Combined Proteomic and Metabolomic Profiling of the Arabidopsis thaliana vps29 Mutant Reveals Pleiotropic Functions of the Retromer in Seed Development

 ,
, 
Abstract
:
1. Introduction
2. Results
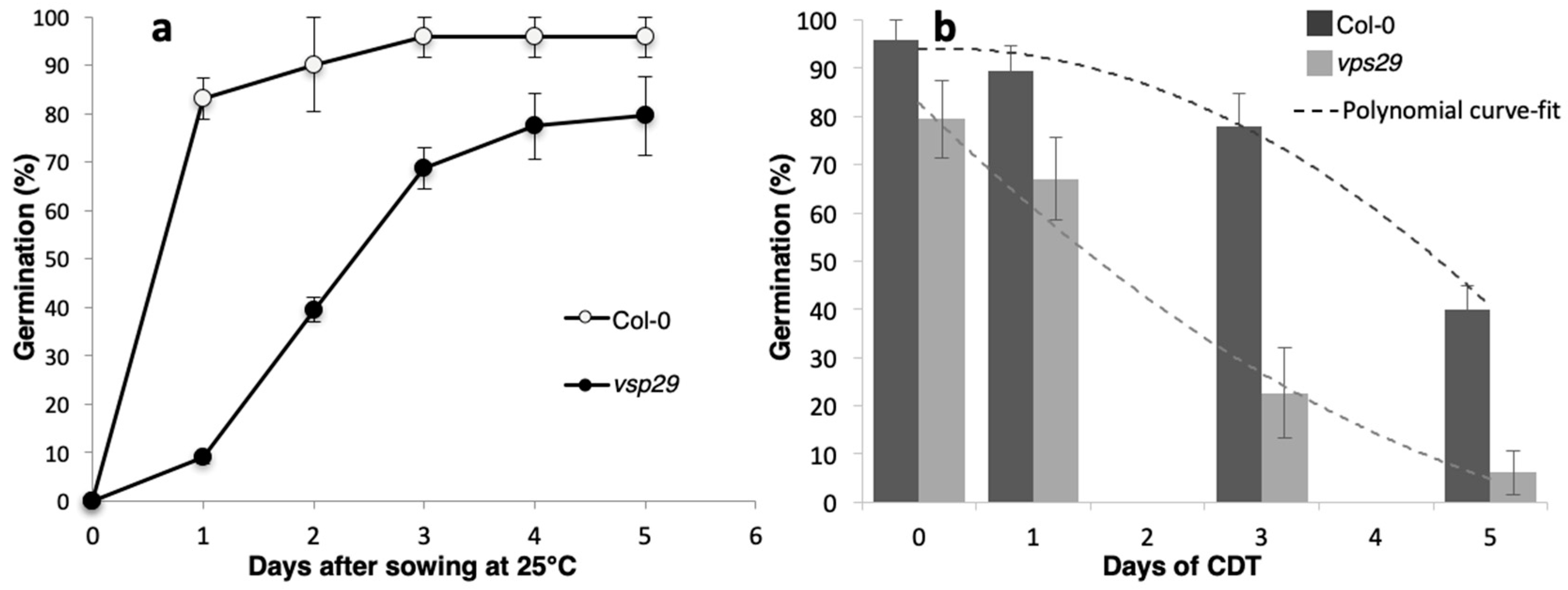
2.1. Impact of the vps29 Mutation on Seed Physiology
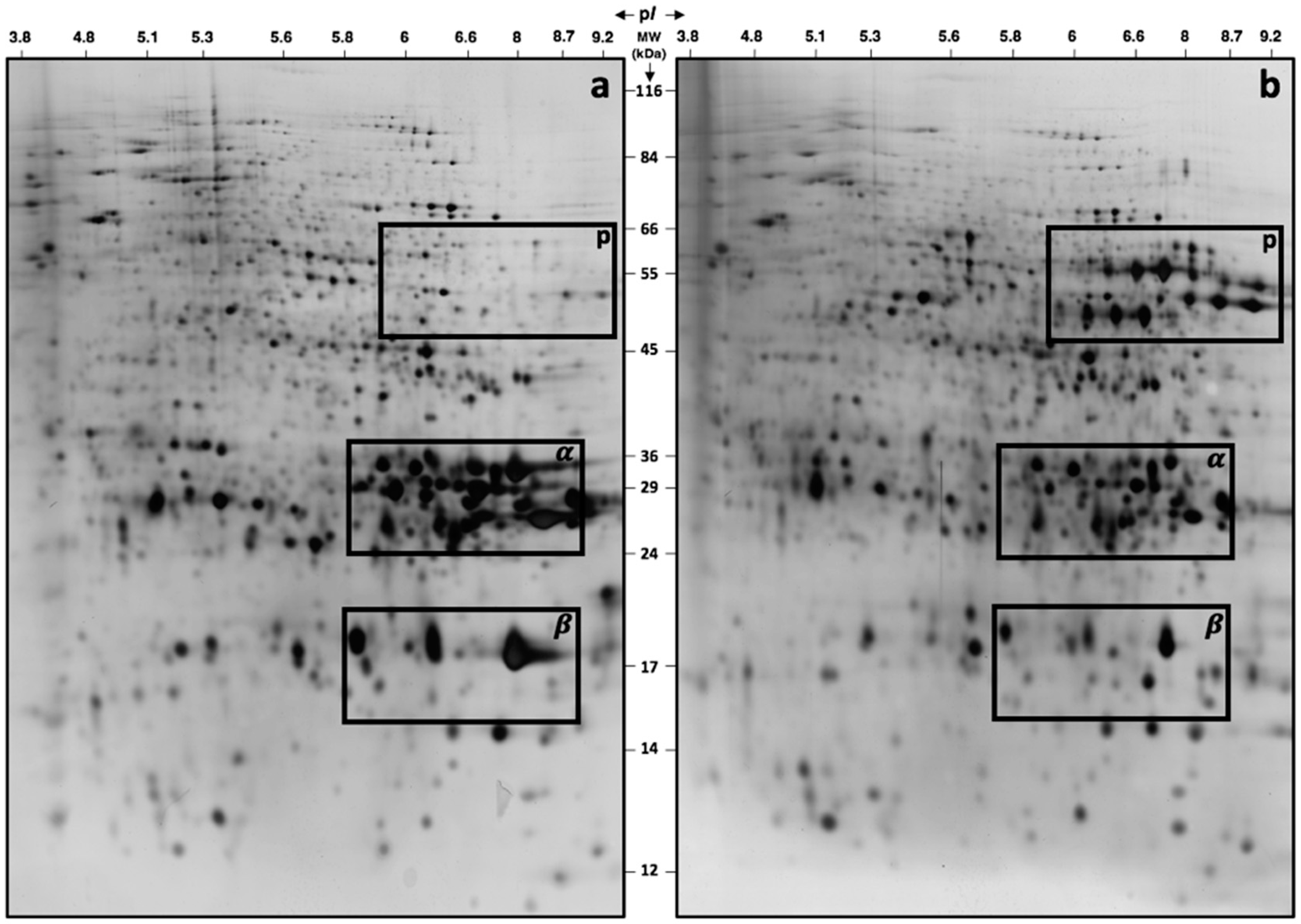
2.2. Gel-Based Proteomics between Col-0 and vps29 Dry Mature Seeds
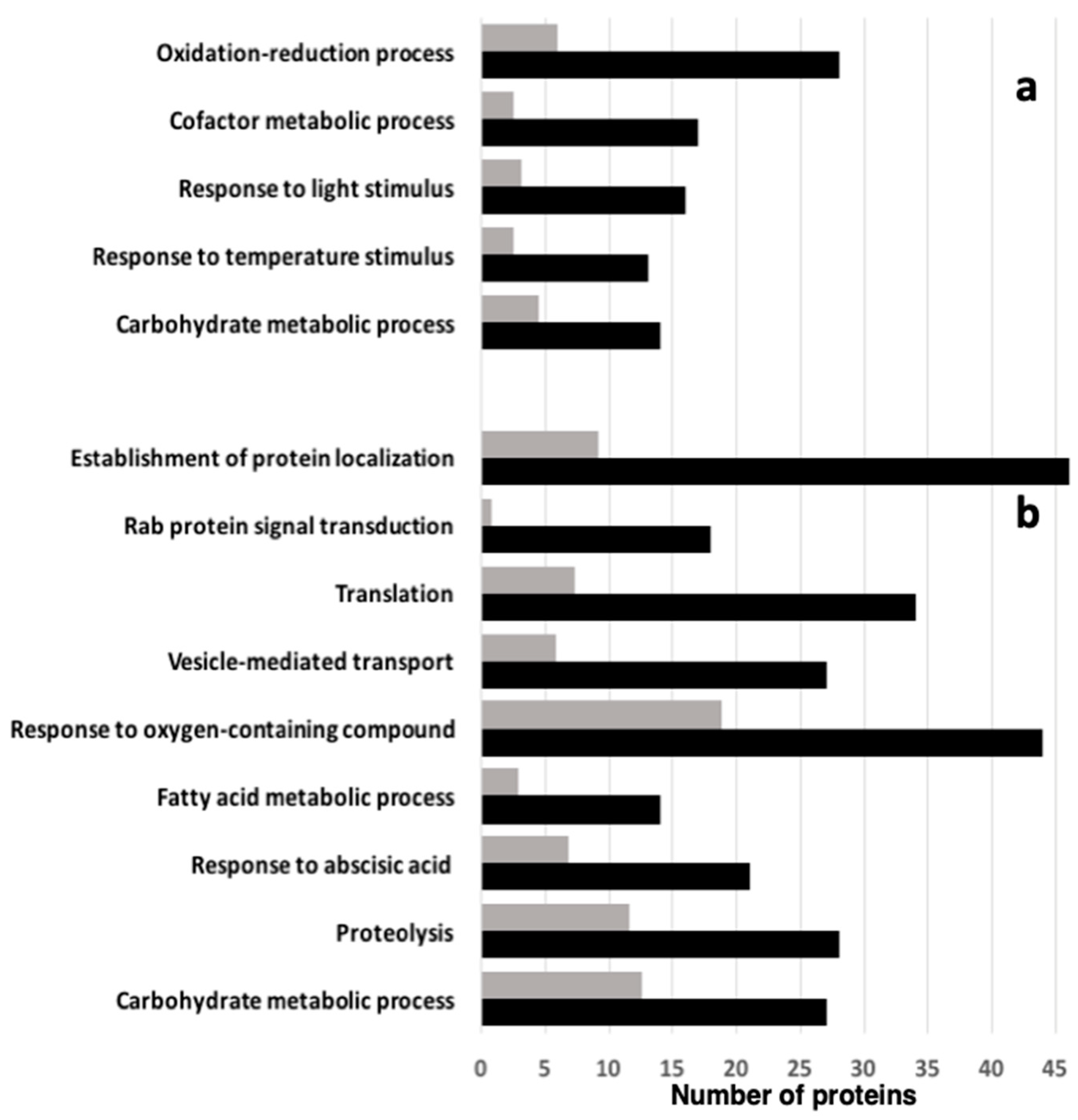
2.3. Quantitative Shotgun Proteomics between Col-0 and vps29 Dry Mature Seeds
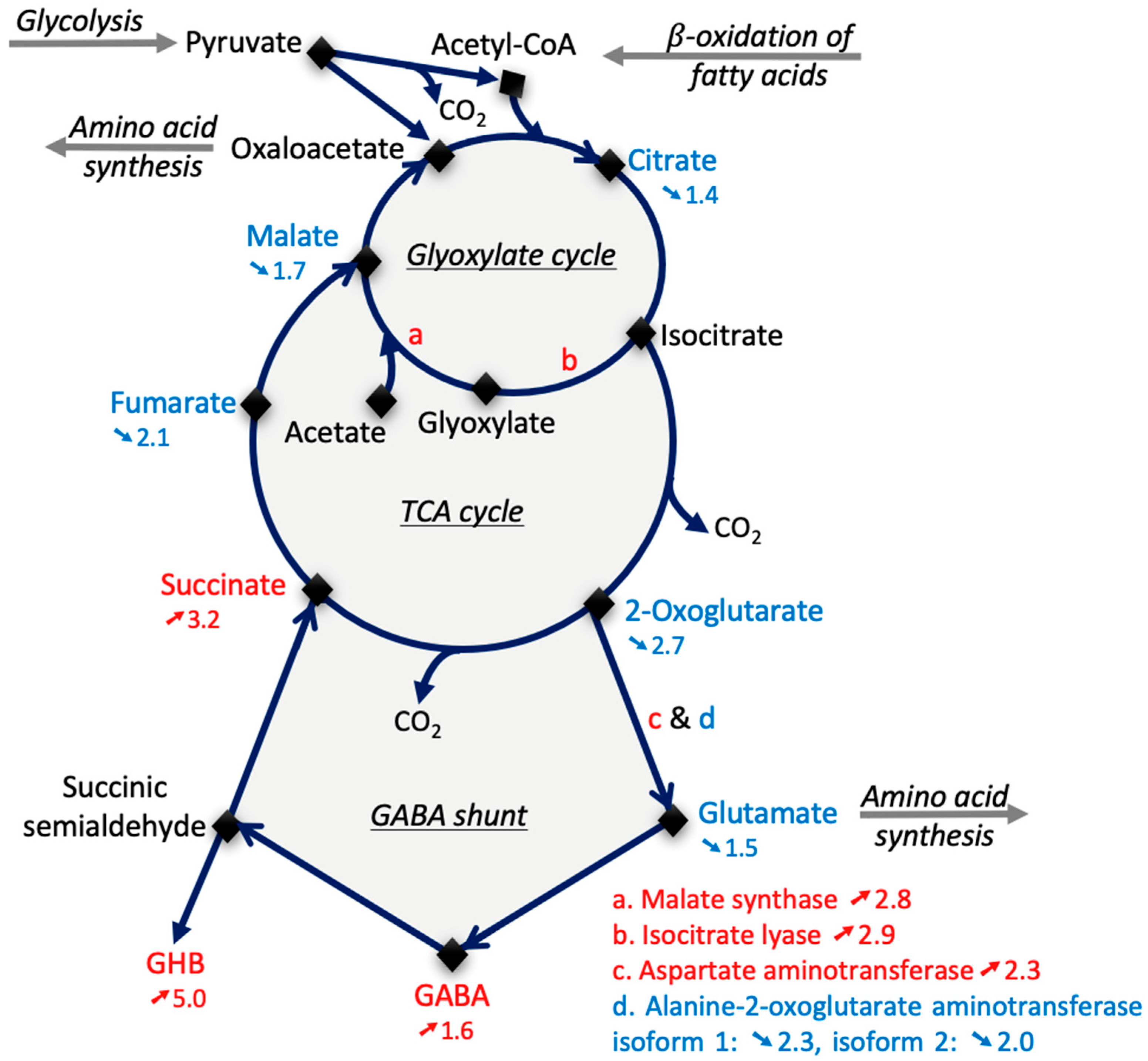
2.4. Quantitative GC/MS-Based Metabolic Profiling of Arabidopsis Col-0 and vps29 Dry Mature Seeds
3. Discussion
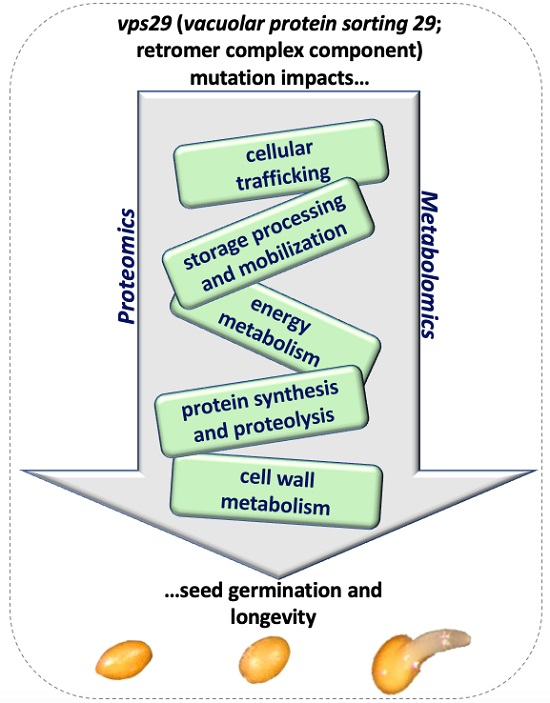
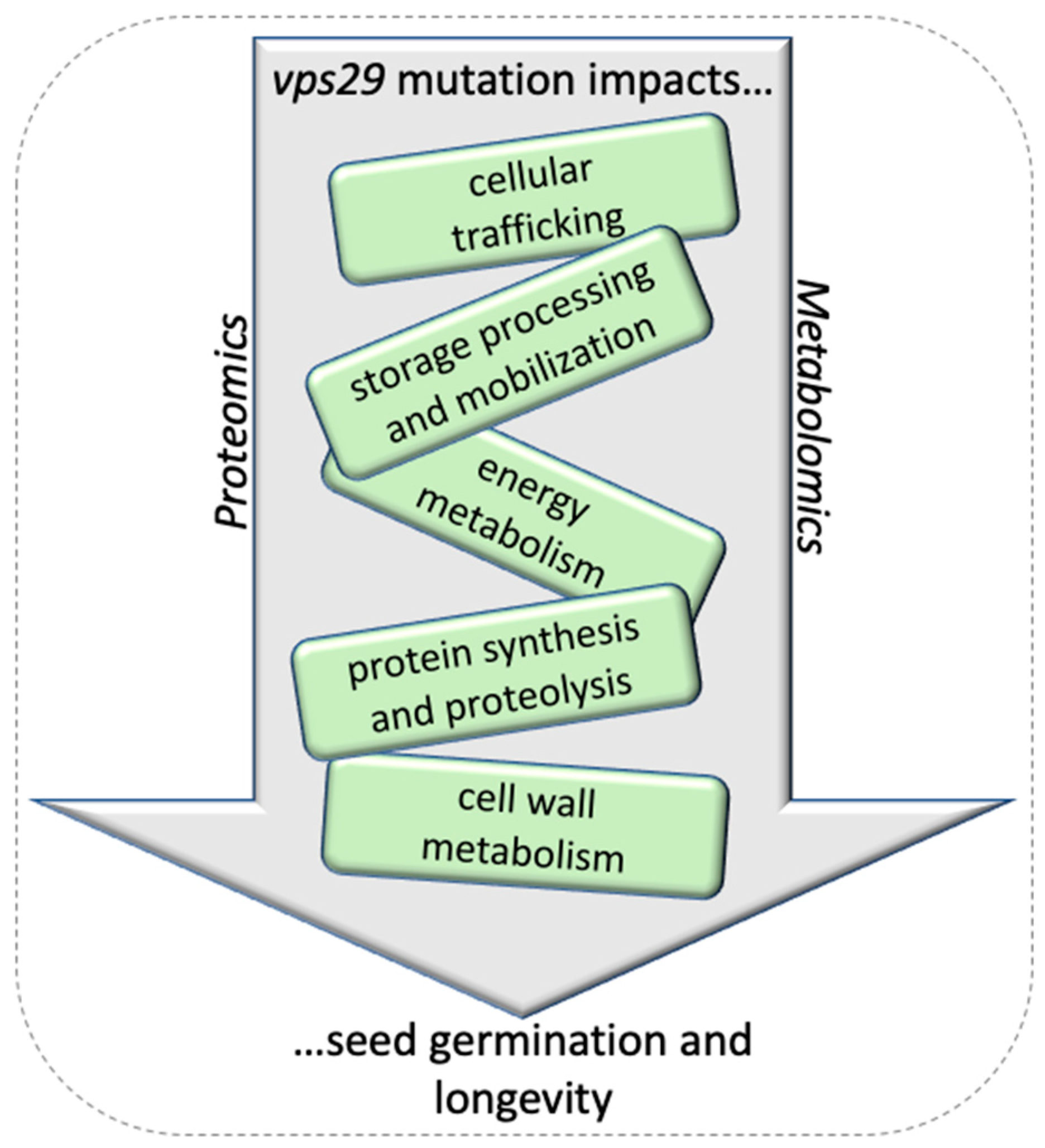
3.1. Seed Germination and Longevity: An Integrative View Based on vps29 Proteomics and Metabolomics
3.2. The vps29 Mutation Affects Energy Metabolism in Dry Mature Seeds
3.3. Cellular Trafficking in vps29 Seeds
3.4. Storage Processing and Mobilization
3.5. Changes in Cell Wall Metabolism
4. Materials and Methods
4.1. Plant Material and Culture Conditions
4.2. Germination Assays
4.3. Total Soluble Protein Extraction
4.4. 2D Gel-Based Proteomics and Image Processing
4.5. Shotgun Proteomic Analysis
4.6. Protein Identification and Quantification
4.7. Metabolome Analysis by Gas Chromatography Coupled to Mass Spectrometry (GC–MS)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| MVB | multivesicular body |
| PVC | prevacuolar compartment |
| PSV | protein storage vacuole |
| SSP | seed storage protein |
| TGN | trans-Golgi network |
| VSR | vacuolar sorting receptor |
References
- Baud, S.; Dubreucq, B.; Miquel, M.; Rochat, C.; Lepiniec, L. Storage reserve accumulation in Arabidopsis: Metabolic and developmental control of seed filling. Arabidopsis Book 2008, 6, e0113. [Google Scholar] [CrossRef] [PubMed]
- Baud, S.; Boutin, J.-P.; Miquel, M.; Lepiniec, L.; Rochat, C. An integrated overview of seed development in Arabidopsis thaliana ecotype WS. Plant Physiol. Biochem. 2002, 40, 151–160. [Google Scholar] [CrossRef]
- Baud, S.; Kelemen, Z.; Thévenin, J.; Boulard, C.; Blanchet, S.; To, A.; Payre, M.; Berger, N.; Effroy-Cuzzi, D.; Franco-Zorrilla, J.M.; et al. Deciphering the molecular mechanisms underpinning the transcriptional control of gene expression by L-AFL proteins in Arabidopsis seed. Plant Physiol. 2016, 171, 1099–1112. [Google Scholar] [CrossRef] [PubMed]
- Fatihi, A.; Boulard, C.; Bouyer, D.; Baud, S.; Dubreucq, B.; Lepiniec, L. Deciphering and modifying LAFL transcriptional regulatory network in seed for improving yield and quality of storage compounds. Plant Sci. 2016, 250, 198–204. [Google Scholar] [CrossRef]
- North, H.; Baud, S.; Debeaujon, I.; Dubos, C.; Dubreucq, B.; Grappin, P.; Jullien, M.; Lepiniec, L.; Marion-Poll, A.; Miquel, M.; et al. Arabidopsis seed secrets unravelled after a decade of genetic and omics-driven research. Plant J. 2010, 61, 971–981. [Google Scholar] [CrossRef] [Green Version]
- Robinson, D.G.; Hinz, G. Golgi-mediated transport of seed storage proteins. Seed Sci. Res. 1999, 9, 267–283. [Google Scholar] [CrossRef]
- Otegui, M.S.; Spitzer, C. Endosomal functions in plants. Traffic 2008, 9, 1589–1598. [Google Scholar] [CrossRef]
- Shimada, T.; Fuji, K.; Tamura, K.; Kondo, M.; Nishimura, M.; Hara-Nishimura, I. Vacuolar sorting receptor for seed storage proteins in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2003, 100, 16095–16100. [Google Scholar] [CrossRef]
