
Pro-Arrhythmic Potential of Accumulated Uremic Toxins Is Mediated via Vulnerability of Action Potential Repolarization
 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Action Potential Prolongation Caused by Chronic Toxin Exposure
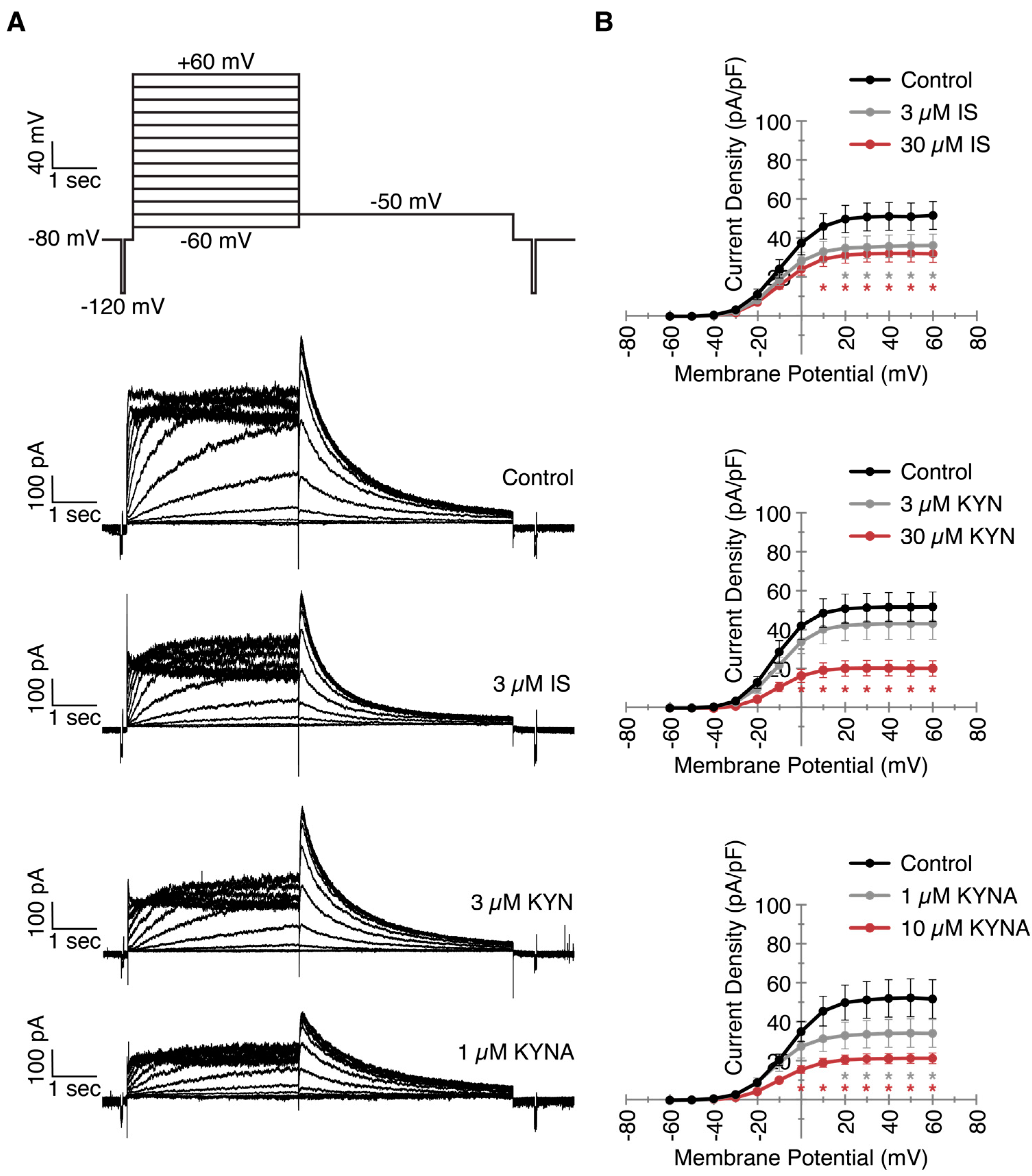
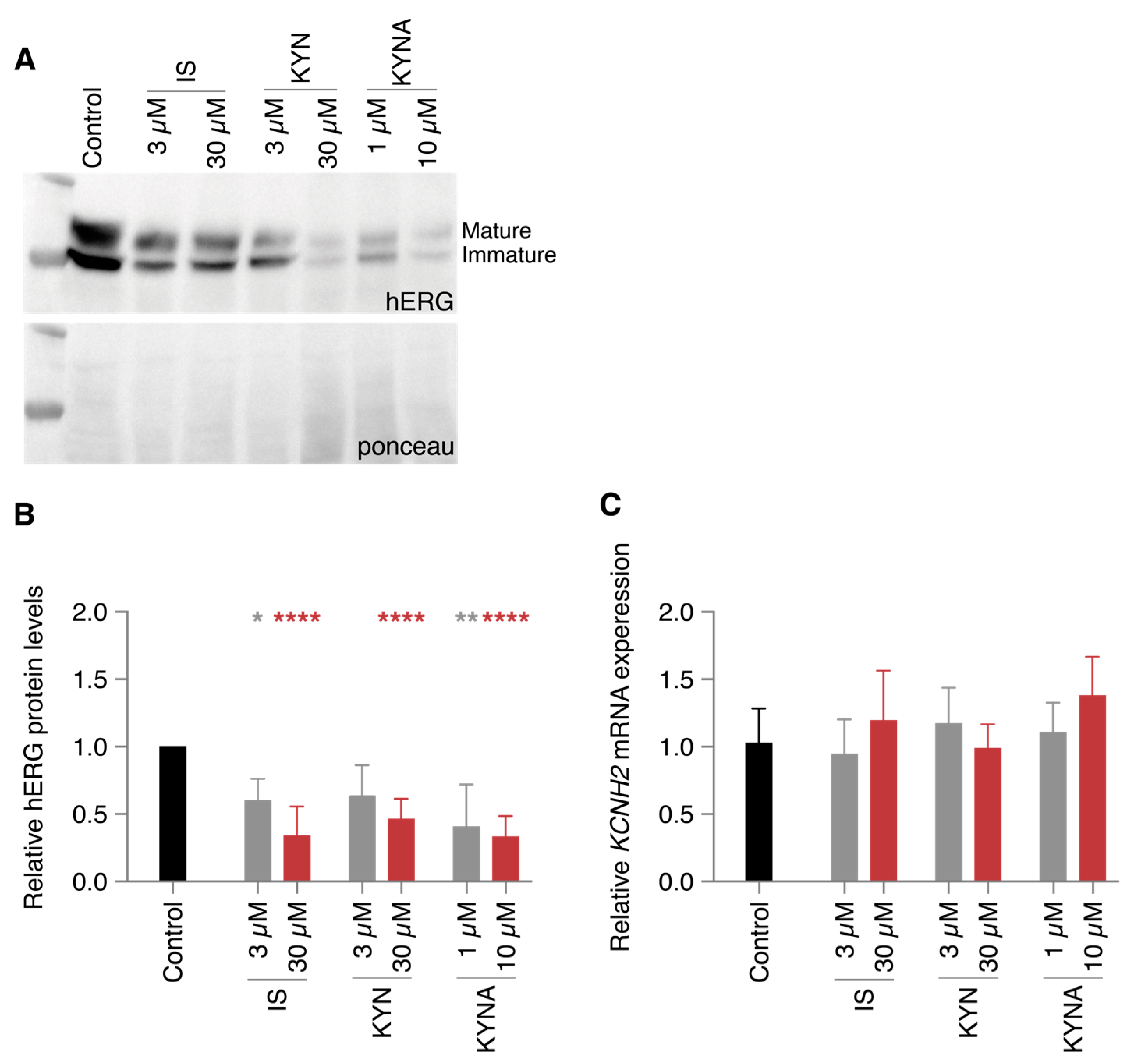
2.2. Decreased IKr Current Density After Exposure to Clinically Relevant UT Concentrations
2.3. Action Potential Prolongation Can Be Reversed by LUF7244
3. Discussion
4. Materials and Methods
4.1. Cell Lines
4.1.1. hiPSCs Culture
4.1.2. hiPSC Cardiomyocyte Differentiation
4.1.3. HEK-hERG Cells
4.2. Drugs and Toxins
4.3. Electrophysiology
4.3.1. Optical Action Potential Measurements
4.3.2. Patch Clamp Electrophysiology
4.4. Western Immunoblotting
4.5. Quantitative PCR
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roth, G.A.; Mensah, G.A.; Johnson, C.O.; Addolorato, G.; Ammirati, E.; Baddour, L.M.; Barengo, N.C.; Beaton, A.Z.; Benjamin, E.J.; Benziger, C.P.; et al. Global Burden of Cardiovascular Diseases and Risk Factors, 1990-2019: Update From the GBD 2019 Study. J. Am. Coll. Cardiol. 2020, 76, 2982–3021. [Google Scholar] [CrossRef]
- Hill, N.R.; Fatoba, S.T.; Oke, J.L.; Hirst, J.A.; O’Callaghan, C.A.; Lasserson, D.S.; Hobbs, F.D. Global Prevalence of Chronic Kidney Disease—A Systematic Review and Meta-Analysis. PLoS ONE 2016, 11, e0158765. [Google Scholar] [CrossRef] [PubMed]
- Kovesdy, C.P. Epidemiology of chronic kidney disease: An update 2022. Kidney Int. Suppl. 2022, 12, 7–11. [Google Scholar] [CrossRef]
- Ronco, C.; Haapio, M.; House, A.A.; Anavekar, N.; Bellomo, R. Cardiorenal syndrome. J. Am. Coll. Cardiol. 2008, 52, 1527–1539. [Google Scholar] [CrossRef] [PubMed]
- Savira, F.; Magaye, R.; Liew, D.; Reid, C.; Kelly, D.J.; Kompa, A.R.; Sangaralingham, S.J.; Burnett, J.C., Jr.; Kaye, D.; Wang, B.H. Cardiorenal syndrome: Multi-organ dysfunction involving the heart, kidney and vasculature. Br. J. Pharmacol. 2020, 177, 2906–2922. [Google Scholar] [CrossRef]
