
Biological Roles and Clinical Applications of Exosomes in Breast Cancer: A Brief Review

Department of General Surgery, Comprehensive Breast Health Center, Ruijin Hospital, Shanghai Jiao Tong University School of Medicine, Shanghai 200025, China
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally.
Int. J. Mol. Sci. 2024, 25(9), 4620; https://doi.org/10.3390/ijms25094620
Submission received: 14 March 2024
/
Revised: 20 April 2024
/
Accepted: 22 April 2024
/
Published: 24 April 2024
(This article belongs to the Special Issue Molecular Research and Treatment of Breast Cancer 3.0)
Abstract
:Breast cancer (BC) is a global health risk for women and has a high prevalence rate. The drug resistance, recurrence, and metastasis of BC affect patient prognosis, thus posing a challenge to scientists. Exosomes are extracellular vesicles (EVs) that originate from various cells; they have a double-layered lipid membrane structure and contain rich biological information. They mediate intercellular communication and have pivotal roles in tumor development, progression, and metastasis and drug resistance. Exosomes are important cell communication mediators in the tumor microenvironment (TME). Exosomes are utilized as diagnostic and prognostic biomarkers for estimating the treatment efficacy of BC and have the potential to function as tools to enable the targeted delivery of antitumor drugs. This review introduces recent progress in research on how exosomes influence tumor development and the TME. We also present the research progress on the application of exosomes as prognostic and diagnostic biomarkers and drug delivery tools.
1. Introduction
Breast cancer (BC), one of the most commonly detected malignant tumors in women, is a major cause of cancer-associated deaths globally [1,2,3,4]. BC exhibits high heterogeneity and is classified into several subtypes according to genetic and clinical characteristics, including luminal A, luminal B, triple-negative breast cancer (TNBC), and HER2 enrichment [5,6,7,8,9]. Despite advances in the early detection of BC and the availability of effective treatment methods, including radiotherapy, mastectomy, and chemotherapy, BC has been reported to show high metastasis, invasion, and recurrence rates and high drug resistance [10,11,12,13,14,15]. Poor prognosis due to distant organ metastasis and treatment failure are the major hindrances to successfully treating patients with advanced-stage BC [16,17,18]. Hence, the key molecular mechanisms underlying the poor prognosis of BC patients need to be clarified.
Exosomes are cell-synthesized lipid bilayer vesicles [19], with a 30–150 nm particle size [20]. Exosome biogenesis and secretion into the extracellular space involve initiation, endocytosis, multivesicular body formation, and final release [21]. Exosomes are commonly detected in several body fluids, including urine, blood, saliva, amniotic fluids, and breast milk, under normal physiological and pathological conditions [22,23]. A critical function of exosomes is the transport of various functional components, such as lipids, proteins, mRNAs, microRNAs (miRNAs), noncoding RNAs (ncRNAs), and DNA fragments, to the extracellular space [24]. These exosomal components induce phenotypic changes in receptor cells and mediate intercellular communication locally and globally in the body [25,26,27]. Thus, by affecting adjacent or distant cells, exosomes participate in regulating immune function, antigen presentation, cell proliferation, angiogenesis, and tumor cell invasion and migration [28,29,30]. Extracellular vesicles (EVs) are critically involved in the immune suppression and chemotherapeutic resistance of BC cells [31,32]. Because of the rich biological information available on this topic, exosomes, which are involved in liquid biopsies, have the potential for their utilization as biomarkers for the early diagnosis, efficacy detection, and prognosis prediction of BC patients [33,34,35]. Additionally, because of the unique structure of exosomes, drugs delivered through exosomes have better biocompatibility than other delivery methods, thus making them a potential treatment option [36].
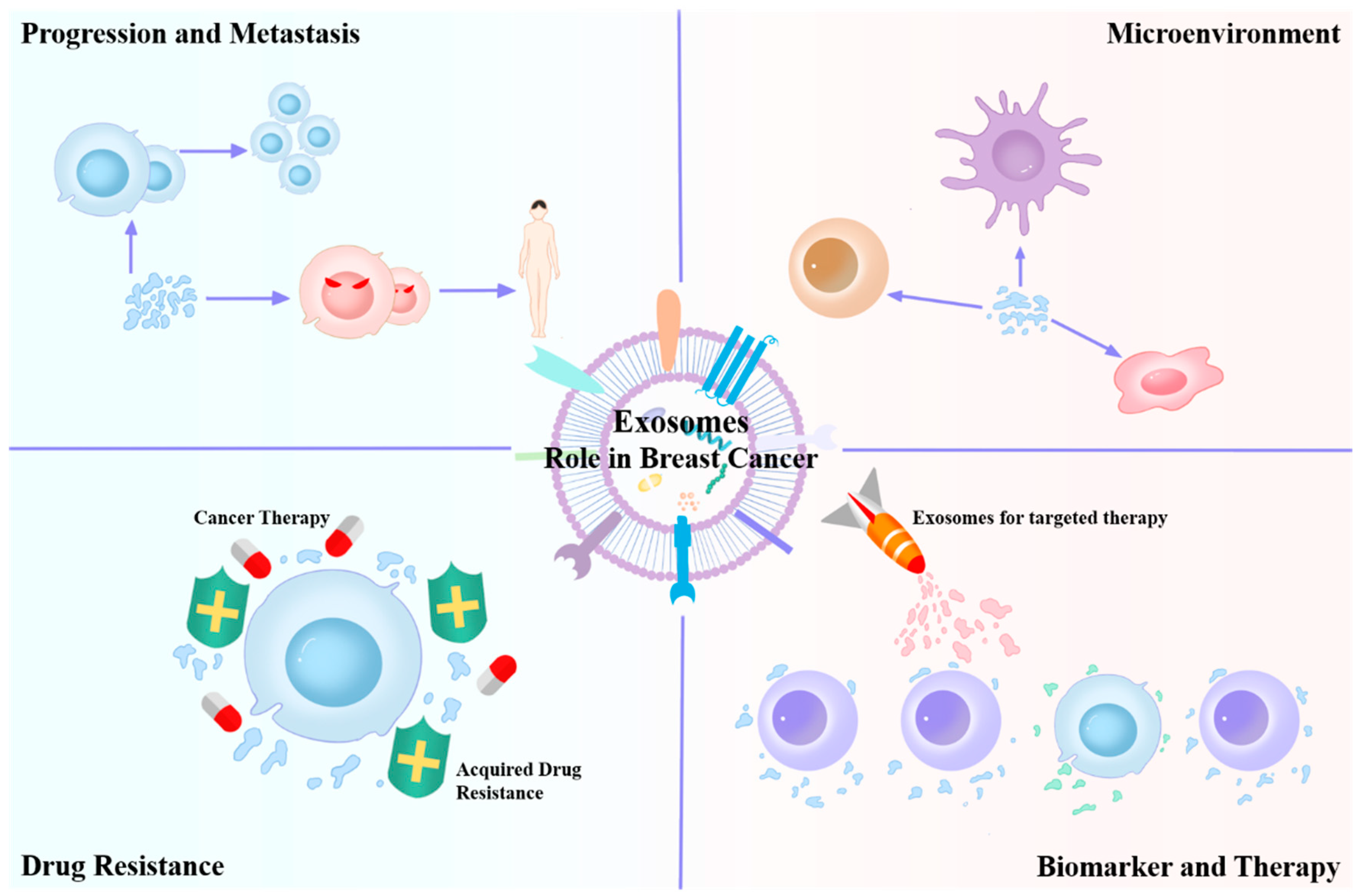
This paper presents a brief review on the research on the impact of exosomes (Figure 1) on tumor development and the tumor microenvironment (TME). We also review recent findings on the application of exosomes as prognostic and diagnostic biomarkers and drug delivery tools. The literature review method can be found in the Supplementary Materials, Table S1.
2. Cancer-Derived Exosomes Can Advance Tumor Progression and Metastasis
Exosomes exhibit pleiotropic roles in BC metastasis and progression, including angiogenesis, invasion, and immune regulation. According to Deng et al., BC cell-derived exosomes can enhance the in vivo metastasis of BC, activate focal adhesion kinase/Src-dependent proteins, and increase proinflammatory cytokine and MMP9 production [37]. Kong et al. detected miR-130a-3p downregulation in BC tissues and circulating exosomes; they also observed an association between the decreased exosome-derived miR-130a-3p level and lymph node (LN) metastasis and advanced TNM stage [38]. According to in vitro and in vivo experiments, exosomal miR-222 contributes to BC cell tumorigenicity and metastasis, possibly through the downregulation of the tumor suppressor genes PDZ and PDLIM2 as well as the subsequent activation of NF-kB [39]. BC cell-derived exosomes are enriched in miR-1910-3p, which can be transferred to BC and breast epithelial cells through MTMR3 regulation and NF-κB signaling pathway activation, thereby facilitating BC cell growth and metastasis [40]. Pan et al. reported that exosomal miR-145 derived from MDA-MB-231 cells targets IRS1 and inhibits HUVEC angiogenesis by controlling the IRS1/Raf/ERK pathways and IRS1/PI3K/Akt/mTOR [41]. TNBC cell-derived exosomal miR-939 enhanced the cross-endothelial migration of tumor cells and targeted vascular endothelial cadherin directly in endothelial cells. Exosomal miR-939 from BC cells participates in extracellular tumor-promoting features and shows an association with the poor prognosis of TNBC patients [42]. A close interaction of the TME with cancer cells is necessary for metastatic potential development. Previous research has shown that cancer-derived exosomes deliver proteins, lipids, and ncRNAs to surrounding cells or cells present in distant metastatic niches through premetastatic TME regulation. As confirmed by Wang et al., exosomal miR-4443 overexpression strongly enhanced liver metastasis in a mouse BC metastasis model. Exosomal miR-4443 delivery can disrupt the natural barrier, accompanied by damage to TIMP2, leading to MMP-2 upregulation in the liver metastatic lesion and the primary tumor, resulting in transplantation into a new microenvironment [43]. In a mouse lung metastasis model, exosomal miR-183-5p inhibited the transfer of PPP1CA to macrophages, thereby promoting IL-2 and TNF-α (proinflammatory cytokines) secretion [44]. The exosomal AnxA2 level in malignant cells was significantly greater than that in normal cells as well as premetastatic BC cells. Additionally, an in vivo analysis revealed that depletion of the EV AnxA2 reduced the number of metastatic sites in the lungs and brain, which confirmed the critical role of AnxA2 in forming a TME that favors metastasis [45].
