Mitochondria Are Important Determinants of the Aging of Seeds



1
Institute of Dendrology, Polish Academy of Sciences, Parkowa 5, 62-035 Kórnik, Poland
2
Department of Biotechnology, Institute of Molecular Biology and Biotechnology, Adam Mickiewicz University, 61-614 Poznań, Poland
3
Plant Physiology Department, Institute of Biology, University of Bialystok, Ciolkowskiego 1J, 15-245 Bialystok, Poland
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2019, 20(7), 1568; https://doi.org/10.3390/ijms20071568
Submission received: 28 February 2019
/
Revised: 26 March 2019
/
Accepted: 26 March 2019
/
Published: 28 March 2019
(This article belongs to the Special Issue Mitochondrial Dysfunction and Oxidative Damage)
Abstract
:Seeds enable plant survival in harsh environmental conditions, and via seeds, genetic information is transferred from parents to the new generation; this stage provides an opportunity for sessile plants to settle in new territories. However, seed viability decreases over long-term storage due to seed aging. For the effective conservation of gene resources, e.g., in gene banks, it is necessary to understand the causes of decreases in seed viability, not only where the aging process is initiated in seeds but also the sequence of events of this process. Mitochondria are the main source of reactive oxygen species (ROS) production, so they are more quickly and strongly exposed to oxidative damage than other organelles. The mitochondrial antioxidant system is also less active than the antioxidant systems of other organelles, thus such mitochondrial ‘defects’ can strongly affect various cell processes, including seed aging, which we discuss in this paper.
1. Aging Seeds
Seed aging, which decreases seed viability during storage, is a major problem for successful plant growth and productivity and leads to seed deterioration. In agricultural production, aged seeds cause commercial and genetic losses. The seed aging process is dependent primarily upon the moisture content of seeds, the oxygen level and the temperature at which seeds are stored [1,2,3,4] but is also associated with various metabolic and biophysical seed conditions [3,5]. Although the mechanisms of seed aging are still under intensive study, reactive oxygen species (ROS) are considered the main factor contributing to seed aging [6]. According to the free radical theory of aging, the generation and quantity of ROS are crucial for the progression of seed aging and other aging-associated disorders [7]. During the storage of seeds, the accumulation of ROS leads to the damage of lipids, DNA and proteins and consequently contributes to decreased germination and the loss of seed vigor [6,8,9,10,11,12,13]. It is claimed that for the effective conservation of genetic resources, it is necessary to identify the factors influencing aging of seeds that differ in sensitivity to drying and long-term storage. Based on their properties, seeds are divided into three categories: Orthodox (tolerating drying to a moisture content <7% and storage at −10 °C), recalcitrant (sensitive to drying to a moisture content <27% and conventional storage conditions), and intermediate (loosing viability relatively quickly, compared to orthodox seeds) [14,15]. These seed categories can also display various aging process timings or mechanisms.
2. Mitochondrial Activity in Aging Seeds
Mitochondria are the most important source of energy for cell growth and metabolism. In addition, mitochondria are the major sites for the production of reactive oxygen species (mtROS), and thus, are involved in the maintenance of ROS homeostasis [16,17,18,19]. ROS such as superoxide, hydrogen peroxide, and hydroxyl radicals, which oxidize cellular constituents, compromise organellar membrane integrity [20] and the main cell components [21,22]. The superoxide anion (O2−) is the precursor of most ROS and a mediator in oxidative chain reactions. Furthermore, O2− can dismute to produce hydrogen peroxide (H2O2) and may react with other radicals, including NO. The product of this reaction, peroxynitrite (ONOO−), is also a very powerful oxidant [23]. Hydrogen peroxide can freely migrate across cell membranes and can generate highly aggressive HO˙ [6,24]. mtROS are generated, in particular, in the context of respiratory electron transport activity. The main mtROS production sites are complex I and complex II of the mitochondrial electron transport chain (ETC). ETC-mediated ROS generation is primarily due to the presence of ubisemiquinone radicals, which can transfer a single electron to oxygen, giving rise to the production of superoxides [25]. In plants, alternative oxidase (AOX) plays a key role during stress by lowering ROS production from the ETC by preventing excessive reduction of the mitochondrial ubiquinone pool [20].
However, the relationship between the AOX pathway and ROS accumulation in the mitochondria of aged seeds has not yet been thoroughly studied [26]. In mitochondria, there are proteins that cause ROS generation: Glycerolphosphate dehydrogenase [27], multi-subunit pyruvate dehydrogenase complex and a structurally similar membrane-bound enzyme complex of alfa-ketoglutarate dehydrogenase (alfa-KGDH) [28]. Researchers have reported that succinate dehydrogenase (SDH) also contributes to mtROS production [26,29]. Aconitase, an enzyme in the mitochondrial matrix, is able to transform hydrogen peroxide into hydroxyl radicals during a Fenton reaction in the presence of iron and sulfur [30]. ROS are generated not only in the mitochondria but also by NADPH oxidase in the conversion of NADPH to NADP+ [23]. ROS are produced during germination in multiple species, where they have been proposed as a signal for release from seed dormancy [20].
Additionally, mitochondria are a source of reactive nitrogen species derived from nitric oxide (NO·). NO· is generated enzymatically by a family of nitric oxide synthases (NOS). These enzymes synthesize NO· using L-arginine as a substrate and NADPH as an electron source in the presence of Ca2+ ions and reduced thiols [21]. Mitochondrial electron transport can also produce NO [31,32]. NO has a short half-life but can react with thiols or/and the catalytic metal center of proteins, which results in the covalent modification of cysteine residues, termed S-nitrosylation. This modification also regulates many cellular processes in seeds/germination [31,32,33,34].
3. Oxidative Damage in Aging Seed Mitochondria
ROS-related mitochondrial dysfunction plays a vital role in seed deterioration, but the detailed mechanism of this role remains largely unknown. Most studies are based on the seeds of herbaceous plants (Table 1), and seeds of one tree species. The past ten years brings much more knowledge in this topic than could have been expected.
The accumulation of ROS causes oxidative damage and dysfunction and membrane system disorders, as well as the oxidative damage of mitochondrial proteins, DNA and lipids [45]. Oxidative damage of mitochondrial proteins include damage to subunits of the pyruvate decarboxylase complex, subunits of ATP synthase, and enzymes of the tricarboxylic acid (TCA) cycle [46]. ROS cause direct oxidation of amino acids, oxidation of Cys residues, which form disulphide bonds, oxidation of Met residues, which forms Met sulphoxide, and oxidation of arginine, lysine, proline, histidine, serine and threonine residues, which creates carbonyl groups in side chains [23,47]. The activity of mitochondrial proteins is regulated at the post-translational level, among others, e.g., S-nitrosylation [48,49]. During oxidative stress, the activity of mitochondrial proteins is lowered by the binding of lipid peroxidation products [50], carbonyl group formation [51] and the oxidation of tryptophan residues [52]. In elm seeds (Ulmus L.) 48 mitochondrial proteins changed during aging and found that these changes were associated with the tricarboxylic acid cycle (TCA) and mitochondrial ETC [44]. During oat seed aging, Moa et al. [26] also showed that proteins in the TCA cycle were down-regulated, and several enzymes related to mitochondrial ETC were up-regulated. Other authors [53] showed that the most recognized source of mtDNA mutagenesis are ROS, that are produced by the ETC. In addition, H2O2 induced strand breaks and abasic sites in mtDNA. The damage of main cell components by ROS leads to mitochondrial dysfunction through Bax induction and cytochrome c release [23].
