
A Comparative Study of the Antiemetic Effects of α2-Adrenergic Receptor Agonists Clonidine and Dexmedetomidine against Diverse Emetogens in the Least Shrew (Cryptotis parva) Model of Emesis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
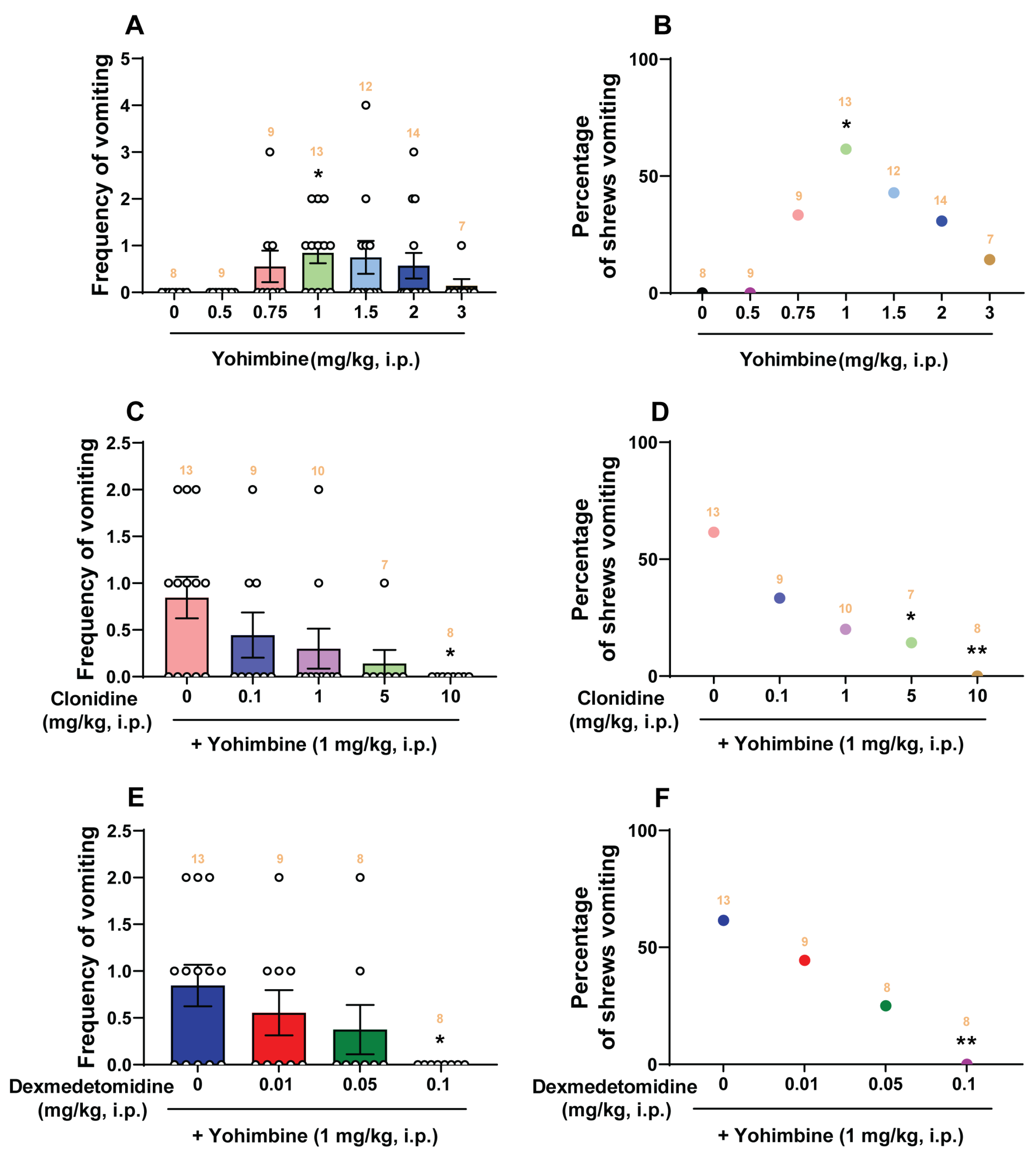
2.1. Dose-Response Emetic Effect of Yohimbine in Least Shrews
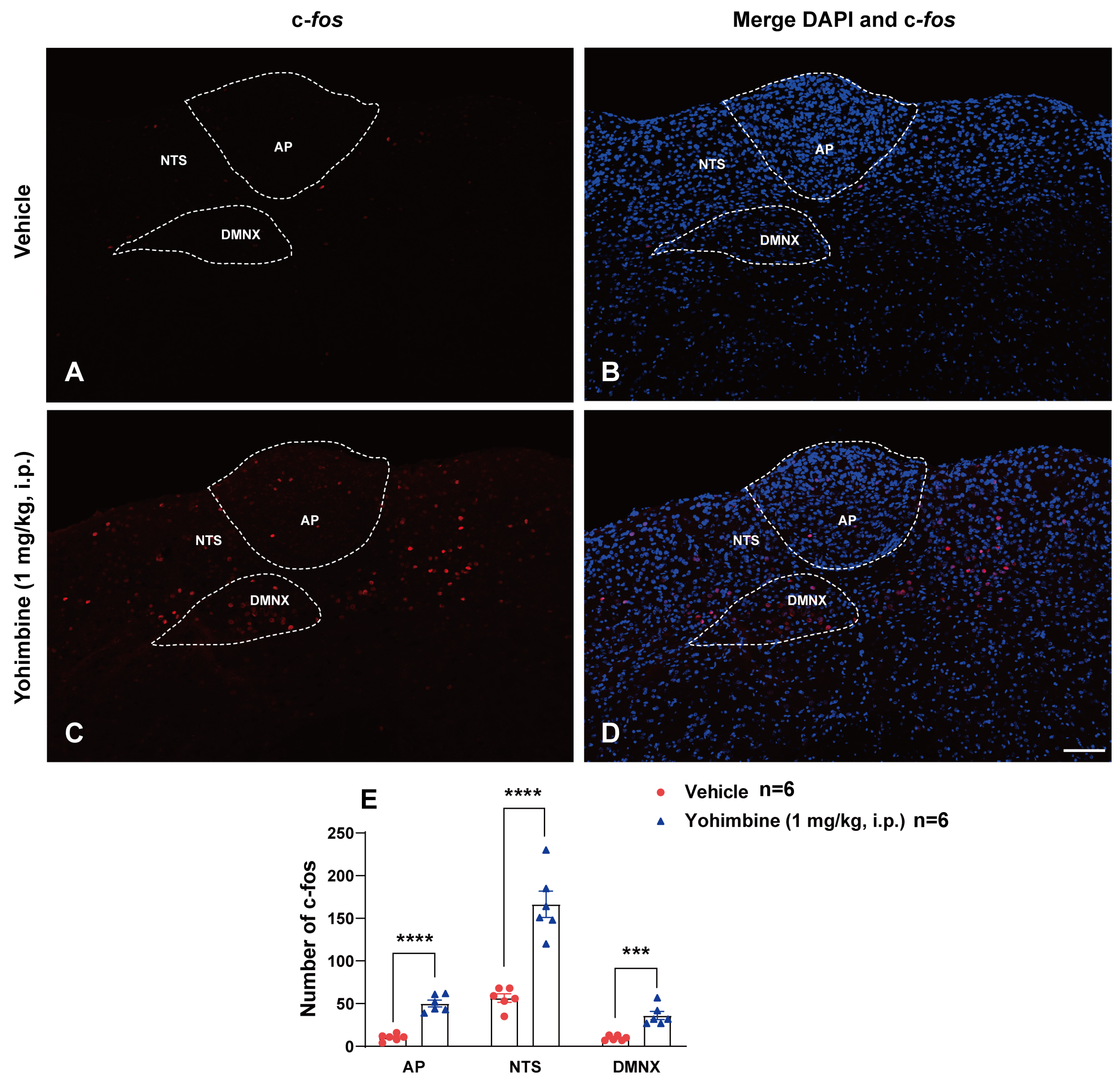
2.2. Yohimbine Activates the Brainstem Emetic Nuclei
2.3. Effect of Yohimbine on 5-HT- and SP-Release in Brainstem Emetic Nuclei
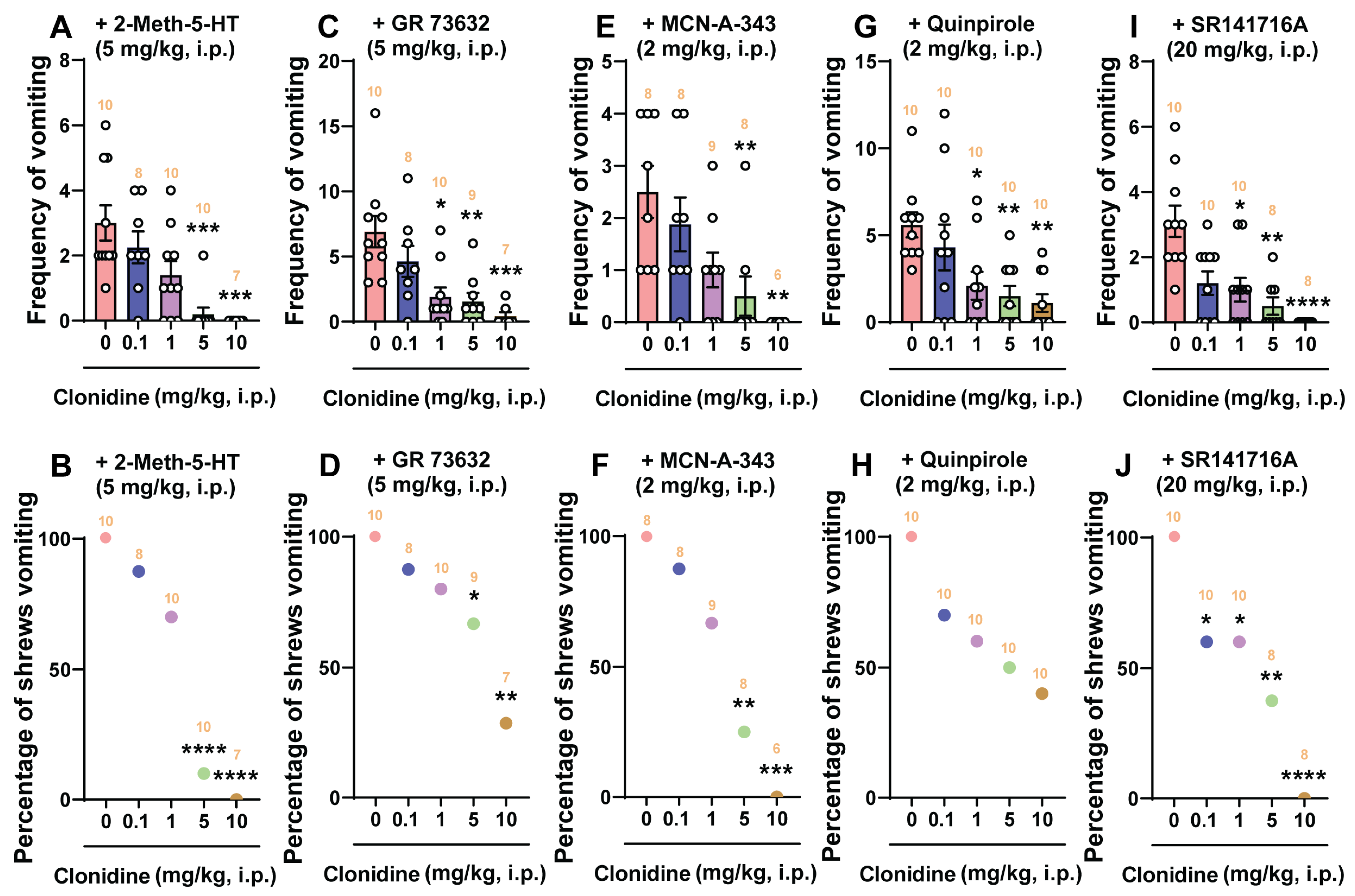
2.4. The Broad-Spectrum Antiemetic Potential of the α2-Adrenergic Receptor Agonists Clonidine and Dexmedetomidine against Vomiting Evoked by Diverse Receptor-Selective Emetogens
2.5. The Antiemetic Potential of the α2-Adrenergic Receptor Agonists Clonidine and Dexmedetomidine against Vomiting Evoked by Ca2+ Channel Regulators
2.6. The Antiemetic Potential of Clonidine and Dexmedetomidine against Vomiting Evoked by Rolipram, the Inhibitor of PDE4 in Shrews
2.7. The Antiemetic Effect of Clonidine and Dexmedetomidine against Vomiting Evoked by the HCN Channel Blocker ZD7288
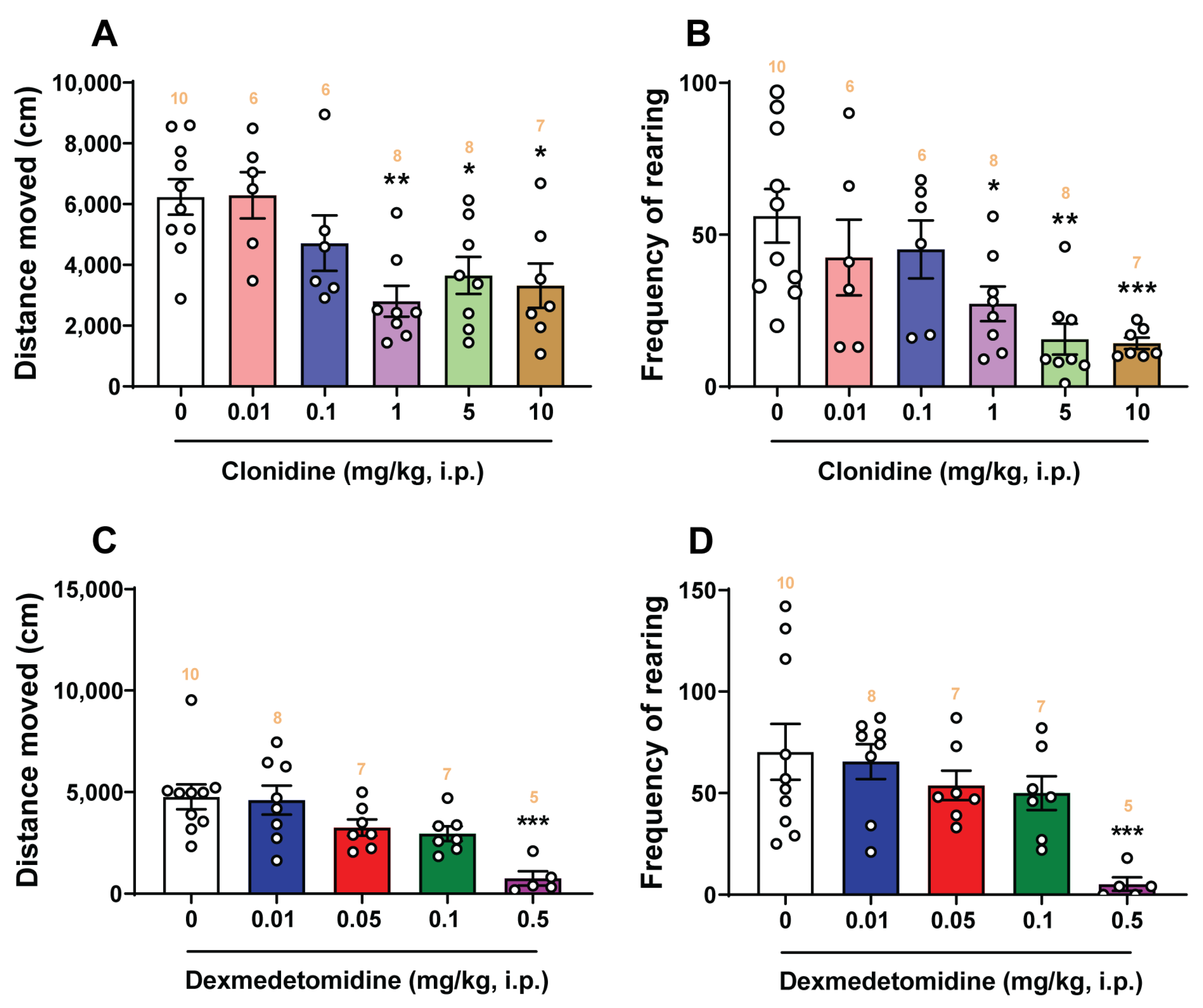
2.8. Open-Field Locomotor Studies
3. Discussion
3.1. Significance of the Present Study
3.2. Emetic Effect of Yohimbine in the Least Shrew
3.3. Yohimbine-Induced Expression of c-fos and Release of 5-HT and SP in the DVC Central Emetic Loci
