1. Introduction
Species diversification is considered as a tool for sustainable development of aquaculture in near future [
1]. The Mediterranean yellowtail (
S. dumerili, Risso 1810) is one of the most interesting candidates for European aquaculture diversification [
2]. This carnivorous fish shows high growth rates (weighs 6 kg within 2.5 years of culture) and has excellent flesh quality and worldwide consumer acceptance [
2,
3].
Fish oil (FO) has been considered as the major lipid source in aquafeeds for carnivorous fish for a long time [
4,
5]. However, FO and fish meal production are no longer sustainable [
6,
7]. During the last couple of decades, the global supply of FO has been tightly regulated and has remained low and stable, whereas the aquaculture industry has expanded rapidly. Consequently, FO prices have increased, prompting researchers and industries to develop alternative lipid sources that can be included in aquafeeds [
8].
Vegetable oils (VO) (e.g., soybean oil, linseed oil, palm oil, and rapeseed oil) have been widely tested as alternatives to FO in aquafeeds, specifically for their impact on fish growth and flesh quality [
9,
10,
11]. However, VO are rich in C18 polyunsaturated fatty acids (PUFA), but are devoid of highly unsaturated fatty acids (HUFA), such as eicosapentaenoic acid (EPA; 20:5 n-3) and docosahexaenoic acid (DHA; 22:6 n-3), which are essentials for growth, health, reproduction, and body functions of fish [
5]. Most freshwater fish are capable of desaturating and elongating C18 fatty acids (i.e., linoleic acid C18:2 n-6 and α-linolenic acid C18:3 n-3) into EPA and DHA [
12], whereas marine carnivorous species have lost this bioconversion ability [
5,
13]. Therefore, these essential fatty acids (EFA) must be included in marine aquafeeds to meet their EFA requirement (approximately 4%) [
12]. According to the literature, the partial substitution (up to 60%) of FO with VO in the diets of marine fish during the grow-out phase does not affect their mortality and growth rates [
6,
10].
Nevertheless, a major limitation of replacing dietary FO with VO is its effect on fillet composition. Since the fatty acid (FA) profile of fish tissues reflects those of the diets consumed by the fish [
5,
14], fillets of fish fed diets containing high VO levels may contain a low amount of n-3 HUFA [
9,
15], which are considered beneficial for human health [
16,
17,
18,
19]. High intakes of EPA and DHA are recommended to prevent premature birth and low birth weight [
20], and also to reduce cardiovascular disease risks [
21,
22]. These FA are also anti-arrhythmic in nature and reduce platelet aggregation and blood triacyl glyceride levels [
23]. For these reasons, health organizations of several countries recommend a daily intake of 1.2-2.0 g/d of n-3 HUFA [
10], adjusted according to eating habits, age, and sex.
Therefore, to restore the healthy FA profile of fish fillets during fish farming, finishing strategies before slaughtering (i.e., wash-out period) have been proposed [
5,
15,
24,
25]. The efficacy of a finishing strategy including FO diets after a grow-out period supplemented with alternative lipid sources has been tested in marine fish species such as gilthead seabream [
6,
10], European seabass [
11,
13], red seabream [
26], and Senegalese sole [
27], as well as in freshwater species such as Murray cod [
25,
28], sunshine bass [
29], and rainbow trout [
30,
31], and in Atlantic salmon [
32,
33,
34,
35].
Finishing strategy can also differently affect the FA composition of the red and the white muscles that make up the fish fillets [
36]. The former (approx. 10% of the fillet) are located in strips along the midline and assure a steady aerobic swimming by an aerobic metabolism based on lipids [
37,
38]; the latter represent the bulk of the fillet [
38] and use carbohydrates for their energy metabolism [
38]. Additionally, white and red muscles show different sensorial traits and fatty acid composition [
39].