Lipid peroxidation in the mitochondrial membranes refers to the free radical peroxidation of the polyunsaturated fatty acids of membrane lipids [54]. Malondialdehyde (MDA) and 4-hydroxy-2-nonenal (HNE) are products of lipid peroxidation and can interact with cells to reduce or even eliminate their functions. MDA can react with DNA bases, resulting in gene mutations, and HNE reacts mostly with proteins, leading to functional alterations [23]. Increases in mitochondrial MDA and H2O2 contents in moist tissues damage the mitochondrial membrane structure and their function [55].
The accumulation of oxidative damage is the basis of Harman’s free radical theory of aging [56]. One of the main sources of ROS in the cell is oxidative phosphorylation within mitochondria, so the free radical theory of aging may essentially be a mitochondrial theory of aging for plant seeds.
4. The Antioxidative System in Aging Seed Mitochondria
The balance between ROS generation and detoxification by antioxidants modulates the redox reactions in plant cells. Respiration and various other metabolic processes, including responses to oxidative stress, are co-controlled by the cellular redox state [57,58]. The plant mitochondrial matrix contains ROS scavenging systems, including enzymatic and nonenzymatic antioxidative systems [40], such as the manganese-superoxide dismutase (Mn-SOD) and ascorbate-glutathione (ASA-GSH) cycles [42], catalase (CAT) and peroxiredoxin (including peroxiredoxin IIF) [59]. During the aging of seeds, a decrease in the mitochondrial ASA-GSH cycle results in less reduced/oxidized forms of ASA and GSH, which might lead to ROS accumulation, affecting mitochondrial dysfunction [40]. Xia et al. showed in aging soybean seed mitochondria, that SOD, ascorbate peroxidases (APX), monodehydroascorbate reductase (MDHAR), and glutathione reductase (GR) activities decreased with prolonged aging.
Mao et al. [26] suggested that mitochondrial structure changes are responsible for decreases in antioxidant enzyme activity in aged seeds. The authors observed decreases in the activities of GR, dehydroascorbate reductase (DHAR) and MDHAR, which were accompanied by damage to the inner mitochondrial membrane in seeds during aging [26,42]. The observed catalase (CAT) is believed to be associated with the mechanism of seed aging [6,60], which actively removes H2O2 [61] and modulates the associated signaling pathways [62,63]. CAT levels decrease in the mitochondria of Fagus sylvatica L. seeds during their natural aging process. The decrease in the activity of this enzyme is accompanied by an increase in the H2O2 level (Ratajczak et al., results not published).
The active adjustment of the redox state in the mitochondria is important for the normal course of the combined metabolic processes of photosynthesis and respiration in green tissues. Ascorbate (ASA) and glutathione (GSH) play an important role in regulating the redox state in the mitochondria. Moreover, they act together in the ASA–GSH cycle, which is a core component of the antioxidant system in plant mitochondria, and have other functions that are important for the mitochondrion. ASA also participates in redox regulation in the modulation of gene expression, in the regulation of enzymatic activity, and in cell signaling [4,64,65,66,67,68]. Changes in mitochondrial ascorbate synthesis may modulate communication between plastids and mitochondria [65,69]. Mitochondrial GSH levels depend on the transport of glutathione into the mitochondria because GSH is mainly synthesized in the cytosol and plastids [70]. There is little information about plant mitochondrial glutathione transporters. The level of mitochondrial GSH is highly dependent on the activity of the enzyme glutathione reductase (GR), which reduces GSH to its oxidized form glutathione (GSSG) [71]. GSH deficiency in the mitochondria can cause mitochondrial damage, affecting changes in the synthesis of thiol proteins and thus changes in redox regulation in mitochondrion cells. The potential E = 2GSH/GSSG redox state in mitochondria, which ranges from –340 to −300 mV, provides a unique environment that affects the corresponding thiol modification of proteins in the mitochondria [69].
ROS that are produced in the mitochondrial matrix are detoxified by thiol-based peroxidase systems, such as peroxiredoxin (Prx) [72]. Prx family members have an important role in regulating and maintaining the redox balance in seed cells [73,74]. Plant mitochondria contain an atypical type II Prx (Prx IIF) [25,59,75]. Prx IIF is a peroxidase that accepts electrons from a broad range of donors and functions principally in the reduction H2O2, which catalyzes the detoxification of ROS in the following order of efficiency: H2O2v > tertiary butyl hydroperoxide (-BOOH) > cumene hydroperoxide (CuCOOH) [75,76].
We found that the level of the protein Prx IIF decreased in the mitochondria of beech (Fagus sylvatica L.) seeds during the natural aging process (Ratajczak, results not published). We also observed differences in Prx IIF transcript and protein levels, as well as in the level of post-translational modification between Norway maple (Acer platanoides L.) seeds (orthodox seeds) and sycamore (A. pseudoplatanus L.) seeds (recalcitrant seeds) during desiccation [77]. It has been proposed that the redox homeostasis of mitochondria in seeds is a necessary feature to maintain high seed viability.
Another possible regulator of the redox state in plant mitochondria is the thioredoxin (Trx) system [78]. The Trx system consists of o-typ Trx (Trx-o) and NADPH-thioredoxin reductase (Figure 1) [79,80]. Thioredoxins (Trx), a regulatory disulfide protein, is a substrate for enzymes that catalyze reactions [78] and regulatory reactions that alter the activity or other functional properties of interaction target proteins [81]. The reduced form of Trx interacts with a variety of target proteins and performs regulatory functions [82].
Members of the Trx system in higher plants are divided into groups: m, f, x, y, o and h [83,84]. Trx o is localized in the mitochondria, and Trx h is typically cytosolic but has also been identified in other cellular compartments, including the mitochondrion [84]. Trx participates in the regulation of 12 mitochondrial processes, ranging from energetics and metabolism reactions to protein synthesis, stress responses and communication with other organelles [79,85]. It has been proposed that Trx functions not only to regulate biochemical processes under optimal conditions but also to restore the function of activities after oxidative stress (adaptation to stress). Sanz-Barrio et al. [86] showed that Trx acts in plant mitochondria as a molecular chaperone. The role of Trx in the regulation of the redox state in tree seeds, which is characterized by a different sensitivity to water loss, is not yet fully understood.
