Spatial Patterns and Short-term Changes of Coral Assemblages Along a Cross-shelf Gradient in the Southwestern Lagoon of New Caledonia


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Coral Sampling
2.3. Data Analysis
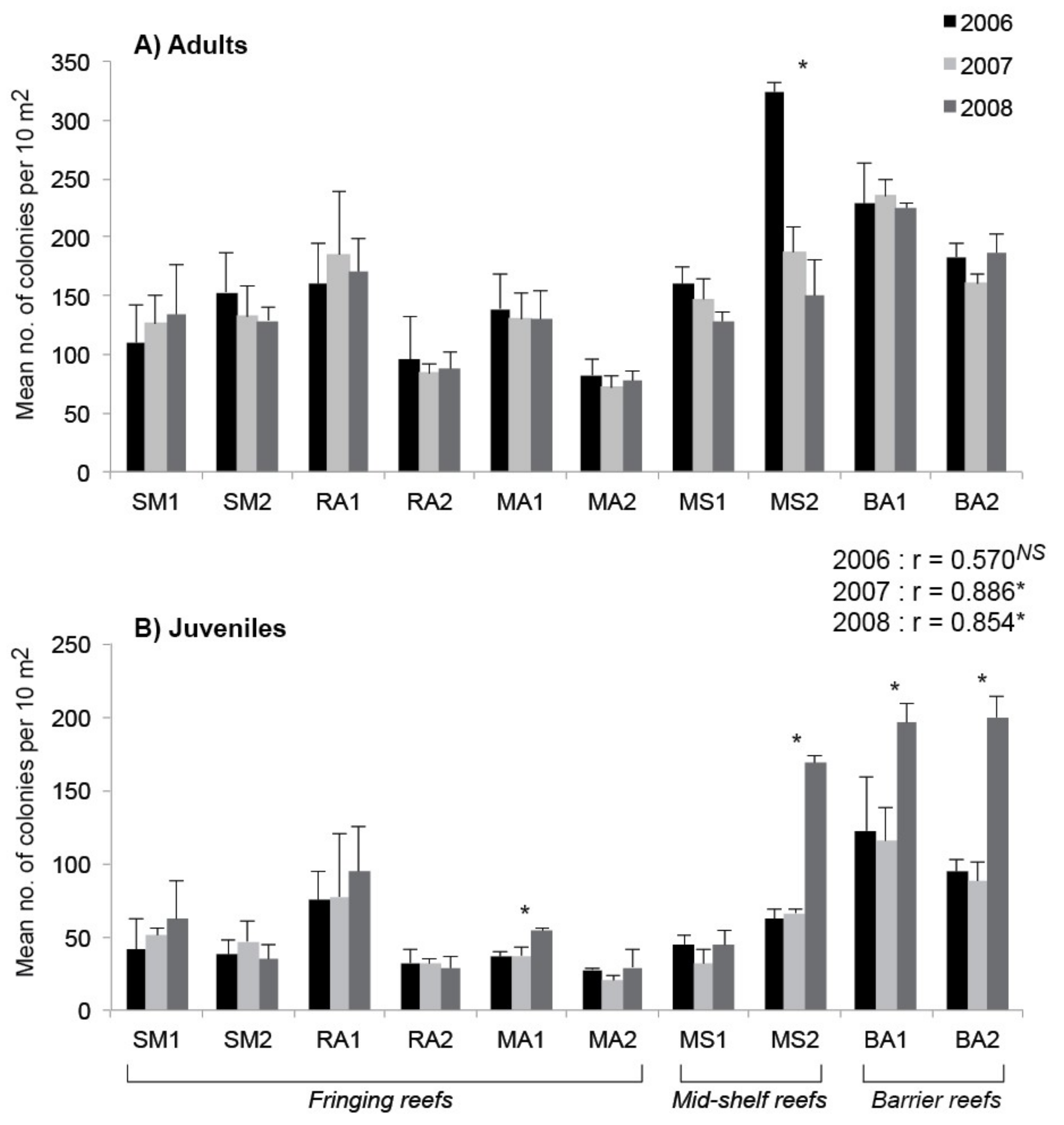
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Reaka-Kudla, M.L. The global biodiversity of coral reefs: A comparison with rain forests. In Biodiversity II: Understanding and Protecting Our Biological Resources; Reaka-Kudla, M.L., Wilson, D.E., Wilson, E.O., Eds.; The National Academy of Science, Joseph Henry Press: Washington, DC, USA, 1997; pp. 83–108. [Google Scholar]
- Fisher, R.; O’Leary, R.A.; Low-Choy, S.; Mengersen, K.; Knowlton, N.; Brainard, R.E.; Caley, M.J. Species richness on coral reefs and the pursuit of convergent global estimates. Curr. Biol. 2015, 25, 500–505. [Google Scholar] [CrossRef] [PubMed]
- Moberg, F.; Folke, C. Ecological goods and services of coral reef ecosystems. Ecol. Econ. 1999, 29, 215–233. [Google Scholar] [CrossRef]
- Kittinger, J.N.; Finkbeiner, E.M.; Glazier, E.W.; Crowder, L.B. Human dimensions of coral reef social-ecological systems. Ecol. Soc. 2012, 17, 17. [Google Scholar] [CrossRef]
- Connell, J.H. Diversity in a tropical rain forests and coral reefs. Science 1978, 199, 1302–1310. [Google Scholar] [CrossRef] [PubMed]
- Done, T.J. Coral zonation: Its nature and significance. In Perspectives on Coral Reefs; Barnes, D.J., Ed.; Australian Institute of Marine Science: Townsville, Australia, 1983; pp. 107–147. [Google Scholar]
- Huston, M.A. Patterns of species diversity on coral reefs. Annu. Rev. Ecol. Syst. 1985, 16, 149–177. [Google Scholar] [CrossRef]
- Cornell, H.V.; Karlson, R.H. Species richness of reef-building corals determined by local and regional processes. J. Anim. Ecol. 1996, 65, 233–241. [Google Scholar] [CrossRef]
- Connell, J.H.; Hughes, T.P.; Wallace, C.C. A 30-year study of coral abundance, recruitment, and disturbance at several scales in space and time. Ecol. Monogr. 1997, 67, 461–488. [Google Scholar] [CrossRef]
- Karlson, R.H.; Cornell, H.V. Species richness of coral assemblages: Detecting regional influences at local spatial scales. Ecology 2002, 83, 452–463. [Google Scholar] [CrossRef]
- Goreau, T.F. The ecology of Jamaican coral reefs. I. Species composition and zonation. Ecology 1959, 40, 67–90. [Google Scholar] [CrossRef]
- Ninio, R.; Meekan, M.G. Spatial patterns in benthic communities and the dynamics of a mosaic ecosystem on the Great Barrier Reef, Australia. Coral Reefs 2002, 21, 95–103. [Google Scholar] [CrossRef]
- Karlson, R.H.; Cornell, H.V.; Hughes, T.P. Coral communities are regionnally enriched along an oceanic biodiversity gradient. Nature 2004, 429, 867–870. [Google Scholar] [CrossRef] [PubMed]