- Shimada, T.; Koumoto, Y.; Li, L.; Yamazaki, M.; Kondo, M.; Nishimura, M.; Hara-Nishimura, I. AtVPS29, a putative component of a retromer complex, is required for the efficient sorting of seed storage proteins. Plant Cell Physiol. 2006, 47, 1187–1194. [Google Scholar] [CrossRef] [PubMed]
- Attar, N.; Cullen, P.J. The retromer complex. Adv. Enzyme Regul. 2010, 50, 216–236. [Google Scholar] [CrossRef] [PubMed]
- McNally, K.E.; Cullen, P.J. Endosomal retrieval of cargo: Retromer is not alone. Trends Cell Biol. 2018, 28, 807–822. [Google Scholar] [CrossRef] [PubMed]
- Seaman, M.N.J.; Marcusson, E.G.; Cereghino, J.L.; Emr, S.D. Endosome to Golgi retrieval of the vacuolar protein sorting receptor, Vps10p, requires the function of the VPS29, VPS30 and VPS35 gene products. J. Cell Biol. 1997, 137, 79–92. [Google Scholar] [CrossRef]
- Seaman, M.N.J. The retromer complex—Endosomal protein recycling and beyond. J. Cell Sci. 2012, 125, 4693–4702. [Google Scholar] [CrossRef] [PubMed]
- Burd, C.; Cullen, P.J. Retromer: A master conductor of endosome sorting. Cold Spring Harb. Perspect. Biol. 2014, 6, a016774. [Google Scholar] [CrossRef]
- Jaillais, Y.; Santambrogio, M.; Rozier, F.; Fobis-Loisy, I.; Miège, C.; Gaude, T. The retromer protein VPS29 links cell polarity and organ initiation in plants. Cell 2007, 130, 1057–1070. [Google Scholar] [CrossRef]
- Pourcher, M.; Santambrogio, M.; Thazar, N.; Thierry, A.-M.; Fobis-Loisy, I.; Miège, C.; Jaillais, Y.; Gaude, T. Analyses of SORTING NEXINs reveal distinct retromer-subcomplex functions in development and protein sorting in Arabidopsis thaliana. Plant Cell 2010, 22, 3980–3991. [Google Scholar] [CrossRef] [PubMed]
- Thazar-Poulot, N.; Miquel, M.; Fobis-Loisy, I.; Gaude, T. Peroxisome extensions deliver the Arabidopsis SDP1 lipase to oil bodies. Proc. Natl. Acad. Sci. USA 2015, 112, 4158–4163. [Google Scholar] [CrossRef]
- Zelazny, E.; Santambrogio, M.; Pourcher, M.; Chambrier, P.; Berne-Dedieu, A.; Fobis-Loisy, I.; Miège, C.; Jaillais, Y.; Gaude, T. Mechanisms governing the endosomal membrane recruitment of the core retromer in Arabidopsis. Biol. Chem. 2013, 288, 8815–8825. [Google Scholar] [CrossRef]
- Rajjou, L.; Duval, M.; Gallardo, K.; Catusse, J.; Bally, J.; Job, C.; Job, D. Seed germination and vigor. Annu. Rev. Plant Biol. 2012, 63, 507–533. [Google Scholar] [CrossRef]
- Finch-Savage, W.E.; Bassel, G.W. Seed vigour and crop establishment: extending performance beyond adaptation. J. Exp. Bot. 2016, 67, 567–591. [Google Scholar] [CrossRef]
- Rajjou, L.; Belghazi, M.; Catusse, J.; Ogé, L.; Arc, E.; Godin, B.; Chibani, K.; Ali-Rachidi, S.; Collet, B.; Grappin, P.; et al. Proteomics and Posttranslational Proteomics of Seed Dormancy and Germination. In Seed Dormancy; Kermode, A.R., Ed.; Humana Press: Totowa, NJ, USA, 2011; Volume 773, pp. 215–236. ISBN 978-1-61779-230-4. [Google Scholar]
- Galland, M.; Job, D.; Rajjou, L. The seed proteome web portal. Front Plant Sci. 2012, 3. [Google Scholar] [CrossRef] [PubMed]
- Galland, M.; Huguet, R.; Arc, E.; Cueff, G.; Job, D.; Rajjou, L. Dynamic proteomics emphasizes the importance of selective mRNA translation and protein turnover during Arabidopsis seed germination. Mol. Cell Proteom. 2014, 13, 252–268. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.Q.; Liu, S.J.; Song, S.Q.; Møller, I.M. Proteomics of seed development, desiccation tolerance, germination and vigor. Plant Physiol. Biochem. 2015, 86, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Durand, T.C.; Sergeant, K.; Renaut, J.; Planchon, S.; Hoffmann, L.; Carpin, S.; Label, P.; Morabito, D.; Hausman, J.F. Poplar under drought: Comparison of leaf and cambial proteomic responses. J. Proteom. 2011, 74, 1396–1410. [Google Scholar] [CrossRef]