3.4. Effects of α2-Adrenergic Receptor Agonists Clonidine and Dexmedetomidine on Locomotor Activity
3.5. Antiemetic Effects of α2-Adrenergic Receptor Agonists Clonidine and Dexmedetomidine against Yohimbine and Diverse Emetogens
- The α2-adrenergic receptor antagonist yohimbine (1 mg/kg, i.p.): Both clonidine and dexmedetomidine reduced the mean frequency and percentage of shrews vomiting in response to yohimbine in a dose-dependent manner with respective ID50 values ranging between 0.21 mg/kg and 0.021 mg/kg, respectively. Thus, dexmedetomidine appears to be ten times more potent antiemetic than clonidine against yohimbine-evoked emesis, and both agents completely protected shrews from vomiting at their maximal tested dose of 0.1 and 10 mg/kg, respectively. Likewise, dexmedetomidine is reported to be eight times more potent than clonidine in the clinic [71]. It is suggested that yohimbine-induced vomiting could be due to blockade of the inhibitory α2-adrenergic receptors, which is associated with the enhanced release of monoamines NE, DA, and 5-HT [50]. Clonidine overrides the ability of yohimbine to release monoamines, and thus prevents vomiting evoked by the monoamine releaser, reserpine [31]. These maximal tested doses of clonidine and dexmedetomidine in least shrews were subsequently used as their utmost tested doses against vomiting produced by the following emetogens.