5. Is Mitochondrial Dysfunction the Cause of Seed Aging?
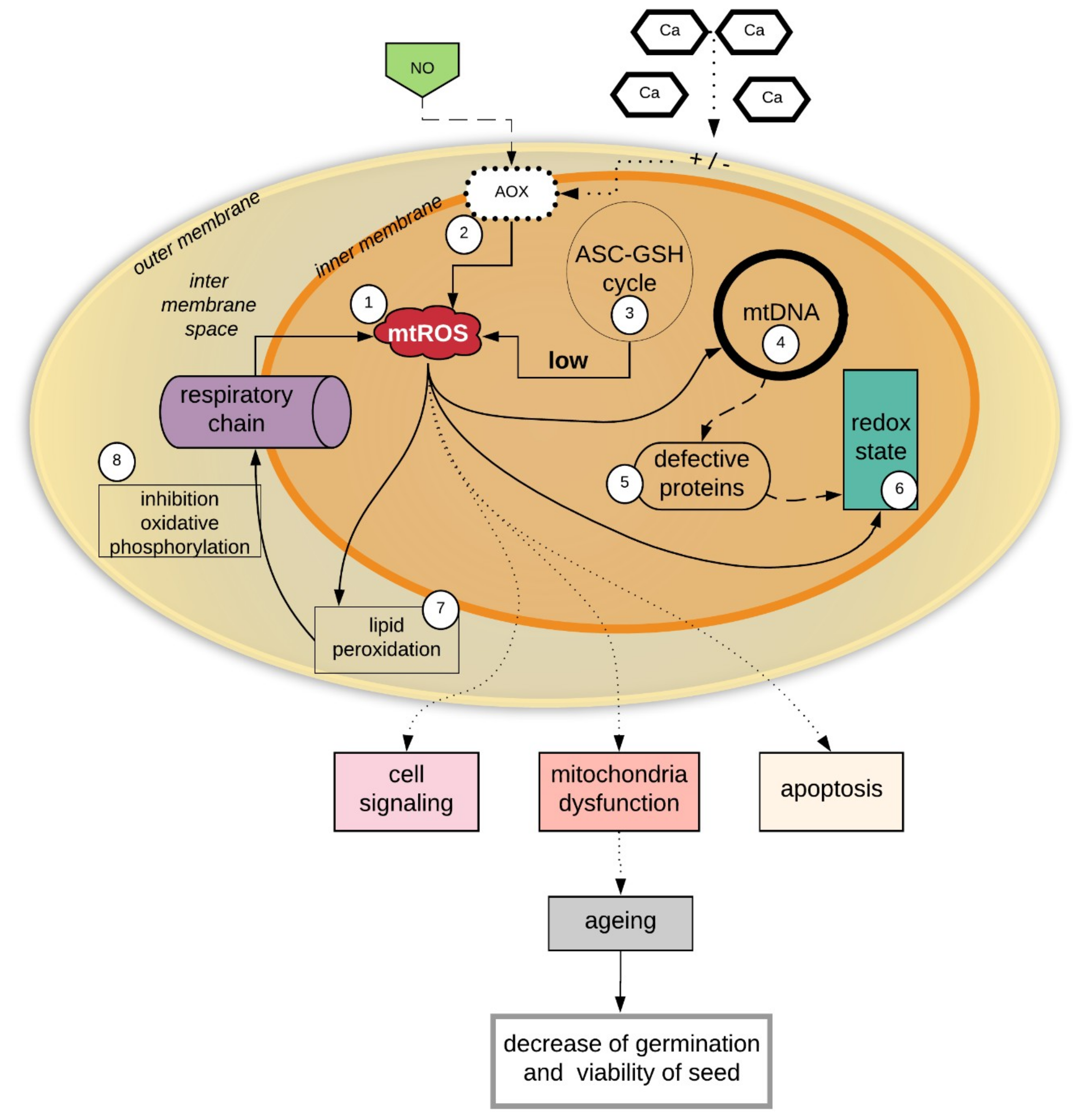
The functions of mitochondria in cell signaling events and inter-organelle communication and aging are already well known, especially in animal cells [87,88]. The role of mitochondria in the plant seed aging processes has not yet been well described. It is important to determine whether mitochondrial dysfunction imitates seed aging and whether mitochondrial dysfunction is the result of seed aging during storage. It is believed that the dysfunction of mitochondria, coupled with plant seed aging, is as is presented in Figure 1.
The effects of mitochondrial damage in seeds during storage, which finally leads to seed aging and the reduction of seed viability, are summarized in Figure 1. (1) Mitochondria are the sites of continuous ROS generation. (3) During the storage of seeds, the ROS level increases, mainly in the mitochondria, and the excess of ROS may not be effectively removed due to the low activity of the mitochondrial antioxidant system (i.e., the ASA-GSH cycle) compared to the antioxidant systems of other cell organelles. The ROS-induced impairment of mitochondria leads to increased oxidant production and oxidative damage. A high level of ROS in the mitochondria causes oxidative damage to the membranes, which affects the inhibition of oxidative phosphorylation. (4) ROS also cause oxidative damage to mtDNA. mtDNA is constantly exposed to oxidative injury, mainly due to the location of mtDNA in the inner mitochondrial membrane, which exposes it to the influence of ROS and makes mtDNA more exposed than nuclear DNA to oxidative damage [89]. Most studies on aging concern human cells, while research on the mtDNA in plant seeds is uncommon. However, damage to plant seed mtDNA is a very important problem for genome reserve conservation because a lack of genome integrity affects seed viability [90]. The stability of mtDNA depends on the production of mitochondrial ROS (mtROS), which are generated during normal electron flux via mitochondrial electron transport [89,91]. mtDNA is not protected by the protein membrane, and histones are not associated with mtDNA it, which makes mtDNA more sensitive to increases in ROS levels [92]. In aging seeds, as was shown in Figure 2, the accumulated mtROS (1) induces damage to the mtDNA (4) and adversely affects the synthesis of mitochondrial proteins (5), including proteins regulating the mitochondrion redox state, e.g., peroxyredoxins (Prxs) and thioredoxins (Trxs). The accumulation of mtROS thus influences changes in signaling and the redox status in the mitochondria (6). (7) A high level of ROS in the mitochondria causes oxidative damage to membranes, which affects the inhibition of oxidative phosphorylation (8). The sum of all adverse reactions (1–8) causes a decrease in seed viability (Figure 1).
The cellular ROS levels and redox status of mitochondria regulate mitochondrial and nuclear gene expression, which are pivotal to seed aging [7,76]. Generally, changes in the mitochondrial redox state will affect not only mitochondrial activity but also cellular processes, such as photosynthesis, stress defenses and the activation of programmed cell death [76].
One potential regulator of mitochondrial activity during aging is the second messenger Ca2+. By stimulating Ca2+-dependent dehydrogenases of the tricarboxylic acid (TCA) cycle, Ca2+ boosts the activity of the mitochondrial respiration chain and, consequently, the mitochondrial adenosine triphosphate (ATP) production via oxidative phosphorylation. By stimulating the mitochondrial respiration chain, matrix Ca2+ also maintains the stability of the mitochondrial membrane potential, which is temporarily dissipated by entering Ca2+. However, in the case of overwhelming mitochondrial Ca2+ accumulation, the permeability of the inner mitochondrial membrane increases drastically, resulting in the dissipation of the mitochondrial membrane potential, the shutdown of mitochondrial respiration and finally, the initiation of cell death signaling pathways [93]. Wang and coauthors [94] indicated that the Ca2+ flux could be part of the AOX retrograde response.
Xin et al. showed that aging severely affects the rate of NADH and succinate-dependent O2 consumption and the respiration control rate, suggesting that aged seeds possess a lower capacity than control seeds for the electron transport chain. Aging directly reduces the efficiency of electron transport chains, thereby reducing ATP production, so aged seeds cannot provide sufficient ATP for germination. Upon stress exposure in mitochondria, an energy deficit signal occurs, which leads to global changes in organellar and nuclear gene expression [20].
6. Mitochondrial Structure in the Process of Seed Aging
The presence of two membranes—the outer membrane (OM) and inner membrane (IM)—with very different compositions and conformations, suggests that the membranes have diverse contributions to mitochondrial function and physiology. Mitochondria modulate their functions and status and allow complex quality control. Recent discoveries have shown a correlation between the modulation of mitochondrial shape and network and the energetic state of the cell. Oxidative stress causes mitochondrial elongation, protecting mitochondria from degradation and promoting mitochondrial ATP production [95]. Xia et al. suggested there are relationships between antioxidative systems and and mitochondrial ultrastructure in aging seeds. The authors of this study used transmission electron microscopy to observe that mitochondrial ultrastructure was damaged in aging and the degree of damage was related to the level of seed moisture. Increasing the moisture content from 4% to 16% in the seeds at 0, 16 and 40 days caused the cristae to no longer be visible. Others authors [44], by using fluorescence microscopy, showed that the mitochondrial distribution and morphology changes gradually with seed aging in Ulmus pumila L. These authors noticed that mitochondrial aggregation in the early aging stage is related to mitochondrial endogenous ROS production. Yin and co-authors indicated that the integrity was highly inhibited in rice embryos aged seven days, in comparison to rice embryos aged 0 and 4 days, and the oldest embryos possessed numerous mature mitochondria with typical structures of well-developed cristae.
