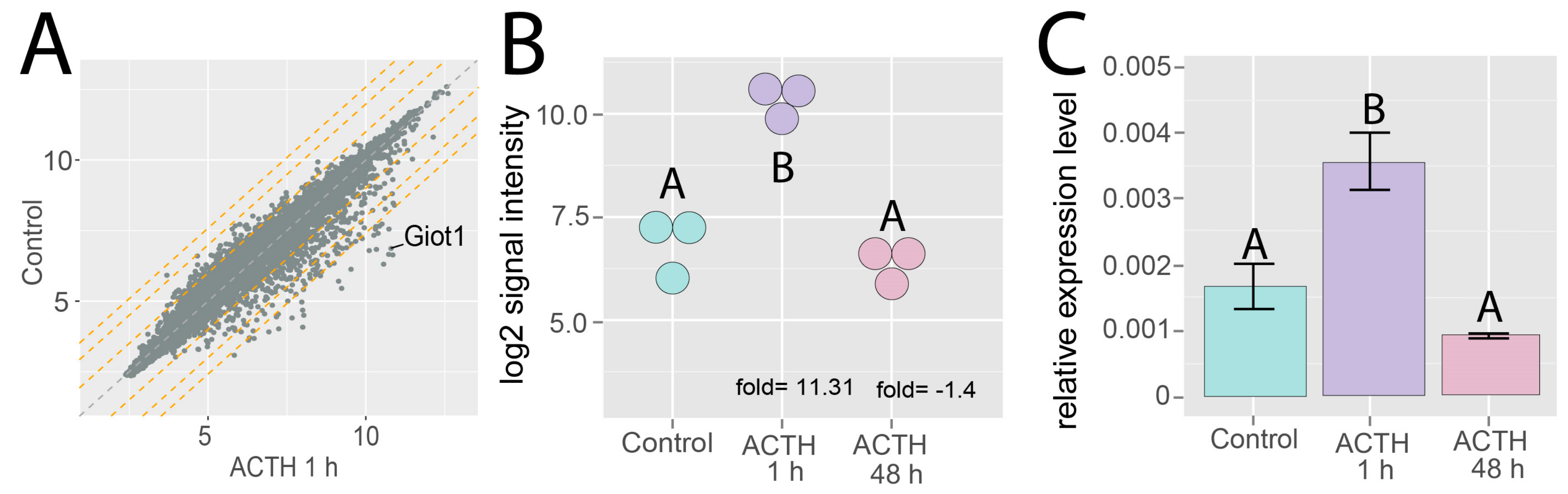
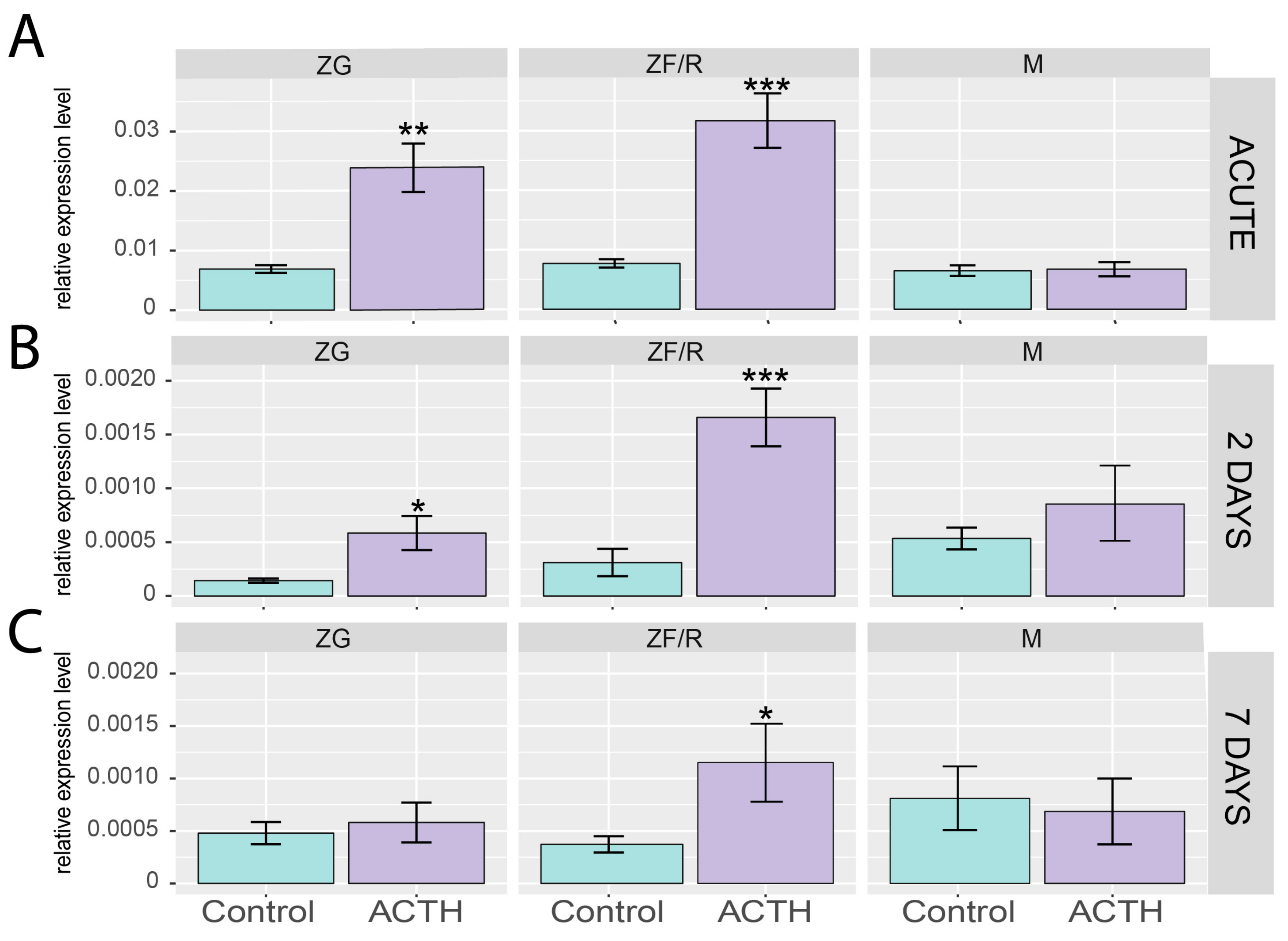
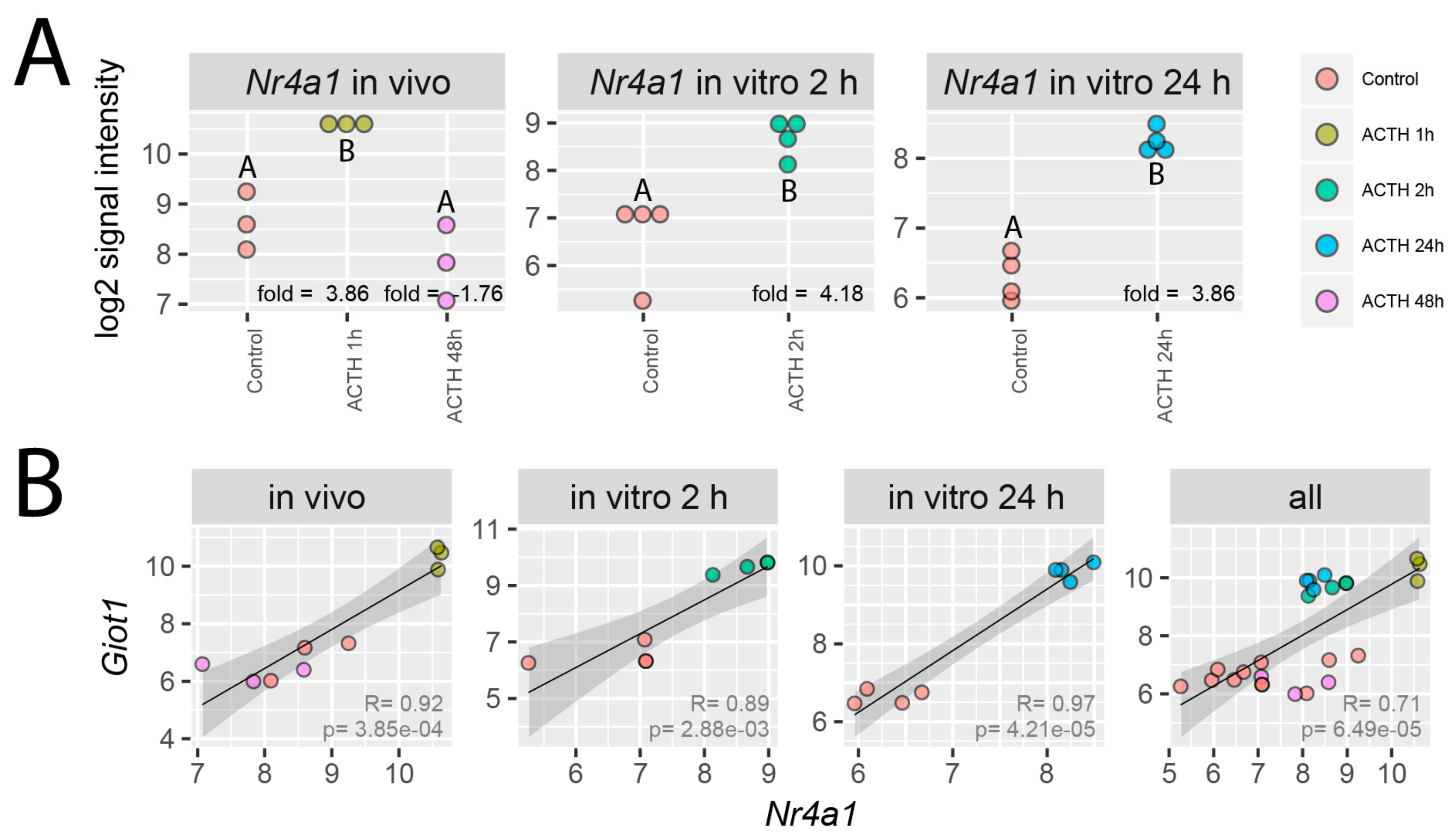
2.2. Effects of ACTH on Giot1 mRNA Level in Rat Adrenal Glands
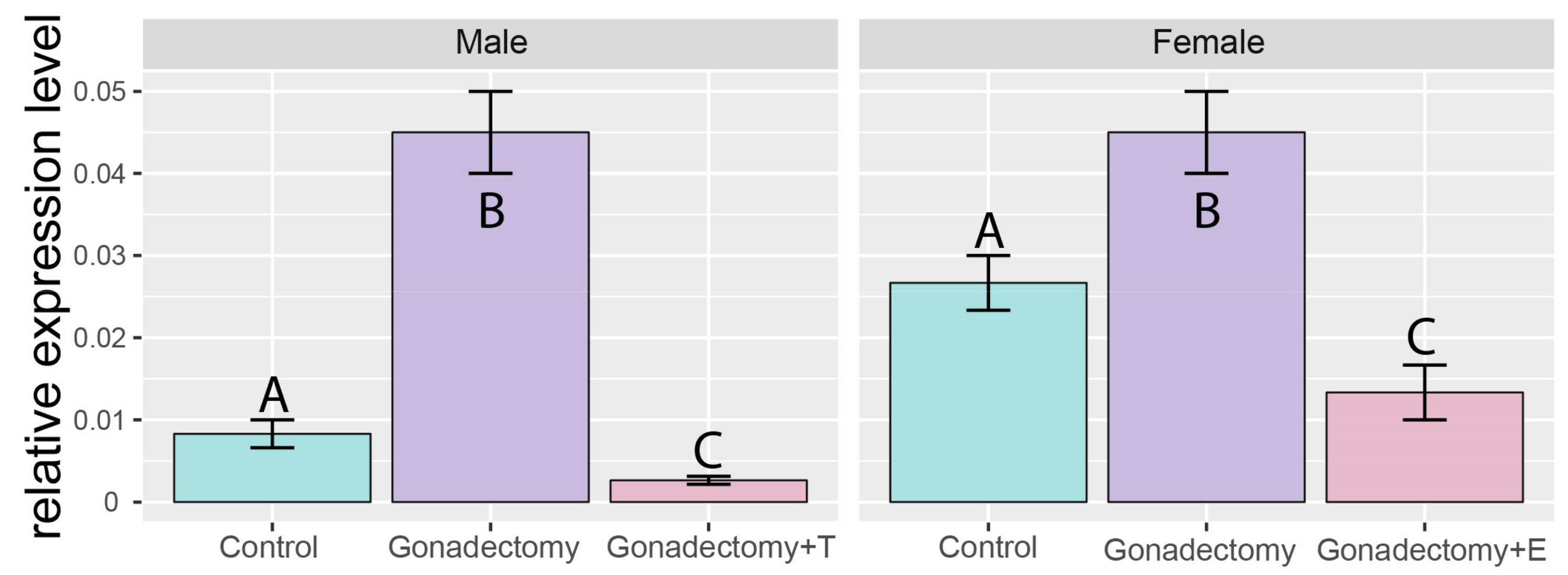
2.3. Effects of Gonadectomy and Testosterone or Estradiol Replacement on the Expression of the Giot1 Gene in Rat Adrenal Gland
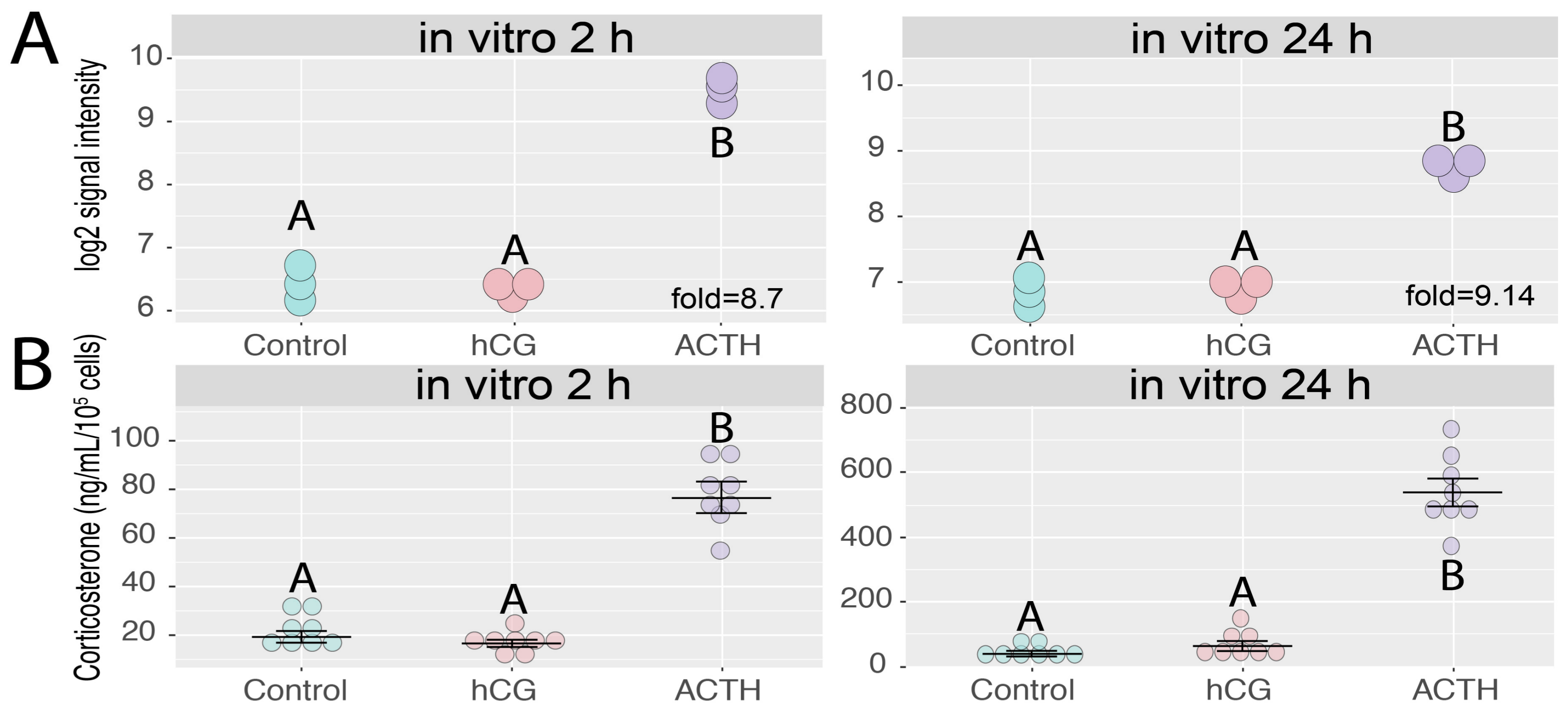
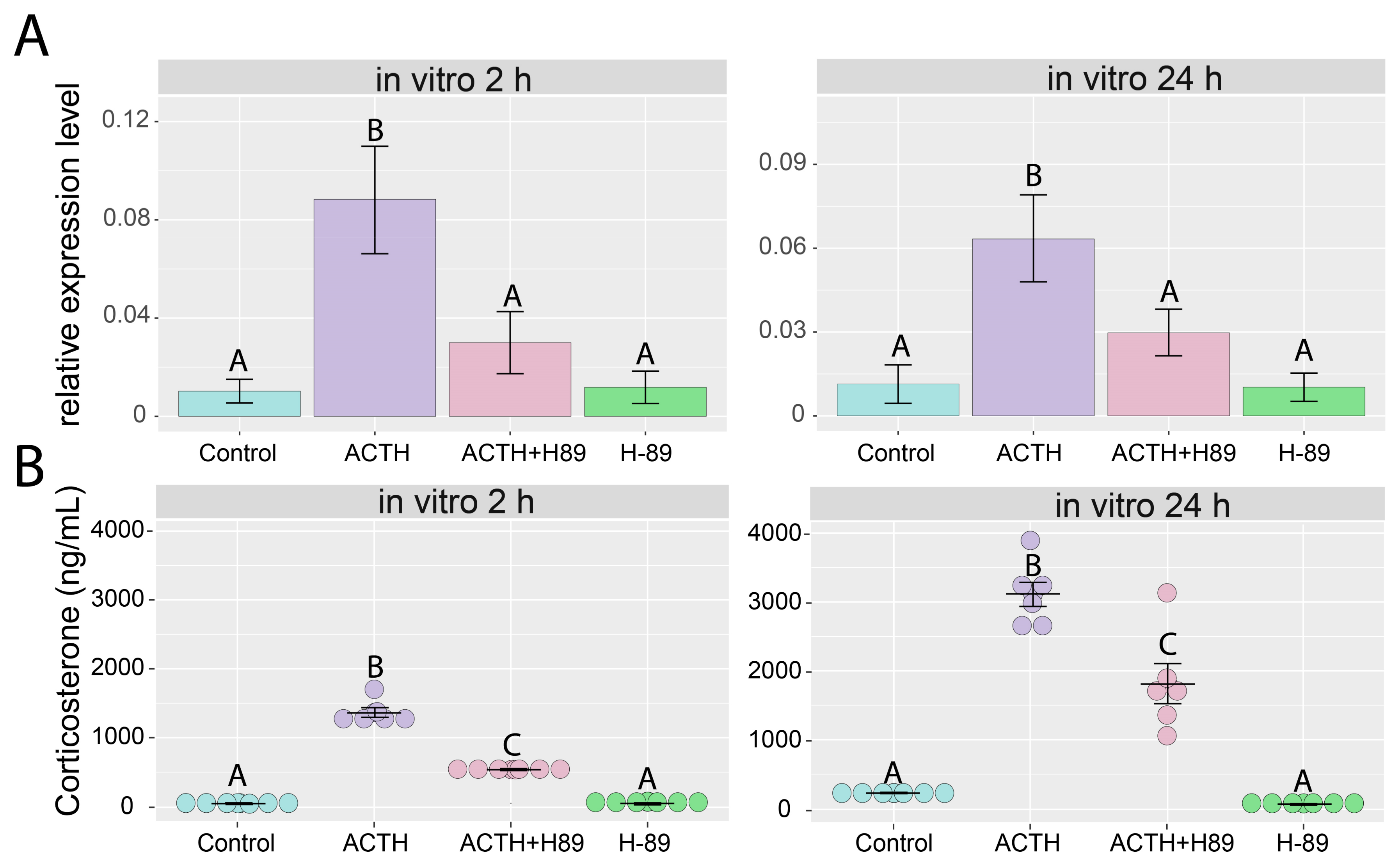
2.4. Effects of ACTH and hCG on the Expression of Giot1 Gene in Rat Primary Adrenocortical Cells
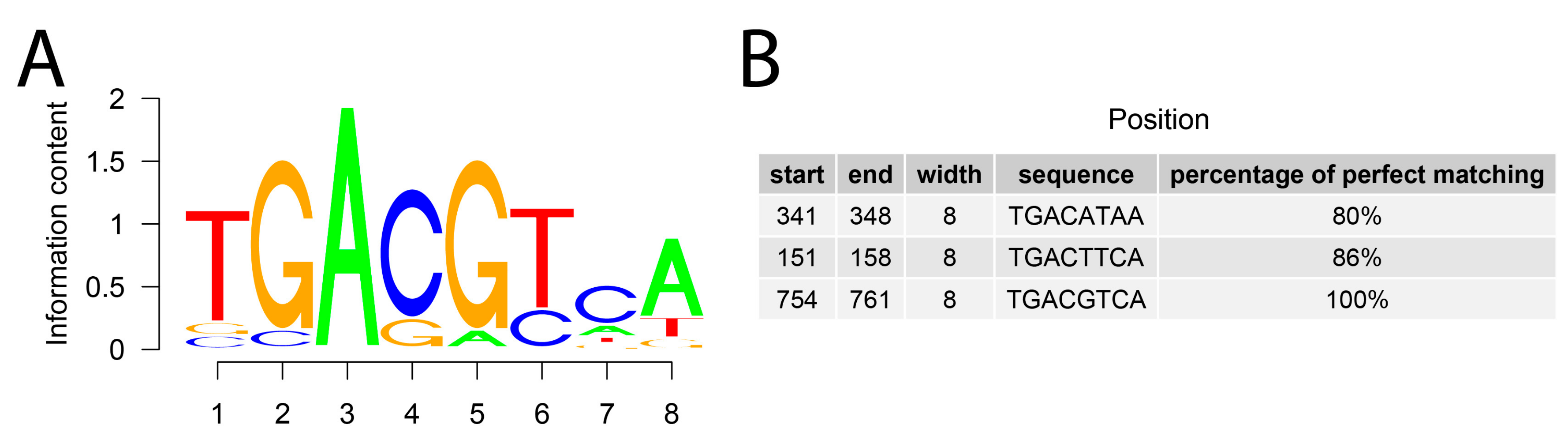
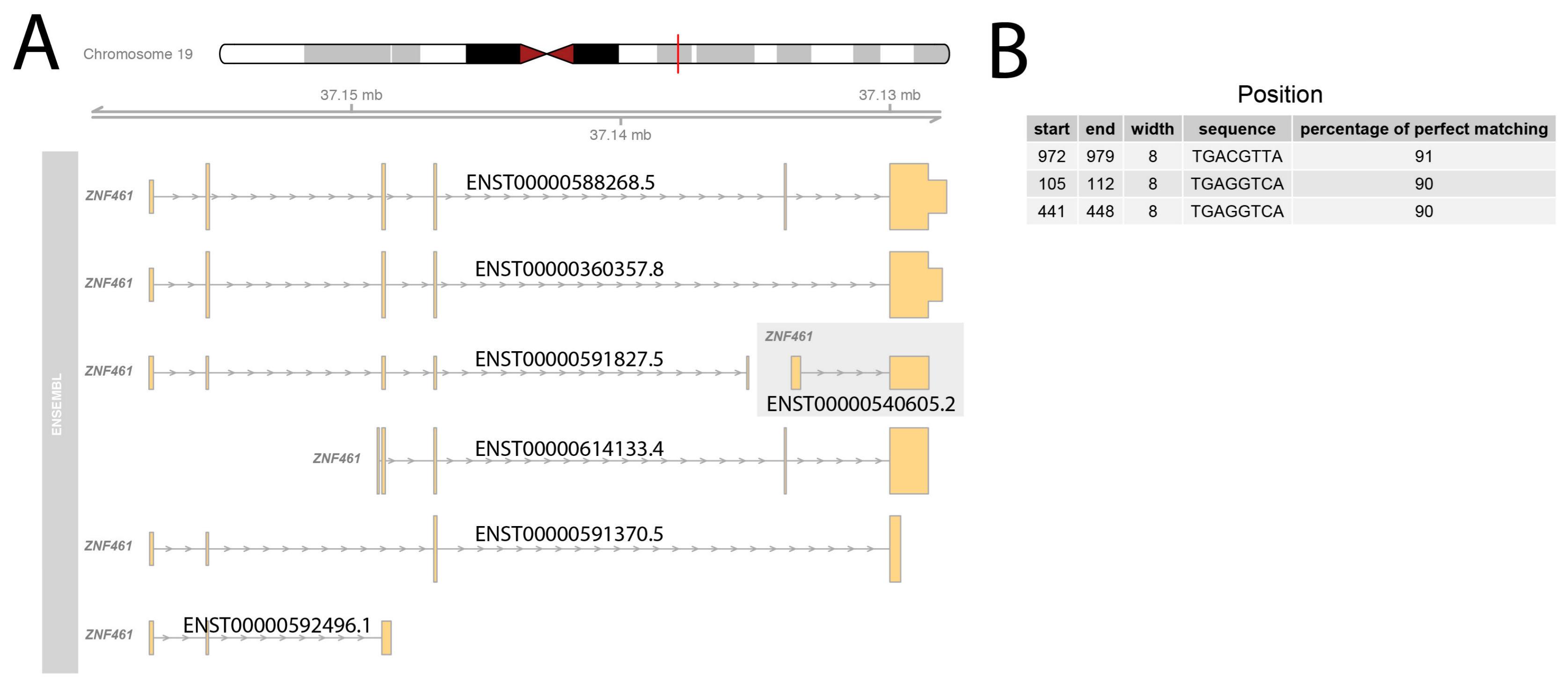
2.5. Bioinformatic Ananalyses Concerning the Other Species
3. Discussion
4. Materials and Methods
4.1. Animals and Reagents
4.2. In Vivo Experiments
4.2.1. Effects of ACTH on Giot1 mRNA Level in Rat Adrenal Glands
4.2.2. Effects of Gonadectomy and Testosteone or Estradiol Replacement on the Expression of the Giot1 Gene in the Adrenal Gland of the Rat
4.3. Primary Adrenocortical Cell Culture
4.4. RNA Isolation
4.5. Reverse Transcription
4.6. Microarray RNA Analysis
4.7. qPCR
4.8. Promoter Analysis
4.9. Multiple Sequence Alignment
4.10. Analysis of RNA-seq Dataset Obtained from Adrenals of Male and Female rats During Postnatal Ontogenesis
4.11. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ACTH | Adrenocorticotropic hormone |
| HCG | Human chorionic gonadotropin |
| GIOT1 | Gonadotropin-inducible ovarian transcription factor-1 |
| cAMP | Cyclic adenosine monophosphate |
| Star | Steroidogenic acute regulatory protein |
| Cyp11a1 | Cytochrome P450 family 11 subfamily A member 1 |
| HSD3B2 | 3β-hydroxysteroid dehydrogenase type 2 |
| SF-1 | Steroidogenic factor-1 |
| DAX-1 | Dosage-sensitive sex reversal, adrenal hypoplasia critical region, on chromosome X, gene 1 |
| PMSG | Pregnant mare’s serum gonadotropin |
| FSH | Follicle-stimulating hormone |
| PKA | Protein kinase A |
| CRE | cAMP response element |
| LHCGR | Luteinizing hormone/choriogonadotropin receptor |
| MC2R | Melanocortin 2 receptor |
| RPKM | Reads Per KilobaseMilion |
| LH | Luteinizing hormone |
| ORX | Orchiectomy |
| OVX | Ovariectomy |
| CREB1 | cAMP responsive element binding protein 1 |
| KRAB-A | Krüppel associated box- A |
| ZF/R | Zona fasciculata/reticularis |
| ZG | Zona glomerulosa |
| M | Medulla |
| ATP | Adenosine triphosphate |
| HPA | Hypothalamic-pituitary-adrenal |
| CRH | Corticotropin-releasing hormone |
| DHEA | Dehydroepiandrosterone |
| NGFIB | Nerve growth factor IB |
| CYP11B2 | Cytochrome P450 family 11 subfamily B member 2 |
| FBS | Fetal bovine serum |
| B2m | Beta-2 microglobulin |
| RMAZNF461 | Robust Multiarray AveragingZinc finger protein 461 |
References
