Phenolic Metabolites from Barley in Contribution to Phenome in soil Moisture Deficit

 , , , and
, , , and
Abstract
:
1. Introduction
2. Results
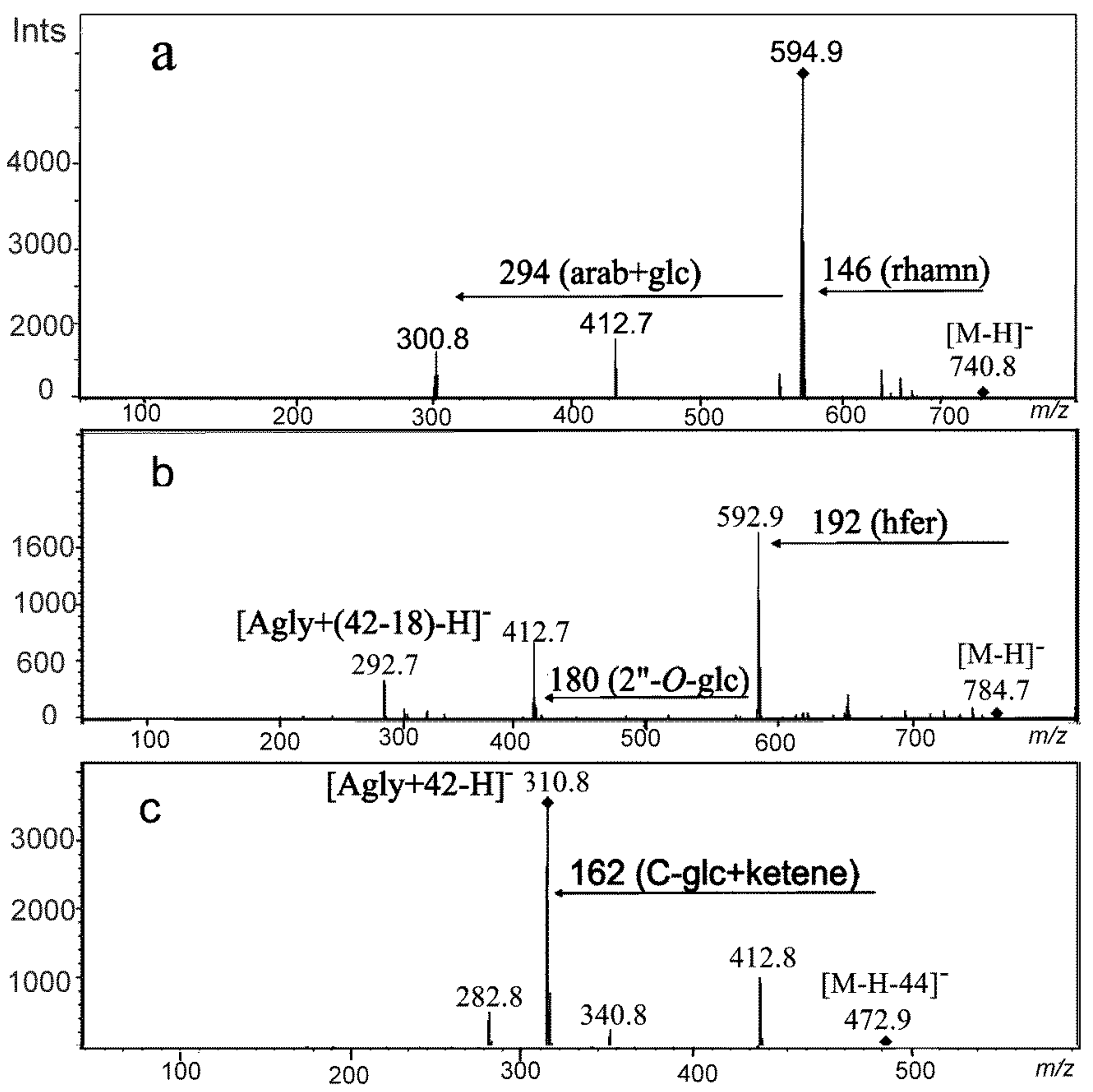
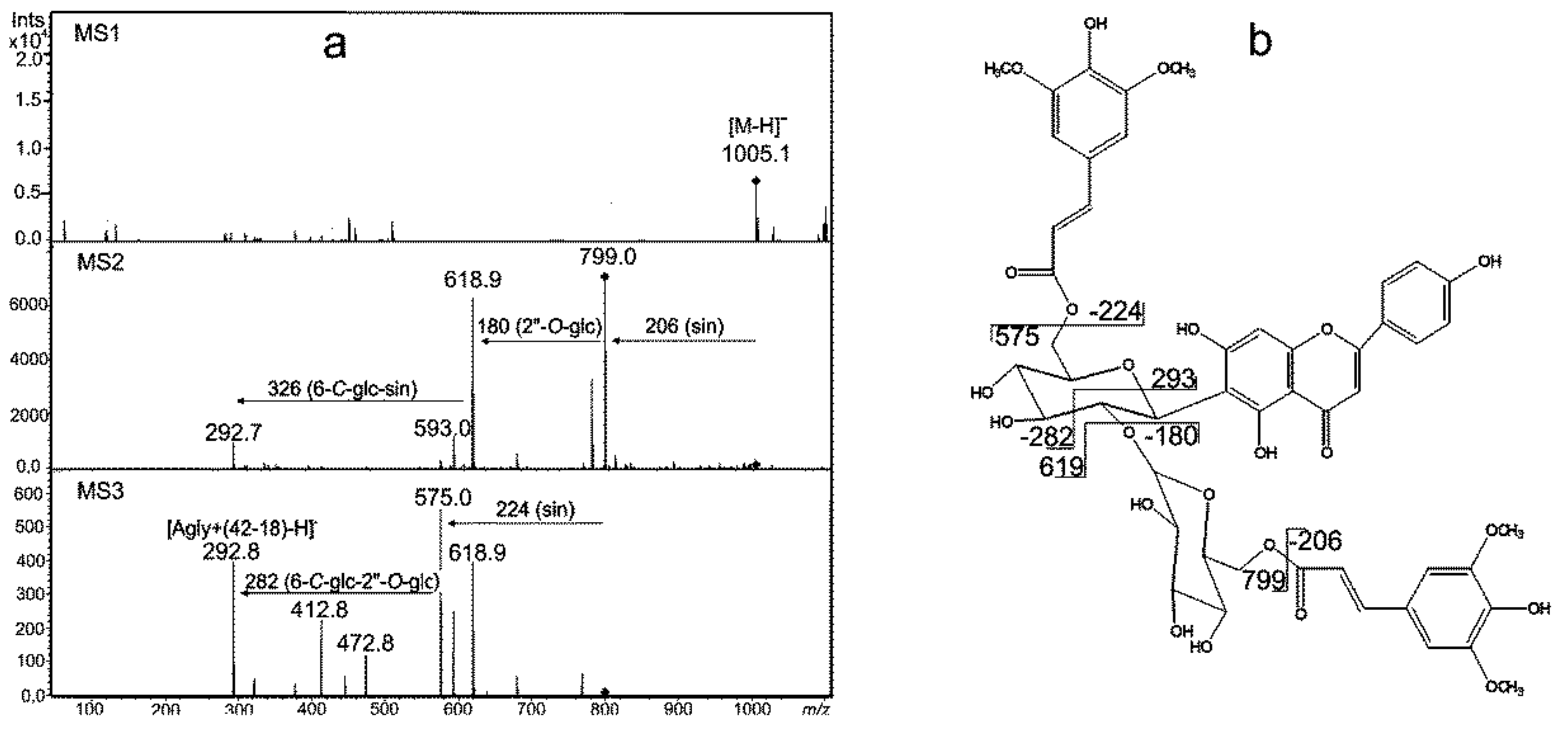
2.1. Peak Annotation
2.2. Changes in Phenolome in Moisture Deficit
2.2.1. Comparison of Effects in Three Treatments
- Observed effect of moisture deficit dependent on treatment (group D),
- Observed effect of moisture deficit dependent on the variety and treatment (VD)
- Observed effect of moisture deficit dependent on treatment and time of observation (treatment duration, TD)
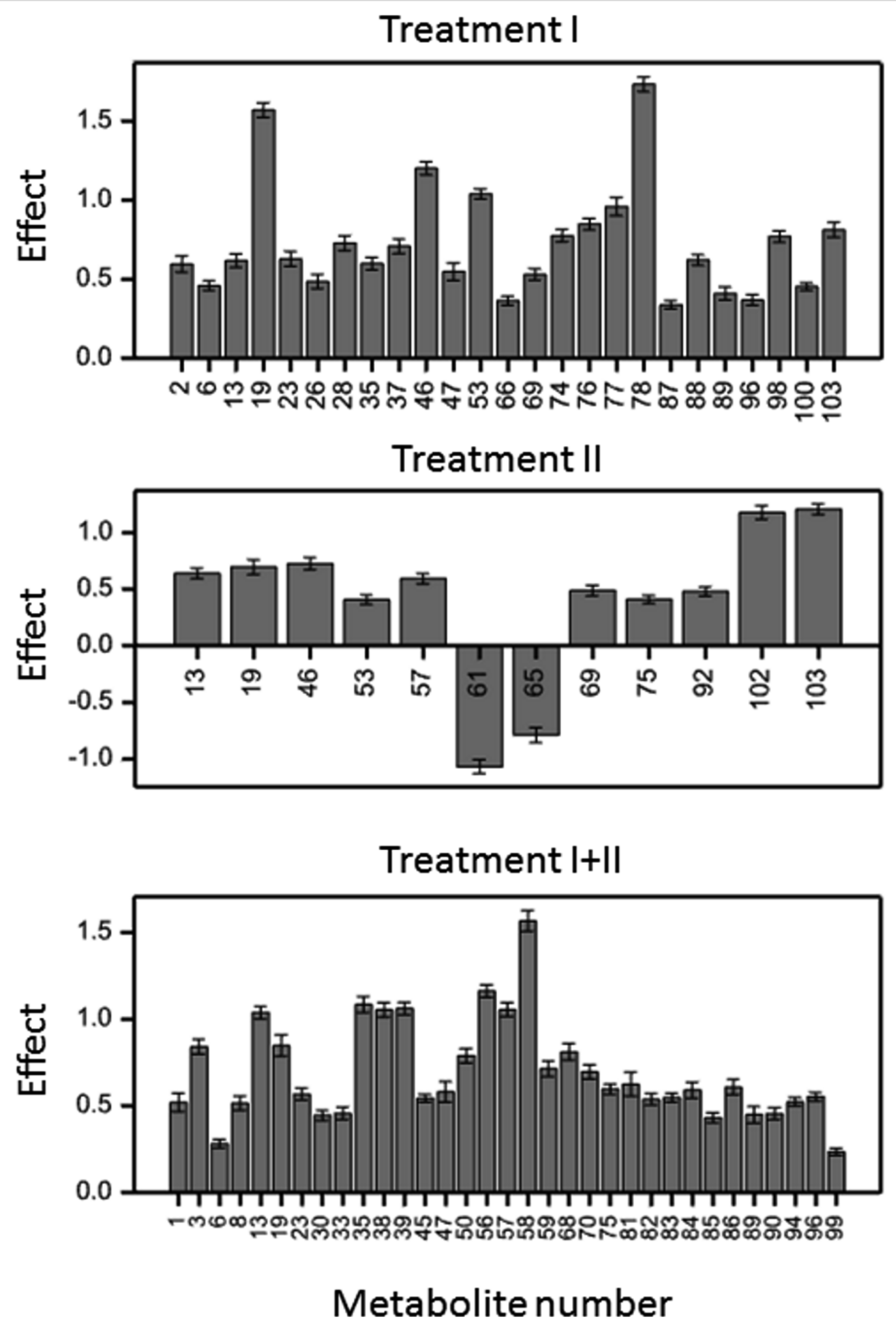
2.2.2. Metabolites in Group D (Treatment Effect Similar for All Variety × Time Variants)
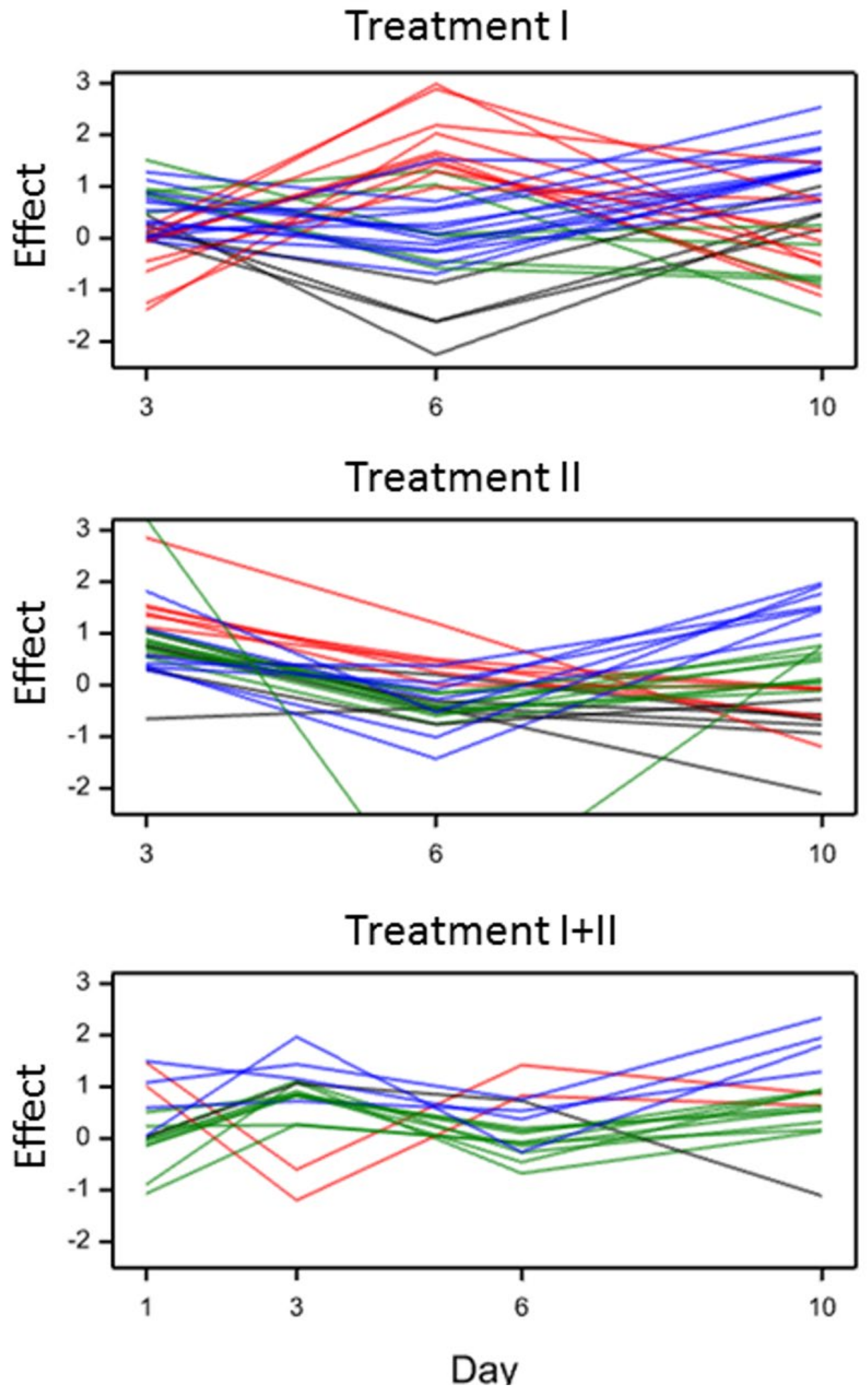
2.2.3. Metabolites in Group TD (Treatment Effect Modified by Time)
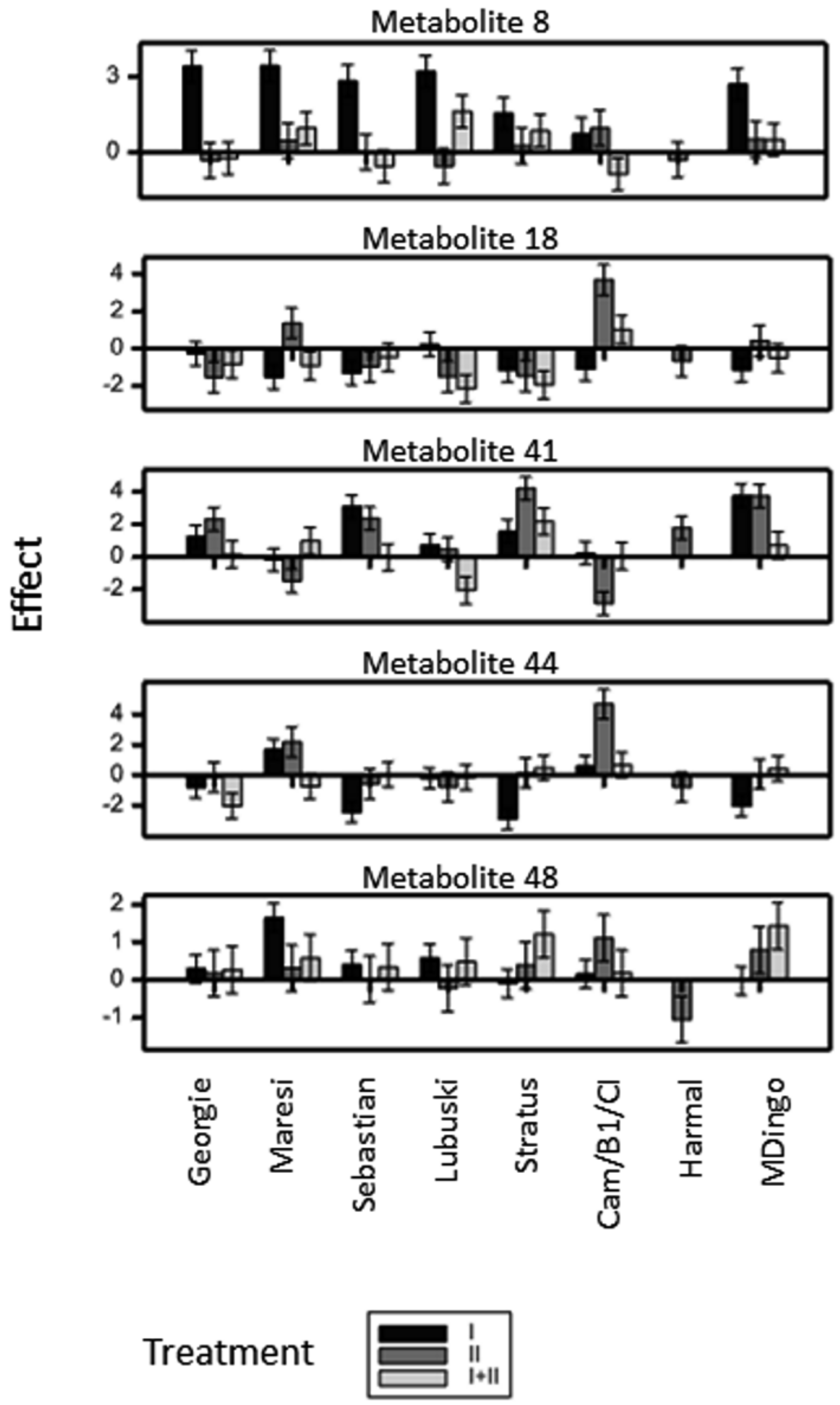
2.2.4. Metabolites in Group VTD (Treatment Effect Modified by Interaction Between Variety and Time)
2.3. Identification of Metabolites Induced in Moisture Deficit
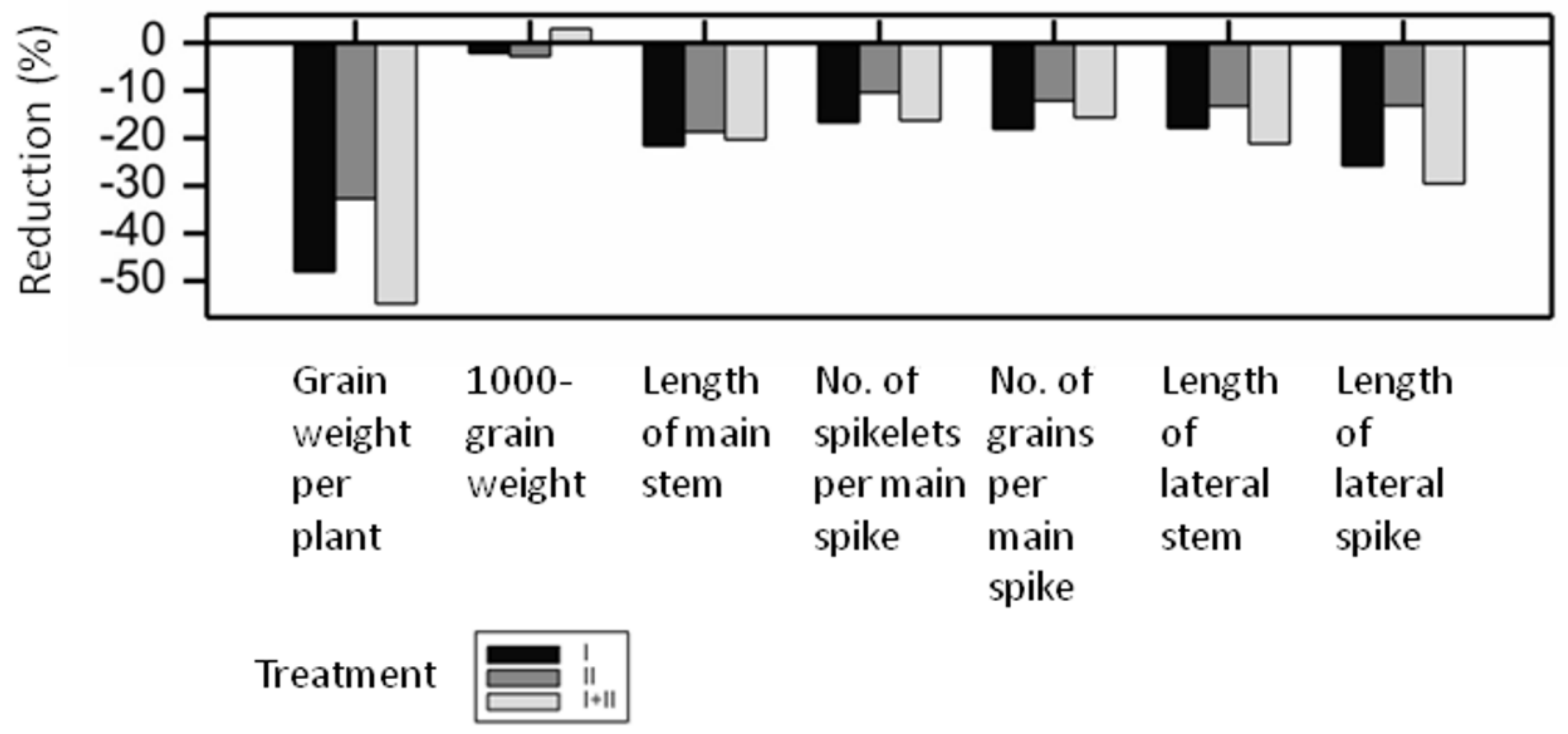
2.4. Moisture Deficit and Phenotypic Assessment at Maturity
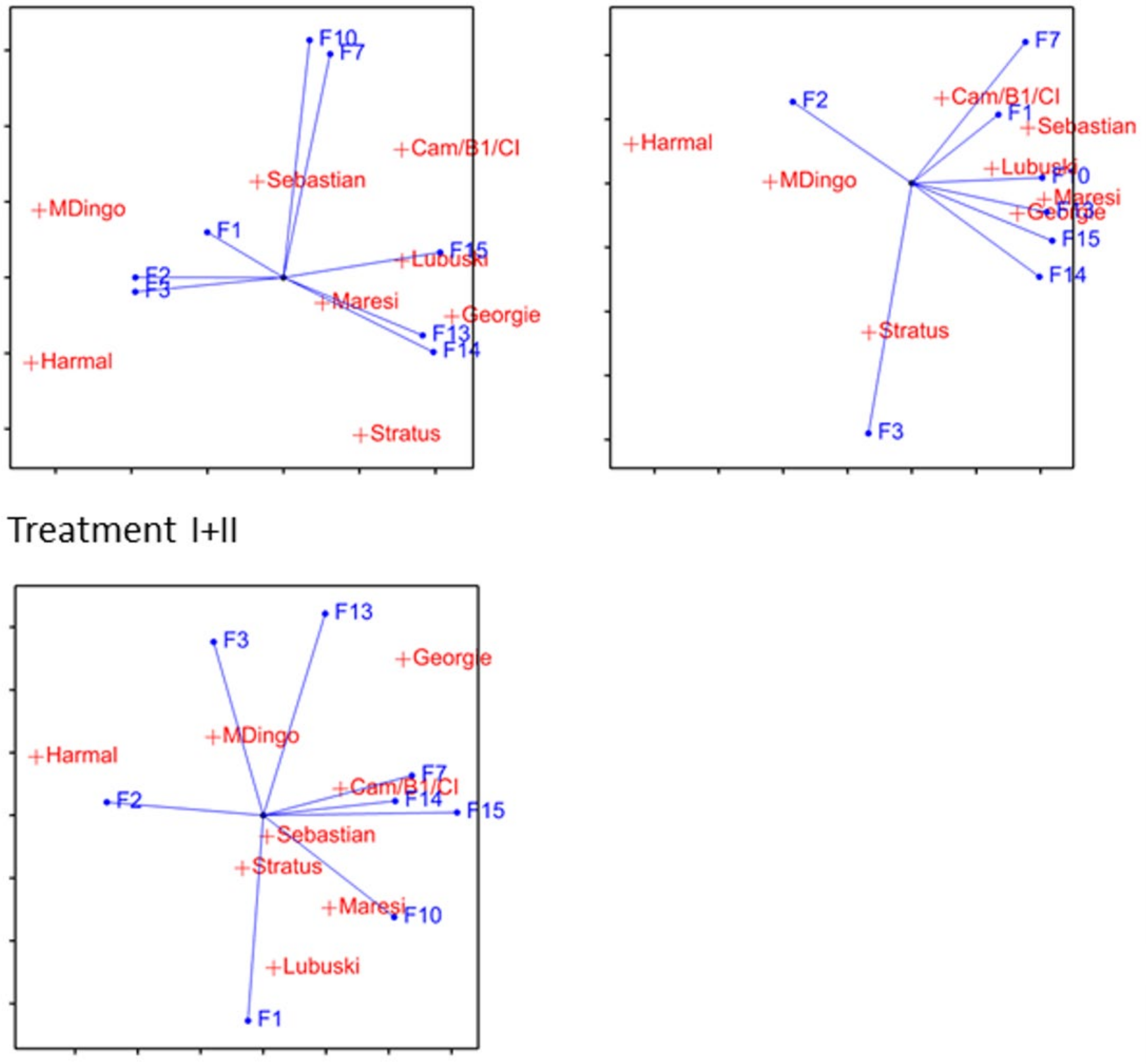
2.5. Correlation between Phenotypic and Metabolomic Traits in Moisture Deficit
3. Discussion
3.1. Design and Data Analysis
3.2. Phenolic Metabolites in Moisture Deficit
3.3. Phenotype and Moisture Deficit
3.4. Correlation between Phenotype and Metabolome in Moisture Deficit
4. Materials and Methods
4.1. Plant Material Cultivation
4.2. Phenolic Compounds Extraction from Plant Material
4.3. Phenolic Compounds Profiling with UPLC-PDA
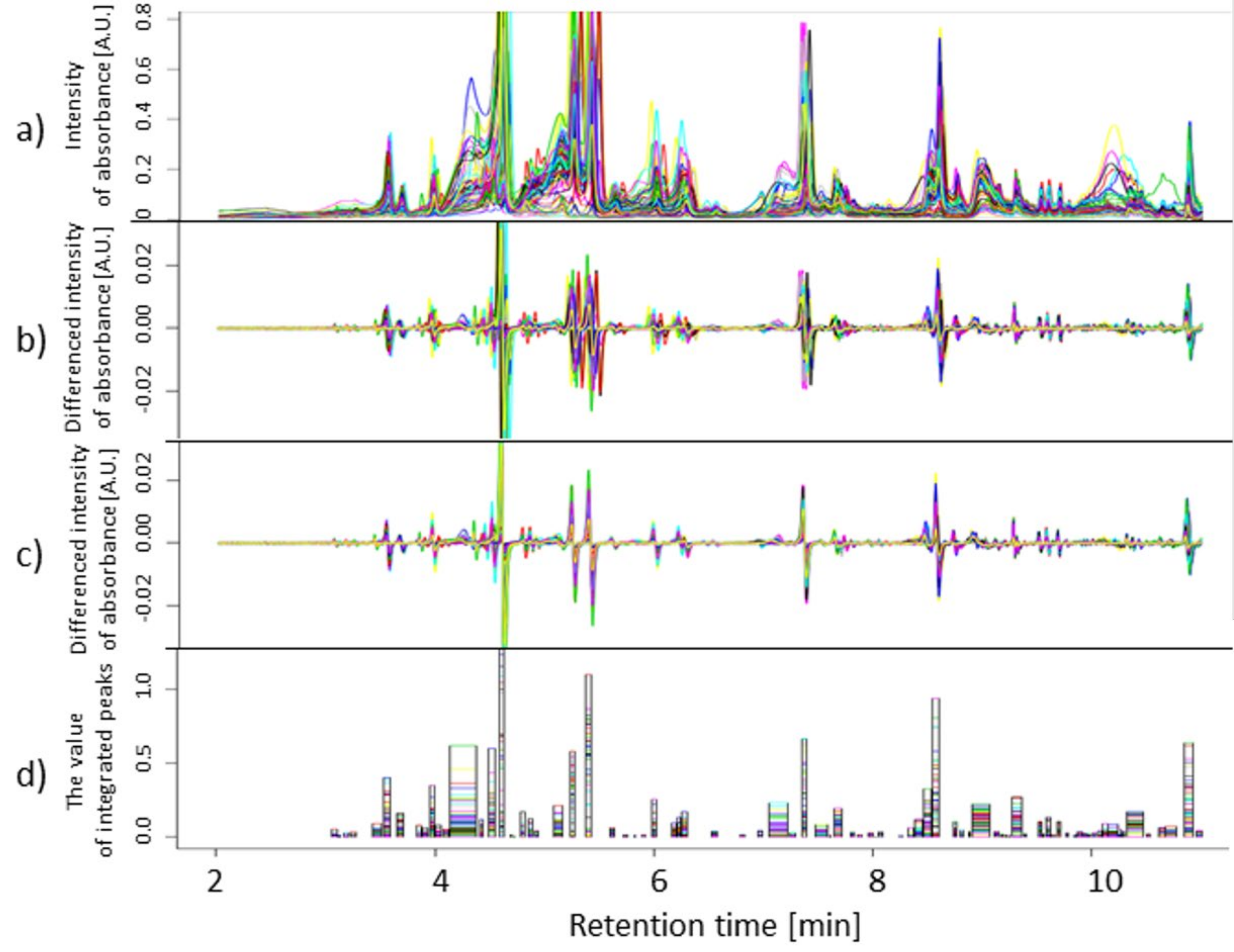
4.4. Chromatographic Data Pre-Processing
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Aroca, R. Plant. Responses to Drought Stress from Morphological to Molecular Features; Springer: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Szira, F.; Bálint, A.F.; Börner, A.; Galiba, G. Evaluation of drought-related traits and screening methods at different developmental stages in spring barley. J. Agron. Crop. Sci. 2008, 194, 334–342. [Google Scholar] [CrossRef]