- The more selective and centrally/peripherally-acting 5-HT3 receptor agonist, 2-methyl-5-HT (5 mg/kg, i.p.): 5-HT3 receptor selective antagonists, such as tropisetron [72] or palonosetron [40], can suppress vomiting caused by 2-Methyl-5-HT. Here, clonidine and dexmedetomidine suppressed the mean frequency of vomiting and protected shrews from emesis in a dose-dependent manner. In the latter case, dexmedetomidine was 25 times more potent than clonidine in suppressing 2-Methyl-5-HT-evoked vomiting. However, clonidine completely protected shrews from vomiting at its highest tested dose (10 mg/kg), whereas the largest dose of dexmedetomidine (0.1 mg/kg) only prevented 75% of shrews from vomiting. This difference probably reflects the nonselective nature of clonidine since the most potent and selective antagonist of 5-HT3 receptors, palonosetron (10 mg/kg, i.p.), could not completely prevent least shrews from 2-Methyl-5-HT-induced vomiting [40]. Ca2+ mobilization via extracellular Ca2+ influx through 5-HT3 receptors and L-type Ca2+ channels, and intracellular Ca2+ release via ryanodine receptors (RyRs) present on the endoplasmic reticulum (ER), initiate Ca2+-dependent sequential activation of CaMKIIa and signal-regulated kinase 1/2 (ERK1/2), which contribute to 2-Methyl-5-HT-evoked emesis [73]. The most studied signaling pathway of α2-adrenergic receptor is through G protein-coupled receptors, which consists of direct inhibition (“membrane-delimited”) by G protein βγ (Gβγ) subunit complexes of Ca2+ entry through voltage-gated Ca2+ (Cav) channels [74]. For α2-adrenergic receptor, the L-type Cav channel is the main effector [75]. Typically, L-type calcium currents are maximally inhibited by 50% upon α2-adrenergic agonist application [74]. We suggest that the antiemetic effects of clonidine and dexmedetomidine on 2-Methyl-5-HT-evoked emesis is probably related to the inhibition of the L-type calcium currents.
- The selective SP neurokinin NK1 receptor agonist GR73632 (5 mg/kg, i.p.): At this dose, GR73632 causes robust vomiting in least shrews through the activation of SP neurokinin NK1 receptors [76], which can be completely blocked by the selective and potent NK1 receptor antagonist netupitant (10 mg/kg, i.p.) in least shrews [77]. In the present study, both clonidine and dexmedetomidine prevented GR73632-evoked emesis in a dose-dependent manner. However, the largest tested dose of clonidine (10 mg/kg, i.p.) reduced the frequency of GR73632-evoked vomiting by 94% and the percentage of least shrews vomiting by 71.4%. Also, dexmedetomidine at its maximum tested dose (0.1 mg/kg) significantly but partially attenuated both the mean vomit frequency (76.3%) and the percentage of least shrews vomiting (62.5%) in response to GR73632. Their ID50 values indicate that dexmedetomidine is more potent than clonidine in preventing GR73632-evoked vomiting. The NK1 receptor is G-protein coupled and can increase cytoplasmic Ca2+ concentration [78,79,80]. In fact, GR73632 evokes an increase in intracellular Ca2+ concentration via both Ca2+ release from intracellular Ca2+ stores, and extracellular Ca2+ influx through the transient receptor potential channels [79]. In addition, we have shown that LTCC blockers amlodipine and nifedipine also suppress vomiting caused by GR73632 in a dose-dependent manner [40]. Per our discussion above, we suggest that the antiemetic effects of clonidine and dexmedetomidine on GR73632-induced emesis may involve suppression of intracellular Ca2+ concentration.
- The cholinergic M1 receptor agonist McN-A-343 (2 mg/kg, i.p.): Clonidine reduced both the mean frequency and the percentage of least shrews vomiting in response to McN-A-343 in a dose-dependent fashion, with complete emesis protection occurring at its 10 mg/kg dose. However, dexmedetomidine significantly but partially reduced the mean frequency of vomiting (66.7%) at its tested maximal dose of 0.1 mg/kg but failed to significantly protect shrews from vomiting. McN-A-343 can activate PKC and PKA phosphorylates to enhance Ca2+ influx through LTCC channels [81,82]. Moreover, the L-type calcium antagonist nifedipine can significantly and completely prevent in a potent and dose-dependent manner both the percentage of shrews vomiting and the mean frequency of emesis evoked by McN-A-343 (2 mg/kg, i.p.) in the least shrew [83]. The antiemetic effects of clonidine and dexmedetomidine on McN-A-343-induced emesis may also involve suppression of intracellular Ca2+ mobilization.
- The dopamine D2/3 receptor preferring agonist quinpirole (2 mg/kg, i.p.): Clonidine only significantly but partially reduced the mean frequency of quinpirole-evoked vomiting by 80.4% at its 10 mg/kg maximal tested dose but failed to completely protect shrews from vomiting. Likewise, dexmedetomidine significantly reduced both the mean frequency (57.8% reduction at 0.1 mg/kg) and the percentage of least shrews vomiting (36.4% protection at 0.1 mg/kg) in response to quinpirole, but these reductions were only partial. Although the dopamine D2 receptor-preferring antagonist sulpiride can completely prevent apomorphine (2 mg/kg, i.p.)-induced vomiting in least shrews at 2 mg/kg (s.c.), but it cannot fully protect shrews from quinpirole (2 mg/kg, i.p.)-evoked emesis even up to 8 mg/kg [84]. Thus, it appears that quinpirole-evoked emesis cannot be easily subdued by sulpiride. Mechanistically, we have previously demonstrated the involvement of the PI3K/mTOR/Akt signaling pathway in dopamine D2 receptor-evoked vomiting [85]. Since the α2-adrenergic receptor agonist tizanidine can reduce the expression levels of PI3K/Akt [86], it is possible that clonidine and dexmedetomidine may also affect this signaling system to suppress the evoked vomiting, but this remains to be investigated.