- Rabilloud, T. How to use 2D gel electrophoresis in plant proteomics. In Plant Proteomics; Jorrin-Novo, J.V., Komatsu, S., Weckwerth, W., Wienkoop, S., Eds.; Humana Press: Totowa, NJ, USA, 2014; Volume 1072, pp. 43–50. ISBN 978-1-62703-630-6. [Google Scholar]
- Arc, E.; Galland, M.; Cueff, G.; Godin, B.; Lounifi, I.; Job, D.; Rajjou, L. Reboot the system thanks to protein post-translational modifications and proteome diversity: How quiescent seeds restart their metabolism to prepare seedling establishment. Proteomics 2011, 11, 1606–1618. [Google Scholar] [CrossRef] [PubMed]
- Laval, V.; Masclaux, F.; Serin, A.; Carrière, M.; Roldan, C.; Devic, M.; Pont-Lezica, R.F.; Galaud, J.-P. Seed germination is blocked in Arabidopsis putative vacuolar sorting receptor (atbp80) antisense transformants. J. Exp. Bot. 2003, 54, 213–221. [Google Scholar] [CrossRef] [Green Version]
- Zouhar, J.; Muñoz, A.; Rojo, E. Functional specialization within the vacuolar sorting receptor family: VSR1, VSR3 and VSR4 sort vacuolar storage cargo in seeds and vegetative tissues: Sorting receptors for storage proteins. Plant J. 2010, 64, 577–588. [Google Scholar] [CrossRef]
- Sattler, S.E. Vitamin E is essential for seed longevity and for preventing lipid peroxidation during germination. Plant Cell 2004, 16, 1419–1432. [Google Scholar] [CrossRef]
- Smirnoff, N. The role of active oxygen in the response of plants to water deficit and desiccation. New Phytol. 1993, 125, 27–58. [Google Scholar] [CrossRef]
- Fait, A.; Fromm, H.; Walter, D.; Galili, G.; Fernie, A.R. Highway or byway: the metabolic role of the GABA shunt in plants. Trends Plant Sci. 2008, 13, 14–19. [Google Scholar] [CrossRef]
- Michaeli, S.; Fait, A.; Lagor, K.; Nunes-Nesi, A.; Grillich, N.; Yellin, A.; Bar, D.; Khan, M.; Fernie, A.R.; Turano, F.J.; et al. A mitochondrial GABA permease connects the GABA shunt and the TCA cycle, and is essential for normal carbon metabolism: Plant mitochondrial GABA permease. Plant J. 2011, 67, 485–498. [Google Scholar] [CrossRef]
- Song, H.; Xu, X.; Wang, H.; Wang, H.; Tao, Y. Exogenous γ-aminobutyric acid alleviates oxidative damage caused by aluminium and proton stresses on barley seedlings. J. Sci. Food Agric. 2010, 90, 1410–1416. [Google Scholar] [CrossRef] [PubMed]
- Sgaravatti, A.M.; Sgarbi, M.B.; Testa, C.G.; Durigon, K.; Pederzolli, C.D.; Prestes, C.C.; Wyse, A.T.S.; Wannmacher, C.M.D.; Wajner, M.; Dutra-Filho, C.S. Gamma-hydroxybutyric acid induces oxidative stress in cerebral cortex of young rats. Neurochem. Int. 2007, 50, 564–570. [Google Scholar] [CrossRef]
- Ben-Izhak Monselise, E.; Parola, A.H.; Kost, D. Low-frequency electromagnetic fields induce a stress effect upon higher plants, as evident by the universal stress signal, alanine. Biochem. Biophys. Res. Commun. 2003, 302, 427–434. [Google Scholar] [CrossRef]