- Cruz, D.N.; Gheorghiade, M.; Palazzuoli, A.; Ronco, C.; Bagshaw, S.M. Epidemiology and outcome of the cardio-renal syndrome. Heart. Fail. Rev. 2011, 16, 531–542. [Google Scholar] [CrossRef]
- Franczyk-Skora, B.; Gluba-Brzozka, A.; Wranicz, J.K.; Banach, M.; Olszewski, R.; Rysz, J. Sudden cardiac death in CKD patients. Int. Urol. Nephrol. 2015, 47, 971–982. [Google Scholar] [CrossRef]
- Green, D.; Roberts, P.R.; New, D.I.; Kalra, P.A. Sudden cardiac death in hemodialysis patients: An in-depth review. Am. J. Kidney. Dis. 2011, 57, 921–929. [Google Scholar] [CrossRef]
- Kumar, U.; Wettersten, N.; Garimella, P.S. Cardiorenal Syndrome: Pathophysiology. Cardiol. Clin. 2019, 37, 251–265. [Google Scholar] [CrossRef]
- Masereeuw, R.; Mutsaers, H.A.; Toyohara, T.; Abe, T.; Jhawar, S.; Sweet, D.H.; Lowenstein, J. The kidney and uremic toxin removal: Glomerulus or tubule? Semin. Nephrol. 2014, 34, 191–208. [Google Scholar] [CrossRef]
- van Gelder, M.K.; Middel, I.R.; Vernooij, R.W.M.; Bots, M.L.; Verhaar, M.C.; Masereeuw, R.; Grooteman, M.P.; Nube, M.J.; van den Dorpel, M.A.; Blankestijn, P.J.; et al. Protein-Bound Uremic Toxins in Hemodialysis Patients Relate to Residual Kidney Function, Are Not Influenced by Convective Transport, and Do Not Relate to Outcome. Toxins 2020, 12, 234. [Google Scholar] [CrossRef]
- Dhondt, A.; Vanholder, R.; Van Biesen, W.; Lameire, N. The removal of uremic toxins. Kidney Int. Suppl. 2000, 76, S47–S59. [Google Scholar] [CrossRef] [PubMed]
- van Ham, W.B.; Cornelissen, C.M.; van Veen, T.A.B. Uremic toxins in chronic kidney disease highlight a fundamental gap in understanding their detrimental effects on cardiac electrophysiology and arrhythmogenesis. Acta Physiol. Oxf. 2022, 236, e13888. [Google Scholar] [CrossRef] [PubMed]