- Wiegmann, M.; Maurer, A.; Pham, A.; March, T.J.; Al-Abdallat, A.; Thomas, W.T.B.; Bull, H.J.; Shahid, M.; Eglinton, J.; Baum, M.; et al. Barley yield formation under abiotic stress depends on the interplay between flowering time genes and environmental cues. Sci. Rep. 2019, 9, 6397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sallam, A.; Alqudah, A.M.; Dawood, M.F.A.; Baenziger, P.S.; Börner, A. Drought stress tolerance in wheat and barley: Advances in physiology, breeding and genetics research. Int. J. Mol. Sci. 2019, 20, 3137. [Google Scholar] [CrossRef] [Green Version]
- Bowler, K. Acclimation, heat shock and hardening. J. Therm. Biol. 2005, 30, 125–130. [Google Scholar] [CrossRef]
- Laxa, M.; Liebthal, M.; Telman, W.; Chibani, K.; Dietz, K.J. The role of the plant antioxidant system in drought tolerance. Antioxidants 2019, 8, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rapacz, M.; Kościelniak, J.; Jurczyk, B.; Adamska, A.; Wójcik, M. Different patterns of physiological and molecular response to drought in seedlings of malt- and feed-type barleys (Hordeum vulgare). J. Agron. Crop. Sci. 2010, 196, 9–19. [Google Scholar] [CrossRef]
- Croft, K.D. The chemistry and biological effects of flavonoids and phenolic acids. Ann. N. Y. Acad. Sci. 1998, 854, 435–442. [Google Scholar] [CrossRef] [PubMed]
- Tohge, T.; Ramos, M.S.; Nunes-Nesi, A.; Mutwil, M.; Giavalisco, P.; Steinhauser, D.; Schellenberg, M.; Willmitzer, L.; Persson, S.; Martinoia, E.; et al. Toward the storage metabolome: Profiling the barley vacuole. Plant Physiol. 2011, 157, 1469–1482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arora, A.; Nair, M.G.; Strasburg, G.M. Structure activity relationship for antioxidant activities of series of flavonoids in liposomal system. Free Rad. Biol. Med. 1998, 24, 1355–1363. [Google Scholar] [CrossRef]
- Kanakis, D.; Nafisi, S.; Rajabi, M.; Shadaloi, A.; Tarantilis, P.A.; Polissiou, M.G.; Bariyanga, J.; Tajmir-Riahi, H.A. Structural analysis of DNA and RNA interactions with antioxidant flavonoids. Spectroscopy. 2009, 23, 29–43. [Google Scholar] [CrossRef]
- Yamasaki, H.; Sakihama, Y.; lkehara, N. Flavonoid-peroxidase reaction as a detoxification mechanism of plant cells against H2O2. Plant Physiol. 1997, 11, 1405–1412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hura, K.; Ostrowska, A.; Dziurka, K.; Hura, T. Photosynthetic apparatus activity in relation to high and low contents of cell wall-bound phenolics in triticale under drought stress. Photosynthetica. 2017, 55, 698–704. [Google Scholar] [CrossRef]
