Antibody Light Chains Dictate the Specificity of Contact Hypersensitivity Effector Cell Suppression Mediated by Exosomes


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
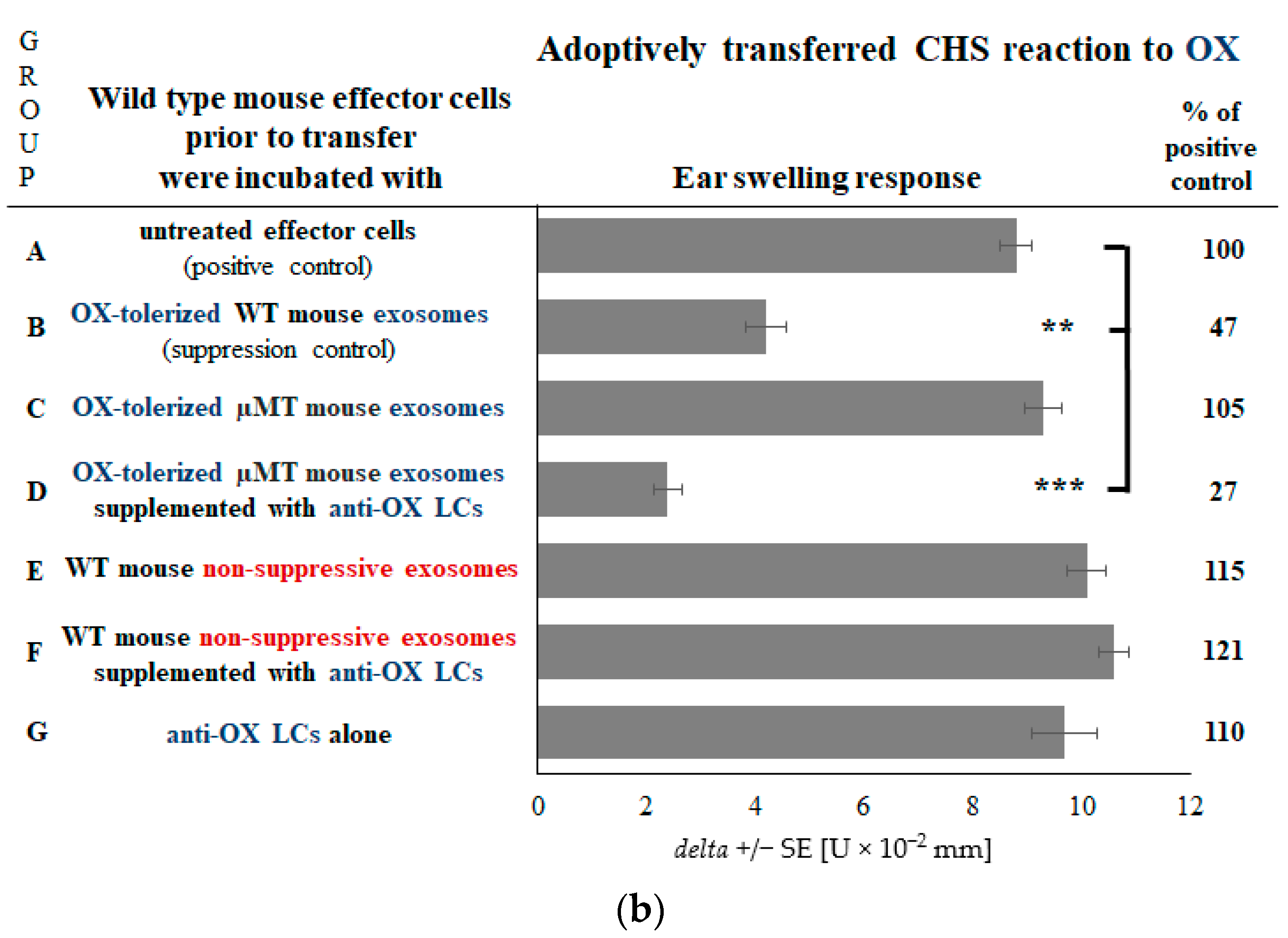
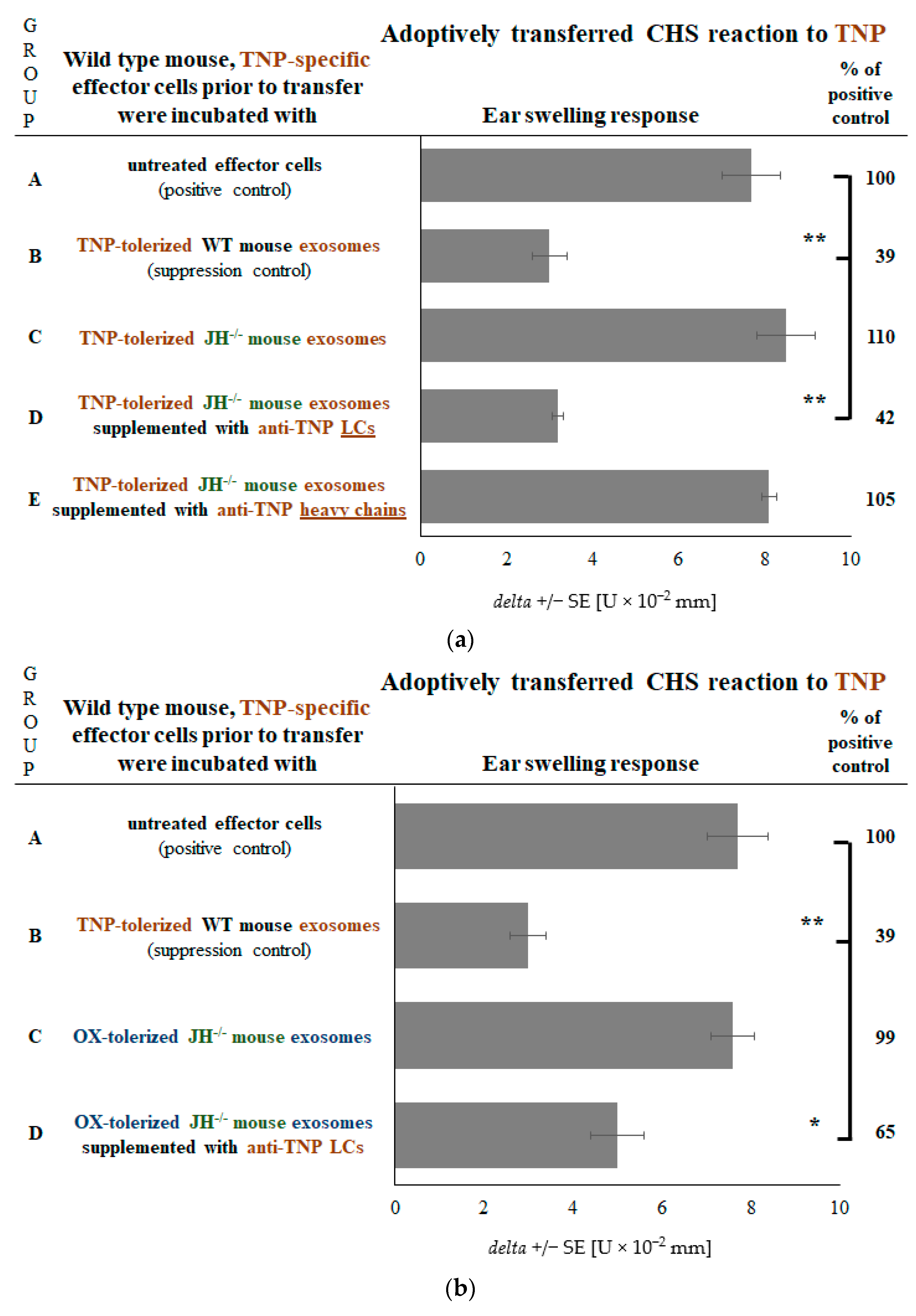
2.1. B Cells Deliver Hapten-Specific LCs for Coating of Ts Cell-Derived Exosomes to Enable Their Suppressive Function
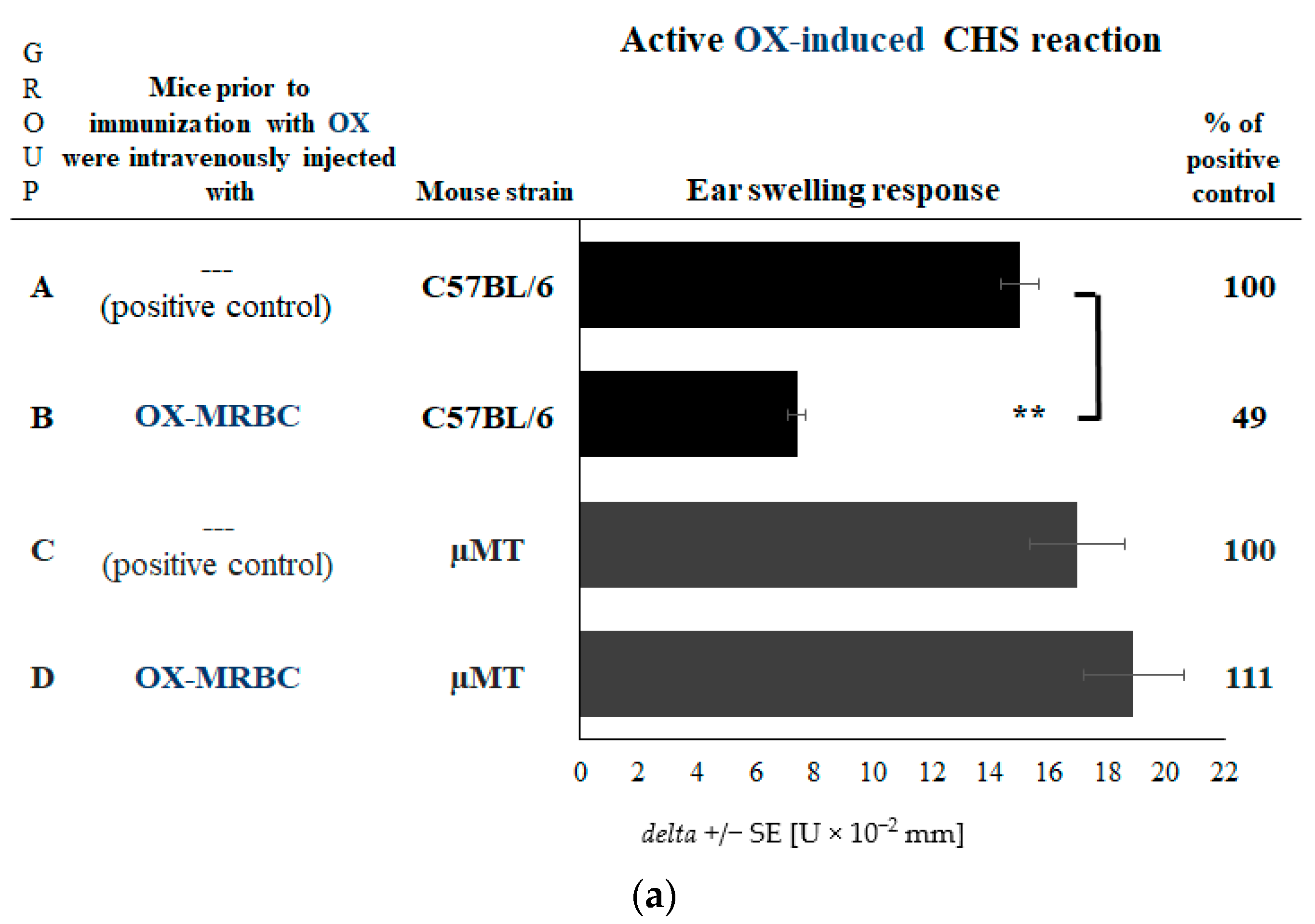
2.2. CHS Reaction Is Not Suppressed in the Absence of NKT Cells
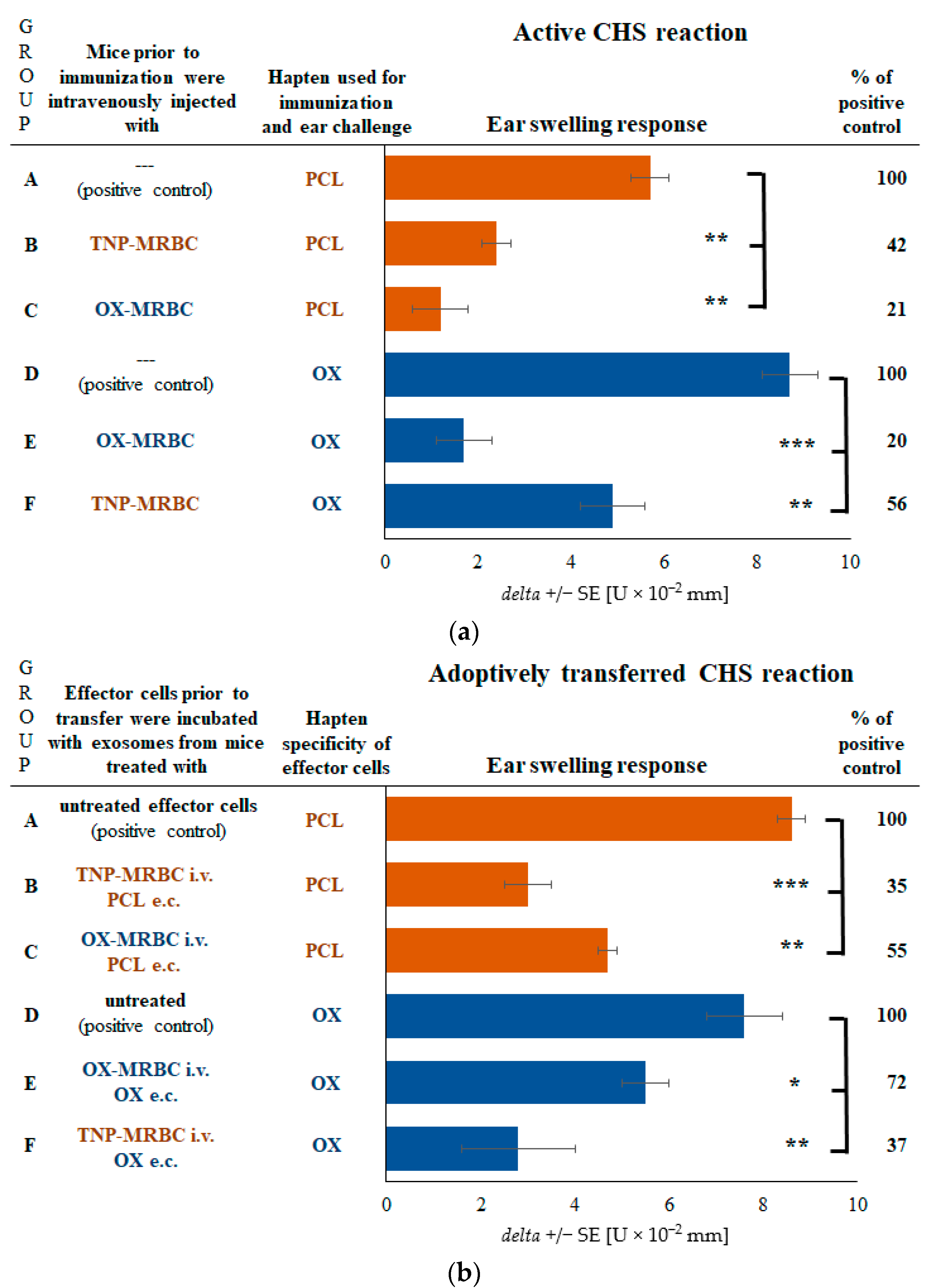
2.3. CHS Reaction Is Suppressed by Exosomes in Mice Tolerized with Syngeneic MRBCs Regardless of Their Hapten Coupling
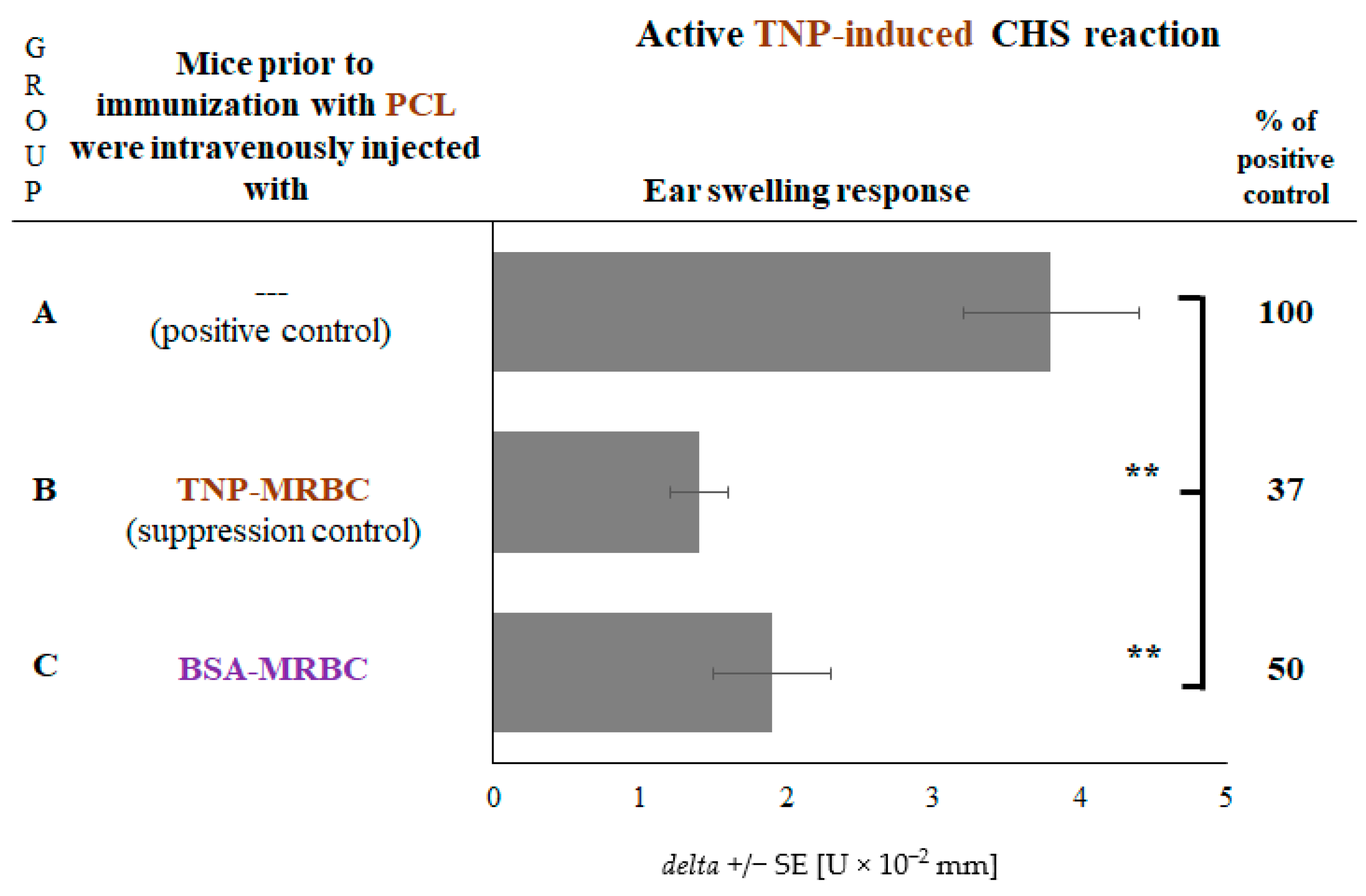
2.4. CHS Reaction to TNP Hapten Is Suppressed in Mice Administered with Non-Self-Protein-Coupled Syngeneic MRBCs
2.5. Coating of Exosomes with LCs Ensures the Antigen-Specificity of Suppression of CHS Effector Cells
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Ethics Statement
4.3. Coupling of Mouse Red Blood Cells with Hapten or Protein Antigen
4.4. Induction of Tolerance by Intravenous Administration of Antigen-Coupled Syngeneic MRBCs Prior to Elicitation of CHS Reaction
4.5. Isolation of Suppressive Exosomes Generated by Suppressor T Cells of Tolerized Mice and Adoptive Transfer of CHS Effector Cells
4.6. Data Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BSA | Bovine serum albumin |
| BSA-MRBCs | Bovine serum albumin-coupled mouse red blood cells |
| CHS | Contact hypersensitivity |
| LCs | Light chains |
| MRBCs | Mouse red blood cells |
| OX | Oxazolone |
| OX-MRBCs | Oxazolone-coupled mouse red blood cells |
| PCL | Picryl chloride (trinitrophenol chloride) |
| TNBSA | 2,4,6-trinitrobenzene sulfonic acid |