- King, B.M.N.; Mintz, S.; Lin, X.; Morley, G.E.; Schlamp, F.; Khodadadi-Jamayran, A.; Fishman, G.I. Chronic Kidney Disease Induces Proarrhythmic Remodeling. Circ. Arrhythm. Electrophysiol. 2023, 16, e011466. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.Y.; Chen, Y.C.; Kao, Y.H.; Hsieh, M.H.; Lin, Y.K.; Chen, S.A.; Chen, Y.J. Redox and Activation of Protein Kinase A Dysregulates Calcium Homeostasis in Pulmonary Vein Cardiomyocytes of Chronic Kidney Disease. J. Am. Heart. Assoc. 2017, 6, e005701. [Google Scholar] [CrossRef] [PubMed]
- Ke, H.Y.; Chin, L.H.; Tsai, C.S.; Lin, F.Z.; Chen, Y.H.; Chang, Y.L.; Huang, S.M.; Chen, Y.C.; Lin, C.Y. Cardiac calcium dysregulation in mice with chronic kidney disease. J. Cell. Mol. Med. 2020, 24, 3669–3677. [Google Scholar] [CrossRef] [PubMed]
- Redfern, W.S.; Carlsson, L.; Davis, A.S.; Lynch, W.G.; MacKenzie, I.; Palethorpe, S.; Siegl, P.K.; Strang, I.; Sullivan, A.T.; Wallis, R.; et al. Relationships between preclinical cardiac electrophysiology, clinical QT interval prolongation and torsade de pointes for a broad range of drugs: Evidence for a provisional safety margin in drug development. Cardiovasc. Res. 2003, 58, 32–45. [Google Scholar] [CrossRef]
- Kiehn, J.; Karle, C.; Thomas, D.; Yao, X.; Brachmann, J.; Kubler, W. HERG potassium channel activation is shifted by phorbol esters via protein kinase A-dependent pathways. J. Biol. Chem. 1998, 273, 25285–25291. [Google Scholar] [CrossRef]
- Yu, Z.; van Veldhoven, J.P.; ME’t Hart, I.M.; Kopf, A.H.; Heitman, L.H.; IJzerman, A.P. Synthesis and biological evaluation of negative allosteric modulators of the Kv11.1(hERG) channel. Eur. J. Med. Chem. 2015, 106, 50–59. [Google Scholar] [CrossRef]
- Vanholder, R.; Pletinck, A.; Schepers, E.; Glorieux, G. Biochemical and Clinical Impact of Organic Uremic Retention Solutes: A Comprehensive Update. Toxins 2018, 10, 33. [Google Scholar] [CrossRef]