Using electron microscopy, Noctor et al. [96] showed that mitochondria, which have elaborate cristae in young and mature Arabidopsis rosette tissues, only lose their internal structure with swollen cristae at the final programed cell death (PCD) stage of senescence, when most cellular proteins and other reserves have been degraded and exported. Electron cryotomography of mitochondria isolated from aging Podospora anserina revealed a sequence of events, namely, the progressive vesiculation of the mitochondrial inner membrane, the collapse of the cristae, disassembly of ATP synthase dimers, and formation of large contact sites between the inner and outer mitochondrial membranes [97].
It is likely that changes in the structure of mitochondria caused by ROS generation and mitochondrial dysfunctions trigger different responses that regulate mitochondrial and nuclear gene expression, which are pivotal to seed aging.
7. Conclusions
We believe that a thorough analysis of seed mitochondria in various tissues will bring us closer to understanding the causes of the seed aging process. It is important to perform these analyses on seeds that have different sensitivities to water loss, storage conditions and long-term storage, i.e., in orthodox, recalcitrant and intermediate seeds. We suggest that more damage to mitochondria will occur in the embryonic axes than in the cotyledons of seeds.
The mitochondria are very important cellular organelles, whose main purpose is to generate energy in the process of cellular respiration—in the form of ATP—during the process of cellular respiration. In mitochondria, seed aging increases the level of ROS, which causes numerous organelle dysfunctions. This contributes to increases in the level of oxidative damage to the main cellular components, decreases in enzyme activities due to the oxidation of the functional groups, and increases membrane lipid peroxidation. During the aging of seeds, ROS initiates gene expression, which is responsible for programmed cell death. In addition, physiological and biochemical changes affect changes in the structures of mitochondria, which determines the action of these organelles. Mitochondria are well-suited for sensing functional imbalances. Their respiratory machinery, which is based on redox chemistry, can react sensitively to changing conditions. At the final stage of mitochondrial aging, the disruption of this arrangement results in the ability of mitochondria to produce ATP. In effect, aging cells with an increasing proportion of dysfunctional mitochondria are less fit than non-aging cells and eventually die. Understanding the mechanisms of seed aging will lead to new methods for seed conservation and longevity.
Author Contributions
Conceptualization E.R., Formal analysis E.R.; A.M.; A.M.S; Investigation, E.R.; A.M.; A.M.S; Writing-Original Draft Preparation, E.R.; A.M.; A.M.S.; Writing-Review & Editing, E.R.; A.M.; A.M.S; I.C.; Visualization, E.R.; A.M.S.; Supervision, E.R.; Funding Acquisition, E.R.
Funding
This research funding was provided by the Institute of Dendrology of the Polish Academy of Sciences.
Conflicts of Interest
The authors declare that they have no conflict of interests.
References
- Bewley, J.D.; Bradford, K.J.; Hilhorst, H.M.W.; Nonogaki, H. Seeds: Physiology of Development, Germination and Dormancy; Springer: New York, NY, USA, 2013. [Google Scholar]
- Groot, S.P.C.; Surki, A.A.; de Vos, R.C.H.; Kodde, J. Seed storage at elevated partial pressure of oxygen, a fast method for analysing seed aging under dry conditions. Ann. Bot. 2012, 110, 1149–1159. [Google Scholar] [CrossRef] [PubMed]
- Walters, C.; Ballesteros, D.; Vertucci, V.A. Structural mechanics of seed deterioration: Standing the test of time. Plant Sci. 2010, 6, 565–573. [Google Scholar] [CrossRef]
- Pukacka, S.; Ratajczak, E. Age-related biochemical changes during storage of beech (Fagus sylvatica L.) seeds. Seed Sci. Res. 2007, 17, 45–53. [Google Scholar] [CrossRef]
- Ballesteros, D.; Walters, C. Detailed characterization of mechanical properties and molecular mobility within dry seed glasses: Relevance to the physiology of dry biological systems. Plant J. 2011, 68, 607–619. [Google Scholar] [CrossRef]
- Ratajczak, E.; Małecka, A.; Bagniewska-Zadworna, A.; Kalemba, E.M. The production, localization and spreading of reactive oxygen species contributes to the low vitality of long-term stored common beech (Fagus sylvatica L.) seeds. J. Plant Physiol. 2015, 174, 147–156. [Google Scholar] [CrossRef]
- Yin, G.K.; Whelan, J.; Wu, S.H.; Zhou, J.; Chen, B.; Chen, X.L.; Zhang, J.M.; He, X.; Xin, J.J.; Lu, X.X. Comprehensive mitochondrial metabolic shift during the critical node of seed aging in rice. PLoS ONE 2016, 11, 1–19. [Google Scholar] [CrossRef]
- Bailly, C.; El-Maarouf-Bouteau, H.; Corbineau, F. From intracellular signaling networks to cell death: the dual role of reactive oxygen species in seed physiology. C RBiol. 2008, 331, 806–814. [Google Scholar] [CrossRef]
- Rajjou, L.; Lovigny, Y.; Grott, S.P.C.; Belghazi, M.; Job, C.; Job, D. Proteome-wide characterization of seed aging in Arabidopsis. A comparison between artificial and natural aging protocols. Plant Physiol. 2008, 148, 20–41. [Google Scholar] [CrossRef]
- Bailly, C.; Kranner, I. Analyses of reactive oxygen species and antioxidants in relation to seed longevity and germination. Methods Mol. Biol. 2011, 773, 343–367. [Google Scholar] [PubMed]
- Bellani, M.; Salvini, L.; Dell’Aquila, A.; Scialabba, A. Reactive oxygen species release, vitamin E, fatty acid and phytosterol content of artificially aged radish (Raphanus sativus L.) seeds during germination. Acta. Physiol. Plant. 2012, 34, 1789–1799. [Google Scholar] [CrossRef]
- Hu, D.; Ma, G.; Wang, Q.; Yao, J.; Wang, Y.; Pritchard, H.W.; Wang, X. Spatial and temporal nature of reactive oxygen species production and programmed cell death in elm (Ulmus pumila L.) seeds during controlled deterioration. Plant Cell Environ. 2012, 35, 2045–2059. [Google Scholar] [CrossRef] [PubMed]
- Yao, Z.; Liu, L.; Gao, F.; Rampitsch, C.; Reinecke, D.M.; Ozga, J.A.; Ayele, B.T. Developmental and seed aging mediated regulation of antioxidative genes and differential expression of proteins during pre- and post-germinative phases in pea. J. Plant Physiol. 2012, 169, 1477–1488. [Google Scholar] [CrossRef] [PubMed]
- Ellis, R.H.; Hong, D.; Roberts, E.H. An Intermediate Category of Seed Storage Behaviour?: I. Coffee. J. Exp. Bot. 1990, 41, 1167–1174. [Google Scholar] [CrossRef]