Table 1 and Figure 2 depict an overview of exosome progression and metastasis in BC. In fact, although a considerable extent of research focuses on how exosomes promote breast cancer progression or metastasis, not all exosomes are harmful, and miR-145 is a typical example. Therefore, subsequent research directions should focus on the study of exosomes, which is beneficial for treating diseases to develop better tools for BC treatment. We also noticed that the role of exosomes seems to not be limited to the original tumor itself but also affects other organs. This effect is currently suggested to be detrimental to patients in the current study, but could it be beneficial? Therefore, this reminds us of the necessity of systematic research in in vivo studies. It is worth noting that although some studies have suggested proteomic differences in exosomes, a large number of studies have focused on differences in exosome content rather than exosome quantity [46,47]. This may suggest potential future research possibilities.
3. Roles of Cancer-Derived Exosomes in the BC Microenvironment
BC-derived exosomes contain rich biological information and have substantial involvement in the cellular communication and biological regulation of tumor cell interactions with the surrounding microenvironment [48,49]. Following induction by BC-derived exosomes, cells residing in the BC microenvironment may influence the biological characteristics of BC through various ways.
Fibroblasts exhibit high abundance in the typical TME [50]. They are spindle-shaped and secrete various extracellular matrix (ECM) proteins that maintain the connective tissue framework and assist in wound healing [51,52]. In the TME, fibroblasts are significantly involved in tumor cell interactions with surrounding cells [53]. Cancer-associated fibroblasts (CAFs) are a crucial cell subtype involved in the coevolution of BC cells and their microenvironment through cellular compartment modification and stromal–epithelial interaction-mediated cancer cell regulations [54]. Tumor cell-derived exosomes can transform normal fibroblasts (NFs) to CAFs. According to previous studies, exosomal miR-9, miR-125b, etc., from BC cells, can induce fibroblasts to produce CAF-like characteristics [55,56]. miR-105 exerts a protumor effect by mediating stromal cell metabolism reprogramming. miR-105 also mediates the metabolic reprogramming of CAFs, thereby providing energy to neighboring cancer cells through the enhancement of glucose and glutamine metabolism under nutrient-rich conditions. Under nutrient-deficient conditions, metabolic waste, such as lactic acid, is converted into energy to support tumor growth [57]. Ahn et al. noted that luminal A cancer cell-derived exosomes delivered miR-130b-3p to CAFs and promoted BC progression by downregulating SPIN90 [58]. Wang et al. revealed that, in BC lung metastasis, tumor cells transfer Cav-1 to the metastatic organ microenvironment through exosomes, which promotes tenascin-C secretion in lung fibroblasts and induces ECM deposition. It also inhibited the PTEN/CCL2/VEGF-A signaling pathway in lung macrophages and promoted angiogenesis and M2-type macrophage polarization, thereby mediating intercellular communication and regulating the premetastatic niche of lung metastasis [59]. BC cells show a high expression of survivin, which is delivered to fibroblasts through exosomes to upregulate SOD-1 expression in CAFs; this provides a feedback to tumor cells, resulting in the enhancement of their metastasis and proliferation [60].
Tumor-associated macrophages (TAMs) are an important group of immune response-associated cells in the BC microenvironment that have recently received considerable attention [61,62]. Macrophages can transform from a tumor-suppressing state to a tumor-promoting state through M1 to M2 polarization [63,64,65]. BC cell-derived exosomes can influence the microenvironment by affecting the polarization process. According to in vitro studies, BC cell-derived exosomes can transfer GP130 to macrophages and alter their polarization through STAT3 pathway activation [66]. miR-138-5p, from BC cells, is delivered to TAMs through exosomes and can decrease KDM6B expression, suppress M1 polarization, activate M2 polarization, and promote lung metastasis in BC patients [67]. Lin28B facilitates the lung metastasis of BC cells. It shows an upregulated expression in the exosomes of TNBC patients and facilitates neutrophil recruitment and M2 polarization, which is critical for PD-L1 upregulation in lungs and for immunosuppression due to a cytokine imbalance [68]. The tumor exosomal circRNA cSERPINE2 is significantly elevated in BC and controls the MALT1-NF-κB-IL-6 axis in TAMs; it also increases IL-6 secretion that facilitates BC cell proliferation and invasion [69]. Endoplasmic reticulum (ER) stress stimulates exosomal Circ_0001142 release from BC cells, which regulates tumor progression by triggering M2 macrophage polarization through the Circ_0001142/miR-361-3p/PIK3CB pathway [70]. ER stress also induces an increase in miR-27a-3p, which is then transferred to macrophages through exosomes and induces the MAGI2/PTEN/PI3K axis-mediated PD-L1 upregulation in TAMs [71]. BC cell-derived exosomes transfer miR-63 to TAMs by binding to the CD375 receptor. miR-375 also directly targets TNS3 and PXN, which increases tumor migration and infiltration by TAMs [72]. In vivo models of BC cell-derived exosomes can also transmit CCL5 to affect immune infiltration in the TME and promote lung metastasis in patients with BC [73].
Cancer-related adipocytes (CAAs) have also recently received a considerable amount of attention from researchers based on the analysis of patient serum-derived exosomes [62]. Zhao et al. noted that BC cell-derived exosomes activate CAA by delivering miR-1304-3p, accelerating adipocyte differentiation and lipid accumulation, targeting the antiadipogenic gene GATA2 to exert a pro-carcinogenic effect, and potentially enabling the prediction of the prognosis of African American patients [74]. BC cell-derived exosomes deliver miR-155 to target UBQLN1 in adipocytes, inhibit white adipose tissue browning, and promote fat reduction related to cancer cachexia [75]. Exosomal miR-122 suppresses the uptake of glucose by premetastatic ecotone cells through the attenuation of pyruvate kinase (a glycolytic enzyme) expression, thus promoting disease progression [76]. In ER-positive BC, exosomal miR-19a is significantly upregulated and is delivered to osteoclasts through IBSP to induce the generation of osteoclasts, thus forming a microenvironment conducive to tumor cell colonization [77]. BC cell-derived exosomes also stimulate bone marrow cell differentiation into MDSCs by downregulating CXCR4 expression and activating the STAT3 signaling pathway, which is closely associated with the drug resistance and poor prognosis of BC patients [78].
Table 2 and Figure 3 show the relevant mechanisms underlying the effect of BC-driven exosomes on the TME. Current research on the roles of cancer-derived exosomes in the BC microenvironment mainly focuses on CAF, TAM, CAA, and other cells. However, the mechanism by which cancer-derived exosomes produce CAA cells is still unclear. In addition, there are a large number of different tumor-related cells in the TME, and the relationship between these cells and roles of cancer-derived exosomes deserves to be explored. Based on current research progress, future research should focus on the following: (1) are there any beneficial effects of BC-driven exosomes on the TME? and (2) can a specific target be developed to block exosomes for therapeutic purposes?
4. Roles of Microenvironment-Derived Exosomes in BC
4.1. Effects of Fibroblast-Derived Exosomes
Exosomal miRNAs are essential mediators of the interaction of tumor cells with CAFs. CAF-derived exosomes containing miR-181d-5p increase the aggressive behavior of BC cells by targeting the CDX2/HOXA5 axis [79]. miR-1b and miR-3-7p are delivered to BC cells from CAFs through exosomes, which enhance tumor cell metastasis and invasion by downregulating TCEAL1 and GLIS232 [80]. The upregulation of miR-18b in CAF-derived exosomes stimulates NF-kB activation through specific binding to TCEAL7, promotes nuclear Snail ectopic activation to induce EMT, and induces cellular metastasis and invasion [81]. CAF-derived exosomes target LATS2 through the miR-92/PD-L1 axis, which enhances BC cell proliferation and migration and impairs tumor-infiltrating immune cell functions [82]. Chen et al. validated miR-500a-5p’s role in CAF-derived exosomes and confirmed that its effect on promoting proliferation was achieved by targeting USP28 [83].
The proteins enriched in exosomes also participate in intercellular communication. Fibroblasts with a low p85α expression can change the TME and promote BC metastasis by the exosome-mediated parasecretion of Wnt10b [84]. ADAM10-rich exosomes derived from CAFs can facilitate cellular motility and activate the RhoA and Notch signaling pathways in BC cells, thus affecting tumor progression [85]. Oxidized ATM promotes the phosphorylation of BNIP3, thus inducing the accumulation of the autophagy-related protein GPR64 in hypoxia CAF-derived exosomes, promoting IL-8 and MMP9 upregulation in BC cells and enhancing cancer cell invasiveness [86].
4.2. Effects of Immune Cell-Derived Exosomes
Macrophages, a highly heterogeneous group of immune cells, exhibit strong plasticity in the TME. As shown previously, TAMs promote BC metastasis, and high-density TAMs are an adverse prognostic factor for BC. Exosomal miR-503-3p generated from macrophages upregulated DACT2, inhibited glycolysis through the Wnt/β-catenin signaling pathway, and promoted oxidative phosphorylation in the mitochondria of BC cells [87]. In vivo experiments revealed that TAM-derived exosomal miR-223-3p can be delivered to 4T1 cells and facilitate lung metastasis in BC patients by targeting Cbx5 [88]. Yu et al. investigated how TAM-derived exosomes influence tumor recurrence and metastasis after chemotherapy. Following the coculturing of THP-1-derived macrophages and apoptotic BC cells, TAM-secreted exosomes enhanced BC cell invasion, migration, and proliferation by activating STAT3 and its target genes CyclinD1, MMP2, and MMP9 [89]. Evidence from other studies also suggests that TAMs facilitate tumor progression [64,90]. TAM-derived exosomes delivered Circ_0020256, which targets miR-432-5p and E2F3 to increase the activity of cholangiocarcinoma cells and promote cholangiocarcinoma development [91]. Prostate cancer (PCa) cells can uptake miR-95 delivered by TAM-derived exosomes, and the binding of miR-95 with its downstream target gene JunB promotes PCa cell invasion, proliferation, and epithelial-mesenchymal transition (EMT); moreover, the high miR-95 expression level is associated with worse clinicopathological features [92].
4.3. Effects of Adipocyte-Derived Exosomes
Compared to that of immune cells and fibroblasts, the role of adipocytes in tumor progression has been less studied. Recent studies have increasingly appreciated the role of adipocytes in the TME. Wang et al. noted that adipocyte-derived exosomes can stimulate BC cell proliferation and migration through the Hippo signaling pathway [93]. Studies on BC patients with type 2 diabetes revealed that CAA-derived exosomes promote the expression of genes associated with cancer stem cell and EMT traits. TSP5 was upregulated in exosomes and related to EMT. COMP co-expression with BRD2 or BRD3 indicated poor patient prognosis [94]. Gernapudi et al. confirmed that preadipocyte-derived exosomes can affect the differentiation, migration, and stemness of BC cells through the miR-140/SOX2/SOX9 axis [95]. Related studies on other cancers confirmed the role of CAA-derived exosomes in tumor progression. Adipocyte-derived exosomes are specifically enriched in proteins involved in fatty acid oxidation (FAO) and can induce the metabolic reprogramming of tumor cells, which is beneficial for FAO to stimulate melanoma cell invasion and migration [96]. Adipocytes increase MMP9 activity in tumor tissues by producing exosomes with a high MMP3 level, thereby promoting lung cancer cell invasion in vivo and in vitro [97].