- Cleary, D.F.R.; Polonia, A.R.M.; Renema, W.; Hoeksema, B.W.; Rachello-Dolmen, P.G.; Moolenbeek, R.G.; Budiyanto, A.; Tufi, Y.; Draisma, S.G.A.; Rud’homme van Reine, W.F.; et al. Variation in the composition of corals, fishes, sponges, echinoderms, ascidians, molluscs, foraminifera and macroalgae across a pronounced in-to-offshore environmental gradient in the Jakarta Bay-Thousand Islands coral reef complex. Mar. Pollut. Bull. 2016, 110, 701–717. [Google Scholar] [CrossRef] [PubMed]
- Teichberg, M.; Wild, C.; Bednarz, V.N.; Kegler, H.F.; Lukman, M.; Gärdes, A.A.; Heiden, J.P.; Weiand, L.; Abu, N.; Nasir, A.; et al. Spatio-temporal patterns in coral reef communities of the Spermonde Archipelago, 2012–2014, I: Comprehensive reef monitoring of water and benthic indicators reflect changes in reef health. Front. Mar. Sci. 2018, 5, 33. [Google Scholar] [CrossRef]
- Adjeroud, M. Factors influencing spatial patterns on coral reefs around Moorea, French Polynesia. Mar. Ecol. Prog. Ser. 1997, 159, 105–119. [Google Scholar] [CrossRef] [Green Version]
- Koop, K.; Booth, D.; Broadbent, A.; Brodie, J.; Bucher, D.; Capone, D.; Coll, J.; Dennison, W.; Erdmann, M.; Harrison, P.; et al. ENCORE: The effects of nutrient enrichment on coral reefs. Synthesis of results and conclusions. Mar. Pollut. Bull. 2001, 42, 91–120. [Google Scholar] [CrossRef]
- McCook, L.; Jompa, J.; Diaz-Pullido, G. Competition between corals and algae on coral reefs: A review of evidence and mechanisms. Coral Reefs 2001, 19, 400–417. [Google Scholar] [CrossRef]
- Fabricius, K.; De’ath, G.; McCook, L.; Turak, E.; Williams, D.Mc.B. Changes in algal, coral and fish assemblages along water quality gradients on the inshore Great Barrier Reef. Mar. Pollut. Bull. 2005, 51, 384–398. [Google Scholar] [CrossRef]
- Caley, M.J.; Carr, M.H.; Hixon, M.A.; Hughes, T.P.; Jones, G.P.; Menge, B.A. Recruitment and the local dynamics of open marine populations. Annu. Rev. Ecol. Syst. 1996, 27, 477–500. [Google Scholar] [CrossRef]
- Hughes, T.P.; Baird, A.H.; Dinsdale, E.A.; Moltschaniwskyj, N.A.; Pratchett, M.S.; Tanner, J.E.; Willis, B.L. Supply-side ecology works both ways: The link between benthic adults, fecundity, and larval recruits. Ecology 2000, 81, 2241–2249. [Google Scholar] [CrossRef]
- Doropoulos, C.; Ward, S.; Roff, G.; Gonzales-Rivero, M.; Mumby, P.J. Linking demographic processes of juvenile corals to benthic recovery trajectories in two common reef habitats. PLoS ONE 2015, 10, e0128535. [Google Scholar] [CrossRef]
- Adjeroud, M.; Kayal, M.; Penin, L. The importance of recruitment processes in the dynamics and resilience of reef coral assemblages. In Marine Animal Forests; Rossi, S., Bramanti, L., Gori, A., Orejas, C., Eds.; Springer: Cham, Switzerland, 2017; pp. 549–569. [Google Scholar]