- Morohashi, K. The ontogenesis of the steroidogenic tissues. Genes Cells 1997, 2, 95–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keegan, C.E.; Hammer, G.D. Recent insights into organogenesis of the adrenal cortex. Trends Endocrinol. Metab. 2002, 13, 200–208. [Google Scholar] [CrossRef]
- Bernichtein, S.; Alevizaki, M.; Huhtaniemi, I. Is the adrenal cortex a target for gonadotropins? Trends Endocrinol. Metab. 2008, 19, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Maniatis, T.; Goodbourn, S.; Fischer, J.A. Regulation of inducible and tissue-specific gene expression. Science 1987, 236, 1237–1245. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, P.J.; Tjian, R. Transcriptional regulation in mammalian cells by sequence-specific DNA binding proteins. Science 1989, 245, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Ptashne, M.; Gann, A.A. Activators and targets. Nature 1990, 346, 329–331. [Google Scholar] [CrossRef] [PubMed]
- Yazawa, T.; Mizutani, T.; Yamada, K.; Kawata, H.; Sekiguchi, T.; Yoshino, M.; Kajitani, T.; Shou, Z.; Miyamoto, K. Involvement of cyclic adenosine 5′-monophosphate response element-binding protein, steroidogenic factor 1, and Dax-1 in the regulation of gonadotropin-inducible ovarian transcription factor 1 gene expression by follicle-stimulating hormone in ovarian granulosa cells. Endocrinology 2003, 144, 1920–1930. [Google Scholar] [PubMed]
- Mizutani, T.; Yamada, K.; Yazawa, T.; Okada, T.; Minegishi, T.; Miyamoto, K. Cloning and characterization of gonadotropin-inducible ovarian transcription factors (Giot1 and -2) that are novel members of the (Cys)(2)-(His)(2)-type zinc finger protein family. Mol. Endocrinol. 2001, 15, 1693–1705. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.; Yao, S.; Hindmarch, C.; Antunes, V.; Paton, J.; Murphy, D. Transcription factor expression in the hypothalamo-neurohypophyseal system of the dehydrated rat: Upregulation of gonadotrophin inducible transcription factor 1 mRNA is mediated by cAMP-dependent protein kinase A. J. Neurosci. 2007, 27, 2196–2203. [Google Scholar] [CrossRef] [PubMed]
- Morohashi, K.I.; Omura, T. Ad4BP/SF-1, a transcription factor essential for the transcription of steroidogenic cytochrome P450 genes and for the establishment of the reproductive function. FASEB J. 1996, 10, 1569–1577. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, Y.; Swain, A.; Weber, T.J.; Hentges, K.E.; Zanaria, E.; Lalli, E.; Tamai, K.T.; Sassone-Corsi, P.; Lovell-Badge, R.; Camerino, G.; et al. Steroidogenic factor 1 and Dax-1 colocalize in multiple cell lineages: Potential links in endocrine development. Mol. Endocrinol. 1996, 10, 1261–1272. [Google Scholar] [PubMed]