Table 3 and Figure 4 show the effects of microenvironment-driven exosomes on BC cells. To summarize, in the BC microenvironment, microenvironment-driven exosomes or BC cell-driven exosomes jointly form a closed loop between BC and the microenvironment; however, many studies have analyzed only one aspect. Therefore, future studies should investigate its overall regulatory mechanism.
5. Roles of Exosomes in BC Drug Resistance
5.1. Exosomes in Chemoresistance
Chemotherapy is a critical component of comprehensive BC treatment. Efflux and the inactivation of drugs are the primary mechanisms of drug resistance caused by exosomes in BC. According to previous studies, the chemotherapeutic drug doxorubicin is excreted through vesicles secreted by cells [98]. P-gp is transported by exosomes from docetaxel-resistant BC cells to docetaxel-sensitive cells, thereby inducing acquired docetaxel resistance [99]. Adriamycin-resistant MCF-7 cell-derived exosomes have a rich content of the proteins P-gp and UCH-L1. The treatment of sensitive MCF-57444 cells with LDN-1 (a UCH-L1-specific inhibitor) can prevent EV internalization-induced resistance in adriamycin-resistant MCF-7 cells [100]. Paclitaxel (PTX) treatment induces MDA-MB-231 cells to secrete survivin-rich exosomes and strongly promotes the survival of PTX-treated SK-BR-3 cells and fibroblasts [101]. Exosomes generated from MDA-MB-231 cells resistant to cisplatin show high miR-423-5p expression. Cisplatin-resistant cells are transferred to recipient cells through the promotion of proliferation, metastasis, and antiapoptotic signaling [102]. According to Wang et al., high exosomal lncRNA-H19 levels induced a resistance to doxorubicin in BC cells. Furthermore, lncRNA-H19 inhibition remarkably reduced a resistance to doxorubicin [103]. Exosome miR-1246 promotes a resistance to docetaxel, gemcitabine, and epirubicin through Cyclin-G2 inhibition in BC cells [104].
5.2. Exosomes in Hormone Resistance
Approximately 70% of patients with BC show tumors containing a high level of estrogen receptor-α (ERα); therefore, a hormonal therapy that targets ERα is an effective approach. Semina et al. reported that TAM-resistant MCF-7 cell-derived exosomes caused horizontal hormone resistance in MCF-7 cells with an estrogen dependency. A coculture of sensitive MCF-7 cells for 14 days with exosomes derived from drug-resistant cells caused the sensitive cells to show antiestrogen drug resistance [105]. An exosomal transfer of the lncRNA UCA1 induced ER-MCF-7 cells to exhibit a resistance to TAM through the mTOR signaling pathway [106]. A downregulation of miR-222 increases ERα and p27 expressions at the mRNA and protein levels and restores the sensitivity of cells to TAM; furthermore, exosomal miR-222 may enhance the resistance to TAM by upregulating ERα and p27 in ER-BC cells [107]. Trastuzumab-resistant BC cells secrete exosomes containing SNHG14, which inhibits apoptosis through the Bcl-2-associated pathway and induces trastuzumab resistance [108]. Exosomes containing the AGAP2-AS1 lncRNA also enhance BC cell resistance to trastuzumab [109].
5.3. Resistance of Exosomes to HER2-Targeted Therapy
Targeting HER2 is an important part of the comprehensive treatment of BC; however, targeted resistance is also an important reason for HER2-positive BC progression. HER2-targeted drug resistance is linked with the elevated levels of PD-L1 and TGF-β1. Martinez observed that exosomes transfer these molecules to characterize their cell of origin in drug-sensitive cells. Exosome-derived TGF-β1 levels were related with the response of patients with HER2-related BC to HER2-targeted therapy, thus suggesting the utilization of this cytokine as a treatment response biomarker [110]. The miR-567 expression was downregulated during trastuzumab resistance, and exosomal miR-567 reversed the resistance to trastuzumab by autophagy inhibition; however, miR-567 knockdown induced trastuzumab resistance [111].
Unlike research studies, the clinical treatment approach commonly involves the combination of multiple treatments (e.g., a chemotherapy combination or sequential targeted therapy). Therefore, exosome-related resistance mechanisms are worth exploring in the combination of multiple treatments. The second possible research direction is that, currently, many research investigations focus on cancer-driven exosomes, and a more holistic view is the related role of microenvironment-driven exosomes in drug resistance. In addition, in advanced BC treatments, including fulvestrant, a resistance to CDK4/6 inhibitors cannot be ignored. There may be an exosome-related mechanism, which deserves further studies.
6. Potential Clinical Applications of Exosomes in Treating BC
6.1. Exosomes as Potential Biomarkers of BC
Exosomes contain rich biological information, and exosome testing is an effective method for liquid biopsies with the advantages of noninvasiveness and dynamic monitoring. An increasing number of tumor diagnostic or prognostic markers are being developed. An increase in the concentration of the Hsp70 protein is a characteristic of tumor exosomes. Del-1, fibronectin, 20S proteasome, and spliced survivin are also present in the blood exosomes of patients with BC [112]. Compared to healthy women, BC patients show a lower CD82 level on the circulating exocrine body surface. The low expression of the aforementioned four transmembrane proteins in exosomes is associated with the metastatic characteristics of tumors [113]. Studies of serum-derived exosomes have shown a higher expression of molecules such as CD24, FAK, EGFR, and GPC-1 in BC patients than in healthy individuals; thus, these molecules have the potential to function as diagnostic and prognostic biomarkers [114,115,116]. Several miRNAs in circulating exosomes have emerged as potential biomarkers to enable the early detection of BC and to differentiate BC subtypes. An Au nanoprobe can differentiate BC patients from healthy people by detecting miRNA-1246 [117]. The analysis of the miRNA expression profiles of exosomes revealed the following findings: (1) the different subtypes of BC populations had specific miRNA expression profiles, (2) HER2-positive BC patients and TNBC patients had specific miRNA expression patterns in plasma exosomes, and (3) the miRNA expression was associated with clinicopathological parameters and pCR [118]. Studies have also shown that long RNAs in exosomes can predict clinical outcomes. Among BC patients receiving neoadjuvant therapy, those who achieved pCR showed a differential expression of 2573 exosomal long RNAs as compared to patients with residual lesions. Further analyses revealed that MSMO1 expression levels in exosomes were linked with the clinical outcomes of BC patients [119].
Research on exosome biomarkers is progressing rapidly. However, current traditional exosome extraction and isolation technologies do not seem to meet the clinical requirements for the highly sensitive extraction and isolation of exosomes required for liquid biopsies. Therefore, future research may need to focus on industrialized exosome extraction and isolation methods.
6.2. Exosomes in BC Treatment
Although a variety of treatments are already used for the treatment of cancer, more treatments still need to be developed [120,121,122,123]. Exosomes are independent units with a complete membrane structure. Surface membrane proteins can enhance the endocytosis of receptor cells. Exosomes can be designed to target the delivery of anticancer drugs or other customized molecules without causing toxicity to target cells. PTX-loaded exosomes can be obtained after sufficient incubation with MSCs; these exosomes effectively inhibit cancer cell growth and prevent tumor angiogenesis in bones [124]. Metastatic BC 4T1 cell-derived exosomes can effectively deliver doxorubicin to mice lungs and inhibit lung metastasis [125]. Tumor-derived exosomes can use transcytosis to cross the blood–brain barrier, thus showing that exosomes could be used as drug delivery vehicles to target the brain for treating BC brain metastases [126]. Park et al. designed exosomes that deliver three miRNAs, namely, miR-19a-3p, miR-19b-3p, and miR-1226-3p, which inhibit BC cell migration by targeting AQP5 [127]. α-Lactalbumin-engineered BC exosomes loaded with human neutrophil elastase and hiltonol enhanced immunogenic cell death processes and promoted antitumor immune properties in TNBC [128]. Li et al. modified the exosome surface with an anti-HER2 antibody and an anti-CD3 antibody to target HER2-positive BC cells and activate T-cell immunity [129]. Activated tumor-associated effector T cell-derived exosomes carry membrane-bound PD-1, which enhances T-cell cytotoxicity to TNBC cells by occupying PD-L1 and weakening the subsequent dysfunction of T cells [130]. Engineered macrophage-secreted exosomes were developed, modified, and loaded with DOX using peptides targeting the mesenchymal-epithelial transition factor. These engineered exosomes significantly prolong the DOX circulation time, which specifically targets tumors, promotes apoptosis, and exhibits low hepatotoxicity [131]. Macrophage-tumor chimeric exosomes can be enriched in LNs through a direct interaction with exosomes, effectively leading to tumor regression and extending the survival of a mouse tumor recurrence model when combined with anti-PD-1 therapy [132].
Although exosomes have great promise as a therapeutic measure, there are still many issues that limit their future applications: (1) What kind of cells should be selected as the donor of exosomes? (2) How should the targeting of exosomes be modified? (3) How should drug loading efficiency be improved? (4) How should the engineered production of exosomes be realized?
7. Conclusions and Outlook
Exosomes contribute to tumor progression, TME remodeling, tumor metastasis promotion, and drug resistance and have been studied in depth in BC. Recent investigations have partially elucidated the mechanism through which exosomes participate in intercellular communication and TME remodeling. Active components in exosomes, including miRNAs, proteins, and lipids, are involved in tumorigenesis, which is achieved through the promotion of EMT, angiogenesis, vascular permeability, and metastatic prebiotic regulation. However, because of the complex composition of the TME, which contains multiple cell types and secreted factors, EVs secreted by different cells can synergistically promote tumor occurrence and metastasis. Therefore, it is difficult to determine which cell types of exosomes or cytokines play a major role. Exosomal miRNAs are considered critical mediators of intercellular communication. However, only the differences in the expression levels of these genes in tissue samples have been investigated, and there is a lack of in-depth mechanistic research.
Traditional screening methods for BC include ultrasounds, mammography, MRIs, etc. Compared to traditional methods, a liquid biopsy serves as a noninvasive and rapid detection method. Tumor-derived exosomes widely occur in various body fluids. Exploring exosome biomarkers is helpful for an early BC diagnosis. In addition, EV biomarkers can dynamically reflect disease conditions and predict patient prognosis. However, there is no clear method to determine the molecular subtype from exosomes. Tumor-derived exosomes can also serve as therapeutic targets, and blocking the delivery or release of exosomes or their key components is a promising therapeutic approach. The construction of engineered exosomes used for targeted drug delivery provides a new direction for future personalized therapy.
Supplementary Materials
The supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ijms25094620/s1.