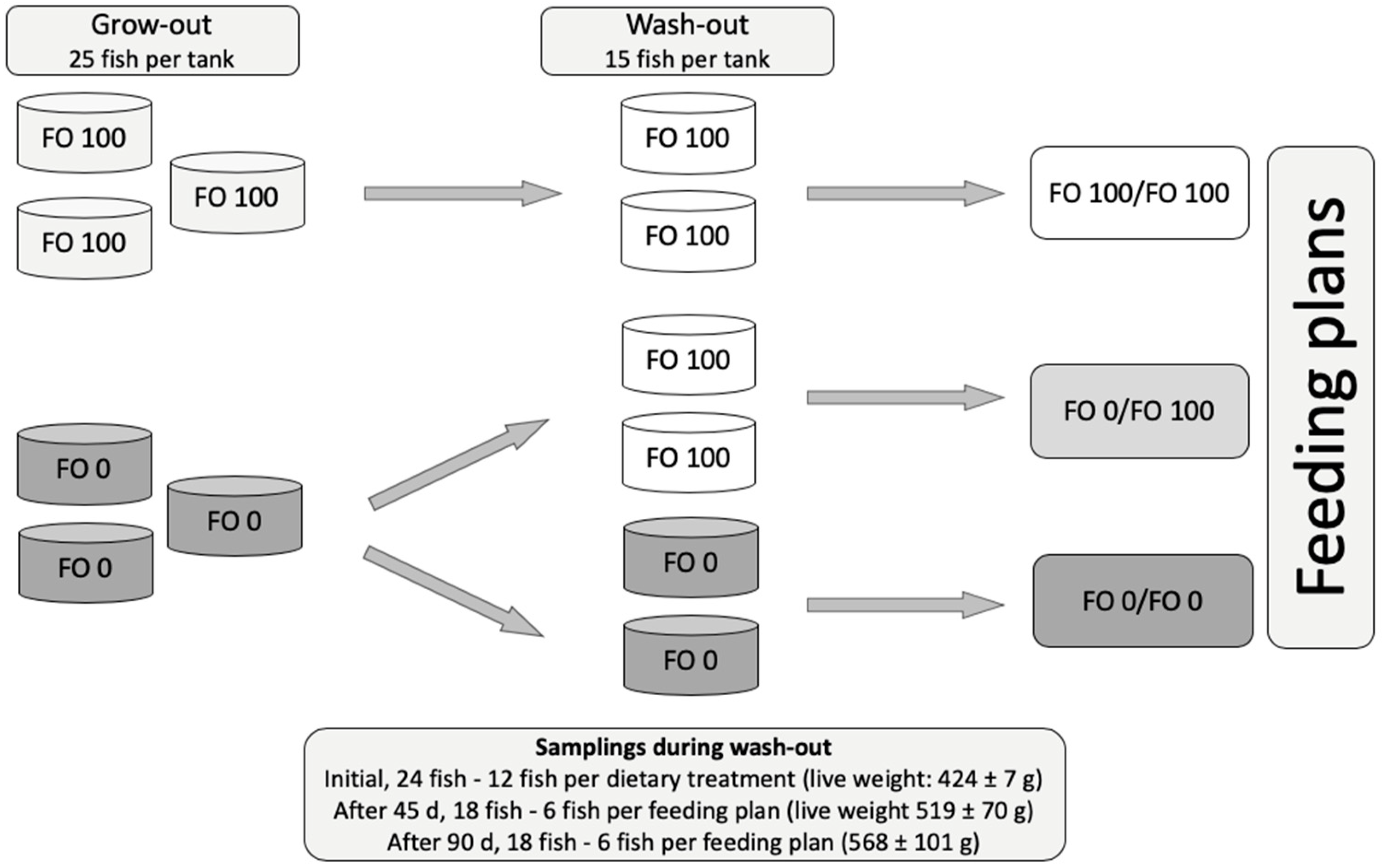
To date, there is no information about the effects of a finishing/wash-out strategy in Mediterranean yellowtail with special emphasis on the time required to restore the FA profile and the sensitivity of selected FA to a wash-out diet. Thus, the present study aimed to evaluate the effects of a wash-out diet and the time required to restore the FA composition of the white and red muscles in Mediterranean yellowtail that was previously fed a vegetable oil-based diet.
2. Results
The feeding plan did not affect fish growth (
Table 1). Fish reached an average weight of 490 g and 624 g after 45 and 90 d of wash-out, respectively, corresponding to SGR (specific growth rate) equal to 0.45% and 0.55% per day (
Table 1).
At the beginning of the wash-out period, the FA profile in the white muscles of the fish fed FO 0 diet showed lower levels of saturated FA (SFA) (−6.1%), arachidonic acid (AA; C20:4 n-6) (−38%), EPA (C20:5 n-3) (−34%), DHA (C22:6 n-3) (−44%) but showed higher ALA (C18:3 n-3) (+176%;
p < 0.001), and total n-6 (+11%) when compared to those seen in fish fed FO 100 diet. The higher total n-6 could be attributed to changes in the levels of LA (C18:2 n-6) (+16%) (
Table 2). Similarly, the red muscles of the fish fed FO 0 diet showed lower levels of AA (−41%), EPA (−35%), and DHA (−47%), and higher ALA (+238%;
p < 0.001) and total n-6 (+14%) (+21%), when compared to those seen in fish that were fed FO 100 diet (
Table 2). The increase in total n-6 (+14%) could be attributed to the changes in LA.