- Song, K.H.; Park, Y.Y.; Kee, H.J.; Hong, C.Y.; Lee, Y.S.; Ahn, S.W.; Kim, H.J.; Lee, K.; Kook, H.; Lee, I.K.; et al. Orphan nuclear receptor Nur77 induces zinc finger protein GIOT-1 gene expression, and GIOT-1 acts as a novel corepressor of orphan nuclear receptor SF-1 via recruitment of HDAC2. J. Biol. Chem. 2006, 281, 15605–15614. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Zhao, C.; Su, Z.; Wang, C.; Fuscoe, J.C.; Tong, W.; Shi, L. Comprehensive RNA-Seq transcriptomic profiling across 11 organs, 4 ages, and 2 sexes of Fischer 344 rats. Sci. Data 2014, 1, 140013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trejter, M.; Hochol, A.; Tyczewska, M.; Ziolkowska, A.; Jopek, K.; Szyszka, M.; Malendowicz, L.K.; Rucinski, M. Sex-related gene expression profiles in the adrenal cortex in the mature rat: Microarray analysis with emphasis on genes involved in steroidogenesis. Int. J. Mol. Med. 2015, 35, 702–714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruggiero, C.; Lalli, E. Impact of ACTH Signaling on Transcriptional Regulation of Steroidogenic Genes. Front. Endocrinol. (Lausanne) 2016, 7, 24. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Li, Y.; Ji, C.; Jin, F.; Zheng, Z.; Wang, X.; Sun, X.; Xu, X.; Gu, S.; Xie, Y.; et al. Characterization of two novel KRAB-domain-containing zinc finger genes, ZNF460 and ZNF461, on human chromosome 19q13.1-->q13.4. Cytogenet. Genome Res. 2003, 103, 74–78. [Google Scholar] [CrossRef] [PubMed]
- Margolin, J.F.; Friedman, J.R.; Meyer, W.K.; Vissing, H.; Thiesen, H.J.; Rauscher, F.J., III. Kruppel-associated boxes are potent transcriptional repression domains. Proc. Natl. Acad. Sci. USA 1994, 91, 4509–4513. [Google Scholar] [CrossRef] [PubMed]
- Witzgall, R.; O’Leary, E.; Leaf, A.; Onaldi, D.; Bonventre, J.V. The Kruppel-associated box-A (KRAB-A) domain of zinc finger proteins mediates transcriptional repression. Proc. Natl. Acad. Sci. USA 1994, 91, 4514–4518. [Google Scholar] [CrossRef] [PubMed]
- Gorrigan, R.J.; Guasti, L.; King, P.; Clark, A.J.; Chan, L.F. Localisation of the melanocortin-2-receptor and its accessory proteins in the developing and adult adrenal gland. J. Mol. Endocrinol. 2011, 46, 227–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinson, G.P. Functional Zonation of the Adult Mammalian Adrenal Cortex. Front. Neurosci. 2016, 10, 238. [Google Scholar] [CrossRef] [PubMed]
- Handa, R.J.; Weiser, M.J. Gonadal steroid hormones and the hypothalamo-pituitary-adrenal axis. Front. Neuroendocrinol. 2014, 35, 197–220. [Google Scholar] [CrossRef] [PubMed]
- Goel, N.; Workman, J.L.; Lee, T.T.; Innala, L.; Viau, V. Sex differences in the HPA axis. Compr. Physiol. 2014, 4, 1121–1155. [Google Scholar] [PubMed]