Author Contributions
H.W. and R.W. conceived the review and wrote the main manuscript text and prepared the figures in this manuscript. Z.W., R.H. and K.S. revised the manuscript. All authors reviewed the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (82072897 and 82002773).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| BC | Breast cancer |
| TNBC | Triple-negative breast cancer |
| ncRNAs | Noncoding RNAs |
| miRNAs | MicroRNAs |
| TME | Tumor microenvironment |
| CAFs | Cancer-associated fibroblasts |
| NFs | Normal fibroblasts |
| TAMs | Tumor-associated macrophages |
| CAAs | Cancer-associated adipocytes |
| FAO | Fatty acid oxidation |
| Erα | Estrogen receptor-α |
References
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Parkin, D.M.; Piñeros, M.; Znaor, A.; Bray, F. Cancer statistics for the year 2020: An overview. Int. J. Cancer, 2021; online ahead of print. [Google Scholar] [CrossRef]
- Hong, R.; Xu, B. Breast cancer: An up-to-date review and future perspectives. Cancer Commun. 2022, 42, 913–936. [Google Scholar] [CrossRef] [PubMed]
- Giaquinto, A.N.; Sung, H.; Miller, K.D.; Kramer, J.L.; Newman, L.A.; Minihan, A.; Jemal, A.; Siegel, R.L. Breast Cancer Statistics, 2022. CA Cancer J. Clin. 2022, 72, 524–541. [Google Scholar] [CrossRef] [PubMed]
- Xia, C.; Dong, X.; Li, H.; Cao, M.; Sun, D.; He, S.; Yang, F.; Yan, X.; Zhang, S.; Li, N.; et al. Cancer statistics in China and United States, 2022: Profiles, trends, and determinants. Chin. Med. J. 2022, 135, 584–590. [Google Scholar] [CrossRef] [PubMed]
- Nolan, E.; Lindeman, G.J.; Visvader, J.E. Deciphering breast cancer: From biology to the clinic. Cell 2023, 186, 1708–1728. [Google Scholar] [CrossRef]
- Leon-Ferre, R.A.; Goetz, M.P. Advances in systemic therapies for triple negative breast cancer. BMJ 2023, 381, e071674. [Google Scholar] [CrossRef]
- Cucciniello, L.; Gerratana, L.; Del Mastro, L.; Puglisi, F. Tailoring adjuvant endocrine therapy in early breast cancer: When, how, and how long? Cancer Treat. Rev. 2022, 110, 102445. [Google Scholar] [CrossRef] [PubMed]
- Joseph, C.; Papadaki, A.; Althobiti, M.; Alsaleem, M.; Aleskandarany, M.A.; Rakha, E.A. Breast cancer intratumour heterogeneity: Current status and clinical implications. Histopathology 2018, 73, 717–731. [Google Scholar] [CrossRef] [PubMed]
- Aleskandarany, M.A.; Vandenberghe, M.E.; Marchiò, C.; Ellis, I.O.; Sapino, A.; Rakha, E.A. Tumour Heterogeneity of Breast Cancer: From Morphology to Personalised Medicine. Pathobiol. J. Immunopathol. Mol. Cell. Biol. 2018, 85, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Swain, S.M.; Shastry, M.; Hamilton, E. Targeting HER2-positive breast cancer: Advances and future directions. Nat. Rev. Drug Discov. 2023, 22, 101–126. [Google Scholar] [CrossRef]
- Kumar, H.; Gupta, N.V.; Jain, R.; Madhunapantula, S.V.; Babu, C.S.; Kesharwani, S.S.; Dey, S.; Jain, V. A review of biological targets and therapeutic approaches in the management of triple-negative breast cancer. J. Adv. Res. 2023, 54, 271–292. [Google Scholar] [CrossRef]
- Bianchini, G.; De Angelis, C.; Licata, L.; Gianni, L. Treatment landscape of triple-negative breast cancer—Expanded options, evolving needs. Nat. Rev. Clin. Oncol. 2022, 19, 91–113. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Wang, Z.; Hong, J.; Wu, J.; Huang, O.; He, J.; Chen, W.; Li, Y.; Chen, X.; Shen, K. Targeting cancer-associated adipocyte-derived CXCL8 inhibits triple-negative breast cancer progression and enhances the efficacy of anti-PD-1 immunotherapy. Cell Death Dis. 2023, 14, 703. [Google Scholar] [CrossRef] [PubMed]
- Ye, F.; Dewanjee, S.; Li, Y.; Jha, N.K.; Chen, Z.-S.; Kumar, A.; Vishakha; Behl, T.; Jha, S.K.; Tang, H. Advancements in clinical aspects of targeted therapy and immunotherapy in breast cancer. Mol. Cancer 2023, 22, 105. [Google Scholar] [CrossRef]
- Xu, K.; Liu, Y.; Wang, R.; Cai, P.; Fang, Y. Design, synthesis, and anticancer activities of benzofuran–isatin hybrids tethered by pentylene and hexylene. J. Heterocycl. Chem. 2019, 56, 2052–2055. [Google Scholar] [CrossRef]
- Zhou, C.; Wang, D.; Li, J.; Wang, Q.; Wo, L.; Zhang, X.; Hu, Z.; Wang, Z.; Zhan, M.; He, M. TGFB2-AS1 inhibits triple-negative breast cancer progression via interaction with SMARCA4 and regulating its targets TGFB2 and SOX2. Proc. Natl. Acad. Sci. USA 2022, 119, e2117988119. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wang, H.; Sun, X.; Fang, Y.; Lu, S.S.; Ding, S.N.; Chen, X.S.; Shen, K.W. A Risk Stratification Model for Predicting Overall Survival and Surgical Benefit in Triple-Negative Breast Cancer Patients With de novo Distant Metastasis. Front. Oncol. 2020, 10, 14. [Google Scholar] [CrossRef] [PubMed]
- Harbeck, N.; Gnant, M. Breast cancer. Lancet 2017, 389, 1134–1150. [Google Scholar] [CrossRef] [PubMed]
- Arya, S.B.; Collie, S.P.; Parent, C.A. The ins-and-outs of exosome biogenesis, secretion, and internalization. Trends Cell Biol. 2023, 34, 90–108. [Google Scholar] [CrossRef]
- Alzhrani, G.N.; Alanazi, S.T.; Alsharif, S.Y.; Albalawi, A.M.; Alsharif, A.A.; Abdel-Maksoud, M.S.; Elsherbiny, N. Exosomes: Isolation, characterization, and biomedical applications. Cell Biol. Int. 2021, 45, 1807–1831. [Google Scholar] [CrossRef]
- Xu, M.; Ji, J.; Jin, D.; Wu, Y.; Wu, T.; Lin, R.; Zhu, S.; Jiang, F.; Ji, Y.; Bao, B.; et al. The biogenesis and secretion of exosomes and multivesicular bodies (MVBs): Intercellular shuttles and implications in human diseases. Genes Dis. 2023, 10, 1894–1907. [Google Scholar] [CrossRef]
- De Sousa, K.P.; Rossi, I.; Abdullahi, M.; Ramirez, M.I.; Stratton, D.; Inal, J.M. Isolation and characterization of extracellular vesicles and future directions in diagnosis and therapy. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnology 2023, 15, e1835. [Google Scholar] [CrossRef] [PubMed]
- Mathivanan, S.; Simpson, R.J. ExoCarta: A compendium of exosomal proteins and RNA. Proteomics 2009, 9, 4997–5000. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, K.; Breyne, K.; Ughetto, S.; Laurent, L.C.; Breakefield, X.O. RNA delivery by extracellular vesicles in mammalian cells and its applications. Nat. Rev. Mol. Cell Biol. 2020, 21, 585–606. [Google Scholar] [CrossRef] [PubMed]
- Hánělová, K.; Raudenská, M.; Masařík, M.; Balvan, J. Protein cargo in extracellular vesicles as the key mediator in the progression of cancer. Cell Commun. Signal. CCS 2024, 22, 25. [Google Scholar] [CrossRef] [PubMed]
- Corrado, C.; Raimondo, S.; Chiesi, A.; Ciccia, F.; De Leo, G.; Alessandro, R. Exosomes as intercellular signaling organelles involved in health and disease: Basic science and clinical applications. Int. J. Mol. Sci. 2013, 14, 5338–5366. [Google Scholar] [CrossRef] [PubMed]
- Dixson, A.C.; Dawson, T.R.; Di Vizio, D.; Weaver, A.M. Context-specific regulation of extracellular vesicle biogenesis and cargo selection. Nat. Rev. Mol. Cell Biol. 2023, 24, 454–476. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Yu, D. Exosomes in cancer development, metastasis, and immunity. Biochim. Biophys. Acta. Rev. Cancer 2019, 1871, 455–468. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; He, X.; Mei, C.; Ou, C. Exosome derived from tumor-associated macrophages: Biogenesis, functions, and therapeutic implications in human cancers. Biomark. Res. 2023, 11, 100. [Google Scholar] [CrossRef] [PubMed]
- Nail, H.M.; Chiu, C.C.; Leung, C.H.; Ahmed, M.M.M.; Wang, H.D. Exosomal miRNA-mediated intercellular communications and immunomodulatory effects in tumor microenvironments. J. Biomed. Sci. 2023, 30, 69. [Google Scholar] [CrossRef] [PubMed]
- Das, K.; Paul, S.; Ghosh, A.; Gupta, S.; Mukherjee, T.; Shankar, P.; Sharma, A.; Keshava, S.; Chauhan, S.C.; Kashyap, V.K.; et al. Extracellular Vesicles in Triple-Negative Breast Cancer: Immune Regulation, Biomarkers, and Immunotherapeutic Potential. Cancers 2023, 15, 4879. [Google Scholar] [CrossRef]
- Yang, Q.; Xu, J.; Gu, J.; Shi, H.; Zhang, J.; Zhang, J.; Chen, Z.S.; Fang, X.; Zhu, T.; Zhang, X. Extracellular Vesicles in Cancer Drug Resistance: Roles, Mechanisms, and Implications. Adv. Sci. 2022, 9, e2201609. [Google Scholar] [CrossRef]
- Wang, H.; Wang, R.; Luo, L.; Hong, J.; Chen, X.; Shen, K.; Wang, Y.; Huang, R.; Wang, Z. An exosome-based specific transcriptomic signature for profiling regulation patterns and modifying tumor immune microenvironment infiltration in triple-negative breast cancer. Front. Immunol. 2023, 14, 1295558. [Google Scholar] [CrossRef] [PubMed]
- Aheget, H.; Mazini, L.; Martin, F.; Belqat, B.; Marchal, J.A.; Benabdellah, K. Exosomes: Their Role in Pathogenesis, Diagnosis and Treatment of Diseases. Cancers 2020, 13, 84. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Bai, X.; Ni, J.; Zhang, H.; Duan, W.; Graham, P.; Li, Y. Exosomes and breast cancer drug resistance. Cell Death Dis. 2020, 11, 987. [Google Scholar] [CrossRef] [PubMed]