- Kayal, M.; Lenihan, H.S.; Brooks, A.J.; Holbrook, S.J.; Schmitt, R.J.; Kendall, B.E. Predicting coral community recovery using multi-species population dynamics models. Ecol. Lett. 2018, 21, 1790–1799. [Google Scholar] [CrossRef]
- Hughes, T.P.; Connell, J.H. Multiple stressors on coral reefs: A long-term perspective. Limnol. Oceanogr. 1999, 44, 932–940. [Google Scholar] [CrossRef] [Green Version]
- Bellwood, D.R.; Hughes, T.P.; Folke, C.; Nyström, M. Confronting the coral reef crisis. Nature 2004, 429, 827–833. [Google Scholar] [CrossRef]
- Hoegh-Guldberg, O.; Mumby, P.J.; Hooten, A.J.; Steneck, R.S.; Greenfield, P.; Gomez, E.; Harvell, C.D.; Sale, P.F.; Edwards, A.J.; Caldeira, K.; et al. Coral reefs under rapid climate change and ocean acidification. Science 2007, 318, 1737–1742. [Google Scholar] [CrossRef]
- Edmunds, P.J.; Adjeroud, M.; Baskett, M.L.; Baums, I.B.; Budd, A.F.; Carpenter, R.C.; Fabina, N.S.; Fan, T.Y.; Franklin, E.C.; Gross, K.; et al. Persistence and change in community composition of reef corals through present, past, and future climates. PLoS ONE 2014, 9, e107525. [Google Scholar] [CrossRef] [PubMed]
- Hughes, T.P.; Barnes, M.L.; Bellwood, D.R.; Cinner, J.E.; Cumming, G.S.; Jackson, J.B.C.; Kleypas, J.; van de Leemput, I.A.; Lough, J.M.; Morrison, T.H.; et al. Coral reefs in the Anthropocene. Nature 2017, 546, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Hughes, T.P.; Baird, A.H.; Bellwood, D.R.; Card, M.; Connolly, S.R.; Folke, C.; Grosberg, R.; Hoegh-Guldberg, O.; Jackson, J.B.C.; Kleypas, J.; et al. Climate change, human impacts, and the resilience of coral reefs. Science 2003, 301, 929–933. [Google Scholar] [CrossRef] [PubMed]
- Hughes, T.P.; Kerry, J.T.; Baird, A.H.; Connolly, S.R.; Dietzel, A.; Eakin, C.M.; Heron, S.F.; Hoey, A.S.; Hoogenboom, M.; Lui, G.; et al. Global warming transforms coral reef assemblages. Nature 2018, 556, 492–496. [Google Scholar] [CrossRef] [PubMed]
- Anthony, K.R.; Marshall, P.A.; Abdulla, A.; Beeden, R.; Bergh, C.; Black, R.; Eakin, C.M.; Game, E.T.; Gooch, M.; Graham, N.A.; et al. Operationalizing resilience for adaptive coral reef management under global environmental change. Glob. Chang. Biol. 2015, 21, 48–61. [Google Scholar] [CrossRef] [PubMed]
- Adjeroud, M.; Kayal, M.; Iborra-Cantonnet, C.; Vercelloni, J.; Bosserelle, P.; Liao, V.; Chancerelle, Y.; Claudet, J.; Penin, L. Recovery of coral assemblages despite acute and recurrent disturbances on a South Central Pacific reef. Sci. Rep. 2018, 8, 9680. [Google Scholar] [CrossRef] [PubMed]
- Done, T.J. Patterns in the distribution of coral communities across the central Great Barrier Reef. Coral Reefs 1982, 1, 95–107. [Google Scholar] [CrossRef]