- Lesniewska, B.; Miskowiak, B.; Nowak, M.; Malendowicz, L.K. Sex differences in adrenocortical structure and function. XXVII. The effect of ether stress on ACTH and corticosterone in intact, gonadectomized, and testosterone- or estradiol-replaced rats. Res. Exp. Med. (Berl.) 1990, 190, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Schillebeeckx, M.; Pihlajoki, M.; Gretzinger, E.; Yang, W.; Thol, F.; Hiller, T.; Lobs, A.K.; Rohrig, T.; Schrade, A.; Cochran, R.; et al. Novel markers of gonadectomy-induced adrenocortical neoplasia in the mouse and ferret. Mol. Cell. Endocrinol. 2015, 399, 122–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jopek, K.; Celichowski, P.; Szyszka, M.; Tyczewska, M.; Milecka, P.; Malendowicz, L.K.; Rucinski, M. Transcriptome Profile of Rat Adrenal Evoked by Gonadectomy and Testosterone or Estradiol Replacement. Front. Endocrinol. (Lausanne) 2017, 8, 26. [Google Scholar] [CrossRef] [PubMed]
- Parker, L.N.; Odell, W.D. Control of adrenal androgen secretion. Endocr. Rev. 1980, 1, 392–410. [Google Scholar] [CrossRef] [PubMed]
- Seron-Ferre, M.; Lawrence, C.C.; Jaffe, R.B. Role of hCG in regulation of the fetal zone of the human fetal adrenal gland. J. Clin. Endocrinol. Metab. 1978, 46, 834–837. [Google Scholar] [CrossRef] [PubMed]
- Keymolen, V.; Dor, P.; Borkowski, A. Output of oestrogens, testosterone and their precursors by isolated human adrenal cells as compared with that of glucocorticosteroids. J. Endocrinol. 1976, 71, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Apter, D.; Pakarinen, A.; Hammond, G.L.; Vihko, R. Adrenocortical function in puberty. serum ACTH, cortisol and dehydroepiandrosterone in girls and boys. Acta Paediatr. Scand. 1979, 68, 599–604. [Google Scholar] [CrossRef] [PubMed]
- O’Connell, Y.; McKenna, T.J.; Cunningham, S.K. The effect of prolactin, human chorionic gonadotropin, insulin and insulin-like growth factor 1 on adrenal steroidogenesis in isolated guinea-pig adrenal cells. J. Ster. Biochem. Mol. Biol. 1994, 48, 235–240. [Google Scholar] [CrossRef]
- Kero, J.; Poutanen, M.; Zhang, F.P.; Rahman, N.; McNicol, A.M.; Nilson, J.H.; Keri, R.A.; Huhtaniemi, I.T. Elevated luteinizing hormone induces expression of its receptor and promotes steroidogenesis in the adrenal cortex. J. Clin. Investig. 2000, 105, 633–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Couzinet, B.; Meduri, G.; Lecce, M.G.; Young, J.; Brailly, S.; Loosfelt, H.; Milgrom, E.; Schaison, G. The postmenopausal ovary is not a major androgen-producing gland. J. Clin. Endocrinol. Metab. 2001, 86, 5060–5066. [Google Scholar] [CrossRef] [PubMed]