- Uemura, T.; Ueda, T. Plant vacuolar trafficking driven by RAB and SNARE proteins. Curr. Opin. Plant Biol. 2014, 22, 116–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira-Leal, J.B.; Seabra, M.C. Evolution of the rab family of small GTP-binding proteins. J. Mol. Biol. 2001, 313, 889–901. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Zhao, Q.; Gao, C.; Ding, Y.; Zeng, Y.; Ueda, T.; Nakano, A.; Jiang, L. Activation of the Rab7 GTPase by the MON1-CCZ1 complex is essential for PVC-to-vacuole trafficking and plant growth in Arabidopsis. Plant Cell 2014, 26, 2080–2097. [Google Scholar] [CrossRef]
- Ebine, K.; Inoue, T.; Ito, J.; Ito, E.; Uemura, T.; Goh, T.; Abe, H.; Sato, K.; Nakano, A.; Ueda, T. Plant vacuolar trafficking occurs through distinctly regulated pathways. Curr. Biol. 2014, 24, 1375–1382. [Google Scholar] [CrossRef] [PubMed]
- Mazel, A. Induction of Salt and Osmotic Stress Tolerance by Overexpression of an Intracellular Vesicle Trafficking Protein AtRab7 (AtRabG3e). Plant Physiol. 2004, 134, 118–128. [Google Scholar] [CrossRef]
- Kang, H.; Hwang, I. Vacuolar Sorting Receptor-Mediated Trafficking of Soluble Vacuolar Proteins in Plant Cells. Plants 2014, 3, 392–408. [Google Scholar] [CrossRef]
- Yamazaki, M.; Shimada, T.; Takahashi, H.; Tamura, K.; Kondo, M.; Nishimura, M.; Hara-Nishimura, I. Arabidopsis VPS35, a Retromer Component, is Required for Vacuolar Protein Sorting and Involved in Plant Growth and Leaf Senescence. Plant Cell Physiol. 2008, 49, 142–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, H.; Kim, S.Y.; Song, K.; Sohn, E.J.; Lee, Y.; Lee, D.W.; Hara-Nishimura, I.; Hwang, I. Trafficking of vacuolar proteins: The crucial role of Arabidopsis vacuolar protein sorting 29 in recycling vacuolar sorting receptor. Plant Cell 2012, 24, 5058–5073. [Google Scholar] [CrossRef] [PubMed]
- Fuji, K.; Shirakawa, M.; Shimono, Y.; Kunieda, T.; Fukao, Y.; Koumoto, Y.; Takahashi, H.; Hara-Nishimura, I.; Shimada, T. The adaptor complex AP-4 regulates vacuolar protein sorting at the trans-Golgi network by interacting with VACUOLAR SORTING RECEPTOR1. Plant Physiol. 2016, 170, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Shimada, T.; Yamada, K.; Kataoka, M.; Nakaune, S.; Koumoto, Y.; Kuroyanagi, M.; Tabata, S.; Kato, T.; Shinozaki, K.; Seki, M.; et al. Vacuolar processing enzymes are essential for proper processing of seed storage proteins in Arabidopsis thaliana. J. Biol. Chem. 2003, 278, 32292–32299. [Google Scholar] [CrossRef] [PubMed]
- Chae, K.; Gonong, B.J.; Kim, S.-C.; Kieslich, C.A.; Morikis, D.; Balasubramanian, S.; Lord, E.M. A multifaceted study of stigma/style cysteine-rich adhesin (SCA)-like Arabidopsis lipid transfer proteins (LTPs) suggests diversified roles for these LTPs in plant growth and reproduction. J. Exp. Bot. 2010, 61, 4277–4290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Svenning, S.; Lamark, T.; Krause, K.; Johansen, T. Plant NBR1 is a selective autophagy substrate and a functional hybrid of the mammalian autophagic adapters NBR1 and p62/SQSTM1. Autophagy 2011, 7, 993–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, B.; Sinkevicius, K.W.; Selinger, D.A.; Tarczynski, M.C. The homeobox gene GLABRA2 affects seed oil content in Arabidopsis. Plant Mol. Biol. 2006, 60, 377–387. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Katavic, V.; Yu, Y.; Kunst, L.; Haughn, G. Arabidopsis glabra2 mutant seeds deficient in mucilage biosynthesis produce more oil: Mucilage-deficient gl2 seeds produce more oil. Plant J. 2012, 69, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Du, X.; Zhu, Y.; Wang, Z.; Hua, S.; Li, Z.; Guo, W.; Zhang, G.; Peng, J.; Jiang, L. Seed Fatty Acid Reducer acts downstream of gibberellin signalling pathway to lower seed fatty acid storage in Arabidopsis: GA regulates seed fatty acid storage via SFARs. Plant Cell Environ. 2012, 35, 2155–2169. [Google Scholar] [CrossRef]