| TNP | 2,4,6-trinitrophenol |
| TNP-MRBCs | trinitrophenol-coupled mouse red blood cells |
| Ts | T suppressor (cells) |
| WT | Wild type |
References
- Basile, U.; Gulli, F.; Gragnani, L.; Napodano, C.; Pocino, K.; Rapaccini, G.L.; Mussap, M.; Zignego, A.L. Free light chains: Eclectic multipurpose biomarker. J. Immunol. Methods 2017, 451, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Bhole, M.V.; Sadler, R.; Ramasamy, K. Serum-free light-chain assay: Clinical utility and limitations. Ann. Clin. Biochem. 2014, 51, 528–542. [Google Scholar] [CrossRef] [PubMed]
- Jenner, E. Serum free light chains in clinical laboratory diagnostics. Clin. Chim. Acta 2014, 427, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Groot Kormelink, T.; Askenase, P.W.; Redegeld, F.A. Immunobiology of antigen-specific immunoglobulin free light chains in chronic inflammatory diseases. Curr. Pharm. Des. 2012, 18, 2278–2289. [Google Scholar] [CrossRef] [PubMed]
- Groot Kormelink, T.; Powe, D.G.; Kuijpers, S.A.; Abudukelimu, A.; Fens, M.H.; Pieters, E.H.; Kassing van der Ven, W.W.; Habashy, H.O.; Ellis, I.O.; Blokhuis, B.R.; et al. Immunoglobulin free light chains are biomarkers of poor prognosis in basal-like breast cancer and are potential targets in tumor-associated inflammation. Oncotarget 2014, 5, 3159–3167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Askenase, P.W.; Bryniarski, K.; Paliwal, V.; Redegeld, F.; Groot Kormelink, T.; Kerfoot, S.; Hutchinson, A.T.; van Loveren, H.; Campos, R.; Itakura, A.; et al. A subset of AID-dependent B-1a cells initiates hypersensitivity and pneumococcal pneumonia resistance. Ann. N. Y. Acad. Sci. 2015, 1362, 200–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Askenase, P.W.; Szczepanik, M.; Itakura, A.; Kiener, C.; Campos, R.A. Extravascular T-cell recruitment requires initiation begun by Valpha14+ NKT cells and B-1 B cells. Trends Immunol. 2004, 25, 441–449. [Google Scholar] [CrossRef] [PubMed]