- Pabon, J.E.; Li, X.; Lei, Z.M.; Sanfilippo, J.S.; Yussman, M.A.; Rao, C.V. Novel presence of luteinizing hormone/chorionic gonadotropin receptors in human adrenal glands. J. Clin. Endocrinol. Metab. 1996, 81, 2397–2400. [Google Scholar] [PubMed]
- Dall’Asta, C.; Ballare, E.; Mantovani, G.; Ambrosi, B.; Spada, A.; Barbetta, L.; Colombo, P.; Travaglini, P.; Loli, P.; Beck-Peccoz, P. Assessing the presence of abnormal regulation of cortisol secretion by membrane hormone receptors: In vivo and in vitro studies in patients with functioning and non-functioning adrenal adenoma. Horm. Metab. Res. 2004, 36, 578–583. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, M.A.; Lei, Z.M.; Li, X.; Greenwold, N.; Nakajima, S.T.; Jauniaux, E.; Rao Ch, V. Human fetal nongonadal tissues contain human chorionic gonadotropin/luteinizing hormone receptors. J. Clin. Endocrinol. Metab. 2004, 89, 952–956. [Google Scholar] [CrossRef] [PubMed]
- Feelders, R.A.; Lamberts, S.W.; Hofland, L.J.; van Koetsveld, P.M.; Verhoef-Post, M.; Themmen, A.P.; de Jong, F.H.; Bonjer, H.J.; Clark, A.J.; van der Lely, A.J.; et al. Luteinizing hormone (LH)-responsive Cushing’s syndrome: The demonstration of LH receptor messenger ribonucleic acid in hyperplastic adrenal cells, which respond to chorionic gonadotropin and serotonin agonists in vitro. J. Clin. Endocrinol. Metab. 2003, 88, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Bugalho, M.J.; Li, X.; Rao, C.V.; Soares, J.; Sobrinho, L.G. Presence of a Gs alpha mutation in an adrenal tumor expressing LH/hCG receptors and clinically associated with Cushing’s syndrome. Gynecol. Endocrinol. 2000, 14, 50–54. [Google Scholar] [CrossRef] [PubMed]
- Miyamura, N.; Taguchi, T.; Murata, Y.; Taketa, K.; Iwashita, S.; Matsumoto, K.; Nishikawa, T.; Toyonaga, T.; Sakakida, M.; Araki, E. Inherited adrenocorticotropin-independent macronodular adrenal hyperplasia with abnormal cortisol secretion by vasopressin and catecholamines: Detection of the aberrant hormone receptors on adrenal gland. Endocrine 2002, 19, 319–326. [Google Scholar] [CrossRef]
- Miyamura, N.; Tsutsumi, A.; Senokuchi, H.; Nakamaru, K.; Kawashima, J.; Sakai, K.; Taguchi, T.; Tokunaga, H.; Nishida, K.; Uehara, M.; et al. A case of ACTH-independent macronodular adrenal hyperplasia: Simultaneous expression of several aberrant hormone receptors in the adrenal gland. Endocr. J. 2003, 50, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Wy, L.A.; Carlson, H.E.; Kane, P.; Li, X.; Lei, Z.M.; Rao, C.V. Pregnancy-associated Cushing’s syndrome secondary to a luteinizing hormone/human chorionic gonadotropin receptor-positive adrenal carcinoma. Gynecol. Endocrinol. 2002, 16, 413–417. [Google Scholar] [CrossRef] [PubMed]