- Berjak, P.; Pammenter, N.W. From Avicennia to Zizania: Seed recalcitrance in perspective. Ann. Bot. Lond. 2008, 101, 213–228. [Google Scholar] [CrossRef] [PubMed]
- Møller, I.M. Plant mitochondria and oxidative stress. Electron transport, NADPH turnover and metabolism of reactive oxygen species. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2001, 52, 561–569. [Google Scholar] [CrossRef]
- Howell, K.A.; Millar, A.H.; Whelan, J. Ordered assembly of mitochondria during rice germination begins with pro-mitochondrial structures rich in components of the protein import apparatus. Plant Mol. Biol. 2006, 60, 201–223. [Google Scholar] [CrossRef] [PubMed]
- Navrot, N.; Rouhier, N.; Gelhaye, E.; Jacquot, J.-P. ROS generation and antioxidant systems in plant mitochondria. Physiol. Plant. 2007, 129, 185–195. [Google Scholar] [CrossRef]
- Dunn, J.D.; Alvarez, L.A.J.; Zhang, X.; Soldati, T. Reactive oxygen species and mitochondria: A nexus of cellular homeostasis. Redox Biol. 2015, 6, 472–485. [Google Scholar] [CrossRef]
- Liberatore, K.L.; Dukowic-Schulze, S.; Miller, M.E.; Chen, C.; Kianian, S.F. The role of mitochondria in plant development and stress tolerance. Free Rad. Biol. Med. 2016, 100, 238–256. [Google Scholar] [CrossRef] [PubMed]
- Kowaltowski, A.J.; de Souza-Pinto, N.C.; Castilho, R.F.; Vercesi, A.E. Mitochondria and reactive oxygen species. Free Rad. Biol. Med. 2009, 47, 333–343. [Google Scholar] [CrossRef] [PubMed]
- Lismont, C.; Nordgren, M.; VanVeldhoven, P.P.; Franse, M. Redox interplay between mitochondria and peroxisomes. Front Cell Dev. Biol. 2015, 3. [Google Scholar] [CrossRef] [PubMed]
- Slimen, I.B.; Najar, T.; Ghram, A.; Dabbebi, H.; Mrad, M.B.; Abdrabbah, M. Reactive oxygen species, heat stress and oxidative induced mitochondrial damage. A review. Int. J. Hyperth. 2014, 30, 513–523. [Google Scholar] [CrossRef] [PubMed]
- Bienert, G.P.; Schjoerring, J.K.; Jahn, T.P. Membrane transport of hydrogen peroxide. Biochim. Biophys. Acta. 2006, 1758, 994–1003. [Google Scholar] [CrossRef]
- Barranco-Medina, S.; Krell, T.; Finkemeier, I.; Sevilla, F.; Lazaro, J.J.; Dietz, K.J. Biochemical and molecular characterization of the mitochondrial peroxiredoxin PsPrxII F from Pisum sativum. Plant Physiol. Biochem. 2007, 45, 729–739. [Google Scholar] [CrossRef] [PubMed]
- Mao, C.; Zhu, Y.; Cheng, H.; Yan, H.; Zhao, L.; Tang, J.; Ma, X.; Mao, P. Nitric oxide regulates seedling growth and mitochondrial responses in aged oat seeds. Int. J. Mol. Sci. 2018, 19, 1052. [Google Scholar] [CrossRef]
- Drahota, Z.; Chowdhury, S.K.R.; Floryk, D.; Mracek, T.; Wilhelm, J.; Rauchova, H. Glycerophosphate-dependent hydrogen peroxide production by brown adiposetissue mitochondria and its activation by ferricyanide. J. Bioenerg. Biomembr. 2002, 34, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Starkov, A.A.; Fiskum, G.; Chinopoulos, C.; Lorenzo, B.J.; Browne, S.E.; Patel, M.S. Mitochondrial alpha-ketoglutarate dehydrogenase complex generates reactive oxygen species. J. Neurosci. 2004, 24, 7779–7788. [Google Scholar]
- Jardim-Messeder, D.; Caverzan, A.; Rauber, R.; de Souza Ferreira, E.; Margis-Pinheiro, M.; Galina, A. Galina Succinate dehydrogenase (mitochondrial complex II) is a source of reactive oxygen species in plants and regulates development and stress responses. New Phytol. 2015, 208, 776–789. [Google Scholar] [CrossRef] [PubMed]
- Holzerová, E.; Prokisch, H. Mitochondria: Much ado about nothing? How dangerous is reactive oxygen species production? Int. J. Biochem. Cell Biol. 2015, 63, 16–20. [Google Scholar] [CrossRef]
- Gupta, K.J. Protein S-nitrosylation in plants: photorespiratory metabolism and NO signaling. Sci. Signal. 2011, 154, jc1. [Google Scholar] [CrossRef] [PubMed]
- Gupta, K.J.; Igamberdiev, A.U. Reactive nitrogen species in mitochondria and their implications in plant energy status and hypoxic stress tolerance. Front Plant Sci. 2016, 7, 369. [Google Scholar] [CrossRef]
- Krasuska, U.; Ciacka, K.; Andryka, P.; Bogatek, R.; Gniazdowska, A. „Nitrosative door” in seed dormancy alleviation and germination. In Reactive oxygen and nitrogen species signaling and communication in plants; Gupta, K.J., Igamberdiev, A.U., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 215–237. [Google Scholar]
- Krasuska, U.; Ciacka, K.; Orzechowski, S.; Fettke, J.; Bogatek, R.; Gniazdowska, A. Modification of the endogenous NO level influences apple embryos dormancy by alterations of nitrated and biotinylated protein patterns. Planta 2016, 244, 877–891. [Google Scholar] [CrossRef] [PubMed]
- Harman, G.; Granett, A.L. Deterioration of stored pea seed: Changes in germination, membrane permeability and ultrastructure resulting from infection by Aspergillus ruber and from aging. Physiol. Plant Path. 1972, 3, 271–276. [Google Scholar] [CrossRef]
- Hallam, N.D.; Roberts, B.E.; Osborne, D.J. Embryogenesis and germination in rye (Secale cereale L.)—III. Fine structure and biochemistry of the non-viable embryo. Planta 1973, 110, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Parrish, D.J.; Leopold, A.C. On the mechanism of aging in soybean seeds. Plant Physiol. 1978, 61, 365–368. [Google Scholar] [CrossRef] [PubMed]
- Yin, G.; Sun, H.; Xin, X.; Qin, G.; Liang, Z.; Jing, X. Mitochondrial damage in the soybean seed axis during imbibition at chilling temperatures. Plant Cell Physiol. 2009, 50, 1305–1318. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.-Q.; Cheng, H.-Y.; Møller, I.M.; Song, S.-Q. The role of recovery of mitochondrial structure and function in desiccation tolerance of pea seeds. Physiol. Plant. 2012, 144, 20–34. [Google Scholar] [CrossRef] [PubMed]
- Xin, X.; Tian, Q.; Yin, G.; Chen, X.; Zhang, J.; Ng, S.; Lu, X. Reduced mitochondrial and ascorbate-glutathione activity after artificial aging in soybean seed. J. Plant Physiol. 2014, 171, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, Y.; Xue, H.; Pritchard, H.W.; Wang, X. Reactive oxygen species—provoked mitochondria—dependent cell death during aging of elm (Ulmus pumila L.) seeds. Plant J. 2015, 81, 438–452. [Google Scholar] [CrossRef] [PubMed]