The dietary treatment did not affect the average levels of C16:0 (14.47%), C18:0 (6.29%,), and total n-3 (21.39%) in the white muscles after 45 days of wash-out (
Table 3). The FA profile of fish that were fed the finishing fish-oil diet (FO 0/FO 100) was similar to that of fish which were fed the VO diet exclusively. Fish that were previously fed FO 0 diet (groups FO 0/FO 0 and FO 0/FO 100) showed lower total SFA (−4%;
p < 0.01), AA (−36%), DHA (−42%), but higher LA (+19%), total n-6 (+12%), and ALA (+193%) (
p < 0.001) levels when compared with those seen in fish which were fed FO 100 diet exclusively (
Table 3). On the other hand, the FA profile of the white muscles was partially restored as EPA showed an increase (+13%) when fish from FO 0/FO 0 dietary treatment were compared with fish that were fed FO 0/100 diet. The EPA levels of fish from FO 0/FO 0 dietary treatment were lower (−33%) than those seen in fish which were fed FO 100 diet exclusively (
p < 0.001).
The dietary treatment did not affect the average levels of C16:0 (14.16%) and C18:0 proportions (6.78%) in the red muscles of the fish after 45 days of wash-out. Fish that were exclusively fed FO 0 diet and fish that were fed FO 0/FO 100 both showed lower total SFA (−4%) and DHA (−36%), but showed elevated proportions of LA (+18%) and total n-6 (+12%) (
p < 0.001) when compared with those seen in fish fed FO 100 diet exclusively (
Table 3). As seen in the white muscle, the FA profile of the red muscles was also partially restored as EPA showed an increase (+16%) when fish from FO 0/FO 0 dietary treatment were compared with fish that were fed FO 0/FO 100 diet. The EPA levels of fish from FO 0/FO 0 dietary treatment were lower (−21%) than those seen in fish which were always fed FO 100 diet (
p < 0.001). Moreover, ALA decreased (−25%) in fish fed FO 0 diet exclusively, when compared with those seen in fish fed FO 0/FO 100 diets (
p < 0.001) (
Table 3).
At the end of wash-out phase, the dietary treatment did not affect the average levels of C16:0 (14.58%), C18:0 (6.13%), and total n-3 (20.91%) in the white muscles of the fish (
Table 4). The differences recorded after 45 d of wash-out between fish that were exclusively fed FO 100 diet and those that were exclusively fed FO 0 diet or fed FO 0/FO 100 diets were confirmed for total SFA (+5% in FO 100;
p < 0.001) and total
n-6 (−6% in FO 100;
p < 0.001). On the other hand, a partial restoration of the FA profile was observed for 74% of fatty acids in fish that were previously fed the FO 0 diet, due to the wash-out phase with FO 100 diet. Fish subjected to wash-out treatment showed higher ratios of AA (+33%), EPA (+16%), and DHA (+43%) (
p < 0.001) and lower ratios of LA (−6%) and ALA (−29%) (
p < 0.001) when compared with those seen in the fish that were not subjected to wash-out treatment (
Table 4).
After 90 d of wash-out, the SFA and EPA ratios seen in the red muscles of fish that were exclusively fed FO 0 diet or in those which were subjected to wash-out treatment with FO 100 diet were lower than that seen in fish that were exclusively fed FO 100 diet [total SFA (−7%;
p < 0.001) and EPA (−20%;
p < 0.05)]. On the other hand, a partial restoration for 65% of fatty acids was observed in fish that were previously fed the FO 0 diet due to the wash-out treatment with FO 100 diet. Fish subjected to wash-out treatment showed higher ratios of AA (+33%) and DHA (+41%) (
p < 0.001), but lower ratios of LA (−7%), and ALA (−29%) (
p < 0.001) compared to fish exclusively fed with FO 0 diet (
Table 4).
As for the nutritional quality of lipids, the index of atherogenicity (IA) was higher in white muscles of fish that were exclusively fed FO 100 diet when compared with those seen in the other groups of fish at the first (+23%), 45th (+17%), and 90th day of wash-out (+16%) (
p < 0.001). No differences among treatments were recorded for the average value of the index of thrombogenicity (IT) at the first, 45th, and 90th day of wash-out treatment (0.23) (
Table 5).