- Chepyshko, H.; Lai, C.-P.; Huang, L.-M.; Liu, J.-H.; Shaw, J.-F. Multifunctionality and diversity of GDSL esterase/lipase gene family in rice (Oryza sativa L. japonica) genome: new insights from bioinformatics analysis. BMC Genom. 2012, 13, 309. [Google Scholar] [CrossRef]
- Durand, T.C.; Sergeant, K.; Carpin, S.; Label, P.; Morabito, D.; Hausman, J.-F.; Renaut, J. Screening for changes in leaf and cambial proteome of Populus tremula×P. alba under different heat constraints. J. Plant Physiol. 2012, 169, 1698–1718. [Google Scholar] [CrossRef]
- Eastmond, P.J.; Quettier, A.-L.; Kroon, J.T.M.; Craddock, C.; Adams, N.; Slabas, A.R. PHOSPHATIDIC ACID PHOSPHOHYDROLASE1 and 2 eegulate phospholipid synthesis at the endoplasmic reticulum in Arabidopsis. Plant Cell 2010, 22, 2796–2811. [Google Scholar] [CrossRef] [PubMed]
- Sampedro, J.; Gianzo, C.; Iglesias, N.; Guitian, E.; Revilla, G.; Zarra, I. AtBGAL10 is the main xyloglucan-Galactosidase in Arabidopsis, and its absence results in unusual xyloglucan subunits and growth defects. Plant Physiol. 2012, 158, 1146–1157. [Google Scholar] [CrossRef]
- Strohmeier, M.; Hrmova, M.; Fischer, M.; Harvey, A.J.; Fincher, G.B.; Pleiss, J. Molecular modeling of family GH16 glycoside hydrolases: Potential roles for xyloglucan transglucosylases/hydrolases in cell wall modification in the poaceae. Protein Sci. 2009, 13, 3200–3213. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.B.; Cosgrove, D.J. Changes in cell wall biomechanical properties in the xyloglucan-deficient xxt1/xxt2 mutant of Arabidopsis. Plant Physiol. 2012, 158, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-B.; Lu, S.-M.; Zhang, J.-F.; Liu, S.; Lu, Y.-T. A xyloglucan endotransglucosylase/hydrolase involves in growth of primary root and alters the deposition of cellulose in Arabidopsis. Planta 2007, 226, 1547–1560. [Google Scholar] [CrossRef]
- Vázquez-Lobo, A.; Roujol, D.; Zuñiga-Sánchez, E.; Albenne, C.; Piñero, D.; de Buen, A.G.; Jamet, E. The highly conserved spermatophyte cell wall DUF642 protein family: Phylogeny and first evidence of interaction with cell wall polysaccharides in vitro. Mol. Phylogenet. Evol. 2012, 63, 510–520. [Google Scholar]
- Zúñiga-Sánchez, E.; Gamboa-de Buen, A. The Two DUF642 At5g11420 and At4g32460-Encoded Proteins Interact in Vitro with the AtPME3 Catalytic Domain; InTech: Rijeka, Croatia, 2012; ISBN 978-953-51-0244-1. [Google Scholar]
- Garza-Caligaris, L.E.; Avendaño-Vázquez, A.O.; Alvarado-López, S.; Zúñiga-Sánchez, E.; Orozco-Segovia, A.; Pérez-Ruíz, R.V.; Gamboa-deBuen, A. At3g08030 transcript: A molecular marker of seed ageing. Ann. Bot. 2012, 110, 1253–1260. [Google Scholar] [CrossRef]
- Hayashi, S.; Ishii, T.; Matsunaga, T.; Tominaga, R.; Kuromori, T.; Wada, T.; Shinozaki, K.; Hirayama, T. The glycerophosphoryl diester phosphodiesterase-like proteins SHV3 and its homologs play important roles in cell wall organization. Plant Cell Physiol. 2008, 49, 1522–1535. [Google Scholar] [CrossRef] [PubMed]
- Hong, M.J.; Kim, D.Y.; Lee, T.G.; Jeon, W.B.; Seo, Y.W. Functional characterization of pectin methylesterase inhibitor (PMEI) in wheat. Genes Genet. Syst. 2010, 85, 97–106. [Google Scholar] [CrossRef] [Green Version]