- Adjeroud, M. Zonation of coral assemblages in the Ryukyu islands (southern Japan): The importance of land-ocean gradients in reef habitats. Proceedings of 10th International Coral Reef Symposium, Okinawa, Japan, 28 June–2 July 2004; pp. 310–318. [Google Scholar]
- Cleary, D.F.R.; Becking, L.E.; de Voogd, N.J.; Renema, W.; de Beer, M.; van Soest, R.W.M.; Hoeksema, B.W. Variation in the diversity and composition of benthic taxa as a function of distance offshore, depth and exposure in the Spermonde Archipelago, Indonesia. Estuar. Coast. Shelf Sci. 2005, 65, 557–570. [Google Scholar] [CrossRef]
- Smith, T.B.; Nemeth, R.S.; Blondeau, J.; Calnan, J.M.; Kadison, E.; Herzlieb, S. Assessing coral reef health across onshore to offshore stress gradients in the US Virgin Islands. Mar. Pollut. Bull. 2008, 56, 1983–1991. [Google Scholar] [CrossRef] [PubMed]
- Ellis, J.; Anlauf, H.; Kürten, S.; Lozano-Cortés, D.; Alsaffar, Z.; Curdia, J.; Jones, B.; Carvalho, S. Cross shelf benthic biodiversity patterns in the Southern Red Sea. Sci. Rep. 2017, 7, 437. [Google Scholar] [CrossRef] [PubMed]
- Adjeroud, M.; Salvat, B. Spatial patterns in biodiversity of a fringing reef community along Opunohu Bay, Moorea, French Polynesia. Bull. Mar. Sci. 1996, 59, 175–187. [Google Scholar]
- Adjeroud, M. Zonation des communautés macrobenthiques le long de deux baies d’un écosystème corallien insulaire (Moorea, Polynésie française). Comptes Rendus Acad. Sci. Ser. III-Sci. Vie-Life Sci. 2000, 323, 305–313. [Google Scholar] [CrossRef]
- Adjeroud, M.; Fernandez, J.M.; Carroll, A.G.; Harrison, P.L.; Penin, L. Spatial patterns and recruitment processes of coral assemblages among contrasting environmental conditions in the southwestern lagoon of New Caledonia. Mar. Pollut. Bull. 2010, 61, 375–386. [Google Scholar] [CrossRef]
- Pichon, M. Scleractinia of New Caledonia: Check list of reef dwelling species. In Compendium of Marine Species from New Caledonia, second edition; Payri, C., Richer de Forges, B., Eds.; Doc. Sci. Tech. II7; IRD Editions: Nouméa, New Caledonia, 2007; pp. 149–157. [Google Scholar]
- Chevalier, J.P. Coral reefs of New Caledonia. In Biology and Geology of Coral Reefs; Jones, O.A., Endean, R., Eds.; Academic Press: New York, NY, USA; London, UK, 1973; pp. 143–167. [Google Scholar]
- Andréfouët, S.; Cabioch, G.; Flamand, B.; Pelletier, B. A reappraisal of the diversity of geomorphological and genetic processes of New Caledonian coral reefs: A synthesis from optical remote sensing, coring and acoustic multibeam observations. Coral Reefs 2009, 28, 691–707. [Google Scholar] [CrossRef]