- Campos, R.A.; Szczepanik, M.; Itakura, A.; Akahira-Azuma, M.; Sidobre, S.; Kronenberg, M.; Askenase, P.W. Cutaneous immunization rapidly activates liver invariant Valpha14 NKT cells stimulating B-1 B cells to initiate T cell recruitment for elicitation of contact sensitivity. J. Exp. Med. 2003, 198, 1785–1796. [Google Scholar] [CrossRef] [PubMed]
- Campos, R.A.; Szczepanik, M.; Itakura, A.; Lisbonne, M.; Dey, N.; Leite-de-Moraes, M.C.; Askenase, P.W. Interleukin-4-dependent innate collaboration between iNKT cells and B-1 B cells controls adaptative contact sensitivity. Immunology 2006, 117, 536–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campos, R.A.; Szczepanik, M.; Lisbonne, M.; Itakura, A.; Leite-de-Moraes, M.C.; Askenase, P.W. Invariant NKT cells rapidly activated via immunization with diverse contact antigens collaborate in vitro with B-1 cells to initiate contact sensitivity. J. Immunol. 2006, 177, 3686–3694. [Google Scholar] [CrossRef] [PubMed]
- Zemelka-Wiącek, M.; Szczepanik, M. Tc1-mediated contact sensitivity reaction, its mechanism and regulation. Postepy Hig. Med. Dosw. (Online) 2014, 68, 955–969. [Google Scholar] [CrossRef] [PubMed]
- Almishri, W.; Deans, J.; Swain, M.G. Rapid activation and hepatic recruitment of innate-like regulatory B cells after invariant NKT cell stimulation in mice. J. Hepatol. 2015, 63, 943–951. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, N.; Kerfoot, S.M.; Hutchinson, A.T.; Dela Cruz, C.S.; Nakazawa, N.; Szczepanik, M.; Majewska-Szczepanik, M.; Nazimek, K.; Ohana, N.; Bryniarski, K.; et al. Expression of activation-induced cytidine deaminase enhances the clearance of pneumococcal pneumonia: Evidence of a subpopulation of protective anti-pneumococcal B1a cells. Immunology 2016, 147, 97–113. [Google Scholar] [CrossRef] [PubMed]
- Ptak, W.; Rozycka, D.; Rewicka, M. Induction of suppressor cells and cells producing antigen-specific suppressor factors by haptens bound to self carriers. Immunobiology 1980, 156, 400–409. [Google Scholar] [CrossRef]
- Ptak, W.; Nazimek, K.; Askenase, P.W.; Bryniarski, K. From mysterious supernatant entity to miRNA-150 in antigen-specific exosomes: A history of hapten-specific T suppressor factor. Arch. Immunol. Ther. Exp. (Warsz.) 2015, 63, 345–356. [Google Scholar] [CrossRef] [PubMed]
- Bryniarski, K.; Ptak, W.; Jayakumar, A.; Püllmann, K.; Caplan, M.J.; Chairoungdua, A.; Lu, J.; Adams, B.D.; Sikora, E.; Nazimek, K.; et al. Antigen-specific, antibody-coated, exosome-like nanovesicles deliver suppressor T-cell microRNA-150 to effector T cells to inhibit contact sensitivity. J. Allergy Clin. Immunol. 2013, 132, 170–181. [Google Scholar] [CrossRef] [PubMed]
- Bryniarski, K.; Ptak, W.; Martin, E.; Nazimek, K.; Szczepanik, M.; Sanak, M.; Askenase, P.W. Free extracellular miRNA functionally targets cells by transfecting exosomes from their companion cells. PLoS ONE 2015, 10, e0122991. [Google Scholar] [CrossRef] [PubMed]