- Mehryab, F.; Rabbani, S.; Shahhosseini, S.; Shekari, F.; Fatahi, Y.; Baharvand, H.; Haeri, A. Exosomes as a next-generation drug delivery system: An update on drug loading approaches, characterization, and clinical application challenges. Acta Biomater. 2020, 113, 42–62. [Google Scholar] [CrossRef]
- Deng, Z.; Cheng, Z.; Xiang, X.; Yan, J.; Zhuang, X.; Liu, C.; Jiang, H.; Ju, S.; Zhang, L.; Grizzle, W. Tumor cell cross talk with tumor-associated leukocytes leads to induction of tumor exosomal fibronectin and promotes tumor progression. Am. J. Pathol. 2012, 180, 390–398. [Google Scholar] [CrossRef]
- Kong, X.; Zhang, J.; Li, J.; Shao, J.; Fang, L. MiR-130a-3p inhibits migration and invasion by regulating RAB5B in human breast cancer stem cell-like cells. Biochem. Biophys. Res. Commun. 2018, 501, 486–493. [Google Scholar] [CrossRef]
- Ding, J.; Xu, Z.; Zhang, Y.; Tan, C.; Hu, W.; Wang, M.; Xu, Y.; Tang, J. Exosome-mediated miR-222 transferring: An insight into NF-κB-mediated breast cancer metastasis. Exp. Cell. Res. 2018, 369, 129–138. [Google Scholar]
- Wang, B.; Mao, J.-H.; Wang, B.-Y.; Wang, L.-X.; Wen, H.-Y.; Xu, L.-J.; Fu, J.-X.; Yang, H. Exosomal miR-1910-3p promotes proliferation, metastasis, and autophagy of breast cancer cells by targeting MTMR3 and activating the NF-κB signaling pathway. Cancer Lett. 2020, 489, 87–99. [Google Scholar] [CrossRef]
- Pan, S.; Zhao, X.; Shao, C.; Fu, B.; Huang, Y.; Zhang, N.; Dou, X.; Zhang, Z.; Qiu, Y.; Wang, R. STIM1 promotes angiogenesis by reducing exosomal miR-145 in breast cancer MDA-MB-231 cells. Cell Death Dis. 2021, 12, 38. [Google Scholar] [CrossRef]
- Di Modica, M.; Regondi, V.; Sandri, M.; Iorio, M.V.; Zanetti, A.; Tagliabue, E.; Casalini, P.; Triulzi, T. Breast cancer-secreted miR-939 downregulates VE-cadherin and destroys the barrier function of endothelial monolayers. Cancer Lett. 2017, 384, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, Q.; Wang, D.; Yang, S.; Zhou, S.; Xu, H.; Zhang, H.; Zhong, S.; Feng, J. Microenvironment-induced TIMP2 loss by cancer-secreted exosomal miR-4443 promotes liver metastasis of breast cancer. J. Cell. Physiol. 2020, 235, 5722–5735. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Duan, Z.; Zhang, C.; Wang, W.; He, H.; Liu, Y.; Wu, P.; Wang, S.; Song, M.; Chen, H. Mouse 4T1 breast cancer cell–derived exosomes induce proinflammatory cytokine production in macrophages via miR-183. J. Immunol. 2020, 205, 2916–2925. [Google Scholar] [CrossRef] [PubMed]
- Maji, S.; Chaudhary, P.; Akopova, I.; Nguyen, P.M.; Hare, R.J.; Gryczynski, I.; Vishwanatha, J.K. Exosomal annexin II promotes angiogenesis and breast cancer metastasis. Mol. Cancer Res. 2017, 15, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Wen, S.W.; Lima, L.G.; Lobb, R.J.; Norris, E.L.; Hastie, M.L.; Krumeich, S.; Möller, A. Breast Cancer-Derived Exosomes Reflect the Cell-of-Origin Phenotype. Proteomics 2019, 19, e1800180. [Google Scholar] [CrossRef]
- Risha, Y.; Minic, Z.; Ghobadloo, S.M.; Berezovski, M.V. The proteomic analysis of breast cell line exosomes reveals disease patterns and potential biomarkers. Sci. Rep. 2020, 10, 13572. [Google Scholar] [CrossRef] [PubMed]
- Berti, F.C.B.; Tofolo, M.V.; Nunes-Souza, E.; Marchi, R.; Okano, L.M.; Ruthes, M.; Rosolen, D.; Malheiros, D.; Fonseca, A.S.; Cavalli, L.R. Extracellular vesicles-associated miRNAs in triple-negative breast cancer: From tumor biology to clinical relevance. Life Sci. 2024, 336, 122332. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Peng, F.; Chen, J. The Role of Exosomal MicroRNAs in the Tumor Microenvironment of Breast Cancer. Int. J. Mol. Sci. 2019, 20, 3884. [Google Scholar] [CrossRef]
- Zhang, H.; Yue, X.; Chen, Z.; Liu, C.; Wu, W.; Zhang, N.; Liu, Z.; Yang, L.; Jiang, Q.; Cheng, Q.; et al. Define cancer-associated fibroblasts (CAFs) in the tumor microenvironment: New opportunities in cancer immunotherapy and advances in clinical trials. Mol. Cancer 2023, 22, 159. [Google Scholar] [CrossRef]
- Mayer, S.; Milo, T.; Isaacson, A.; Halperin, C.; Miyara, S.; Stein, Y.; Lior, C.; Pevsner-Fischer, M.; Tzahor, E.; Mayo, A.; et al. The tumor microenvironment shows a hierarchy of cell-cell interactions dominated by fibroblasts. Nat. Commun. 2023, 14, 5810. [Google Scholar] [CrossRef]
- Ma, C.; Yang, C.; Peng, A.; Sun, T.; Ji, X.; Mi, J.; Wei, L.; Shen, S.; Feng, Q. Pan-cancer spatially resolved single-cell analysis reveals the crosstalk between cancer-associated fibroblasts and tumor microenvironment. Mol. Cancer 2023, 22, 170. [Google Scholar] [CrossRef]
- Arima, Y.; Matsueda, S.; Saya, H. Significance of Cancer-Associated Fibroblasts in the Interactions of Cancer Cells with the Tumor Microenvironment of Heterogeneous Tumor Tissue. Cancers 2023, 15, 2536. [Google Scholar] [CrossRef] [PubMed]
- Chhabra, Y.; Weeraratna, A.T. Fibroblasts in cancer: Unity in heterogeneity. Cell 2023, 186, 1580–1609. [Google Scholar] [CrossRef] [PubMed]
- Baroni, S.; Romero-Cordoba, S.; Plantamura, I.; Dugo, M.; D’ippolito, E.; Cataldo, A.; Cosentino, G.; Angeloni, V.; Rossini, A.; Daidone, M.G. Exosome-mediated delivery of miR-9 induces cancer-associated fibroblast-like properties in human breast fibroblasts. Cell Death Dis. 2016, 7, e2312. [Google Scholar] [CrossRef] [PubMed]
- Vu, L.T.; Peng, B.; Zhang, D.X.; Ma, V.; Mathey-Andrews, C.A.; Lam, C.K.; Kiomourtzis, T.; Jin, J.; McReynolds, L.; Huang, L. Tumor-secreted extracellular vesicles promote the activation of cancer-associated fibroblasts via the transfer of microRNA-125b. J. Extracell. Vesicles 2019, 8, 1599680. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.; Wu, X.; Zhou, W.; Fong, M.Y.; Cao, M.; Liu, J.; Liu, X.; Chen, C.-H.; Fadare, O.; Pizzo, D.P. Cancer-cell-secreted exosomal miR-105 promotes tumour growth through the MYC-dependent metabolic reprogramming of stromal cells. Nat. Cell Biol. 2018, 20, 597–609. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.; Kwon, A.; Huh, Y.H.; Rhee, S.; Song, W.K. Tumor-derived miR-130b-3p induces cancer-associated fibroblast activation by targeting SPIN90 in luminal A breast cancer. Oncogenesis 2022, 11, 47. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, Y.; Zhong, J.; Li, M.; Zhou, Y.; Lin, Q.; Zong, S.; Luo, W.; Wang, J.; Wang, K. Tumor-derived Cav-1 promotes pre-metastatic niche formation and lung metastasis in breast cancer. Theranostics 2023, 13, 1684. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Liu, T.; Chen, J.; Ni, H.; Li, W. Survivin in breast cancer–derived exosomes activates fibroblasts by up-regulating SOD1, whose feedback promotes cancer proliferation and metastasis. J. Biol. Chem. 2020, 295, 13737–13752. [Google Scholar] [CrossRef]
- Ji, X.; Huang, X.; Li, C.; Guan, N.; Pan, T.; Dong, J.; Li, L. Effect of tumor-associated macrophages on the pyroptosis of breast cancer tumor cells. Cell Commun. Signal. CCS 2023, 21, 197. [Google Scholar] [CrossRef]
- Song, J.; Xiao, T.; Li, M.; Jia, Q. Tumor-associated macrophages: Potential therapeutic targets and diagnostic markers in cancer. Pathol. Res. Pract. 2023, 249, 154739. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Bao, X.; Zou, Y.; Wang, L.; Li, Y.; Yang, L.; Liao, A.; Zhang, X.; Jiang, X.; Liang, D.; et al. d-lactate modulates M2 tumor-associated macrophages and remodels immunosuppressive tumor microenvironment for hepatocellular carcinoma. Sci. Adv. 2023, 9, eadg2697. [Google Scholar] [CrossRef] [PubMed]
- Basak, U.; Sarkar, T.; Mukherjee, S.; Chakraborty, S.; Dutta, A.; Dutta, S.; Nayak, D.; Kaushik, S.; Das, T.; Sa, G. Tumor-associated macrophages: An effective player of the tumor microenvironment. Front. Immunol. 2023, 14, 1295257. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Huang, R.; Lu, Z.; Wang, Z.; Chen, X.; Huang, D. Exosomes from M1-polarized macrophages promote apoptosis in lung adenocarcinoma via the miR-181a-5p/ETS1/STK16 axis. Cancer Sci 2022, 113, 986–1001. [Google Scholar] [CrossRef] [PubMed]
- Ham, S.; Lima, L.G.; Chai, E.P.Z.; Muller, A.; Lobb, R.J.; Krumeich, S.; Wen, S.W.; Wiegmans, A.P.; Möller, A. Breast cancer-derived exosomes alter macrophage polarization via gp130/STAT3 signaling. Front. Immunol. 2018, 9, 871. [Google Scholar] [CrossRef] [PubMed]
- Xun, J.; Du, L.; Gao, R.; Shen, L.; Wang, D.; Kang, L.; Zhang, Z.; Zhang, Y.; Yue, S.; Feng, S. Cancer-derived exosomal miR-138-5p modulates polarization of tumor-associated macrophages through inhibition of KDM6B. Theranostics 2021, 11, 6847. [Google Scholar] [CrossRef] [PubMed]