- The cannabinoid CB1 receptor-selective inverse agonist/antagonist SR141716A (20 mg/kg, i.p.): SR141716A can induce vomiting in the least shrew at large doses (20–40 mg/kg, i.p.) which can be fully prevented by CB1 receptor agonists, including Δ9-THC [87]. Clonidine significantly reduced both the mean frequency and the percentage of shrews vomiting in response to SR141716A in a dose-dependent manner with complete emesis protection at it 10 mg/kg dose. Dexmedetomidine reduced the mean frequency of vomiting by 88% and protected shrews from vomiting by 75% at its maximal tested dose of 0.1 mg/kg. SR141716A increases the release and turnover of monoamines DA, NE, and 5-HT [88] in several brain regions of rodents and least shrews, and can enhance capsaicin-evoked release of SP in the mouse spinal cord [89]. As discussed earlier, since clonidine can prevent vomiting caused by the monoamine releaser reserpine [31], it probably prevents SR141716A-evoked vomiting via a similar mechanism.
- The selective LTCC agonist FPL64176 (10 m/kg, i.p.): The LTCC regulates extracellular Ca2+ influx into the cytosol [90]. FPL64176 is an extracellular Ca2+-mobilizing agent and evokes vomiting in all tested shrews at 10 mg/kg [40,83]. Clonidine significantly and completely reduced the mean vomiting frequency and the percentage of shrews vomiting in response to FPL64176 challenge in a dose-dependent manner. Dexmedetomidine significantly and dose-dependently reduced the mean vomit frequency by 92.3% following its maximal tested dose (0.1 mg/kg), and protected 71.4% of shrews from FPL64176 (10 mg/kg, i.p.)-induced vomiting. However, their ID50 values indicate that dexmedetomidine is more potent than clonidine in preventing FPL64176-evoked vomiting. As was discussed earlier, the antiemetic effects of clonidine and dexmedetomidine on FPL64176-evoked emesis may also involve inhibition of L-type calcium currents.
- Specific inhibitor of the SERCA pump thapsigargin (0.5 mg/kg, i.p.): The SERCA pump is a main mechanism that transports free cytosolic Ca2+ into endoplasmic reticulum (ER) Ca2+ stores. Release of Ca2+ from the ER stores into the cytosol occurs through the inositol trisphosphate (IP3)- and ryanodine (RyR)-receptor ion channels localized on the ER membrane [91,92]. Thapsigargin is a specific and potent inhibitor of SERCA pumps and causes a rapid elevation in cytosolic Ca2+ concentrations. Thapsigargin can also release Ca2+ from the ER stores into the cytosol via IP3 and RyR calcium channels [93,94,95]. Thapsigargin causes vomiting by triggering an initial elevation in the cytoplasmic Ca2+ concentration by inhibiting the SERCA as well as releasing Ca2+ from the ER into the cytoplasm via both RyR- and IP3-receptors (IP3Rs), which is followed by an extracellular Ca2+ influx through LTCCs prior to the intracellular activation of the Ca2+-CaMKII-ERK1/2 cascade [41]. In the present study, clonidine significantly reduced the mean frequency of thapsigargin-evoked vomiting by 93.3% at its 10 mg/kg dose, but only partially protected 66.7% of shrews at this maximal dose. Moreover, dexmedetomidine only reduced the mean frequency of the evoked vomiting by 70.8% at its 0.1 mg/kg maximal dose but failed to significantly protect shrews from vomiting. Since the LTCC inhibitor nifedipine can dose-dependently inhibit thapsigargin-induced vomiting [41], we speculate that clonidine and dexmedetomidine may partially suppress extracellular Ca2+ influx through LTCCs, which could then attenuate the above-discussed signaling cascade, leading to a limited reduction in thapsigargin-evoked vomiting.
- The PDE4 inhibitor rolipram (1 mg/kg, i.p.): PDE4 inhibitors prevent metabolism of second messengers such as cAMP and increase their tissue levels. They have procognitive and antidepressant properties [96]. The emetic effect of some PDE4 inhibitors is thought to be a consequence of inhibition of PDE4 and the subsequent increase in cAMP levels in the brainstem DVC [30,42]. PDE4 inhibitors may mimic the pharmacological effect of α2-adrenergic receptor antagonists, which elevate intracellular levels of cAMP in noradrenergic neurons [30]. In contrast, α2-adrenergic receptor activation decreases intracellular levels of cAMP in noradrenergic neurons. PDE4 inhibitors are thought to modulate the release of emetic mediators including 5-HT, SP, and noradrenaline which are involved in the onset of the emetic reflex [30]. We have previously shown that the PDE4 inhibitor rolipram (1 mg/kg, i.p.) evokes both vomiting as well as significant increases in shrew brainstem cAMP levels, while pretreatment with SQ22536, an inhibitor of adenylyl cyclase, prevented the evoked emesis [42]. In the present study, clonidine dose-dependently and significantly, albeit partially, reduced both the mean vomit frequency (80.3%) and the percentage (62.5%) of shrews vomiting in response to rolipram at its highest tested dose, 10 mg/kg. Likewise, dexmedetomidine produced similar dose-dependent but partial reductions in both the mean vomit frequency (73.7%) and the percentage of shrews vomiting (60%) at its maximal tested dose, 0.1 mg/kg. The antiemetic effect of clonidine and dexmedetomidine against rolipram-induced vomiting might be due to suppression of increased intracellular tissue levels of cAMP evoked by rolipram in the shrew brainstem. In fact, administration of cAMP analogs such as 8-chloro-cAMP cause vomiting in cancer patients [97].
- The HCN blocker ZD7288 (1 mg/kg, i.p.): The HCN channels are a class of voltage-gated ion-channels permeable to Na+ and K+ and constitutively open at voltages near the resting membrane potential [98,99]. The hyperpolarization-activated currents (Ih) mediated by HCN channels elicit membrane depolarization toward a threshold for action potential generation, which plays a pivotal role in controlling neuronal excitability [98,100]. The HCN channel blocker ZD7288 can reduce apomorphine-induced conditioned taste aversion to saccharin preference and depress the excitability of the AP since it blocks HCN channel activation [101]. We have recently demonstrated that ZD7288 (1 mg/kg, i.p.) evokes both vomiting in a dose-dependent manner as well as a robust expression of c-fos and ERK1/2 phosphorylation in the shrew brainstem DVC, indicating a central contribution to the evoked vomiting [43]. In the present study, clonidine significantly and dose-dependently reduced ZD7288-evoked mean vomit frequency (92.4% reduction at 10 mg/kg), but only partially protected shrews from vomiting (55.6% protection at 10 mg/kg). Dexmedetomidine only significantly reduced the mean frequency (63.1% protection at 0.1 mg/kg) and failed to significantly protect shrews from ZD7288-evoked vomiting. Given the well-known negative coupling of α2-adrenergic receptors to adenylate cyclase via a heterotrimeric G protein [102], any reduction in the cAMP level would decrease Ih. Dexmedetomidine via α2-adrenergic receptors activates G-protein-coupled K+ channels and inhibits Ih, which leads to membrane hyperpolarization [103]. In addition, clonidine can directly inhibit If current [11]. Hence, we speculate that administration of clonidine and dexmedetomidine reverse the inhibitory effect of ZD7288 on the HCN channels and consequently suppresses ZD7288-induced vomiting in shrews.