- Nazimek, K.; Ptak, W.; Nowak, B.; Ptak, M.; Askenase, P.W.; Bryniarski, K. Macrophages play an essential role in antigen-specific immune suppression mediated by T CD8+ cell-derived exosomes. Immunology 2015, 146, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Nazimek, K.; Bryniarski, K.; Askenase, P.W. Functions of exosomes and microbial extracellular vesicles in allergy and contact and delayed-type hypersensitivity. Int. Arch. Allergy Immunol. 2016, 171, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Szczepanik, M.; Akahira-Azuma, M.; Bryniarski, K.; Tsuji, R.F.; Kawikova, I.; Ptak, W.; Kiener, C.; Campos, R.A.; Askenase, P.W. B-1 B cells mediate required early T cell recruitment to elicit protein-induced delayed-type hypersensitivity. J. Immunol. 2003, 171, 6225–6235. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, R.F.; Szczepanik, M.; Kawikova, I.; Paliwal, V.; Campos, R.A.; Itakura, A.; Akahira-Azuma, M.; Baumgarth, N.; Herzenberg, L.A.; Askenase, P.W. B cell-dependent T cell responses: IgM antibodies are required to elicit contact sensitivity. J. Exp. Med. 2002, 196, 1277–1290. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Bedel, R.; Krovi, S.H.; Tuttle, K.D.; Zhang, B.; Gross, J.; Gapin, L.; Matsuda, J.L. Mutation of the Traj18 gene segment using TALENs to generate Natural Killer T cell deficient mice. Sci. Rep. 2016, 6, 27375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaplan, D.H.; Igyártó, B.Z.; Gaspari, A.A. Early immune events in the induction of allergic contact dermatitis. Nat. Rev. Immunol. 2012, 12, 114–124. [Google Scholar] [CrossRef] [PubMed]
- Theoharides, T.C.; Kalogeromitros, D. The critical role of mast cells in allergy and inflammation. Ann. N. Y. Acad. Sci. 2006, 1088, 78–99. [Google Scholar] [CrossRef] [PubMed]
- Esparvarinha, M.; Nickho, H.; Mohammadi, H.; Aghebati-Maleki, L.; Abdolalizadeh, J.; Majidi, J. The role of free kappa and lambda light chains in the pathogenesis and treatment of inflammatory diseases. Biomed. Pharmacother. 2017, 91, 632–644. [Google Scholar] [CrossRef] [PubMed]