- Qi, M.; Xia, Y.; Wu, Y.; Zhang, Z.; Wang, X.; Lu, L.; Dai, C.; Song, Y.; Xu, K.; Ji, W. Lin28B-high breast cancer cells promote immune suppression in the lung pre-metastatic niche via exosomes and support cancer progression. Nat. Commun. 2022, 13, 897. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Mo, Z.; Lai, G.; Chen, X.; Li, R.; Wu, R.; Zhu, J.; Zheng, F. Targeting tumor exosomal circular RNA cSERPINE2 suppresses breast cancer progression by modulating MALT1-NF-κB-IL-6 axis of tumor-associated macrophages. J. Exp. Clin. Cancer Res. 2023, 42, 48. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Shi, W.; Hu, W.; Zhao, Y.; Zhao, X.; Dong, F.; Xin, Y.; Peng, T.; Liu, C. Endoplasmic reticulum stress promotes breast cancer cells to release exosomes circ_0001142 and induces M2 polarization of macrophages to regulate tumor progression. Pharmacol. Res. 2022, 177, 106098. [Google Scholar] [CrossRef]
- Yao, X.; Tu, Y.; Xu, Y.; Guo, Y.; Yao, F.; Zhang, X. Endoplasmic reticulum stress-induced exosomal miR-27a-3p promotes immune escape in breast cancer via regulating PD-L1 expression in macrophages. J. Cell. Mol. Med. 2020, 24, 9560–9573. [Google Scholar] [CrossRef]
- Frank, A.-C.; Ebersberger, S.; Fink, A.F.; Lampe, S.; Weigert, A.; Schmid, T.; Ebersberger, I.; Syed, S.N.; Brüne, B. Apoptotic tumor cell-derived microRNA-375 uses CD36 to alter the tumor-associated macrophage phenotype. Nat. Commun. 2019, 10, 1135. [Google Scholar] [CrossRef] [PubMed]
- Rabe, D.C.; Walker, N.D.; Rustandy, F.D.; Wallace, J.; Lee, J.; Stott, S.L.; Rosner, M.R. Tumor extracellular vesicles regulate macrophage-driven metastasis through CCL5. Cancers 2021, 13, 3459. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Wu, K.; Sharma, S.; Xing, F.; Wu, S.-Y.; Tyagi, A.; Deshpande, R.; Singh, R.; Wabitsch, M.; Mo, Y.-Y. Exosomal miR-1304-3p promotes breast cancer progression in African Americans by activating cancer-associated adipocytes. Nat. Commun. 2022, 13, 7734. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Wang, Z.; Yao, F.; Sun, K.; Li, Z.; Sun, S.; Li, C. Breast cancer cell-derived exosome-delivered microRNA-155 targets UBQLN1 in adipocytes and facilitates cancer cachexia-related fat loss. Hum. Mol. Genet. 2023, 32, 2219–2228. [Google Scholar] [CrossRef] [PubMed]
- Fong, M.Y.; Zhou, W.; Liu, L.; Alontaga, A.Y.; Chandra, M.; Ashby, J.; Chow, A.; O’Connor, S.T.F.; Li, S.; Chin, A.R. Breast-cancer-secreted miR-122 reprograms glucose metabolism in premetastatic niche to promote metastasis. Nat. Cell Biol. 2015, 17, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; Feng, J.; Lyu, F.; Xing, F.; Sharma, S.; Liu, Y.; Wu, S.-Y.; Zhao, D.; Tyagi, A.; Deshpande, R.P. Exosomal miR-19a and IBSP cooperate to induce osteolytic bone metastasis of estrogen receptor-positive breast cancer. Nat. Commun. 2021, 12, 5196. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.W.; Chen, Y.; Li, J.Y.; Xiao, L.; Zhang, W.J.; Zhao, J.L.; Gu, H.C.; Wu, H.Y.; Zuo, G.S.L.; Deng, K.Y. Bone marrow cells are differentiated into MDSCs by BCC-Ex through down-regulating the expression of CXCR4 and activating STAT3 signalling pathway. J. Cell. Mol. Med. 2021, 25, 5497–5510. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wei, H.; Wang, J.; Li, L.; Chen, A.; Li, Z. MicroRNA-181d-5p-containing exosomes derived from CAFs promote EMT by regulating CDX2/HOXA5 in breast cancer. Mol. Ther. Nucleic Acids 2020, 19, 654–667. [Google Scholar] [CrossRef] [PubMed]
- Tao, S.; Li, H.; Ma, X.; Ma, Y.; He, J.; Gao, Y.; Li, J. Elevating microRNA-1-3p shuttled by cancer-associated fibroblasts-derived extracellular vesicles suppresses breast cancer progression and metastasis by inhibiting GLIS1. Cancer Gene Ther. 2021, 28, 634–648. [Google Scholar] [CrossRef]
- Yan, Z.; Sheng, Z.; Zheng, Y.; Feng, R.; Xiao, Q.; Shi, L.; Li, H.; Yin, C.; Luo, H.; Hao, C. Cancer-associated fibroblast-derived exosomal miR-18b promotes breast cancer invasion and metastasis by regulating TCEAL7. Cell Death Dis. 2021, 12, 1120. [Google Scholar] [CrossRef]
- Dou, D.; Ren, X.; Han, M.; Xu, X.; Ge, X.; Gu, Y.; Wang, X. Cancer-associated fibroblasts-derived exosomes suppress immune cell function in breast cancer via the miR-92/PD-L1 pathway. Front. Immunol. 2020, 11, 2026. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Sang, Y.; Song, X.; Zhang, D.; Wang, L.; Zhao, W.; Liang, Y.; Zhang, N.; Yang, Q. Exosomal miR-500a-5p derived from cancer-associated fibroblasts promotes breast cancer cell proliferation and metastasis through targeting USP28. Theranostics 2021, 11, 3932. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zeng, C.; Zhan, Y.; Wang, H.; Jiang, X.; Li, W. Aberrant low expression of p85α in stromal fibroblasts promotes breast cancer cell metastasis through exosome-mediated paracrine Wnt10b. Oncogene 2017, 36, 4692–4705. [Google Scholar] [CrossRef] [PubMed]
- Shimoda, M.; Principe, S.; Jackson, H.W.; Luga, V.; Fang, H.; Molyneux, S.D.; Shao, Y.W.; Aiken, A.; Waterhouse, P.D.; Karamboulas, C. Loss of the Timp gene family is sufficient for the acquisition of the CAF-like cell state. Nat. Cell Biol. 2014, 16, 889–901. [Google Scholar] [CrossRef] [PubMed]
- Xi, L.; Peng, M.; Liu, S.; Liu, Y.; Wan, X.; Hou, Y.; Qin, Y.; Yang, L.; Chen, S.; Zeng, H. Hypoxia-stimulated ATM activation regulates autophagy-associated exosome release from cancer-associated fibroblasts to promote cancer cell invasion. J. Extracell. Vesicles 2021, 10, e12146. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Fan, P.; Zhang, C.; Xie, J.; Gu, X.; Lei, S.; Chen, Z.; Huang, Z. Exosomal microRNA-503-3p derived from macrophages represses glycolysis and promotes mitochondrial oxidative phosphorylation in breast cancer cells by elevating DACT2. Cell Death Discov. 2021, 7, 119. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhang, C.; Guo, J.; Wang, W.; Si, Q.; Chen, C.; Luo, Y.; Duan, Z. Exosomal miRNA-223-3p derived from tumor associated macrophages promotes pulmonary metastasis of breast cancer 4T1 cells. Transl. Oncol. 2023, 35, 101715. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Zhang, Q.; Zhang, X.; Han, Q.; Li, H.; Mao, Y.; Wang, X.; Guo, H.; Irwin, D.M.; Niu, G. Exosomes from macrophages exposed to apoptotic breast cancer cells promote breast cancer proliferation and metastasis. J. Cancer 2019, 10, 2892. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.Y.; Zhu, W.W.; Wang, Z.; Huang, J.B.; Wang, S.H.; Bai, F.M.; Li, T.E.; Zhu, Y.; Zhao, J.; Yang, X.; et al. Driver mutations of intrahepatic cholangiocarcinoma shape clinically relevant genomic clusters with distinct molecular features and therapeutic vulnerabilities. Theranostics 2022, 12, 260–276. [Google Scholar] [CrossRef]
- Chen, S.; Chen, Z.; Li, Z.; Li, S.; Wen, Z.; Cao, L.; Chen, Y.; Xue, P.; Li, H.; Zhang, D. Tumor-associated macrophages promote cholangiocarcinoma progression via exosomal Circ_0020256. Cell Death Dis. 2022, 13, 94. [Google Scholar] [CrossRef]
- Guan, H.; Peng, R.; Fang, F.; Mao, L.; Chen, Z.; Yang, S.; Dai, C.; Wu, H.; Wang, C.; Feng, N. Tumor-associated macrophages promote prostate cancer progression via exosome-mediated miR-95 transfer. J. Cell. Physiol. 2020, 235, 9729–9742. [Google Scholar] [CrossRef]
- Wang, S.; Su, X.; Xu, M.; Xiao, X.; Li, X.; Li, H.; Keating, A.; Zhao, R.C. Exosomes secreted by mesenchymal stromal/stem cell-derived adipocytes promote breast cancer cell growth via activation of Hippo signaling pathway. Stem Cell Res. Ther. 2019, 10, 1–12. [Google Scholar] [CrossRef]
- Jafari, N.; Kolla, M.; Meshulam, T.; Shafran, J.S.; Qiu, Y.; Casey, A.N.; Pompa, I.R.; Ennis, C.S.; Mazzeo, C.S.; Rabhi, N. Adipocyte-derived exosomes may promote breast cancer progression in type 2 diabetes. Sci. Signal. 2021, 14, eabj2807. [Google Scholar] [CrossRef]
- Gernapudi, R.; Yao, Y.; Zhang, Y.; Wolfson, B.; Roy, S.; Duru, N.; Eades, G.; Yang, P.; Zhou, Q. Targeting exosomes from preadipocytes inhibits preadipocyte to cancer stem cell signaling in early-stage breast cancer. Breast Cancer Res. Treat. 2015, 150, 685–695. [Google Scholar] [CrossRef]
- Lazar, I.; Clement, E.; Dauvillier, S.; Milhas, D.; Ducoux-Petit, M.; LeGonidec, S.; Moro, C.; Soldan, V.; Dalle, S.; Balor, S. Adipocyte exosomes promote melanoma aggressiveness through fatty acid oxidation: A novel mechanism linking obesity and cancer. Cancer Res. 2016, 76, 4051–4057. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wu, Y.; Guo, J.; Fei, X.; Yu, L.; Ma, S. Adipocyte-derived exosomes promote lung cancer metastasis by increasing MMP9 activity via transferring MMP3 to lung cancer cells. Oncotarget 2017, 8, 81880. [Google Scholar] [CrossRef] [PubMed]
- Shedden, K.; Xie, X.T.; Chandaroy, P.; Chang, Y.T.; Rosania, G.R. Expulsion of small molecules in vesicles shed by cancer cells: Association with gene expression and chemosensitivity profiles. Cancer Res. 2003, 63, 4331–4337. [Google Scholar]
- Lv, M.-M.; Zhu, X.-Y.; Chen, W.-X.; Zhong, S.-L.; Hu, Q.; Ma, T.-F.; Zhang, J.; Chen, L.; Tang, J.-H.; Zhao, J.-H. Exosomes mediate drug resistance transfer in MCF-7 breast cancer cells and a probable mechanism is delivery of P-glycoprotein. Tumor Biol. 2014, 35, 10773–10779. [Google Scholar] [CrossRef] [PubMed]