4. Materials and Methods
4.1. Animals
4.2. Chemicals
4.3. Behavioral Emesis Studies
4.4. Immunohistochemistry and Image Analysis
4.4.1. c-fos Staining and Image Analysis
4.4.2. 5-HT and SP Immunohistochemistry
4.5. Locomotor Activity Studies
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Carpenter, D.O. Neural mechanisms of emesis. Can. J. Physiol. Pharmacol. 1990, 68, 230–236. [Google Scholar] [CrossRef]
- Horn, C.C. Why is the neurobiology of nausea and vomiting so important? Appetite 2008, 50, 430–434. [Google Scholar] [CrossRef]
- Darmani, N.A.; Ray, A.P. Evidence for a re-evaluation of the neurochemical and anatomical bases of chemotherapy-induced vomiting. Chem. Rev. 2009, 109, 3158–3199. [Google Scholar] [CrossRef] [PubMed]
- Hesketh, P.J. Chemotherapy-induced nausea and vomiting. N. Engl. J. Med. 2008, 358, 2482–2494. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Yoon, S.S.; Kuo, B. Nausea: A review of pathophysiology and therapeutics. Therap. Adv. Gastroenterol. 2016, 9, 98–112. [Google Scholar] [CrossRef] [PubMed]
- Zhong, W.; Shahbaz, O.; Teskey, G.; Beever, A.; Kachour, N.; Venketaraman, V.; Darmani, N.A. Mechanisms of Nausea and Vomiting: Current Knowledge and Recent Advances in Intracellular Emetic Signaling Systems. Int. J. Mol. Sci. 2021, 22, 5797. [Google Scholar] [CrossRef]
- Babic, T.; Browning, K.N. The role of vagal neurocircuits in the regulation of nausea and vomiting. Eur. J. Pharmacol. 2014, 722, 38–47. [Google Scholar] [CrossRef]
- Pertovaara, A. The noradrenergic pain regulation system: A potential target for pain therapy. Eur. J. Pharmacol. 2013, 716, 2–7. [Google Scholar] [CrossRef] [PubMed]
- Phillips, J.K.; McLean, A.J.; Hill, C.E. Receptors involved in nerve-mediated vasoconstriction in small arteries of the rat hepatic mesentery. Br. J. Pharmacol. 1998, 124, 1403–1412. [Google Scholar] [CrossRef]
- Leao, R.M.; Von Gersdorff, H. Noradrenaline increases high-frequency firing at the calyx of Held synapse during development by inhibiting glutamate release. J. Neurophysiol. 2002, 87, 2297–2306. [Google Scholar] [CrossRef]
- Knaus, A.E.; Muthig, V.; Schickinger, S.; Moura, E.; Beetz, N.; Gilsbach, R.; Hein, L. Alpha2-adrenoceptor subtypes--unexpected functions for receptors and ligands derived from gene-targeted mouse models. Neurochem. Int. 2007, 51, 277–281. [Google Scholar] [CrossRef] [PubMed]
- Davis, R.; Wilde, M.I. Mirtazapine: A Review of its Pharmacology and Therapeutic Potential in the Management of Major Depression. CNS Drugs 1996, 5, 389–402. [Google Scholar] [CrossRef] [PubMed]
- Van Bockstaele, E.J. Morphological substrates underlying opioid, epinephrine and gamma-aminobutyric acid inhibitory actions in the rat locus coeruleus. Brain Res. Bull. 1998, 47, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Kirouac, G.J. Placing the paraventricular nucleus of the thalamus within the brain circuits that control behavior. Neurosci. Biobehav. Rev. 2015, 56, 315–329. [Google Scholar] [CrossRef] [PubMed]
- Van Bockstaele, E.J.; Peoples, J.; Telegan, P. Efferent projections of the nucleus of the solitary tract to peri-locus coeruleus dendrites in rat brain: Evidence for a monosynaptic pathway. J. Comp. Neurol. 1999, 412, 410–428. [Google Scholar] [CrossRef]
- McCune, S.K.; Voigt, M.M.; Hill, J.M. Expression of multiple alpha adrenergic receptor subtype messenger RNAs in the adult rat brain. Neuroscience 1993, 57, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Nicholas, A.P.; Pieribone, V.; Hokfelt, T. Distributions of mRNAs for alpha-2 adrenergic receptor subtypes in rat brain: An in situ hybridization study. J. Comp. Neurol. 1993, 328, 575–594. [Google Scholar] [CrossRef] [PubMed]
- Robertson, S.D.; Plummer, N.W.; de Marchena, J.; Jensen, P. Developmental origins of central norepinephrine neuron diversity. Nat. Neurosci. 2013, 16, 1016–1023. [Google Scholar] [CrossRef]
- Hikasa, Y.; Takase, K.; Ogasawara, S. Evidence for the involvement of alpha 2-adrenoceptors in the emetic action of xylazine in cats. Am. J. Vet. Res. 1989, 50, 1348–1351. [Google Scholar]
- Hikasa, Y.; Ogasawara, S.; Takase, K. Alpha adrenoceptor subtypes involved in the emetic action in dogs. J. Pharmacol. Exp. Ther. 1992, 261, 746–754. [Google Scholar]
- Hikasa, Y.; Akiba, T.; Iino, Y.; Matsukura, M.; Takase, K.; Ogasawara, S. Central alpha-adrenoceptor subtypes involved in the emetic pathway in cats. Eur. J. Pharmacol. 1992, 229, 241–251. [Google Scholar]
- Japundzic-Zigon, N.; Samardzic, R.; Beleslin, D.B. Clonidine-induced emesis: A multitransmitter pathway concept. Pharmacol. Res. 1997, 35, 287–297. [Google Scholar] [CrossRef]
- McSweeney, P.M.; Martin, D.D.; Ramsey, D.S.; McKusick, B.C. Clinical efficacy and safety of dexmedetomidine used as a preanesthetic prior to general anesthesia in cats. J. Am. Vet. Med. Assoc. 2012, 240, 404–412. [Google Scholar] [CrossRef] [PubMed]
- Santos, L.C.; Ludders, J.W.; Erb, H.N.; Martin-Flores, M.; Basher, K.L.; Kirch, P. A randomized, blinded, controlled trial of the antiemetic effect of ondansetron on dexmedetomidine-induced emesis in cats. Vet. Anaesth. Analg. 2011, 38, 320–327. [Google Scholar] [CrossRef] [PubMed]
- Granholm, M.; McKusick, B.C.; Westerholm, F.C.; Aspegren, J.C. Evaluation of the clinical efficacy and safety of dexmedetomidine or medetomidine in cats and their reversal with atipamezole. Vet. Anaesth. Analg. 2006, 33, 214–223. [Google Scholar] [CrossRef]
- Brioschi, F.A.; Gioeni, D.; Jacchetti, A.; Carotenuto, A.M. Effect of metoclopramide on nausea and emesis in dogs premedicated with morphine and dexmedetomidine. Vet. Anaesth. Analg. 2018, 45, 190–194. [Google Scholar] [CrossRef] [PubMed]
- Nystrom, M.R.; Odunayo, A.; Okafor, C.C. Assessment of hydromorphone and dexmedetomidine for emesis induction in cats. J. Vet. Emerg. Crit. Care 2019, 29, 360–365. [Google Scholar] [CrossRef]
- Papastefanou, A.K.; Galatos, A.D.; Pappa, E.; Lymperis, A.G.; Kostoulas, P. The effect of butorphanol on the incidence of dexmedetomidine-induced emesis in cats. Vet. Anaesth. Analg. 2015, 42, 608–613. [Google Scholar] [CrossRef] [PubMed]
- Hassen, K.M.; Posner, L.P.; Campbell, N.B. The effect of aquapuncture at Pericardium 6 (PC-6) on dexmedetomidine-induced nausea and vomiting in cats. Vet. Anaesth. Analg. 2019, 46, 308–314. [Google Scholar] [CrossRef]
- Robichaud, A.; Savoie, C.; Stamatiou, P.B.; Tattersall, F.D.; Chan, C.C. PDE4 inhibitors induce emesis in ferrets via a noradrenergic pathway. Neuropharmacology 2001, 40, 262–269. [Google Scholar] [CrossRef]