- Xia, F.S.; Wang, M.Y.; Li, M.L.; Mao, P.S. Mitochondrial structural and antioxidant system responses to aging in oat (Avena sativa L.) seeds with different moisture contents. Plant Physiol. Biochem. 2015, 94, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Fu, H.; Zhou, X.; Chen, Z.; Luo, Y.; Cui, B.; Chen, G.; Liu, J. Comparative proteomic analysis of seed embryo proteins associated with seed storability in rice (Oryza sativa L.) during natural aging. Plant Physiol. Biochem. 2016, 103, 31–44. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, Y.; Xue, H.; Pritchard, H.W.; Wang, X. Changes in the mitochondrial protein profile due to ROS eruption during aging of elm (Ulmus pumila L.) seeds. Plant Physiol Biochem. 2017, 114, 72–87. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Osuna, D.; Colville, L.; Lorenzo, O.; Graeber, K.; Dennis, E.S.; Peacock, W.J. Transcriptome-wide mapping of pea seed aging reveals a pivotal role for genes related to oxidative stress and programmed cell death. PLoS ONE 2013. [Google Scholar] [CrossRef]
- Rhoads, D.M.; Umbach, A.L.; Subbaiah, C.C.; Siedow, J.N. Mitochondrial reactive oxygen species. Contribution to oxidative stress and interorganellar signaling. Plant Physiol. 2006, 141, 357–366. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Prabhakaran, S.A. Mechanism, measurement and prevention of oxidative stress in male reproductive physiology. Indian J. Exp. Biol. 2005, 43, 963–974. [Google Scholar] [PubMed]
- Bykova, N.V.; Egsgaard, H.; Møller, I.M. Identification of 14 new phosphoproteins involved in important plant mitochondrial processes. FEBS Lett. 2003, 540, 141–146. [Google Scholar] [CrossRef]
- Ito, J.; Heazlewood, J.L.; Millar, A.H. The plant mitochondrial proteome and the challenge of defining the post-translational modifications responsible for signalling and stress effects on respiratory functions. Physiol. Plant. 2007, 129, 207–224. [Google Scholar] [CrossRef]
- Winger, A.M.; Taylor, N.L.; Heazlewood, J.H.; Day, D.A.; Millar, A.H. The cytotoxic lipid peroxidation product 4-hydroxy-2-nomenal covalently modifies a selective range of proteins linked to respiratory function in plant mitochondria. J. Biol. Chem. 2007, 282, 37436–37447. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, B.K.; Askerlund, P.; Bykova, N.V.; Egsgaard, H.; Møller, I.M. Identification of oxidised proteins in the matrix of rice leaf mitochondria by immunoprecipitation and two-dimensional liquid chromatography-tandem mass spectrometry. Phytochemistry 2004, 65, 1839–1851. [Google Scholar] [CrossRef] [PubMed]
- Moller, I.M.; Kristensen, B.K. Protein oxidation in plant mitochondria as a stress indicator. Photochem. Photobiol. Sci. 2004, 3, 730–735. [Google Scholar] [CrossRef]
- Shokolenko, I.; Venediktova, N.; Bochkareva, A.; Wilson, G.L.; Alexeyev, M.F. Oxidative stress induces degradation of mitochondrial DNA. Nucleic Acids Res. 2009, 37, 2539–2548. [Google Scholar] [CrossRef] [PubMed]
- Jacoby, RP.; Li, L.; Huang, S.; Pong Lee, C.; Millar, A.H.; Taylor, N.L. Mitochondrial composition, function and stress response in plants. J. Integr. Plant Biol. 2012, 11, 887–906. [Google Scholar] [CrossRef] [PubMed]
- Zhan, J.; Li, W.; He, H.Y.; Li, C.Z.; He, L.F. Mitochondrial alterations during Al-induced PCD in peanut root tips. Plant Physiol. Biochem. 2014, 7, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Harman, D. Free radical theory of aging: an update: increasing the functional lifespan. Ann. N. Y. Acad. Sci. 2006, 1067, 10–21. [Google Scholar] [CrossRef] [PubMed]
- Oelze, M.L.; Kandlbinder, A.; Dietz, K.J. Redox regulation and overreduction control in the photosynthesizing cell: Complexity in redox regulatory networks. Bioch. Bioph. Acta. 2008, 1780, 1261–1272. [Google Scholar] [CrossRef] [PubMed]
- Ratajczak, E.; Dietz, K.J. Regulatory redox state in tree seeds. Acta Soc. Bot. Poloniae 2017, 86, 35–67. [Google Scholar] [CrossRef]
- Gama, F.; Keech, O.; Eymery, F.; Finkemeier, I.; Gelhaye, E.; Gardeströ>m, P.; Dietz, K.J.; Rey, P.; Jacquot, J.-P.; Rouhie, N. The mitochondrial type II peroxiredoxin from poplar. Physiol. Plant 2007, 129, 196–206. [Google Scholar] [CrossRef]
- Bailly, C. Active oxygen species and antioxidants in seed biology. Seed Sci. Res. 2004, 14, 93–107. [Google Scholar] [CrossRef]
- Kibinza, S.; Bazin, J.; Bailly, C.; Farrant, J.M.; Corbineau, F.; El-Maarouf-Bouteau, H. Catalase is a key enzyme in seed recovery from aging during priming. Plant Sci. 2011, 181, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 9, 405–410. [Google Scholar] [CrossRef]
- Neill, S.J.; Desikan, R.; Clarke, A.; Hurst, R.D.; Hancock, J.T. Hydrogen peroxide and nitric oxide as signalling molecules in plants. J. Exp. Bot. 2002, 372, 1237–1247. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Redox regulation and photosynthetic organisms: Signaling, acclimation and practical implications. Antiox. Redox. Signal. 2009, 11, 861–905. [Google Scholar] [CrossRef] [PubMed]
- Talla, S.; Riazunnisa, K.; Padmavathi, L.; Sunil, B.; Rajsheel, P.; Raghavendra, A.S. Ascorbic acid is a key participant during the interactions between chloroplasts and mitochondria to optimize photosynthesis and protect against photoinhibition. J. Biosci. 2011, 36, 163–173. [Google Scholar] [CrossRef]
- Kerchev, P.I.; Pellny, T.K.; Vivancos, P.D.; Kiddle, G.; Hedden, P.; Driscoll, S.; Vanacker, H.; Verrier, P.; Hancock, R.D.; Foyer, C.H. The transcription factor ABI4 Is required for the ascorbic acid-dependent regulation of growth and regulation of jasmonate-dependent defense signaling pathways in Arabidopsis. Plant Cell. 2011, 23, 3319–3334. [Google Scholar] [CrossRef] [PubMed]
- Zechmann, B. Subcellular distribution of ascorbate in plants. Plant Sign. Behav. 2011, 6, 360–363. [Google Scholar] [CrossRef]
- Akram, N.A.; Shafiq, F.; Ashraf, M. Ascorbic Acid-A potential oxidant scavenger and its role in plant development and abiotic stress tolerance. Front Plant Sci. 2017, 26, 613. [Google Scholar] [CrossRef] [PubMed]
- Schwarzländer, M.; Finkemeier, I. Mitochondrial energy and redox signaling in plants. Antioxid. Redox. Signal. 2013, 18, 2122–2144. [Google Scholar] [CrossRef] [PubMed]
- Chew, O.; Whelan, J.; Millar, A.H. Molecular definition of the ascorbate-glutathione cycle in Arabidopsis mitochondria reveals dual targeting of antioxidant defenses in plants. J. Biol. Chem. 2003, 278, 46869–46877. [Google Scholar] [CrossRef]
- Mailloux, R.J.; Jin, X.; Willmore, W.G. Redox regulation of mitochondrial function with emphasis on cysteine oxidation reactions. Redox Biol. 2013, 19, 123–139. [Google Scholar] [CrossRef]
- Barranco-Medina, S.; Kakorin, S.; Lázaro, J.J.; Dietz, K.J. Thermodynamics of the dimer–decamer transition of reduced human and plant 2-Cys peroxiredoxin. Biochemistry 2008, 47, 7196–7204. [Google Scholar] [CrossRef] [PubMed]