- Atmodjo, M.A.; Sakuragi, Y.; Zhu, X.; Burrell, A.J.; Mohanty, S.S.; Atwood, J.A.; Orlando, R.; Scheller, H.V.; Mohnen, D. Galacturonosyltransferase (GAUT)1 and GAUT7 are the core of a plant cell wall pectin biosynthetic homogalacturonan:galacturonosyltransferase complex. Proc. Natl. Acad. Sci. USA 2011, 108, 20225–20230. [Google Scholar] [CrossRef]
- Rautengarten, C.; Usadel, B.; Neumetzler, L.; Hartmann, J.; Büssis, D.; Altmann, T. A subtilisin-like serine protease essential for mucilage release from Arabidopsis seed coats. Plant J. 2008, 54, 466–480. [Google Scholar] [CrossRef]
- Rajjou, L.; Lovigny, Y.; Groot, S.P.C.; Belghazi, M.; Job, C.; Job, D. Proteome-Wide Characterization of Seed Aging in Arabidopsis: A Comparison between Artificial and Natural Aging Protocols. Plant Physiol. 2008, 148, 620–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradford, M. A rapid and sensitive method for the quantitation of microgram quantities of protein using the principle of protein dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Arc, E.; Chibani, K.; Grappin, P.; Jullien, M.; Godin, B.; Cueff, G.; Valot, B.; Balliau, T.; Job, D.; Rajjou, L. Cold Stratification and Exogenous Nitrates Entail Similar Functional Proteome Adjustments during Arabidopsis Seed Dormancy Release. J. Proteome Res. 2012, 11, 5418–5432. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680. [Google Scholar] [CrossRef] [PubMed]
- Ishihama, Y.; Oda, Y.; Tabata, T.; Sato, T.; Nagasu, T.; Rappsilber, J.; Mann, M. Exponentially Modified Protein Abundance Index (emPAI) for Estimation of Absolute Protein Amount in Proteomics by the Number of Sequenced Peptides per Protein. Mol. Cell Proteom. 2005, 4, 1265–1272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rappsilber, J. Large-Scale Proteomic Analysis of the Human Spliceosome. Genome Res. 2002, 12, 1231–1245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Job, C. Patterns of protein oxidation in Arabidopsis seeds and during germination. Plant Physiol. 2005, 138, 790–802. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.-P.; Cueff, G.; Hegedus, D.D.; Rajjou, L.; Bentsink, L. A role for seed storage proteins in Arabidopsis seed longevity. J. Exp. Bot. 2015, 66, 6399–6413. [Google Scholar] [CrossRef]
- Xiao, C.; Zhang, T.; Zheng, Y.; Cosgrove, D.J.; Anderson, C.T. Xyloglucan deficiency disrupts microtubule stability and cellulose biosynthesis in Arabidopsis, altering cell growth and morphogenesis. Plant Physiol. 2016, 170, 234–249. [Google Scholar] [CrossRef] [PubMed]
- Wolf, S. Plant cell wall signalling and receptor-like kinases. Biochem. J. 2017, 474, 471–492. [Google Scholar] [CrossRef] [PubMed]
- Jeevan Kumar, S.P.; Rajendra Prasad, S.; Banerjee, R.; Thammineni, C. Seed birth to death: dual functions of reactive oxygen species in seed physiology. Ann. Bot. 2015, 116, 663–668. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolites | Abundance in Col-0 Seeds (Mean) | Abundance in vps29 Seeds (Mean) | Fold Change (Col-o/vps29) | p-Value | FDR-Adjusted p-Value |
|---|---|---|---|---|---|
| β-Alanine | 4.84 | 5.35 × 10−1 | 9.1 | 0.002 | 0.007 |
| Hexacosanoate | 2.14 × 10−4 | 3.19 × 10−5 | 6.7 | 0.009 | 0.022 |
| β-Aminoisobutyrate | 6.39 × 10−2 | 1.14 × 10−2 | 5.6 | 0.002 | 0.007 |
| Dibenzoyltartrateanhydre | 3.06 × 10−1 | 8.48 × 10−2 | 3.6 | 0.003 | 0.007 |
| Monosearin | 6.63 × 10−4 | 1.89 × 10−4 | 3.5 | 0.003 | 0.007 |
| Glutamine | 1.00 | 3.25 × 10−1 | 3.1 | 0.002 | 0.007 |