- Mortaz, E.; Adcock, I.M.; Jamaati, H.; Khosravi, A.; Movassaghi, M.; Garssen, J.; Alavi Mogadam, M.; Redegeld, F.A. Immunoglobulin free light chains in the pathogenesis of lung disorders. Iran. J. Allergy Asthma Immunol. 2017, 16, 282–288. [Google Scholar] [PubMed]
- Ramirez-Alvarado, M.; Barnidge, D.R.; Murray, D.L.; Dispenzieri, A.; Marin-Argany, M.; Dick, C.J.; Cooper, S.A.; Nasr, S.H.; Ward, C.J.; Dasari, S.; et al. Assessment of renal response with urinary exosomes in patients with AL amyloidosis: A proof of concept. Am. J. Hematol. 2017, 92, 536–541. [Google Scholar] [CrossRef] [PubMed]
- Di Noto, G.; Chiarini, M.; Paolini, L.; Mazzoldi, E.L.; Giustini, V.; Radeghieri, A.; Caimi, L.; Ricotta, D. Immunoglobulin free light chains and gags mediate multiple myeloma extracellular vesicles uptake and secondary NFκB nuclear translocation. Front. Immunol. 2014, 5, 517. [Google Scholar] [CrossRef] [PubMed]
- Nazimek, K.; Bryniarski, K.; Santocki, M.; Ptak, W. Exosomes as mediators of intercellular communication: Clinical implications. Pol. Arch. Med. Wewn. 2015, 125, 370–380. [Google Scholar] [CrossRef] [PubMed]
- Hutchinson, A.T.; Jones, D.R.; Raison, R.L. The ability to interact with cell membranes suggests possible biological roles for free light chain. Immunol. Lett. 2012, 142, 75–77. [Google Scholar] [CrossRef] [PubMed]
- Hutchinson, A.T.; Ramsland, P.A.; Jones, D.R.; Agostino, M.; Lund, M.E.; Jennings, C.V.; Bockhorni, V.; Yuriev, E.; Edmundson, A.B.; Raison, R.L. Free Ig light chains interact with sphingomyelin and are found on the surface of myeloma plasma cells in an aggregated form. J. Immunol. 2010, 185, 4179–4188. [Google Scholar] [CrossRef] [PubMed]