- Savira, F.; Magaye, R.; Hua, Y.; Liew, D.; Kaye, D.; Marwick, T.; Wang, B.H. Molecular mechanisms of protein-bound uremic toxin-mediated cardiac, renal and vascular effects: Underpinning intracellular targets for cardiorenal syndrome therapy. Toxicol. Lett. 2019, 308, 34–49. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.H.; Wang, C.P.; Chung, F.M.; Huang, L.L.; Yu, T.H.; Hung, W.C.; Lu, L.F.; Chen, P.Y.; Luo, C.H.; Lee, K.T.; et al. Uremic retention solute indoxyl sulfate level is associated with prolonged QTc interval in early CKD patients. PLoS ONE 2015, 10, e0119545. [Google Scholar] [CrossRef] [PubMed]
- Tsai, I.T.; Hsu, C.C.; Hung, W.C.; Wang, C.P.; Yu, T.H.; Houng, J.Y.; Lee, K.T.; Tang, W.H. The Arrhythmogenic Effect of Protein-Bound Uremic Toxin p-Cresylsulfate: An In Vitro Study. Acta Cardiol. Sin. 2019, 35, 641–648. [Google Scholar] [CrossRef]
- Duranton, F.; Cohen, G.; De Smet, R.; Rodriguez, M.; Jankowski, J.; Vanholder, R.; Argiles, A.; European Uremic Toxin Work Group. Normal and pathologic concentrations of uremic toxins. J. Am. Soc. Nephrol. 2012, 23, 1258–1270. [Google Scholar] [CrossRef]
- Pavlenko, D.; van Geffen, E.; van Steenbergen, M.J.; Glorieux, G.; Vanholder, R.; Gerritsen, K.G.; Stamatialis, D. New low-flux mixed matrix membranes that offer superior removal of protein-bound toxins from human plasma. Sci. Rep. 2016, 6, 34429. [Google Scholar] [CrossRef]
- Ter Beek, O.E.M.; van Gelder, M.K.; Lokhorst, C.; Hazenbrink, D.H.M.; Lentferink, B.H.; Gerritsen, K.G.F.; Stamatialis, D. In vitro study of dual layer mixed matrix hollow fiber membranes for outside-in filtration of human blood plasma. Acta Biomater. 2021, 123, 244–253. [Google Scholar] [CrossRef]
- Yu, Z.; Liu, J.; van Veldhoven, J.P.; AP, I.J.; Schalij, M.J.; Pijnappels, D.A.; Heitman, L.H.; de Vries, A.A. Allosteric Modulation of Kv11.1 (hERG) Channels Protects Against Drug-Induced Ventricular Arrhythmias. Circ. Arrhythm. Electrophysiol. 2016, 9, e003439. [Google Scholar] [CrossRef]