- Hernández, I.; Van Breusegem, F. Opinion on the possible role of flavonoids as energy escape valves: Novel tools for nature’s Swiss army knife? Plant Sci. 2010, 179, 297–301. [Google Scholar] [CrossRef]
- Neilson, E.H.; Goodger, J.Q.D.; Woodrow, I.E.; Lindberg Møller, B. Plant chemical defense: At what cost? Trends Plant Sci. 2013, 18(5), 250–258. [Google Scholar] [CrossRef]
- De Mezer, M.; Turska-Taraska, A.; Kaczmarek, Z.; Glowacka, K.; Swarcewicz, B.; Rorat, T. Differential physiological and molecular response of barley genotypes to water deficit. Plant Physiol. Biochem. 2014, 80, 234–248. [Google Scholar] [CrossRef]
- Marok, M.A.; Tarrago, L.; Ksas, B.; Henri, P.; Abrous-Belbachir, O.; Havaux, M.; Rey, P. A drought-sensitive barley variety displays oxidative stress and strongly increased contents in low-molecular weight antioxidant compounds during water deficit compared to a tolerant variety. J. Plant Physiol. 2013, 170, 633–645. [Google Scholar] [CrossRef] [PubMed]
- Surma, M.; Kuczyńska, A.; Mikołajczak, K.; Ogrodowicz, P.; Adamski, T.; Ćwiek-Kupczyńska, H.; Sawikowska, A.; Pecio, A.; Wach, D.; Józefaciuk, G.; et al. Barley varieties in semi-controlled and natural conditions—Response to water shortage and changing environment. J. Agron. Crop. Sci. 2019, 205, 295–308. [Google Scholar] [CrossRef]
- Mikołajczak, K.; Kuczyńska, A.; Krajewski, P.; Sawikowska, A.; Surma, M.; Ogrodowicz, P.; Adamski, T.; Krystkowiak, K.; Górny, A.G.; Kempa, M.; et al. Quantitative trait loci for plant height in Maresi × CamB barley population and their associations with yield-related traits under different water regimes. J. Appl. Genet. 2017, 58, 23–35. [Google Scholar] [CrossRef] [Green Version]
- Nagy, Z.; Németh, E.; Guóth, A.; Bona, L.; Wodala, B.; Pécsváradi, A. Metabolic indicators of drought stress tolerance in wheat: Glutamine synthetase isoenzymes and Rubisco. Plant Physiol. Biochem. 2013, 67, 48–54. [Google Scholar] [CrossRef]
- Piasecka, A.; Sawikowska, A.; Kuczyńska, A.; Ogrodowicz, P.; Mikołajczak, K.; Krystkowiak, K.; Gudyś, K.; Guzy-Wróbelska, J.; Krajewski, P.; Kachlicki, P. Drought-related secondary metabolites of barley (Hordeum vulgare L.) leaves and their metabolomic quantitative trait loci. Plant J. 2017, 89, 898–913. [Google Scholar] [CrossRef] [Green Version]