In the red muscles of the fish, IA was higher in fish that were exclusively fed FO 100 diet as compared to that in the fish which were fed FO 0 diet, on the first (+12%;
p < 0.05) and on the 45th day of wash-out treatment (+16%;
p < 0.001). In fish that were fed FO 0/FO 100 diet, IA value on the 45th day of wash-out (0.25, on average) was equal to that seen in fish that were fed FO 0 diet exclusively, whereas it was higher (+8%) at the end of the treatment. As for the white muscles, no differences among treatments were recorded for the average value of IT at the first, 45th, and 90th day of wash-out (0.22) (
Table 5).
3. Discussion
In the last few decades, the widespread use of FO as lipid source in aquafeeds has drastically challenged the sustainability of aquaculture. The substitution of FO with alternative sources (such as vegetable oils) has been successful in terms of fish performance but has faced concerns about fish health and flesh nutritional quality. New feeding strategies are being developed to address those concerns. In fact, the replacement of FO with VO in aqua feed increases dietary oleic acid, LA and ALA and reduces the n-3 fatty acid such as EPA and DHA, which increases the vegetable lipid profile of the whole fish, as well as its organs and flesh [
5]. Since n-3 PUFA play a specific role in inflammatory processes and immune response [
40], the change in the dietary FA profile can affect fish response both during growth and wash-out. Moreover, the presence of vegetable FA leads to a decrease in the nutritional value of fish flesh for humans as a result of changes in FA profile as well as an unbalanced ratio of n-3/n-6 [
41]. This can affect both the white and red muscles of fish, whereas a greater contribution to the nutritional value of the flesh comes from the former due its higher proportion on the whole fillet compared to the latter (in the present trial, red muscles: 6.8–9.8% of total fillet weight).
In the present trial, two indices based on the functional effects of FA were used to evaluate the nutritional quality of the fish fillet lipid fraction, i.e., the indexes of atherogenicity (IA) and thrombogenicity (IT). Briefly, IA indicates the relationship between main classes of saturated FA (considered pro-atherogenic, i.e., favoring lipid adhesion to cells) and those of unsaturated FA (anti-atherogenic; i.e., inhibiting the aggregation of lipid plaque and reducing the levels of cholesterol, phospholipid and esterified FA) [
42]. The IT is defined by the relationship between the pro-thrombogenic saturated FA and the anti-thrombogenic FA (MUFA, PUFA n-6, and PUFA n-3) [
42]. The IT of flesh in our trial was consistent with values (0.22–0.23) found in most common marine species [
43], whereas the higher IA in fish fed FO 100 diet is related to the higher content of myristic acid (C14:0) in their red and white muscles. In fact, myristic acid in FO 100 diet was almost twice than in FO 0 one. Nevertheless, for all fish, IA values were within recommended values for human health (<1.0) [
44,
45].
Moreover, in the present trial, after the growth phase, the FA profile of
S. dumerili in the white and red muscles reflected the FA composition of the fish-oil (FO 100) or vegetable-oil (FO 0) diets, ingested by the fish, which agreed with our assumptions. Nevertheless, the differences in AA, EPA and DHA rates in both the white and red muscles of the fish that were fed the two dietary treatments were lower than the difference in the diets, which agrees with previously published studies on gilthead seabream [
6,
10,
46] and rainbow trout [
31].
To restore the fillet nutritional value (in terms of high levels of EPA and DHA) in fish grown on diets containing vegetable oils, specific feeding strategies can be used during the finishing period to wash-out fish that were previously fed VO diets [
4,
15,
31]. However, fish FA levels are also affected by the fish biosynthesis ability for the different FA, besides the dietary supply. Indeed, standards for a successful wash-out and recovery of the desired FA in fish flesh are not yet available for all species including greater amberjack. On the other hand, the available literature about other species is inconsistent.
In gilthead seabream, oleic acid and LA are retained in the flesh of the fish even after 120 d of wash-out [
6]. In European sea bass, Montero et al. [
11] reported that LA was 3-fold higher in fish previously fed vegetable-oil diets compared to those that were fed on fish oil exclusively even after 150 d of wash-out, which agrees with the results stated by Izquierdo et al. [
10] for seabass after a 104-d wash-out. In turbot, even after a wash-out of eight months, high levels of LA in muscle phospholipids have been reported, which could be attributed to the poor LA utilization in a species that can convert LA to 20:2 n-6 [
47].
According to Mourente and Bell [
13] the wash-out (150 d, 160 g LW) treatment in sea bass is insufficient for restoring EFA. On the contrary, Montero et al. [
11] report that a wash-out period of 150 d, after 8 months of a diet containing 60:40 ratio of vegetable oil and fish oil (75–366 g LW), was able to recover flesh DHA, but could not increase EPA level. Similarly, in gilthead seabream, EFA levels could not be restored after a 120-d wash-out [
6]. On the other hand, Izquierdo et al. [
10] report that DHA and AA levels were recovered in gilthead seabream following a wash-out of 60 d (after 7 months of feeding with diets containing vegetable oils at 60% and 80%; 85–452 g LW), but EPA levels were not recovered even after 90 d of wash-out treatment.