- Waterman, M.R.; Bischof, L.J. Cytochromes P450 12: Diversity of ACTH (cAMP)-dependent transcription of bovine steroid hydroxylase genes. FASEB J. 1997, 11, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Schimmer, B.P.; Cordova, M.; Cheng, H.; Tsao, A.; Goryachev, A.B.; Schimmer, A.D.; Morris, Q. Global profiles of gene expression induced by adrenocorticotropin in Y1 mouse adrenal cells. Endocrinology 2006, 147, 2357–2367. [Google Scholar] [CrossRef] [PubMed]
- Song, K.H.; Park, J.I.; Lee, M.O.; Soh, J.; Lee, K.; Choi, H.S. LH induces orphan nuclear receptor Nur77 gene expression in testicular Leydig cells. Endocrinology 2001, 142, 5116–5123. [Google Scholar] [CrossRef] [PubMed]
- Bassett, M.H.; Suzuki, T.; Sasano, H.; De Vries, C.J.; Jimenez, P.T.; Carr, B.R.; Rainey, W.E. The orphan nuclear receptor NGFIB regulates transcription of 3beta-hydroxysteroid dehydrogenase. implications for the control of adrenal functional zonation. J. Biol. Chem. 2004, 279, 37622–37630. [Google Scholar] [CrossRef] [PubMed]
- Shimada, H.; Kogure, N.; Noro, E.; Kudo, M.; Sugawara, K.; Sato, I.; Shimizu, K.; Kobayashi, M.; Suzuki, D.; Parvin, R.; et al. High glucose stimulates expression of aldosterone synthase (CYP11B2) and secretion of aldosterone in human adrenal cells. FEBS Open Bio 2017, 7, 1410–1421. [Google Scholar] [CrossRef] [PubMed]
- Sadovsky, Y.; Crawford, P.A.; Woodson, K.G.; Polish, J.A.; Clements, M.A.; Tourtellotte, L.M.; Simburger, K.; Milbrandt, J. Mice deficient in the orphan receptor steroidogenic factor 1 lack adrenal glands and gonads but express P450 side-chain-cleavage enzyme in the placenta and have normal embryonic serum levels of corticosteroids. Proc. Natl. Acad. Sci. USA 1995, 92, 10939–10943. [Google Scholar] [CrossRef] [PubMed]
- Naville, D.; Penhoat, A.; Marchal, R.; Durand, P.; Begeot, M. SF-1 and the transcriptional regulation of the human ACTH receptor gene. Endocr. Res. 1998, 24, 391–395. [Google Scholar] [CrossRef] [PubMed]
- Caron, K.M.; Ikeda, Y.; Soo, S.C.; Stocco, D.M.; Parker, K.L.; Clark, B.J. Characterization of the promoter region of the mouse gene encoding the steroidogenic acute regulatory protein. Mol. Endocrinol. 1997, 11, 138–147. [Google Scholar] [CrossRef] [PubMed]
- Ozisik, G.; Achermann, J.C.; Meeks, J.J.; Jameson, J.L. SF1 in the development of the adrenal gland and gonads. Horm. Res. 2003, 59 (Suppl. 1), 94–98. [Google Scholar] [CrossRef] [PubMed]
- Trejter, M.; Hochol, A.; Tyczewska, M.; Ziolkowska, A.; Jopek, K.; Szyszka, M.; Malendowicz, L.K.; Rucinski, M. Visinin-like peptide 1 in adrenal gland of the rat. Gene expression and its hormonal control. Peptides 2015, 63, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Cagen, L.M.; Baer, P.G. Effects of gonadectomy and steroid treatment on renal prostaglandin 9-ketoreductase activity in the rat. Life Sci. 1987, 40, 95–100. [Google Scholar] [CrossRef]