- Piasecka, A.; Sawikowska, A.; Krajewski, P.; Kachlicki, P. Combined mass spectrometric and chromatographic methods for in-depth analysis of phenolic secondary metabolites in barley leaves. J. Mass Spectrom. 2015, 50, 513–532. [Google Scholar] [CrossRef] [PubMed]
- Ferreres, F.; Andrade, P.B.; Valentao, P.; Gil-Izquierdo, A. Further knowledge on barley (Hordeum vulgare L.) leaves O-glycosyl-C-glycosyl flavones by liquid chromatography-UV diode–array detection-electrospray ionisation mass spectrometry. J. Chromatogr. A 2008, 1182, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Wojakowska, A.; Piasecka, A.; García-López, P.M.; Zamora-Natera, F.; Krajewski, P.; Marczak, Ł.; Kachlicki, P.; Stobiecki, M. Structural analysis and profiling of phenolic secondary metabolites of Mexican lupine species using LC-MS techniques. Phytochemistry 2013, 92, 71–86. [Google Scholar] [CrossRef]
- Oosterhuis, D.M.; Cartwright, P.M. Spike differentiation and floret survival in semidwarf spring wheat as affected by water stress and photoperiod. Crop. Sci. 1983, 23, 711–717. [Google Scholar] [CrossRef]
- Lafitte, H.R.; Yongsheng, G.; Yan, S.; Li, Z.H. Whole plant responses, key processes, and adaptation to drought stress: The case of rice. J. Exp. Bot. 2007, 58, 169–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomasi, G.; Savorani, F.; Engelsen, S.B. Icoshift: An effective tool for the alignment of chromatographic data. J. Chromatogr. A 2011, 1218, 7832–7840. [Google Scholar] [CrossRef]
- Zhang, Z.M.; Liang, Y.Z.; Lu, H.M.; Tan, B.B.; Xu, X.N.; Ferro, M. Multiscale peak alignment for chromatographic datasets. J. Chromatogr. A 2012, 1223, 93–106. [Google Scholar] [CrossRef]
- Piasecka, A.; Kachlicki, P.; Stobiecki, M. Analytical methods for detection of plant metabolomes changes in response to biotic and abiotic stresses. Int. J. Mol. Sci. 2019, 20, 379. [Google Scholar] [CrossRef] [Green Version]
- Nakabayashi, R.; Yonekura-Sakakibara, K.; Urano, K.; Suzuki, M.; Yamada, Y.; Nishizawa, T.; Matsuda, F.; Kojima, M.; Sakakibara, H.; Shinozaki, K.; et al. Enhancement of oxidative and drought tolerance in Arabidopsis by overaccumulation of antioxidant flavonoids. Plant J. 2014, 77, 367–379. [Google Scholar] [CrossRef]