- Barile, L.; Vassalli, G. Exosomes: Therapy delivery tools and biomarkers of diseases. Pharmacol. Ther. 2017, 174, 63–78. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Vader, P.; Schiffelers, R.M. Extracellular vesicles for nucleic acid delivery: Progress and prospects for safe RNA-based gene therapy. Gene Ther. 2017, 24, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.C.; Gao, J.Q. Exosomes as novel bio-carriers for gene and drug delivery. Int. J. Pharm. 2017, 521, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Stremersch, S.; De Smedt, S.C.; Raemdonck, K. Therapeutic and diagnostic applications of extracellular vesicles. J. Control. Release 2016, 244, 167–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Sun, X.; Zhao, J.; Yang, Y.; Cai, X.; Xu, J.; Cao, P. Exosomes: A novel strategy for treatment and prevention of diseases. Front. Pharmacol. 2017, 8, 300. [Google Scholar] [CrossRef] [PubMed]
- Paszko, E.; Senge, M.O. Immunoliposomes. Curr. Med. Chem. 2012, 19, 5239–5277. [Google Scholar] [CrossRef] [PubMed]
- Ohno, S.; Drummen, G.P.; Kuroda, M. Focus on extracellular vesicles: Development of extracellular vesicle-based therapeutic systems. Int. J. Mol. Sci. 2016, 17, 172. [Google Scholar] [CrossRef] [PubMed]
- Sagini, K.; Costanzi, E.; Emiliani, C.; Buratta, S.; Urbanelli, L. Extracellular vesicles as conveyors of membrane-derived bioactive lipids in immune system. Int. J. Mol. Sci. 2018, 19, 1227. [Google Scholar] [CrossRef] [PubMed]