| Maleate | 1.12 × 10−3 | 3.73 × 10−4 | 3.0 | 0.002 | 0.007 |
| Rhamnose | 2.10 × 10−1 | 7.75 × 10−2 | 2.7 | 0.002 | 0.007 |
| 2-Oxoglutarate | 3.37 × 10−2 | 1.24 × 10−2 | 2.7 | 0.002 | 0.007 |
| Alpha-tocopherol | 1.13 × 10−2 | 4.29 × 10−3 | 2.6 | 0.002 | 0.007 |
| Malonate | 1.38 × 10−1 | 5.41 × 10−2 | 2.6 | 0.040 | 0.079 |
| Phenylalanine | 3.29 × 10−1 | 1.31 × 10−1 | 2.5 | 0.002 | 0.007 |
| Cystine | 1.65 × 10−3 | 7.07 × 10−4 | 2.3 | 0.002 | 0.007 |
| β-Indole-3-acetonitrile | 2.90 × 10−2 | 1.31 × 10−2 | 2.2 | 0.002 | 0.007 |
| Fumarate | 1.34 | 6.40 × 10−1 | 2.1 | 0.002 | 0.007 |
| Unknown sugar (RI = 2550.4; mass = 219) | 2.18 × 10−3 | 1.19 × 10−3 | 1.8 | 0.002 | 0.007 |
| Proline | 2.12 | 1.18 | 1.8 | 0.004 | 0.011 |
| Urea | 7.82 × 10−3 | 4.47 × 10−3 | 1.7 | 0.002 | 0.007 |
| Malate | 2.35 | 1.38 | 1.7 | 0.002 | 0.007 |
| Nicotinate | 7.61 × 10−3 | 4.98 × 10−3 | 1.5 | 0.002 | 0.007 |
| Glutamate | 2.11 | 1.45 | 1.5 | 0.009 | 0.022 |
| Citrate | 4.42 × 10−1 | 3.28 × 10−1 | 1.3 | 0.002 | 0.007 |
| Dibenzoyltartrate | 8.48 × 10−2 | 6.34 × 10−2 | 1.3 | 0.002 | 0.007 |
| Kaempferol | 5.52 × 10−4 | 4.45 × 10−4 | 1.2 | 0.026 | 0.056 |
| Metabolites | Abundance in Col-0 Seeds (Mean) | Abundance in vps29 Seeds (Mean) | Fold Change (Col-o/vps29) | p-Value | FDR-Adjusted p-Value |
|---|---|---|---|---|---|
| Xylitol | 4.14 × 10−3 | 1.08 × 10−1 | 26.15 | 0.002 | 0.007 |
| Sorbitol | 6.43 × 10−2 | 1.42 | 22.11 | 0.002 | 0.007 |
| Trehalose | 7.43 × 10−4 | 1.44 × 10−2 | 19.39 | 0.002 | 0.007 |
| Epicatechin | 2.33 × 10−3 | 3.50 × 10−2 | 15.04 | 0.002 | 0.007 |
| Putrescine | 6.39 × 10−3 | 5.62 × 10−2 | 8.79 | 0.002 | 0.007 |
| Alanine | 4.93 × 10−1 | 3.21 | 6.51 | 0.002 | 0.007 |
| Oleic acid | 1.18 × 10−3 | 7.40 × 10−3 | 6.25 | 0.002 | 0.007 |
| Anhydroglucose | 1.99 × 10−2 | 1.15 × 10−1 | 5.81 | 0.002 | 0.007 |
| Lysine | 8.74 × 10−3 | 4.93 × 10−2 | 5.64 | 0.002 | 0.007 |
| Gluconate | 9.07 × 10−1 | 5.07 | 5.59 | 0.002 | 0.007 |
| Gamma-hydroxybutyric acid | 1.81 × 10−4 | 9.00 × 10−4 | 4.96 | 0.002 | 0.007 |
| Pipecolate | 3.55 × 10−3 | 1.69 × 10−2 | 4.76 | 0.002 | 0.007 |
| Linoleic acid | 2.34 × 10−3 | 9.16 × 10−3 | 3.91 | 0.002 | 0.007 |
| Glycine | 1.51 × 10−1 | 5.63 × 10−1 | 3.73 | 0.002 | 0.007 |
| Myo-inositol | 3.54 × 10−1 | 1.21 | 3.41 | 0.002 | 0.007 |
| Succinate | 1.85 × 10−1 | 6.00 × 10−1 | 3.24 | 0.002 | 0.007 |
| Glycerate | 4.70 × 10−2 | 1.48 × 10−1 | 3.14 | 0.002 | 0.007 |
| Glycerol | 2.62 × 10−2 | 7.81 × 10−2 | 2.98 | 0.002 | 0.007 |
| Linolenic acid | 6.01 × 10−4 | 1.77 × 10−3 | 2.95 | 0.002 | 0.007 |
| Serine | 2.41 × 10−1 | 7.08 × 10−1 | 2.94 | 0.002 | 0.007 |
| Glycerol-3-phosphate | 1.43 × 10−2 | 3.95 × 10−2 | 2.76 | 0.002 | 0.007 |
| Arabitol | 4.38 × 10−2 | 1.11 × 10−1 | 2.53 | 0.002 | 0.007 |
| 7-(Methylthio)heptyl-glucosinolate | 1.78 × 10−3 | 4.29 × 10−3 | 2.41 | 0.039 | 0.079 |
| Threitol | 5.48 × 10−4 | 1.31 × 10−3 | 2.39 | 0.002 | 0.007 |
| Tyrosine | 2.56 × 10−2 | 5.97 × 10−2 | 2.33 | 0.002 | 0.007 |
| Homoserine | 2.44 × 10−3 | 5.55 × 10−3 | 2.27 | 0.002 | 0.007 |
| 8-(Methylthio)octyl-glucosinolate | 2.05 × 10−3 | 4.57 × 10−3 | 2.23 | 0.042 | 0.079 |
| 6-(Methylthio)hexanenitrile | 9.16 × 10−3 | 1.90 × 10−2 | 2.07 | 0.002 | 0.007 |
| Xylose | 5.31 × 10−2 | 1.04 × 10−1 | 1.96 | 0.002 | 0.007 |
| Unknown sugar (RI = 3891.7; Mass = 204) | 6.17 × 10−2 | 1.20 × 10−1 | 1.95 | 0.025 | 0.056 |
| Methionine | 4.39 × 10−2 | 8.49 × 10−2 | 1.93 | 0.002 | 0.007 |