- Brauch, D.; Porzel, A.; Schumann, E.; Pillen, K.; Mock, H.P. Changes in isovitexin-O-glycosylation during the development of young barley plants. Phytochemistry 2018, 148, 11–20. [Google Scholar] [CrossRef]
- Ishihara, A.; Kumeda, R.; Hayashi, N.; Yagi, Y.; Sakaguchi, N.; Kokubo, Y.; Ube, N.; Tebayashi, S.; Ueno, K. Induced accumulation of tyramine, serotonin, and related amines in response to Bipolaris sorokiniana infection in barley. Biosci. Biotechnol. Biochem. 2017, 81, 1090–1098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoessl, A.; Unwin, C.H. The antifungal factors in barley. V. Antifungal activity of the hordatines. Can. J. Bot. 1970, 48, 465–470. [Google Scholar] [CrossRef]
- Commont, D.; Winters, A.; Gwynn-Jones, D. Acclimation and interaction between drought and elevated UV-B in A. thaliana: Differences in response over treatment, recovery and reproduction. Ecol. Evol. 2012, 2, 2695–2709. [Google Scholar] [CrossRef]
- Marraccini, P.; Vinecky, F.; Alves, G.S.C.; Ramos, H.J.O.; Elbelt, S.; Vieira, N.G.; Carneiro, F.A.; Sujii, P.S.; Alekcevetch, J.C.; Silva, V.A.; et al. Differentially expressed genes and proteins upon drought acclimation in tolerant and sensitive genotypes of Coffea canephora. J. Exp. Bot. 2012, 63, 4191–4212. [Google Scholar] [CrossRef] [PubMed]
- Watkinson, J.I.; Sioson, A.A.; Vasquez-Robinet, C.; Shukla, M.; Kumar, D.; Ellis, M.; Heath, L.S.; Ramakrishnan, N.; Chevone, B.; Watson, L.T.; et al. Photosynthetic acclimation is reflected in specific patterns of gene expression in drought-stressed loblolly pine. Plant Physiol. 2003, 133, 1702–1716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agati, G.; Matteini, P.; Goti, A.; Tattini, M. Chloroplast-located flavonoids can scavenge singlet oxygen. New Phytol. 2007, 174, 77–89. [Google Scholar] [CrossRef]
- Stochmal, A.; Oleszek, W. Effect of acylation of flavones with hydroxycinnamic acids on their spectral characteristics. Nat. Prod. Commun. 2007, 2, 571–574. [Google Scholar] [CrossRef] [Green Version]
- Kohyama, N.; Ono, H. Hordatine A ß-D-glucopyranoside from ungerminated barley grains. J. Agric. Food Chem. 2013, 61, 1112–1116. [Google Scholar] [CrossRef]