- Qile, M.; Beekman, H.D.M.; Sprenkeler, D.J.; Houtman, M.J.C.; van Ham, W.B.; Stary-Weinzinger, A.; Beyl, S.; Hering, S.; van den Berg, D.J.; de Lange, E.C.M.; et al. LUF7244, an allosteric modulator/activator of K(v) 11.1 channels, counteracts dofetilide-induced torsades de pointes arrhythmia in the chronic atrioventricular block dog model. Br. J. Pharmacol. 2019, 176, 3871–3885. [Google Scholar] [CrossRef]
- Qile, M.; Ji, Y.; Golden, T.D.; Houtman, M.J.C.; Romunde, F.; Fransen, D.; van Ham, W.B.; AP, I.J.; January, C.T.; Heitman, L.H.; et al. LUF7244 plus Dofetilide Rescues Aberrant K(v)11.1 Trafficking and Produces Functional I(Kv11.1). Mol. Pharmacol. 2020, 97, 355–364. [Google Scholar] [CrossRef]
- Zhou, Z.; Gong, Q.; Epstein, M.L.; January, C.T. HERG channel dysfunction in human long QT syndrome. Intracellular transport and functional defects. J. Biol. Chem. 1998, 273, 21061–21066. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
van Ham, W.B.; Cornelissen, C.M.; Polyakova, E.; van der Voorn, S.M.; Ligtermoet, M.L.; Monshouwer-Kloots, J.; Vos, M.A.; Bossu, A.; van Rooij, E.; van der Heyden, M.A.G.; et al. Pro-Arrhythmic Potential of Accumulated Uremic Toxins Is Mediated via Vulnerability of Action Potential Repolarization. Int. J. Mol. Sci. 2023, 24, 5373. https://doi.org/10.3390/ijms24065373
van Ham WB, Cornelissen CM, Polyakova E, van der Voorn SM, Ligtermoet ML, Monshouwer-Kloots J, Vos MA, Bossu A, van Rooij E, van der Heyden MAG, et al. Pro-Arrhythmic Potential of Accumulated Uremic Toxins Is Mediated via Vulnerability of Action Potential Repolarization. International Journal of Molecular Sciences. 2023; 24(6):5373. https://doi.org/10.3390/ijms24065373
Chicago/Turabian Stylevan Ham, Willem B., Carlijn M. Cornelissen, Elizaveta Polyakova, Stephanie M. van der Voorn, Merel L. Ligtermoet, Jantine Monshouwer-Kloots, Marc A. Vos, Alexandre Bossu, Eva van Rooij, Marcel A. G. van der Heyden, and et al. 2023. "Pro-Arrhythmic Potential of Accumulated Uremic Toxins Is Mediated via Vulnerability of Action Potential Repolarization" International Journal of Molecular Sciences 24, no. 6: 5373. https://doi.org/10.3390/ijms24065373