- Dietz, K.J. Peroxiredoxins in plants and cyanobacteria. Antiox. Redox Sign. 2011, 15, 1129–1159. [Google Scholar] [CrossRef] [PubMed]
- Ratajczak, E.; Dietz, K.J.; Kalemba, E.M. The occurrence of peroxiredoxins and changes in redox state in Acer platanoides and Acer pseudoplatanus during seed development. J. Plant Grow. Regul. 2018. [Google Scholar] [CrossRef]
- Finkemeier, I.; Goodman, M.; Lamkemeyer, P.; Kandlbinder, A.; Sweetlove, L.J.; Dietz, K.J. The mitochondrial type II peroxiredoxin F is essential for redox homeostasis and root growth of Arabidopsis thaliana under stress. J. Biol. Chem. 2005, 280, 12168–12180. [Google Scholar] [CrossRef] [PubMed]
- Rouhier, N.; Jacquot, J.P. The plant multigenic family of thiol peroxidases. Free Radic. Biol. Med. 2005, 38, 1413–1421. [Google Scholar] [CrossRef] [PubMed]
- Ratajczak, E.; Ströher, E.; Oelze, M.L.; Kalemba, E.M.; Pukacka, S.; Dietz, K.J. The involvement of the mitochondrial peroxiredoxin PrxIIF in defining physiological differences between orthodox and recalcitrant seeds of two Acer species. Funct. Plant Biol. 2013, 40, 1005–1017. [Google Scholar] [CrossRef]
- Buchanan, B.B.; Balmer, Y. Redox regulation: A broadening horizon. Ann. Rev. Plant Biol. 2005, 56, 187–220. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K.; Noguchi, K.; Motohash, K.; Hisabori, T. Systematic exploration of thioredoxin target proteins in plant mitochondria. Plant Cell Physiol. 2013, 54, 875–892. [Google Scholar] [CrossRef] [PubMed]
- Rouhier, N.; Cerveau, D.; Couturier, J.; Reichheld, J.P.; Rey, P. Involvement of thiol-based mechanisms in plant development. Biochim. Biophys. Acta. 2015, 1850, 1479–1496. [Google Scholar] [CrossRef] [PubMed]
- Sevilla, F.; Camejo, D.; Ortiz-Espín, A.; Calderón, A.; Lázaro, J.J.; Jiménez, A. The thioredoxin/peroxiredoxin/sulfiredoxin system: current overview on its redox function in plants and regulation by reactive oxygen and nitrogen species. J. Exp. Bot. 2015, 66, 2945–2955. [Google Scholar] [CrossRef] [PubMed]
- Shahpiri, A.; Svensson, B.; Finni, Ch. From Proteomics to Structural Studies of Cytosolic/Mitochondrial-Type Thioredoxin Systems in Barley Seeds. Mol. Plant 2009, 2, 378–389. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.H.; Cai, N.; Tanaka, C.K.; Vensel, W.H.; Hurkman, W.J.; Buchanan, B.B. Thioredoxin reduction alters the solubility of proteins of wheat starchy endosperm: An early event in cereal germination. Plant Cell Physiol. 2004, 45, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Gelhaye, E.; Rouhier, N.; Jacquot, J.P. The thioredoxin h system of higher plants. Plant Physiol. Biochem. 2004, 42, 265–271. [Google Scholar] [CrossRef]
- Balmer, Y.; Vensel, W.H.; Tanaka, C.K.; Hurkman, W.J.; Schurmann, P.; Droux, M.; et al. Thioredoxin links redox to the regulation of fundamental processes of plant mitochondria. PNAS 2004, 101, 2642–2647. [Google Scholar] [CrossRef]
- Sanz-Barrio, R.; Fernández-San Millán, A.; Carballeda, J.; Corral-Martínez, P.; Seguí-Simarro, J.M.; Farran, I. Chaperone-like properties of tobacco plastid thioredoxins f and m. J. Exp. Bot. 2012, 63, 365–379. [Google Scholar] [CrossRef]
- Hill, S.; Van Remmen, H. Mitochondrial stress signaling in longevity: A new role for mitochondrial function in aging. Redox Biol. 2014, 27, 936–944. [Google Scholar] [CrossRef] [PubMed]
- Sun, N.; Youle R., J.; Finkel, T. The mitochondrial basis of aging. Mol. Cell 2016, 61, 654–666. [Google Scholar] [CrossRef]
- Gredilla, R.; Bohr, V.A.; Stevnsner, T. Mitochondrial DNA repair and association with aging—An update. Exp. Gerontol. 2010, 45, 478–488. [Google Scholar] [CrossRef] [PubMed]
- Waterworth, W.M.; Bray, C.M.; West, C.E. The importance of safeguarding genome integrity in germination and seed longevity. J. Exp. Bot. 2015, 66, 3549–3558. [Google Scholar] [CrossRef] [PubMed]
- Paszkiewicz, G.; Gualberto, J.M.; Benamar, A.; Macherel, D.; Logana, D.C. Arabidopsis seed mitochondria are bioenergetically active immediately upon imbibition and specialize via biogenesis in preparation for autotrophic growth. Plant Cell. 2017, 29, 109–128. [Google Scholar] [CrossRef]
- Fu, B.F.; Ahmed, Z.; Diederichsen, A. Towards a better monitoring of seed aging under ex situ seed conservation. Conserv. Physiol. 2015, 3, cov026. [Google Scholar] [CrossRef] [PubMed]
- Madreiter-Sokolowski, C.T.; Waldeck-Weiermair, M.; Bourguignon, M.P.; Villeneuve, N.; Gottschalka, B.; Klec, Ch.; Stryeck, S.; Radulovic, S.; Parichatikan, W.; Madl, T.; et al. Enhanced inter-compartmental Ca2+ flux modulates mitochondrial metabolism and apoptotic threshold during aging. Redox. Biol. 2019, 20, 458–466. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Berkowitz, O.; Selinski, J.; Xu, Y.; Hartmann, A.; Whelan, J. Stress responsive mitochondrial proteins in Arabidopsis thaliana. Free Radic. Biol. Med. 2018, 122, 28–39. [Google Scholar] [CrossRef] [PubMed]
- Cogliati, S.; Enriquez, J.A.; Scorrano, L. Mitochondrial Cristae: Where beauty meets functionality. Trends Biochem. Sci. 2018, 41, 261–273. [Google Scholar] [CrossRef] [PubMed]
- Noctor, G.; Paepe, R.; Foyer, C.H. Mitochondrial redox biology and homeostasis in plants. Trends Plant Sci. 2007, 12, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Daum, B.; Walter, A.; Horst, A.; Osiewacz, H.D.; Kühlbrandt, W. Age-dependent dissociation of ATP synthase dimers and loss of inner-membrane cristae in mitochondria. PNAS 2013, 110, 15301–15306. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Dysfunctional mitochondria and aging of seeds. Due to continuous reactive oxygen species (ROS) production in mitochondria, the level of ROS increases during seed storage (1). The nitrogen oxide (NO) influence on alternative oxidase (AOX) (2), which cause ROS generation (2). Calcium ions change membrane potentials and are also influenced by AOX (2) on the increase of ROS level in mitochondria. The ascorbate-glutathione cycle (3) does not effectively remove ROS (3), which causes oxidative damage to mtDNA (4) and protein synthesis (5) and leads to changes in signaling and the redox status in the mitochondria (6). Increasing ROS levels (7) in the mitochondria cause oxidative damage to the membranes (7), which affects the inhibition of oxidative phosphorylation (8). All of these events cause a decrease in seed viability and show the basis of aspects of seed aging. Solid arrows shows the process that happened in ageing seeds, the doted arrows explain in which processes ROS participate, detailing processes of ageing and decrease of germination and viability of seed.
Figure 1.