- Khandker, S.K.; Mukerjee, D.; Gurtu, S.; Pant, K.K.; Dhawan, K.N.; Sinha, J.N. Modification of reserpine-induced emetic response in pigeons by alpha 2-adrenoceptors. Pharmacol. Res. 1994, 29, 383–387. [Google Scholar] [CrossRef] [PubMed]
- Giampreti, A.; Lonati, D.; Locatelli, C.; Rocchi, L.; Campailla, M.T. Acute neurotoxicity after yohimbine ingestion by a body builder. Clin. Toxicol. 2009, 47, 827–829. [Google Scholar] [CrossRef] [PubMed]
- Sanchez Munoz, M.C.; De Kock, M.; Forget, P. What is the place of clonidine in anesthesia? Systematic review and meta-analyses of randomized controlled trials. J. Clin. Anesth. 2017, 38, 140–153. [Google Scholar] [CrossRef] [PubMed]
- Goyal, S.; Sharma, A.; Goswami, D.; Kothari, N.; Goyal, A.; Vyas, V.; Kirubakaran, R.; Sahu, R.; Singh, S. Clonidine and Morphine as Adjuvants for Caudal Anaesthesia in Children: A Systematic Review and Meta-Analysis of Randomised Controlled Trials. Turk. J. Anaesthesiol. Reanim. 2020, 48, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, X.; Wang, Y.; Zhang, J. Effect of Clonidine on Hemodynamic Responses During Laparoscopic Cholecystectomy: A Systematic Review and Meta-Analysis. Surg. Laparosc. Endosc. Percutan. Tech. 2017, 27, 335–340. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Yu, L.Y.; Zhang, W.S. Clonidine versus other adjuncts added to local anesthetics for pediatric neuraxial blocks: A systematic review and meta-analysis. J. Pain Res. 2018, 11, 1027–1036. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Wang, H.; Zhu, A.; Niu, K.; Wang, G. Meta-analysis of dexmedetomidine on emergence agitation and recovery profiles in children after sevoflurane anesthesia: Different administration and different dosage. PLoS ONE 2015, 10, e0123728. [Google Scholar] [CrossRef] [PubMed]
- Colbert, E.H. Tetrapod Extinctions at the End of the Triassic Period. Proc. Natl. Acad. Sci. USA 1958, 44, 973–977. [Google Scholar] [CrossRef]
- Darmani, N.A. Serotonin 5-HT3 receptor antagonists prevent cisplatin-induced emesis in Cryptotis parva: A new experimental model of emesis. J. Neural. Transm. 1998, 105, 1143–1154. [Google Scholar] [CrossRef]
- Darmani, N.A.; Zhong, W.; Chebolu, S.; Vaezi, M.; Alkam, T. Broad-spectrum antiemetic potential of the L-type calcium channel antagonist nifedipine and evidence for its additive antiemetic interaction with the 5-HT (3) receptor antagonist palonosetron in the least shrew (Cryptotis parva). Eur. J. Pharmacol. 2014, 722, 2–12. [Google Scholar] [CrossRef]
- Zhong, W.; Chebolu, S.; Darmani, N.A. Thapsigargin-induced activation of Ca (2+)-CaMKII-ERK in brainstem contributes to substance P release and induction of emesis in the least shrew. Neuropharmacology 2016, 103, 195–210. [Google Scholar] [CrossRef] [PubMed]
- Alkam, T.; Chebolu, S.; Darmani, N.A. Cyclophosphamide causes activation of protein kinase A (PKA) in the brainstem of vomiting least shrews (Cryptotis parva). Eur. J. Pharmacol. 2014, 722, 156–164. [Google Scholar] [CrossRef]
- Zhong, W.; Darmani, N.A. The HCN Channel Blocker ZD7288 Induces Emesis in the Least Shrew (Cryptotis parva). Front. Pharmacol. 2021, 12, 647021. [Google Scholar] [CrossRef]
- Andrews, P.L.; Naylor, R.J.; Joss, R.A. Neuropharmacology of emesis and its relevance to anti-emetic therapy. Consensus and controversies. Support. Care Cancer 1998, 6, 197–203. [Google Scholar] [CrossRef]
- Zhong, W.; Chebolu, S.; Darmani, N.A. Intracellular emetic signaling evoked by the L-type Ca(2+) channel agonist FPL64176 in the least shrew (Cryptotis parva). Eur. J. Pharmacol. 2018, 834, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Haller, J.; Barna, I.; Kovacs, J.L. Alpha 2-adrenoceptor blockade, pituitary-adrenal hormones, and agonistic interactions in rats. Psychopharmacology 1994, 115, 478–484. [Google Scholar] [CrossRef] [PubMed]
- Cornil, C.A.; Ball, G.F. Interplay among catecholamine systems: Dopamine binds to alpha2-adrenergic receptors in birds and mammals. J. Comp. Neurol. 2008, 511, 610–627. [Google Scholar] [CrossRef]
- Millan, M.J.; Newman-Tancredi, A.; Audinot, V.; Cussac, D.; Lejeune, F.; Nicolas, J.P.; Coge, F.; Galizzi, J.P.; Boutin, J.A.; Rivet, J.M.; et al. Agonist and antagonist actions of yohimbine as compared to fluparoxan at alpha(2)-adrenergic receptors (AR)s, serotonin (5-HT)(1A), 5-HT(1B), 5-HT(1D) and dopamine D(2) and D(3) receptors. Significance for the modulation of frontocortical monoaminergic transmission and depressive states. Synapse 2000, 35, 79–95. [Google Scholar] [PubMed]
- Uhlen, S.; Dambrova, M.; Nasman, J.; Schioth, H.B.; Gu, Y.; Wikberg-Matsson, A.; Wikberg, J.E. [3H]RS79948-197 binding to human, rat, guinea pig and pig alpha2A-, alpha2B- and alpha2C-adrenoceptors. Comparison with MK912, RX821002, rauwolscine and yohimbine. Eur. J. Pharmacol. 1998, 343, 93–101. [Google Scholar] [CrossRef]
- Brannan, T.; Martinez-Tica, J.; Yahr, M.D. Effect of yohimbine on brain monoamines: An in vivo study. J. Neural. Transm. Park. Dis. Dement. Sect. 1991, 3, 81–87. [Google Scholar] [CrossRef]
- Dunbar, J.C.; Clough-Helfman, C.; Barraco, R.A.; Anderson, G.F. Effect of insulin and clonidine on the evoked release of norepinephrine and serotonin from the nucleus tractus solitarius of the diabetic rat. Pharmacology 1995, 51, 370–380. [Google Scholar] [CrossRef] [PubMed]
- Bullitt, E. Expression of c-fos-like protein as a marker for neuronal activity following noxious stimulation in the rat. J. Comp. Neurol. 1990, 296, 517–530. [Google Scholar] [CrossRef] [PubMed]
- Ray, A.P.; Chebolu, S.; Darmani, N.A. Receptor-selective agonists induce emesis and Fos expression in the brain and enteric nervous system of the least shrew (Cryptotis parva). Pharmacol. Biochem. Behav. 2009, 94, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Gershon, M.D.; Tack, J. The serotonin signaling system: From basic understanding to drug development for functional GI disorders. Gastroenterology 2007, 132, 397–414. [Google Scholar] [CrossRef] [PubMed]
- Johnston, K.D.; Lu, Z.; Rudd, J.A. Looking beyond 5-HT (3) receptors: A review of the wider role of serotonin in the pharmacology of nausea and vomiting. Eur. J. Pharmacol. 2014, 722, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Langer, S.Z. Presynaptic regulation of the release of catecholamines. Pharmacol. Rev. 1980, 32, 337–362. [Google Scholar] [CrossRef] [PubMed]
- Westfall, T.C. Neuroeffector mechanisms. Annu. Rev. Physiol. 1980, 42, 383–397. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.H.; Costall, B.; Ge, J.; Naylor, R.J. The profiles of interaction of yohimbine with anxiolytic and putative anxiolytic agents to modify 5-HT release in the frontal cortex of freely-moving rats. Br. J. Pharmacol. 1993, 110, 1079–1084. [Google Scholar] [CrossRef]