- Longhurst, P.A.; Kauer, J.; Leggett, R.E.; Levin, R.M. The influence of ovariectomy and estradiol replacement on urinary bladder function in rats. J. Urol. 1992, 148, 915–919. [Google Scholar] [CrossRef]
- Udagawa, K.; Ogawa, T.; Watanabe, T.; Tamura, Y.; Kita, K.; Kubota, Y. Testosterone administration promotes regeneration of chemically impaired spermatogenesis in rats. Int. J. Urol. 2006, 13, 1103–1108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulte-Beerbuhl, M.; Nieschlag, E. Comparison of testosterone, dihydrotestosterone, luteinizing hormone, and follicle-stimulating hormone in serum after injection of testosterone enanthate of testosterone cypionate. Fertil. Steril. 1980, 33, 201–203. [Google Scholar] [CrossRef]
- Ziolkowska, A.; Macchi, C.; Trejter, M.; Rucinski, M.; Nowak, M.; Nussdorfer, G.G.; Malendowicz, L.K. Effects of neuromedin-U on immature rat adrenocortical cells: In vitro and in vivo studies. Int. J. Mol. Med. 2008, 21, 303–307. [Google Scholar] [CrossRef] [PubMed]
- Tyczewska, M.; Rucinski, M.; Trejter, M.; Ziolkowska, A.; Szyszka, M.; Malendowicz, L.K. Angiogenesis in the course of enucleation-induced adrenal regeneration—Expression of selected genes and proteins involved in development of capillaries. Peptides 2012, 38, 404–413. [Google Scholar] [CrossRef] [PubMed]
- Tyczewska, M.; Rucinski, M.; Ziolkowska, A.; Szyszka, M.; Trejter, M.; Hochol-Molenda, A.; Nowak, K.W.; Malendowicz, L.K. Enucleation-induced rat adrenal gland regeneration: Expression profile of selected genes involved in control of adrenocortical cell proliferation. Int. J. Endocrinol. 2014, 2014, 130359. [Google Scholar] [CrossRef] [PubMed]
- Jopek, K.; Tyczewska, M.; Celichowski, P.; Malendowicz, L.K.; Rucinski, M. Transcriptome Profile in Unilateral Adrenalectomy-Induced Compensatory Adrenal Growth in the Rat. Int. J. Mol. Sci. 2018, 19, 1111. [Google Scholar] [CrossRef] [PubMed]
- Gentleman, R.C.; Carey, V.J.; Bates, D.M.; Bolstad, B.; Dettling, M.; Dudoit, S.; Ellis, B.; Gautier, L.; Ge, Y.; Gentry, J.; et al. Bioconductor: Open software development for computational biology and bioinformatics. Genome Biol. 2004, 5, R80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gautier, L.; Cope, L.; Bolstad, B.M.; Irizarry, R.A. affy--analysis of Affymetrix GeneChip data at the probe level. Bioinformatics 2004, 20, 307–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rucinski, M.; Albertin, G.; Spinazzi, R.; Ziolkowska, A.; Nussdorfer, G.G.; Malendowicz, L.K. Cerebellin in the rat adrenal gland: Gene expression and effects of CER and [des-Ser1] CER on the secretion and growth of cultured adrenocortical cells. Int. J. Mol. Med. 2005, 15, 411–415. [Google Scholar] [CrossRef] [PubMed]