- Ning, K.; Wang, T.; Sun, X.; Zhang, P.; Chen, Y.; Jin, J.; Hua, D. UCH-L1-containing exosomes mediate chemotherapeutic resistance transfer in breast cancer. J. Surg. Oncol. 2017, 115, 932–940. [Google Scholar] [CrossRef]
- Kreger, B.T.; Johansen, E.R.; Cerione, R.A.; Antonyak, M.A. The enrichment of survivin in exosomes from breast cancer cells treated with paclitaxel promotes cell survival and chemoresistance. Cancers 2016, 8, 111. [Google Scholar] [CrossRef]
- Wang, B.; Zhang, Y.; Ye, M.; Wu, J.; Ma, L.; Chen, H. Cisplatin-resistant MDA-MB-231 cell-derived exosomes increase the resistance of recipient cells in an exosomal miR-423-5p-dependent manner. Curr. Drug Metab. 2019, 20, 804–814. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Pei, X.; Guo, G.; Qian, X.; Dou, D.; Zhang, Z.; Xu, X.; Duan, X. Exosome-mediated transfer of long noncoding RNA H19 induces doxorubicin resistance in breast cancer. J. Cell. Physiol. 2020, 235, 6896–6904. [Google Scholar] [CrossRef] [PubMed]
- Li, X.J.; Ren, Z.J.; Tang, J.H.; Yu, Q. Exosomal MicroRNA MiR-1246 promotes cell proliferation, invasion and drug resistance by targeting CCNG2 in breast cancer. Cell. Physiol. Biochem. 2018, 44, 1741–1748. [Google Scholar] [CrossRef] [PubMed]
- Semina, S.E.; Scherbakov, A.M.; Vnukova, A.A.; Bagrov, D.V.; Evtushenko, E.G.; Safronova, V.M.; Golovina, D.A.; Lyubchenko, L.N.; Gudkova, M.V.; Krasil’nikov, M.A. Exosome-mediated transfer of cancer cell resistance to antiestrogen drugs. Molecules 2018, 23, 829. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.G.; Yang, M.F.; Ren, Y.Q.; Wu, C.H.; Wang, L.Q. Exosomes mediated transfer of lncRNA UCA1 results in increased tamoxifen resistance in breast cancer cells. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 4362–4368. [Google Scholar] [PubMed]
- Wei, Y.; Lai, X.; Yu, S.; Chen, S.; Ma, Y.; Zhang, Y.; Li, H.; Zhu, X.; Yao, L.; Zhang, J. Exosomal miR-221/222 enhances tamoxifen resistance in recipient ER-positive breast cancer cells. Breast Cancer Res. Treat. 2014, 147, 423–431. [Google Scholar] [CrossRef]
- Dong, H.; Wang, W.; Chen, R.; Zhang, Y.; Zou, K.; Ye, M.; He, X.; Zhang, F.; Han, J. Exosome-mediated transfer of lncRNA-SNHG14 promotes trastuzumab chemoresistance in breast cancer. Int. J. Oncol. 2018, 53, 1013–1026. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Chen, M.; Xing, P.; Yan, X.; Xie, B. Increased expression of exosomal AGAP2-AS1 (AGAP2 antisense RNA 1) in breast cancer cells inhibits trastuzumab-induced cell cytotoxicity. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2019, 25, 2211. [Google Scholar] [CrossRef]
- Martinez, V.G.; O’Neill, S.; Salimu, J.; Breslin, S.; Clayton, A.; Crown, J.; O’Driscoll, L. Resistance to HER2-targeted anti-cancer drugs is associated with immune evasion in cancer cells and their derived extracellular vesicles. Oncoimmunology 2017, 6, e1362530. [Google Scholar] [CrossRef]
- Han, M.; Hu, J.; Lu, P.; Cao, H.; Yu, C.; Li, X.; Qian, X.; Yang, X.; Yang, Y.; Han, N. Exosome-transmitted miR-567 reverses trastuzumab resistance by inhibiting ATG5 in breast cancer. Cell Death Dis. 2020, 11, 43. [Google Scholar] [CrossRef]
- Chanteloup, G.; Cordonnier, M.; Isambert, N.; Bertaut, A.; Hervieu, A.; Hennequin, A.; Luu, M.; Zanetta, S.; Coudert, B.; Bengrine, L. Monitoring HSP70 exosomes in cancer patients’ follow up: A clinical prospective pilot study. J. Extracell. Vesicles 2020, 9, 1766192. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhong, W.; Bu, J.; Li, Y.; Li, R.; Nie, R.; Xiao, C.; Ma, K.; Huang, X.; Li, Y. Exosomal protein CD82 as a diagnostic biomarker for precision medicine for breast cancer. Mol. Carcinog. 2019, 58, 674–685. [Google Scholar] [CrossRef] [PubMed]
- Stefanska, A.M.; Walsh, P.T. Chronic obstructive pulmonary disease: Evidence for an autoimmune component. Cell. Mol. Immunol. 2009, 6, 81–86. [Google Scholar] [CrossRef]
- Golubovskaya, V.M. Focal adhesion kinase and cross-linked signaling in cancer. Anti-Cancer Agents Med. Chem. 2014, 14, 2. [Google Scholar] [CrossRef] [PubMed]
- Melo, S.A.; Luecke, L.B.; Kahlert, C.; Fernandez, A.F.; Gammon, S.T.; Kaye, J.; LeBleu, V.S.; Mittendorf, E.A.; Weitz, J.; Rahbari, N. Glypican-1 identifies cancer exosomes and detects early pancreatic cancer. Nature 2015, 523, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Zhai, L.-Y.; Li, M.-X.; Pan, W.-L.; Chen, Y.; Li, M.-M.; Pang, J.-X.; Zheng, L.; Chen, J.-X.; Duan, W.-J. In situ detection of plasma exosomal microRNA-1246 for breast cancer diagnostics by a Au nanoflare probe. ACS Appl. Mater. Interfaces 2018, 10, 39478–39486. [Google Scholar] [CrossRef] [PubMed]
- Stevic, I.; Müller, V.; Weber, K.; Fasching, P.A.; Karn, T.; Marmé, F.; Schem, C.; Stickeler, E.; Denkert, C.; van Mackelenbergh, M. Specific microRNA signatures in exosomes of triple-negative and HER2-positive breast cancer patients undergoing neoadjuvant therapy within the GeparSixto trial. BMC Med. 2018, 16, 179. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Li, Y.; Guo, R.; Zhao, J.; Chi, W.; Lai, H.; Wang, J.; Wang, Z.; Li, L.; Sang, Y. Plasma extracellular vesicle long RNA profiles in the diagnosis and prediction of treatment response for breast cancer. NPJ Breast Cancer 2021, 7, 154. [Google Scholar] [CrossRef]
- Obidiro, O.; Battogtokh, G.; Akala, E.O. Triple Negative Breast Cancer Treatment Options and Limitations: Future Outlook. Pharmaceutics 2023, 15, 1796. [Google Scholar] [CrossRef]
- Wang, R. Current perspectives on naturally occurring saponins as anticancer agents. Arch. Pharm. 2022, 355, e2100469. [Google Scholar] [CrossRef]
- Herdiana, Y.; Husni, P.; Nurhasanah, S.; Shamsuddin, S.; Wathoni, N. Chitosan-Based Nano Systems for Natural Antioxidants in Breast Cancer Therapy. Polymers 2023, 15, 2953. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Zhang, Q.; Feng, C.; Zhang, J.; Qin, Y.; Meng, L. Advances in the Pharmacological Activities and Effects of Perilla Ketone and Isoegomaketone. Evid. Based Complement. Altern. Med. Ecam 2022, 2022, 8809792. [Google Scholar] [CrossRef] [PubMed]
- Pascucci, L.; Coccè, V.; Bonomi, A.; Ami, D.; Ceccarelli, P.; Ciusani, E.; Viganò, L.; Locatelli, A.; Sisto, F.; Doglia, S.M. Paclitaxel is incorporated by mesenchymal stromal cells and released in exosomes that inhibit in vitro tumor growth: A new approach for drug delivery. J. Control. Release 2014, 192, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Qiu, X.; Li, Z.; Han, X.; Zhen, L.; Luo, C.; Liu, M.; Yu, K.; Ren, Y. Tumor-derived nanovesicles promote lung distribution of the therapeutic nanovector through repression of Kupffer cell-mediated phagocytosis. Theranostics 2019, 9, 2618. [Google Scholar] [CrossRef]
- Morad, G.; Carman, C.V.; Hagedorn, E.J.; Perlin, J.R.; Zon, L.I.; Mustafaoglu, N.; Park, T.-E.; Ingber, D.E.; Daisy, C.C.; Moses, M.A. Tumor-derived extracellular vesicles breach the intact blood–brain barrier via transcytosis. ACS Nano 2019, 13, 13853–13865. [Google Scholar] [CrossRef]
- Park, E.J.; Jung, H.J.; Choi, H.J.; Jang, H.J.; Park, H.J.; Nejsum, L.N.; Kwon, T.H. Exosomes co-expressing AQP5-targeting miRNAs and IL-4 receptor-binding peptide inhibit the migration of human breast cancer cells. FASEB J. 2020, 34, 3379–3398. [Google Scholar] [CrossRef]
- Huang, L.; Rong, Y.; Tang, X.; Yi, K.; Qi, P.; Hou, J.; Liu, W.; He, Y.; Gao, X.; Yuan, C. Engineered exosomes as an in situ DC-primed vaccine to boost antitumor immunity in breast cancer. Mol. Cancer 2022, 21, 45. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Cheng, Q.; Hou, T.; Han, M.; Smbatyan, G.; Lang, J.E.; Epstein, A.L.; Lenz, H.-J.; Zhang, Y. Genetically engineered cell-derived nanoparticles for targeted breast cancer immunotherapy. Mol. Ther. 2020, 28, 536–547. [Google Scholar] [CrossRef]
- Qiu, Y.; Yang, Y.; Yang, R.; Liu, C.; Hsu, J.-M.; Jiang, Z.; Sun, L.; Wei, Y.; Li, C.-W.; Yu, D. Activated T cell-derived exosomal PD-1 attenuates PD-L1-induced immune dysfunction in triple-negative breast cancer. Oncogene 2021, 40, 4992–5001. [Google Scholar] [CrossRef]
- Li, S.; Wu, Y.; Ding, F.; Yang, J.; Li, J.; Gao, X.; Zhang, C.; Feng, J. Engineering macrophage-derived exosomes for targeted chemotherapy of triple-negative breast cancer. Nanoscale 2020, 12, 10854–10862. [Google Scholar] [CrossRef]
- Wang, S.; Li, F.; Ye, T.; Wang, J.; Lyu, C.; Qing, S.; Ding, Z.; Gao, X.; Jia, R.; Yu, D. Macrophage-tumor chimeric exosomes accumulate in lymph node and tumor to activate the immune response and the tumor microenvironment. Sci. Transl. Med. 2021, 13, eabb6981. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Multiple roles of exosomes in various aspects of breast cancer.