- Alcázar, R.; Altabella, T.; Marco, F.; Bortolotti, C.; Reymond, M.; Koncz, C.; Carrasco, P.; Tiburcio, A.F. Polyamines: Molecules with regulatory functions in plant abiotic stress tolerance. Planta 2010, 231, 1237–1249. [Google Scholar] [CrossRef]
- Sarabia, L.D.; Boughton, B.A.; Rupasinghe, T.; Van De Meene, A.M.L.; Callahan, D.L.; Hill, C.B.; Roessner, U. High-mass-resolution MALDI mass spectrometry imaging reveals detailed spatial distribution of metabolites and lipids in roots of barley seedlings in response to salinity stress. Metabolomics 2018, 14, 63. [Google Scholar] [CrossRef] [Green Version]
- Velikova, V.B.; Edreva, A.M.; Tsonev, T.D.; Jones, H.G.Z. Singlet oxygen quenching by phenylamides and their parent compounds. Zeitschrift für Naturforschung C 2007, 62, 833–838. [Google Scholar] [CrossRef] [PubMed]
- Von Röpenack, E.; Parr, A.; Schulze-Lefert, P. Structural analyses and dynamics of soluble and cell wall-bound phenolics in a broad spectrum resistance to the powdery mildew fungus in barley. J. Biol. Chem. 1998, 273, 9013–9022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Vogt, T.; Schmidt, J.; Parthier, B.; Löbler, M. Methyljasmonate-induced accumulation of coumaroyl conjugates in barley leaf segments. Phytochemistry 1997, 44, 589–592. [Google Scholar] [CrossRef]
- Ogura, Y.; Ishihara, A.; Iwamura, H. Induction of hydroxycinnamic acid amides and tryptophan by jasmonic acid, abscisic acid and osmotic stress in barley leaves. Zeitschrift für Naturforschung C 2001, 56, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Agati, G.; Azzarello, E.; Pollastri, S.; Tattini, M. Flavonoids as antioxidants in plants: Location and functional significance. Plant Sci. 2012, 196, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.Z.; Li, P.; Wang, J.; Rehman, N.U.; Zhao, J. Isoflavone malonyltransferases GmIMaT1 and GmIMaT3 differently modify isoflavone glucosides in soybean (Glycine max) under various stresses. Front. Plant Sci. 2017, 8, 735. [Google Scholar] [CrossRef]
- Kazan, K.; Lyons, R. The link between flowering time and stress tolerance. J. Exp. Botany 2016, 67, 47–60. [Google Scholar] [CrossRef] [Green Version]
- Refay, Y.A. Impact of soil moisture stress and seeding rate on yield variability of barely grown in arid environment of Saudi Arabia. Am-Eurasian J. Agron. 2009, 2, 185–191. [Google Scholar]