- Rucinski, M.; Andreis, P.G.; Ziolkowska, A.; Nussdorfer, G.G.; Malendowicz, L.K. Differential expression and function of beacon in the rat adrenal cortex and medulla. Int. J. Mol. Med. 2005, 16, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Rucinski, M.; Malendowicz, L.K. Precerebellin-related genes and precerebellin 1 peptide in endocrine glands of the rat—Pattern of their expression. Int. J. Mol. Med. 2009, 23, 113–119. [Google Scholar] [PubMed]
- Tyczewska, M.; Rucinski, M.; Ziolkowska, A.; Trejter, M.; Szyszka, M.; Malendowicz, L.K. Expression of selected genes involved in steroidogenesis in the course of enucleation-induced rat adrenal regeneration. Int. J. Mol. Med. 2014, 33, 613–623. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Genbank Accession Number | Primer | Primer Sequence (5′-3′) | Position | PCR Product Size (bp) |
|---|---|---|---|---|---|
| B2m | NM_012512.2 | S A | CTTGCAGAGTTAAACACGTCA CTTGATTACATGTCTCGGTC | 316-336 366-385 | 70 |
| Giot1 | NM_133563.1 | S A | AATAGGAGGGGACACTTCCG CATCCTCATAGGTGACTGCAT | 152-171 294-314 | 163 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jopek, K.; Tyczewska, M.; Ramanjaneya, M.; Szyszka, M.; Celichowski, P.; Milecka, P.; Malendowicz, L.K.; Rucinski, M. Effect of ACTH and hCG on the Expression of Gonadotropin-Inducible Ovarian Transcription Factor 1 (Giot1) Gene in the Rat Adrenal Gland. Int. J. Mol. Sci. 2018, 19, 2285. https://doi.org/10.3390/ijms19082285
Jopek K, Tyczewska M, Ramanjaneya M, Szyszka M, Celichowski P, Milecka P, Malendowicz LK, Rucinski M. Effect of ACTH and hCG on the Expression of Gonadotropin-Inducible Ovarian Transcription Factor 1 (Giot1) Gene in the Rat Adrenal Gland. International Journal of Molecular Sciences. 2018; 19(8):2285. https://doi.org/10.3390/ijms19082285
Chicago/Turabian StyleJopek, Karol, Marianna Tyczewska, Manjunath Ramanjaneya, Marta Szyszka, Piotr Celichowski, Paulina Milecka, Ludwik K. Malendowicz, and Marcin Rucinski. 2018. "Effect of ACTH and hCG on the Expression of Gonadotropin-Inducible Ovarian Transcription Factor 1 (Giot1) Gene in the Rat Adrenal Gland" International Journal of Molecular Sciences 19, no. 8: 2285. https://doi.org/10.3390/ijms19082285
APA StyleJopek, K., Tyczewska, M., Ramanjaneya, M., Szyszka, M., Celichowski, P., Milecka, P., Malendowicz, L. K., & Rucinski, M. (2018). Effect of ACTH and hCG on the Expression of Gonadotropin-Inducible Ovarian Transcription Factor 1 (Giot1) Gene in the Rat Adrenal Gland. International Journal of Molecular Sciences, 19(8), 2285. https://doi.org/10.3390/ijms19082285