Figure 2.
Roles of cancer-derived exosomes in the metastasis and progression of breast cancer.
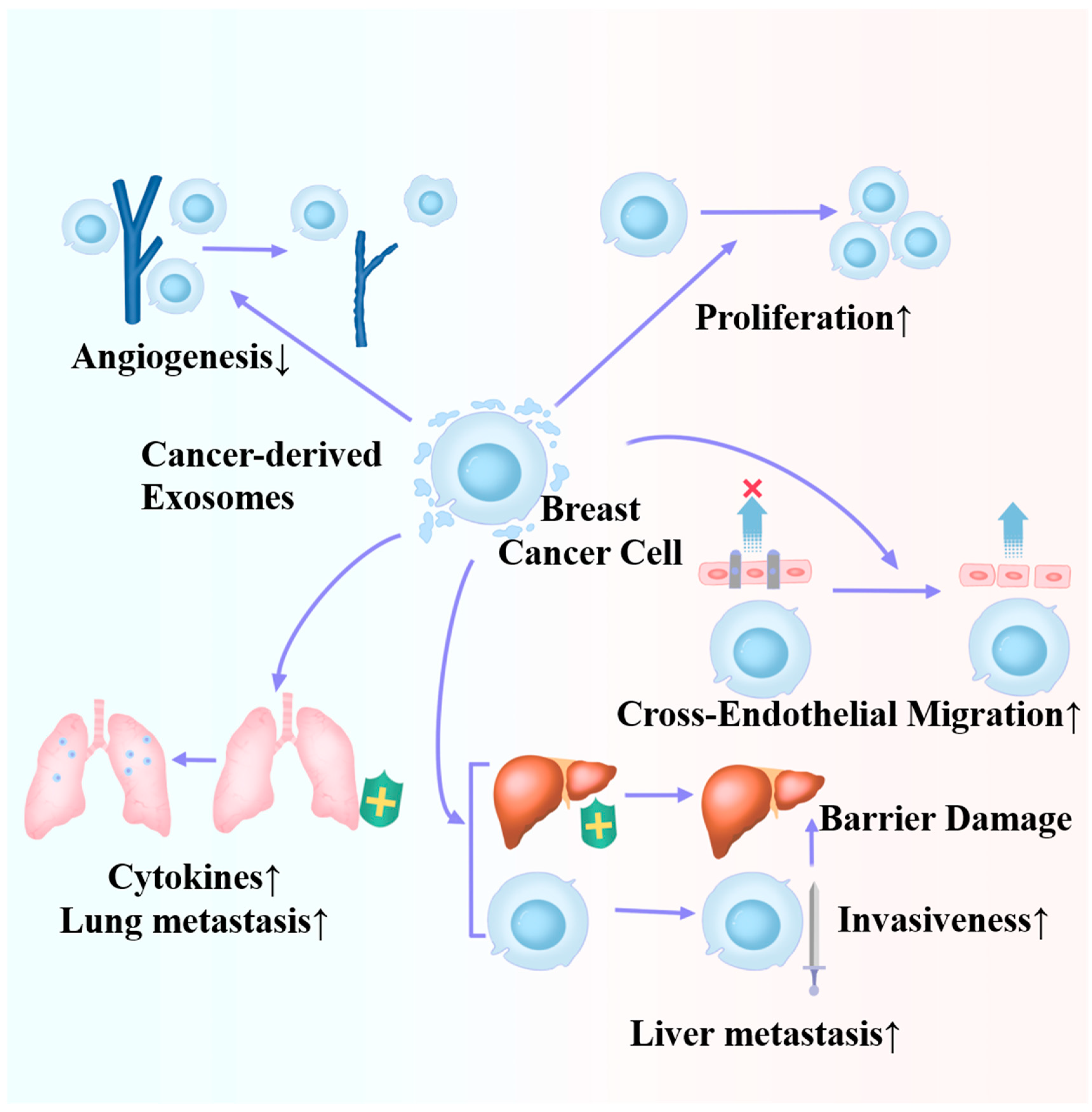
Figure 3.
Roles of cancer-derived exosomes in the microenvironment of breast cancer cells.

Figure 4.
Roles of microenvironment-derived exosomes in the proliferation, invasion, and migration of breast cancer.
Figure 4.
Roles of microenvironment-derived exosomes in the proliferation, invasion, and migration of breast cancer.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Cancer-derived exosomes in the metastasis and progression of breast cancer.
| Biological Roles | Intermediates | Major Mechanisms | References |
|---|---|---|---|
| BC metastasis | / | FAK/Src-dependent proteins | [37] |
| BC progression | miR-130a-3p | RAB5B | [38] |
| BC invasiveness and metastasis | miR-222 | PDZ/PDLIM2/NF-kB | [39] |
| BC progression and metastasis | miR-1910-3p | MTMR3/NF-kB | [40] |
| HUVEC angiogenesis (BC inhibition) | miR-145 | IRS1/Raf/ERK IRS1/PI3K/Akt/mTOR | [41] |
| BC cross-endothelial migration | miR-939 | Vascular endothelial cadherin | [42] |
| BC liver metastasis | miR-4443 | Barrier damage TIMP2/MMP-2 | [43] |
| BC lung metastasis | miR-183-5p | PPP1CA-IL2/TNFα | [44] |
Table 2.
Cancer-derived exosomes in the different tumor microenvironment cell subtype and breast cancer cells.
Table 2.
Cancer-derived exosomes in the different tumor microenvironment cell subtype and breast cancer cells.
| TME Cell Subtype | Biological Roles | Intermediates | Major Mechanisms | References |
|---|---|---|---|---|
| Cancer-associated fibroblasts | ||||
| NF to CAF | miR-9 | / | [55] | |
| NF to CAF | miR-125b | / | [56] | |
| BC tumor growth | miR-105 | Metabolism reprogramming | [57] | |
| BC progression | miR-130b-3p | SPIN90 | [58] | |
| BC lung metastasis | Cav-1 | Extracellular matrix deposition | [59] | |
| BC metastasis and proliferation | Survivin | SOD-1 | [60] | |
| Tumor-associated macrophages | ||||
| M1 to M2 | GP130 | STAT3 pathway | [66] | |
| M1 to M2 | miR-138-5p | KDM6B | [67] | |
| BC lung metastasis | Lin28B | PD-L1/Cytokine imbalance | [68] | |
| M1 to M2 | Circ_0001142 | Endoplasmic reticulum | [70] | |
| BC immune escape | miR-27a-3p | PD-L1 | [71] | |
| BC migration and infiltration | CD375/miR-63 | TNS3PXN | [72] | |
| BC lung metastasis | CCL5 | Immune infiltration | [73] | |
| Cancer-related adipocytes | ||||
| BC progression | miR-1304-3p | GATA2 | [74] | |
| BC inhibition | miR-155 | White adipose tissue browning | [75] | |
| BC progression | miR-122 | Pyruvate kinase | [76] |
Table 3.
Tumor microenvironment-derived exosomes in breast cancer cells.
| Tumor Microenvironment-Derived Exosomes | Biological Roles | Intermediates | Major Mechanisms | References |
|---|---|---|---|---|
| Fibroblast-derived exosomes | ||||
| BC invasiveness | miR-181d-5p | CDX2/HOXA5 axis | [79] | |
| BC metastasis and invasion | miR-1b/miR-3-7p | TCEAL1 GLIS232 | [80] | |
| BC metastasis and invasion | miR-18b | TCEAL7/NF-kB/Snail/EMT | [81] | |
| BC proliferation and migration | miR-92 | PD-L1/LATS2 | [82] | |
| BC tumor-infiltrating immune cell function | ||||
| BC proliferation | miR-500a-5p | USP28 | [83] | |
| BC metastasis | Wnt10b parasecretion | [84] | ||
| BC progression | ADAM10 | RhoA/Notch axis | [85] | |
| BC invasiveness | GPR64 | IL-8/MMP9 | [86] | |
| Immune cell-derived exosomes | ||||
| BC progression | miR-503-3p | DACT2/Wnt/β-Catenin | [87] | |
| BC lung metastasis | miR-223-3p | Cbx5 | [88] | |
| BC recurrence and metastasis after chemotherapy | / | STAT3 | [89] | |
| Cancer progression | Circ_0020256 | miR-432-5p/E2F3 | [91] | |
| Cancer progression | miR-95 | JunB/EMT | [92] | |
| Adipocyte-derived exosomes | ||||
| BC cell proliferation and migration | / | Hippo | [93] | |
| BC cancer stem cell | TSP5 COMP/BRD2 or BRD3 | EMT | [94] | |
| BC differentiation, migration, and stemness | miR-140 | SOX2/SOX9 | [95] | |
| Cancer invasiveness and migration | Proteins involved in fatty acid oxidation | Metabolic reprogramming | [96] | |
| Cancer invasiveness | MMP3 | MMP9 | [97] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wang, H.; Wang, R.; Shen, K.; Huang, R.; Wang, Z. Biological Roles and Clinical Applications of Exosomes in Breast Cancer: A Brief Review. Int. J. Mol. Sci. 2024, 25, 4620. https://doi.org/10.3390/ijms25094620
AMA Style
Wang H, Wang R, Shen K, Huang R, Wang Z. Biological Roles and Clinical Applications of Exosomes in Breast Cancer: A Brief Review. International Journal of Molecular Sciences. 2024; 25(9):4620. https://doi.org/10.3390/ijms25094620
Chicago/Turabian StyleWang, Han, Ruo Wang, Kunwei Shen, Renhong Huang, and Zheng Wang. 2024. "Biological Roles and Clinical Applications of Exosomes in Breast Cancer: A Brief Review" International Journal of Molecular Sciences 25, no. 9: 4620. https://doi.org/10.3390/ijms25094620
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.