- Sanad, M.N.M.E.; Smertenko, A.; Garland-Campbell, K.A. Differential dynamic changes of reduced trait model for analyzing the plastic response to drought phases: A case study in spring wheat. Front. Plant. Sci. 2019, 10, 504. [Google Scholar] [CrossRef]
- Hura, T.; Hura, K.; Grzesiak, S. Possible contribution of cell-wall-bound ferulic acid in drought resistance and recovery in triticale seedlings. Plant Physiol. 2009, 166, 1720–1733. [Google Scholar] [CrossRef]
- Ballizany, W.L.; Hofmann, R.W.; Jahufer, M.; Zulfiqhar, Z.; Barrett, B.A. Multivariate associations of flavonoid and biomass accumulation in white clover (Trifolium repens) under drought. Funct. Plant Biol. 2012, 39, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Handakumbura, P.P.; Stanfill, B.; Rivas-Ubach, A.; Fortin, D.; Vogel, J.P.; Jansson, C. Metabotyping as a stopover in genome-to-phenome mapping. Sci. Rep. 2019, 9, 1858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennett, T.; Sieberer, T.; Willett, B.; Booker, J.; Luschnig, C.; Leyser, O. The Arabidopsis MAX pathway controls shoot branching by regulating auxin transport. Curr. Biol. 2006, 16, 553–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogrodowicz, P.; Adamski, T.; Mikołajczak, K.; Kuczyńska, A.; Surma, M.; Krajewski, P.; Sawikowska, A.; Górny, A.G.; Gudyś, K.; Szarejko, I.; et al. QTLs for earliness and yield-forming traits in the Lubuski × CamB barley RIL population under various water regimes. J. Appl. Genet. 2017, 58, 49–65. [Google Scholar] [CrossRef] [Green Version]
- Nørbæk, R.; Brandt, K.; Kondo, T. Identification of flavone C-glycosides including a new flavonoid chromophore from barley leaves (Hordeum vulgare L.) by improved NMR techniques. J. Agric. Food Chem. 2000, 48, 1703–1707. [Google Scholar] [CrossRef]
- Haug, K.; Cochrane, K.; Nainala, V.C.; Williams, M.; Chang, J.; Jayaseelan, K.V.; O’Donovan, C. MetaboLights: A resource evolving in response to the needs of its scientific community. Nucleic Acids Res. 2020, gkz1019. [Google Scholar] [CrossRef] [Green Version]
- Tomasi, G.; van den Berg, F.; Andersson, C. Correlation optimized warping and dynamic time warping as preprocessing methods for chromatographic data. J. Chemom. 2004, 18, 231–241. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Drought | No Effect | D | VD | TD | VD,TD | VTD | Total |