- Ellison, D.W.; Campbell, I.C. Studies on the role of alpha 2-adrenoceptors in the control of synaptosomal [3H]5-hydroxytryptamine release: Effects of antidepressant drugs. J. Neurochem. 1986, 46, 218–223. [Google Scholar] [CrossRef]
- Takano, M.; Takano, Y.; Yaksh, T.L. Release of calcitonin gene-related peptide (CGRP), substance P (SP), and vasoactive intestinal polypeptide (VIP) from rat spinal cord: Modulation by alpha 2 agonists. Peptides 1993, 14, 371–378. [Google Scholar] [CrossRef]
- Kuraishi, Y.; Hirota, N.; Sato, Y.; Kaneko, S.; Satoh, M.; Takagi, H. Noradrenergic inhibition of the release of substance P from the primary afferents in the rabbit spinal dorsal horn. Brain Res. 1985, 359, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, H.; Koga, T. Non-respiratory neurons in the Botzinger complex exhibiting appropriate firing patterns to generate the emetic act in dogs. Neurosci. Res. 1992, 14, 180–194. [Google Scholar] [CrossRef] [PubMed]
- Cruickshank, M.; Henderson, L.; MacLennan, G.; Fraser, C.; Campbell, M.; Blackwood, B.; Gordon, A.; Brazzelli, M. Alpha-2 agonists for sedation of mechanically ventilated adults in intensive care units: A systematic review. Health Technol. Assess. 2016, 20, v–xx, 1–117. [Google Scholar] [CrossRef] [PubMed]
- Gavino, L.; Willaredt, M.; Chiche, J.D.; Ben-Hamouda, N. Alpha-2 adrenoreceptor agonists for the intensive care physician. Rev. Med. Suisse 2023, 19, 872–877. [Google Scholar] [PubMed]
- Durcan, M.J.; Lister, R.G.; Linnoila, M. Interactions of alpha 2-adrenoceptor antagonists with medetomidine and with ethanol in a holeboard test. Neuropharmacology 1989, 28, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Malinge, M.; Colombel, M.C.; Bourin, M. Mechanism of action of clonidine in the forced-swimming test in mice. Encephale 1989, 15, 37–41. [Google Scholar] [PubMed]
- Smith, D.F. Lithium attenuates clonidine-induced hypoactivity: Further studies in inbred mouse strains. Psychopharmacology 1988, 94, 428–430. [Google Scholar] [CrossRef] [PubMed]
- Hunter, J.C.; Fontana, D.J.; Hedley, L.R.; Jasper, J.R.; Lewis, R.; Link, R.E.; Secchi, R.; Sutton, J.; Eglen, R.M. Assessment of the role of alpha2-adrenoceptor subtypes in the antinociceptive, sedative and hypothermic action of dexmedetomidine in transgenic mice. Br. J. Pharmacol. 1997, 122, 1339–1344. [Google Scholar] [CrossRef]
- Houston, M.C. Clonidine hydrochloride: Review of pharmacologic and clinical aspects. Prog. Cardiovasc. Dis. 1981, 23, 337–350. [Google Scholar] [CrossRef]
- Kumar, A.; Bose, S.; Bhattacharya, A.; Tandon, O.P.; Kundra, P. Oral clonidine premedication for elderly patients undergoing intraocular surgery. Acta Anaesthesiol. Scand. 1992, 36, 159–164. [Google Scholar] [CrossRef]
- Ramadhyani, U.; Park, J.L.; Carollo, D.S.; Waterman, R.S.; Nossaman, B.D. Dexmedetomidine: Clinical application as an adjunct for intravenous regional anesthesia. Anesthesiol. Clin. 2010, 28, 709–722. [Google Scholar] [CrossRef]
- Darmani, N.A.; Chebolu, S.; Amos, B.; Alkam, T. Synergistic antiemetic interactions between serotonergic 5-HT3 and tachykininergic NK1-receptor antagonists in the least shrew (Cryptotis parva). Pharmacol. Biochem. Behav. 2011, 99, 573–579. [Google Scholar] [CrossRef] [PubMed]
- Zhong, W.; Hutchinson, T.E.; Chebolu, S.; Darmani, N.A. Serotonin 5-HT3 receptor-mediated vomiting occurs via the activation of Ca2+/CaMKII-dependent ERK1/2 signaling in the least shrew (Cryptotis parva). PLoS ONE 2014, 9, e104718. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Guijo, J.M.; Carabelli, V.; Gandia, L.; Garcia, A.G.; Carbone, E. Voltage-independent autocrine modulation of L-type channels mediated by ATP, opioids and catecholamines in rat chromaffin cells. Eur. J. Neurosci. 1999, 11, 3574–3584. [Google Scholar] [CrossRef] [PubMed]
- Kleppisch, T.; Ahnert-Hilger, G.; Gollasch, M.; Spicher, K.; Hescheler, J.; Schultz, G.; Rosenthal, W. Inhibition of voltage-dependent Ca2+ channels via alpha 2-adrenergic and opioid receptors in cultured bovine adrenal chromaffin cells. Pflugers Arch. 1992, 421, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Darmani, N.A.; Wang, Y.; Abad, J.; Ray, A.P.; Thrush, G.R.; Ramirez, J. Utilization of the least shrew as a rapid and selective screening model for the antiemetic potential and brain penetration of substance P and NK1 receptor antagonists. Brain Res. 2008, 1214, 58–72. [Google Scholar] [CrossRef] [PubMed]
- Zhong, W.; Chebolu, S.; Darmani, N.A. Intracellular emetic signaling cascades by which the selective neurokinin type 1 receptor (NK(1)R) agonist GR73632 evokes vomiting in the least shrew (Cryptotis parva). Neurochem. Int. 2019, 122, 106–119. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.R.; Kao, P.C.; Chan, M.H. Involvement of Ca2+ signaling in tachykinin-mediated contractile responses in swine trachea. J. Biomed. Sci. 2005, 12, 547–558. [Google Scholar] [CrossRef]
- Miyano, K.; Morioka, N.; Sugimoto, T.; Shiraishi, S.; Uezono, Y.; Nakata, Y. Activation of the neurokinin-1 receptor in rat spinal astrocytes induces Ca2+ release from IP3-sensitive Ca2+ stores and extracellular Ca2+ influx through TRPC3. Neurochem. Int. 2010, 57, 923–934. [Google Scholar] [CrossRef]
- Suzuki, Y.; Inoue, T.; Ra, C. L-type Ca2+ channels: A new player in the regulation of Ca2+ signaling, cell activation and cell survival in immune cells. Mol. Immunol. 2010, 47, 640–648. [Google Scholar] [CrossRef]
- Sculptoreanu, A.; Yoshimura, N.; de Groat, W.C.; Somogyi, G.T. Protein kinase C is involved in M1-muscarinic receptor-mediated facilitation of L-type Ca2+ channels in neurons of the major pelvic ganglion of the adult male rat. Neurochem. Res. 2001, 26, 933–942. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, L.; Correia-de-Sa, P. Protein kinase A and Ca(v)1 (L-Type) channels are common targets to facilitatory adenosine A2A and muscarinic M1 receptors on rat motoneurons. Neurosignals 2005, 14, 262–272. [Google Scholar] [CrossRef] [PubMed]
- Zhong, W.; Chebolu, S.; Darmani, N.A. Broad-spectrum antiemetic efficacy of the L-type calcium channel blocker amlodipine in the least shrew (Cryptotis parva). Pharmacol. Biochem. Behav. 2014, 120, 124–132. [Google Scholar] [CrossRef]
- Darmani, N.A.; Zhao, W.; Ahmad, B. The role of D2 and D3 dopamine receptors in the mediation of emesis in Cryptotis parva (the least shrew). J. Neural. Transm. 1999, 106, 1045–1061. [Google Scholar] [CrossRef] [PubMed]
- Belkacemi, L.; Zhong, W.; Darmani, N.A. Signal transduction pathways involved in dopamine D(2) receptor-evoked emesis in the least shrew (Cryptotis parva). Auton. Neurosci. 2021, 233, 102807. [Google Scholar] [CrossRef] [PubMed]