Dysfunctional mitochondria and aging of seeds. Due to continuous reactive oxygen species (ROS) production in mitochondria, the level of ROS increases during seed storage (1). The nitrogen oxide (NO) influence on alternative oxidase (AOX) (2), which cause ROS generation (2). Calcium ions change membrane potentials and are also influenced by AOX (2) on the increase of ROS level in mitochondria. The ascorbate-glutathione cycle (3) does not effectively remove ROS (3), which causes oxidative damage to mtDNA (4) and protein synthesis (5) and leads to changes in signaling and the redox status in the mitochondria (6). Increasing ROS levels (7) in the mitochondria cause oxidative damage to the membranes (7), which affects the inhibition of oxidative phosphorylation (8). All of these events cause a decrease in seed viability and show the basis of aspects of seed aging. Solid arrows shows the process that happened in ageing seeds, the doted arrows explain in which processes ROS participate, detailing processes of ageing and decrease of germination and viability of seed.

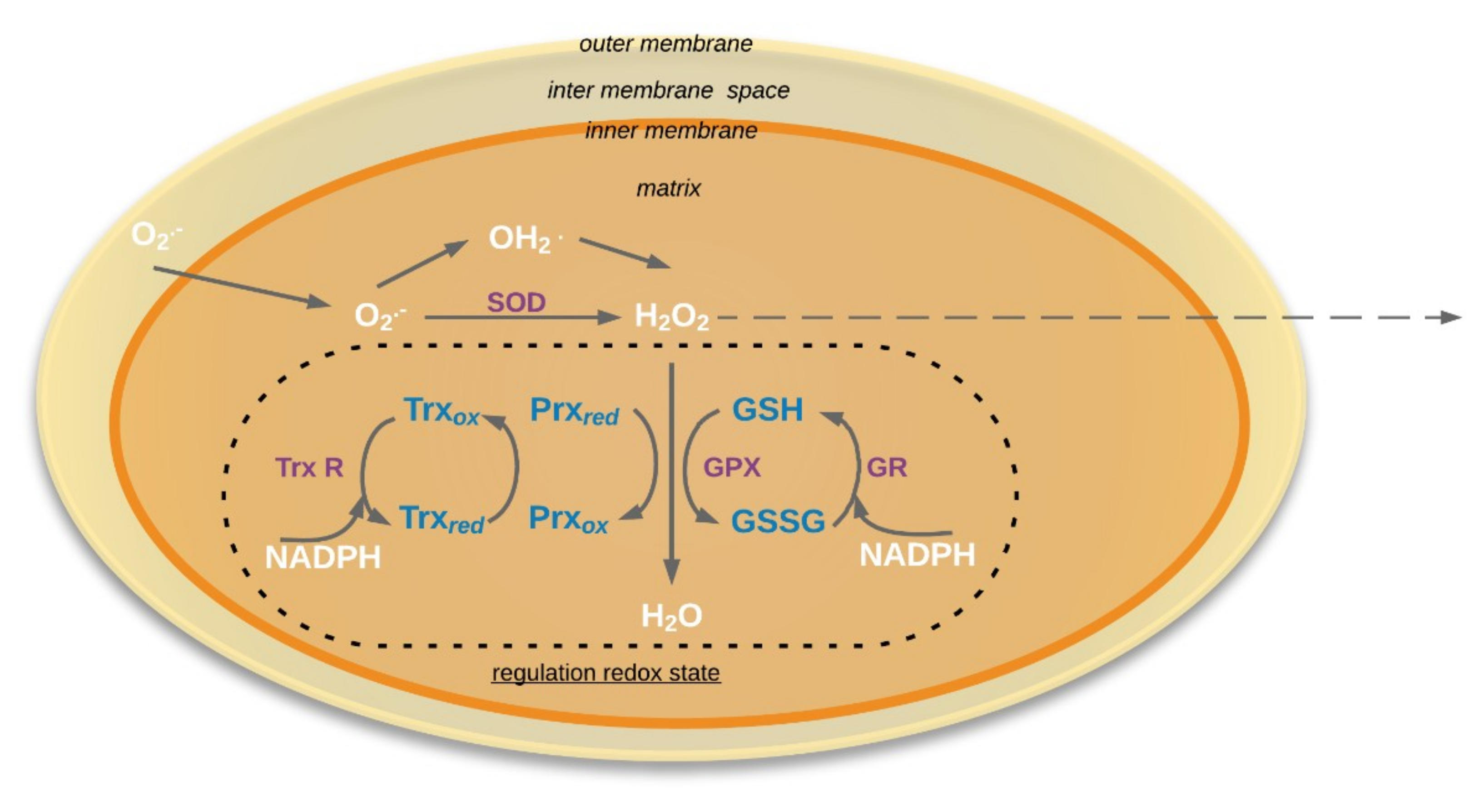
Figure 2.
Schema of regulation redox state in the mitochondria. O2•−: Superoxide; HO2˙: Hydroperoxyl radical; SOD: Superoxide dismutase (E.C. 1.15.1.1); GR: Glutathione reductase (EC 1.6.4.2); GSH: reduced form glutathione; GSSG: oxidized form glutathione; GPX: glutathione peroxidases (EC 1.11.1.9); Prx: peroxiredoxin; Trx: Thioredoxin; TrxR: NADPH-thioredoxin reductase (EC 1.8.1.9). Solid arrows mean that process occurred in mitochondria, dotted arrows mean that molecules could be exported outside the mitochondria.
Figure 2.
Schema of regulation redox state in the mitochondria. O2•−: Superoxide; HO2˙: Hydroperoxyl radical; SOD: Superoxide dismutase (E.C. 1.15.1.1); GR: Glutathione reductase (EC 1.6.4.2); GSH: reduced form glutathione; GSSG: oxidized form glutathione; GPX: glutathione peroxidases (EC 1.11.1.9); Prx: peroxiredoxin; Trx: Thioredoxin; TrxR: NADPH-thioredoxin reductase (EC 1.8.1.9). Solid arrows mean that process occurred in mitochondria, dotted arrows mean that molecules could be exported outside the mitochondria.


{kind=link}
{kind=link}
Table 1.
The role of reactive oxygen species in dysfunction mitochondrion seeds.
| Organism (Species) | Processes | References |
|---|---|---|
| Pea (Pisum sativum L.) | Germination after Aspergillus ruber infection, aging | [35] |
| Rye (Secale cereal L.) | Embryogenesis, germination | [36] |
| Soybean [Glycine max (L.) Merr] | Aging | [37] |
| Pea (Pisum sativum) | Germination | [25] |
| Soybean [Glycine max (L.) Merr] | Germination, imbibition | [38] |
| Pea (Pisum sativum cv. Jizhuang) | Germination | [39] |
| Soybean [G.max (L.) Merr] | Aging | [40] |
| Elm (Ulmus pumila L.) | Aging | [41] |
| Oat (Avena sativa L.) | Aging | [42] |
| Rice (Oryza sativa L.) | Aging | [7] |
| Rice (Oryza sativa L.) | Aging | [43] |
| Elm (Ulmus pumila L.) | Aging | [44] |
| Oat (Avena sativa L.) | Aging | [26] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ratajczak, E.; Małecka, A.; Ciereszko, I.; Staszak, A.M. Mitochondria Are Important Determinants of the Aging of Seeds. Int. J. Mol. Sci. 2019, 20, 1568. https://doi.org/10.3390/ijms20071568
AMA Style
Ratajczak E, Małecka A, Ciereszko I, Staszak AM. Mitochondria Are Important Determinants of the Aging of Seeds. International Journal of Molecular Sciences. 2019; 20(7):1568. https://doi.org/10.3390/ijms20071568
Chicago/Turabian StyleRatajczak, Ewelina, Arleta Małecka, Iwona Ciereszko, and Aleksandra M. Staszak. 2019. "Mitochondria Are Important Determinants of the Aging of Seeds" International Journal of Molecular Sciences 20, no. 7: 1568. https://doi.org/10.3390/ijms20071568
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.