|---|---|---|---|---|---|---|---|
| I | 36 (-) | 25 | 0 | 32 | 1 | 10 (+) | 104 |
| II | 61 | 12 (-) | 1 | 26 | 2 | 2 | 104 |
| I+II | 54 | 33 (+) | 0 | 16 | 0 | 1 | 104 |
| Trait Symbol | Group of Traits | Trait Name [units] | Treatment | ||
|---|---|---|---|---|---|
| I | II | I+II | |||
| F1 | Tillers | Number of tillers per plant [pcs] | – | VD | VD |
| F2 | Number of productive tillers per plant [pcs] a) | VD | D | D | |
| F3 | Yield and biomass | Straw weight per plant [g] | VD | D | D |
| F4 | Grain weight per plant [g] | D | D | D | |
| F5 | 1000-grain weight [g] | – | – | – | |
| F6 | Main stem and spike | Length of main stem [cm] | D | D | D |
| F7 | Length of main spike [cm] | D | VD | VD | |
| F8 | Number of spikelets on main spike [pcs] | D | D | D | |
| F9 | Number of grains on main spike [pcs] | D | D | D | |
| F10 | Grain weight on main spike [g] | D | D | VD | |
| F11 | Lateral stems and spikes | Length of lateral stems [cm] | D | D | D |
| F12 | Length of lateral spikes [cm] | D | D | D | |
| F13 | Number of spikelets per lateral spike [pcs] | VD | D | D | |
| F14 | Number of grains per lateral spike [pcs] | VD | VD | D | |
| F15 | Grain weight per lateral spike [g] | VD | VD | - | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piasecka, A.; Sawikowska, A.; Kuczyńska, A.; Ogrodowicz, P.; Mikołajczak, K.; Krajewski, P.; Kachlicki, P. Phenolic Metabolites from Barley in Contribution to Phenome in soil Moisture Deficit. Int. J. Mol. Sci. 2020, 21, 6032. https://doi.org/10.3390/ijms21176032
Piasecka A, Sawikowska A, Kuczyńska A, Ogrodowicz P, Mikołajczak K, Krajewski P, Kachlicki P. Phenolic Metabolites from Barley in Contribution to Phenome in soil Moisture Deficit. International Journal of Molecular Sciences. 2020; 21(17):6032. https://doi.org/10.3390/ijms21176032
Chicago/Turabian StylePiasecka, Anna, Aneta Sawikowska, Anetta Kuczyńska, Piotr Ogrodowicz, Krzysztof Mikołajczak, Paweł Krajewski, and Piotr Kachlicki. 2020. "Phenolic Metabolites from Barley in Contribution to Phenome in soil Moisture Deficit" International Journal of Molecular Sciences 21, no. 17: 6032. https://doi.org/10.3390/ijms21176032