In rainbow trout, 8–12 weeks of wash-out cannot recover EPA and DHA levels [
30,
31], whereas in Atlantic salmon that has been previously fed linseed oil-based diets for 40 weeks, levels of EPA and DHA in flesh can be restored by 80% after 20 weeks [
34] and by 83% after 24 weeks of wash-out treatment [
32]. On the other hand, Bell et al. [
33] report that in post-smolt salmon (200 g) previously fed a diet containing increasing rates of rapeseed oil for 16 weeks, EPA and DHA levels in flesh can be recovered after 4 and 12 weeks of wash-out, respectively. In Senegalese sole a total restoration of all EFA levels in flesh can be achieved after 26 d of wash-out [
27].
In the present study, EFA levels in the Mediterranean yellowtail did not recover completely after a 90-d wash-out, but some differences between white and red muscles were recorded. In the white muscles, AA, EPA, and DHA levels were partially restored, despite remaining lower when compared with those of fish fed fish-oil diets exclusively. On the other hand, in the red muscle, a partial restoration was observed only for AA and DHA levels, whereas the EPA level remained low.
Based on literature, EFA recovery in fish muscles depends upon several factor such as fish species, fish size, duration for which fish were fed vegetable-oil based diets, duration of the wash-out period, and the specific FA. Additionally, FA incorporation in fish muscles can be altered by different metabolic factors such as FA elongation and desaturation, β-oxidation [
48], preferential incorporation [
49], lipogenic activity, environmental factors [
50], size and age of animals [
51], and their physiological state [
52].
In the present trial, after 90 d of wash-out, DHA was partially recovered and selectively retained by the muscles. This observation agrees with previously published studies on a wide variety of species [
53]. The mechanism of selective deposition has been likely influenced by the high specificity of fatty acid transferases for DHA and the relative resistance of DHA to β-oxidation [
54]. Fish size and fish physiological state can also play a role in DHA recovery. DHA content in flesh of the fish has been found to be negatively affected by the increase in fish size [
55] and the competition between muscles and developing gonads for its incorporation at the time of sexual maturation [
52], which was not the case of our trial.
The recovery of EPA in the flesh of the fish following a wash-out strategy was unsuccessful in the present trial. Our observations agree with previously reported studies done with other species (seabass, [
11]; seabream, [
10]; rainbow trout, [
30,
31]). According to Madsen et al. [
56], a preferential oxidation of EPA occurs over DHA; and EPA is mainly oxidized by mitochondria, whereas DHA seems to be oxidized by the peroxisomes. Thus, the failure of EPA recovery in the white muscle of Mediterranean yellowtail in our trial could be attributed to the fact that mitochondrial β-oxidation prevails over peroxisomal oxidation in white muscles [
56].
The definition of the wash-out duration is also crucial to produce fish with a healthy FA profile. An n-3 HUFA deficiency in the fish muscle lowers the nutritional value of the fish for humans. To overcome this, a dilution model has been proposed [
15] to predict the FA restoration at a given time after a dietary change. This model has been used in some fish species such as gilthead seabream and Atlantic salmon [
15,
57]. Nevertheless, the model does not fully represent changes in all FA and the variation in different species. For instance, in the Murray cod, the mobilization of oleic acid, LA, and ALA during the wash-out has been found to be at a lower rate than the rate predicted by the model, with major changes occurring during the first days of wash-out [
25]. According to some studies [
15,
29], the model can provide misleading results when it is used to study the FA changes of ‘lean’ fish fillets, such as the Mediterranean yellowtail.
Thus, under the conditions of our trial, a wash-out period of 90 d partially improved the final FA profile in muscles of Mediterranean yellowtail that was previously fed VO-based diets. In fact, based on our results, the quantity of muscle necessary to cover the daily-recommended ingestion of EPA and DHA (average requirement of 1.6 g/d of n-3 HUFA) decreased from 90 g/d (fish submitted to feeding plan FO 0/FO 0) to 70 g/d (fish submitted to feeding plan FO 0/FO 100). However, further studies are necessary to define the time and the dietary fish oil level that can restore EFA to the same levels seen in the fish that were exclusively fed on fish oil for S. dumerili, under different conditions of growth and at different sizes.


 ,
, 

{kind=link}