| 8-(Methylthio)octanenitrile | 1.51 × 10−2 | 2.88 × 10−2 | 1.90 | 0.002 | 0.007 |
| Methionine | 4.49 × 10−3 | 8.25 × 10−3 | 1.84 | 0.026 | 0.056 |
| Mannose | 8.82 × 10−2 | 1.61 × 10−1 | 1.82 | 0.039 | 0.079 |
| Arginine | 2.86 × 10−1 | 5.15 × 10−1 | 1.80 | 0.002 | 0.007 |
| Galactonate | 5.81 × 10−2 | 1.01 × 10−1 | 1.75 | 0.008 | 0.020 |
| (Gamma-aminobutyric acid) GABA | 2.12 × 10−1 | 3.42 × 10−1 | 1.62 | 0.004 | 0.011 |
| 9-(Methylthio)nonanenitrile | 2.08 × 10−2 | 3.36 × 10−2 | 1.61 | 0.002 | 0.007 |
| Quercitrin | 2.67 × 10−2 | 4.24 × 10−2 | 1.58 | 0.026 | 0.056 |
| Valine | 5.31 × 10−1 | 8.07 × 10−1 | 1.52 | 0.002 | 0.007 |
| Eicosanoate | 3.60 × 10−3 | 5.45 × 10−3 | 1.51 | 0.041 | 0.079 |
| Allantoin | 2.07 × 10−2 | 3.12 × 10−2 | 1.51 | 0.004 | 0.011 |
| Galactinol | 5.63 × 10−1 | 8.38 × 10−1 | 1.49 | 0.027 | 0.056 |
| Sucrose | 3.36 × 101 | 4.96 × 101 | 1.48 | 0.029 | 0.061 |
| Alpha-aminoadipate | 4.38 × 10−2 | 6.38 × 10−2 | 1.46 | 0.026 | 0.056 |
| Threonate | 7.57 × 10−3 | 1.10 × 10−2 | 1.45 | 0.027 | 0.056 |
| Ribose | 6.02 × 10−2 | 8.62 × 10−2 | 1.43 | 0.009 | 0.022 |
| Allantoin | 4.78 × 10−2 | 6.58 × 10−2 | 1.38 | 0.027 | 0.056 |
| Leucine | 1.88 × 10−1 | 2.57 × 10−1 | 1.36 | 0.002 | 0.007 |
| Isoleucine | 2.32 × 10−1 | 2.98 × 10−1 | 1.28 | 0.008 | 0.021 |
| Cis-sinapinate | 1.03 × 10−2 | 1.28 × 10−2 | 1.25 | 0.041 | 0.079 |
| RAB Name a | AGI Gene b | RAB Abundance Ratio vps29/Col-0 |
|---|---|---|
| AtRABA1a | At1g06400 | 2.3 |
| AtRABA1b | At1g16920 | 2.5 |
| AtRABA1c | At5g45750 | 2.3 |
| AtRABA2a | At1g09630 | 2.1 |
| AtRABA2c | At3g46830 | 2.5 |
| AtRABA4a | At5g65270 | 3.6 |
| AtRABB1b | At4g35860 | 3.3 |
| AtRABB1c | At4g17170 | 2.1 |
| AtRABD1 | At3g11730 | 2.0 |
| AtRABD2a | At1g02130 | 2.4 |
| AtRABD2b | At5g47200 | 3.7 |
| AtRABD2c | At4g17530 | 3.2 |
| AtRABF1 | At3g54840 | 5.3 |
| AtRABF2b | At4g19640 | only detected in vps29 seeds |
| AtRABG3e | At1g49300 | 3.2 |
| AtRABG3f | At3g18820 | 3.1 |
| AtRABH1b | At2g44610 | 5.0 |
| AtRABH1c | At4g39890 | 4.3 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Durand, T.C.; Cueff, G.; Godin, B.; Valot, B.; Clément, G.; Gaude, T.; Rajjou, L. Combined Proteomic and Metabolomic Profiling of the Arabidopsis thaliana vps29 Mutant Reveals Pleiotropic Functions of the Retromer in Seed Development. Int. J. Mol. Sci. 2019, 20, 362. https://doi.org/10.3390/ijms20020362
Durand TC, Cueff G, Godin B, Valot B, Clément G, Gaude T, Rajjou L. Combined Proteomic and Metabolomic Profiling of the Arabidopsis thaliana vps29 Mutant Reveals Pleiotropic Functions of the Retromer in Seed Development. International Journal of Molecular Sciences. 2019; 20(2):362. https://doi.org/10.3390/ijms20020362
Chicago/Turabian StyleDurand, Thomas C, Gwendal Cueff, Béatrice Godin, Benoît Valot, Gilles Clément, Thierry Gaude, and Loïc Rajjou. 2019. "Combined Proteomic and Metabolomic Profiling of the Arabidopsis thaliana vps29 Mutant Reveals Pleiotropic Functions of the Retromer in Seed Development" International Journal of Molecular Sciences 20, no. 2: 362. https://doi.org/10.3390/ijms20020362
APA StyleDurand, T. C., Cueff, G., Godin, B., Valot, B., Clément, G., Gaude, T., & Rajjou, L. (2019). Combined Proteomic and Metabolomic Profiling of the Arabidopsis thaliana vps29 Mutant Reveals Pleiotropic Functions of the Retromer in Seed Development. International Journal of Molecular Sciences, 20(2), 362. https://doi.org/10.3390/ijms20020362