- Wen, C.; Seeger, R.C.; Fabbri, M.; Wang, L.; Wayne, A.S.; Jong, A.Y. Biological roles and potential applications of immune cell-derived extracellular vesicles. J. Extracell. Vesicles 2017, 6, 1400370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prada, I.; Meldolesi, J. Binding and fusion of extracellular vesicles to the plasma membrane of their cell targets. Int. J. Mol. Sci. 2016, 17, 1296. [Google Scholar] [CrossRef] [PubMed]
- Zemelka-Wiącek, M.; Majewska-Szczepanik, M.; Pyrczak, W.; Szczepanik, M. Complementary methods for contact hypersensitivity (CHS) evaluation in mice. J. Immunol. Methods 2013, 387, 270–275. [Google Scholar] [CrossRef] [PubMed]
- Redegeld, F.A.; van der Heijden, M.W.; Kool, M.; Heijdra, B.M.; Garssen, J.; Kraneveld, A.D.; Van Loveren, H.; Roholl, P.; Saito, T.; Verbeek, J.S.; et al. Immunoglobulin-free light chains elicit immediate hypersensitivity-like responses. Nat. Med. 2002, 8, 694–701. [Google Scholar] [CrossRef] [PubMed]








© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nazimek, K.; Askenase, P.W.; Bryniarski, K. Antibody Light Chains Dictate the Specificity of Contact Hypersensitivity Effector Cell Suppression Mediated by Exosomes. Int. J. Mol. Sci. 2018, 19, 2656. https://doi.org/10.3390/ijms19092656
Nazimek K, Askenase PW, Bryniarski K. Antibody Light Chains Dictate the Specificity of Contact Hypersensitivity Effector Cell Suppression Mediated by Exosomes. International Journal of Molecular Sciences. 2018; 19(9):2656. https://doi.org/10.3390/ijms19092656
Chicago/Turabian StyleNazimek, Katarzyna, Philip W. Askenase, and Krzysztof Bryniarski. 2018. "Antibody Light Chains Dictate the Specificity of Contact Hypersensitivity Effector Cell Suppression Mediated by Exosomes" International Journal of Molecular Sciences 19, no. 9: 2656. https://doi.org/10.3390/ijms19092656