- Xing, X.W.; Sun, Y.F.; Zhao, J.; Pan, Z.X.; Jiang, W.X. Tizanidine hydrochloride exhibits a cytotoxic effect on osteosarcoma cells through the PI3K/AKT signaling pathway. J. Int. Med. Res. 2019, 47, 3792–3802. [Google Scholar] [CrossRef]
- Darmani, N.A. Delta (9)-tetrahydrocannabinol and synthetic cannabinoids prevent emesis produced by the cannabinoid CB (1) receptor antagonist/inverse agonist SR 141716A. Neuropsychopharmacology 2001, 24, 198–203. [Google Scholar] [CrossRef] [PubMed]
- Tzavara, E.T.; Davis, R.J.; Perry, K.W.; Li, X.; Salhoff, C.; Bymaster, F.P.; Witkin, J.M.; Nomikos, G.G. The CB1 receptor antagonist SR141716A selectively increases monoaminergic neurotransmission in the medial prefrontal cortex: Implications for therapeutic actions. Br. J. Pharmacol. 2003, 138, 544–553. [Google Scholar] [CrossRef] [PubMed]
- Lever, I.J.; Malcangio, M. CB (1) receptor antagonist SR141716A increases capsaicin-evoked release of Substance P from the adult mouse spinal cord. Br. J. Pharmacol. 2002, 135, 21–24. [Google Scholar] [CrossRef]
- Zamponi, G.W.; Striessnig, J.; Koschak, A.; Dolphin, A.C. The Physiology, Pathology, and Pharmacology of Voltage-Gated Calcium Channels and Their Future Therapeutic Potential. Pharmacol. Rev. 2015, 67, 821–870. [Google Scholar] [CrossRef]
- Garaschuk, O.; Yaari, Y.; Konnerth, A. Release and sequestration of calcium by ryanodine-sensitive stores in rat hippocampal neurones. J. Physiol. 1997, 502 Pt 1, 13–30. [Google Scholar] [CrossRef]
- Gomez-Viquez, N.L.; Guerrero-Serna, G.; Arvizu, F.; Garcia, U.; Guerrero-Hernandez, A. Inhibition of SERCA pumps induces desynchronized RyR activation in overloaded internal Ca2+ stores in smooth muscle cells. Am. J. Physiol. Cell Physiol. 2010, 298, C1038–C1046. [Google Scholar] [CrossRef]
- Beltran-Parrazal, L.; Fernandez-Ruiz, J.; Toledo, R.; Manzo, J.; Morgado-Valle, C. Inhibition of endoplasmic reticulum Ca2+ ATPase in preBotzinger complex of neonatal rat does not affect respiratory rhythm generation. Neuroscience 2012, 224, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Michelangeli, F.; East, J.M. A diversity of SERCA Ca2+ pump inhibitors. Biochem. Soc. Trans. 2011, 39, 789–797. [Google Scholar] [CrossRef] [PubMed]
- Solovyova, N.; Verkhratsky, A. Neuronal endoplasmic reticulum acts as a single functional Ca2+ store shared by ryanodine and inositol-1,4,5-trisphosphate receptors as revealed by intra-ER [Ca2+] recordings in single rat sensory neurones. Pflugers Arch. 2003, 446, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Richter, W.; Menniti, F.S.; Zhang, H.T.; Conti, M. PDE4 as a target for cognition enhancement. Expert. Opin. Ther. Targets 2013, 17, 1011–1027. [Google Scholar] [CrossRef] [PubMed]
- Propper, D.J.; Saunders, M.P.; Salisbury, A.J.; Long, L.; O’Byrne, K.J.; Braybrooke, J.P.; Dowsett, M.; Taylor, M.; Talbot, D.C.; Ganesan, T.S.; et al. Phase I study of the novel cyclic AMP (cAMP) analogue 8-chloro-cAMP in patients with cancer: Toxicity, hormonal, and immunological effects. Clin. Cancer Res. 1999, 5, 1682–1689. [Google Scholar] [PubMed]
- Benarroch, E.E. HCN channels: Function and clinical implications. Neurology 2013, 80, 304–310. [Google Scholar] [CrossRef]
- McGovern, A.E.; Robusto, J.; Rakoczy, J.; Simmons, D.G.; Phipps, S.; Mazzone, S.B. The effect of hyperpolarization-activated cyclic nucleotide-gated ion channel inhibitors on the vagal control of guinea pig airway smooth muscle tone. Br. J. Pharmacol. 2014, 171, 3633–3650. [Google Scholar] [CrossRef]
- Gill, C.H.; Randall, A.; Bates, S.A.; Hill, K.; Owen, D.; Larkman, P.M.; Cairns, W.; Yusaf, S.P.; Murdock, P.R.; Strijbos, P.J.; et al. Characterization of the human HCN1 channel and its inhibition by capsazepine. Br. J. Pharmacol. 2004, 143, 411–421. [Google Scholar] [CrossRef]
- Shinpo, K.; Hirai, Y.; Maezawa, H.; Totsuka, Y.; Funahashi, M. The role of area postrema neurons expressing H-channels in the induction mechanism of nausea and vomiting. Physiol. Behav. 2012, 107, 98–103. [Google Scholar] [CrossRef] [PubMed]
- Docherty, J.R. Subtypes of functional alpha1- and alpha2-adrenoceptors. Eur. J. Pharmacol. 1998, 361, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.C.; Meng, Q.T.; Pan, X.; Xia, Z.Y.; Chen, X.D. Dexmedetomidine produced analgesic effect via inhibition of HCN currents. Eur. J. Pharmacol. 2014, 740, 560–564. [Google Scholar] [CrossRef] [PubMed]
- Vulliemoz, Y.; Shen, H.; Virag, L. Alpha-2 adrenoceptor agonists decrease cyclic guanosine 3′,5′-monophosphate in the mouse brain. Anesthesiology 1996, 85, 544–550. [Google Scholar] [CrossRef] [PubMed]
- Glaess, S.S.; Attridge, R.L.; Christina Gutierrez, G. Clonidine as a strategy for discontinuing dexmedetomidine sedation in critically ill patients: A narrative review. Am. J. Health Syst. Pharm. 2020, 77, 515–522. [Google Scholar] [CrossRef]
- Keating, G.M. Dexmedetomidine: A Review of Its Use for Sedation in the Intensive Care Setting. Drugs 2015, 75, 1119–1130. [Google Scholar] [CrossRef]
- Nguyen, V.; Tiemann, D.; Park, E.; Salehi, A. Alpha-2 Agonists. Anesthesiol. Clin. 2017, 35, 233–245. [Google Scholar] [CrossRef]
- Belkacemi, L.; Sun, Y.; Darmani, N.A. Evidence for Bell-Shaped Dose-Response Emetic Effects of Temsirolimus and Analogs: The Broad-Spectrum Antiemetic Efficacy of a Large Dose of Temsirolimus Against Diverse Emetogens in the Least Shrew (Cryptotis parva). Front. Pharmacol. 2022, 13, 848673. [Google Scholar] [CrossRef]
- Chow, S.-C.; Wang, H.; Shao, J. Sample Size Calculations in Clinical Research, 2nd ed.; Chapman and Hall/CRC: New York, NY, USA, 2007; p. 480. ISBN 9780429144639. [Google Scholar]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Y.; Darmani, N.A. A Comparative Study of the Antiemetic Effects of α2-Adrenergic Receptor Agonists Clonidine and Dexmedetomidine against Diverse Emetogens in the Least Shrew (Cryptotis parva) Model of Emesis. Int. J. Mol. Sci. 2024, 25, 4603. https://doi.org/10.3390/ijms25094603
Sun Y, Darmani NA. A Comparative Study of the Antiemetic Effects of α2-Adrenergic Receptor Agonists Clonidine and Dexmedetomidine against Diverse Emetogens in the Least Shrew (Cryptotis parva) Model of Emesis. International Journal of Molecular Sciences. 2024; 25(9):4603. https://doi.org/10.3390/ijms25094603
Chicago/Turabian StyleSun, Yina, and Nissar A. Darmani. 2024. "A Comparative Study of the Antiemetic Effects of α2-Adrenergic Receptor Agonists Clonidine and Dexmedetomidine against Diverse Emetogens in the Least Shrew (Cryptotis parva) Model of Emesis" International Journal of Molecular Sciences 25, no. 9: 4603. https://doi.org/10.3390/ijms25094603