- Wantiez, L. Les récifs coralliens de Nouvelle-Calédonie en 2006: États des lieux et réseau de suivi. Rev. Ecol. Terre Vie 2008, 63, 117–132. [Google Scholar]
- Wantiez, L.; Château, O.; Le Mouellic, S. Initial and mid-term impacts of cyclone Erica on coral reef fish communities and habitat in the South Lagoon Marine Park of New Caledonia. J. Mar. Biol. Assoc. 2006, 86, 1229–1236. [Google Scholar] [CrossRef]
- Guillemot, N.; Chabanet, P.; Le Pape, O. Cyclone effects on coral reef habitats in New Caledonia (South Pacific). Coral Reefs 2010, 29, 445–453. [Google Scholar] [CrossRef]
- Adjeroud, M.; Kayal, M.; Peignon, C.; Juncker, M.; Mills, S.C.; Beldade, R.; Dumas, P. Ephemeral and localized outbreaks of the coral predator Acanthaster cf. solaris in the southwestern lagoon of New Caledonia. Zool. Stud. 2018, 57, 4. [Google Scholar]
- Payri, C.; Benzoni, F.; André, L.V.; Houlbrèque, F. Le blanchissement corallien de 2016. In Nouvelle-Calédonie, Archipel de Corail; Payri, C.E., Ed.; IRD Editions/Solaris: Marseille, France, 2018; pp. 161–166. [Google Scholar]
- Jouon, A.; Douillet, P.; Ouillon, S.; Fraunié, P. Calculations of hydrodynamic time parameters in a semi-opened coastal zone using a 3D hydrodynamic model. Cont. Shelf Res. 2006, 26, 1395–1415. [Google Scholar] [CrossRef]
- Jacquet, S.; Delesalle, B.; Torréton, J.P.; Blanchot, J. Response of phytoplankton communities to increased anthropogenic influences (southwestern lagoon, New Caledonia). Mar. Ecol. Prog. Ser. 2006, 320, 65–78. [Google Scholar] [CrossRef] [Green Version]
- Mari, X.; Rochelle-Newall, E.; Torréton, J.P.; Pringault, O.; Jouon, A.; Migon, C. Water residence time: A regulatory factor of the DOM to TOM transfer efficiency. Limnol. Oceanogr. 2007, 52, 808–819. [Google Scholar] [CrossRef]
- Migon, C.; Ouillon, S.; Mari, X.; Nicolas, Z. Geochemical and hydrodynamic constraints on the distribution of trace metal concentrations in the lagoon of Nouméa, New Caledonia. Estuar. Coast. Shelf Sci. 2007, 74, 756–765. [Google Scholar] [CrossRef]
- Dalto, A.G.; Grémare, A.; Dinet, A.; Fichet, D. Muddy-bottom meiofauna responses to metal concentrations and organic enrichment in New Caledonia South-West Lagoon. Estuar. Coast. Shelf Sci. 2006, 67, 629–644. [Google Scholar] [CrossRef]
- Fernandez, J.M.; Ouillon, S.; Chevillon, C.; Douillet, P.; Fichez, R.; Le Gendre, R. A combined modelling and geochemical study of the fate of terrigenous inputs from mixed natural and mining sources in a coral reef lagoon (New Caledonia). Mar. Pollut. Bull. 2006, 52, 320–331. [Google Scholar] [CrossRef]
- Dumas, P.; Kulbicki, M.; Chifflet, S.; Fichez, R.; Ferraris, J. Environmental factors influencing urchin spatial distributions on disturbed coral reefs (New Caledonia, South Pacific). J. Exp. Mar. Biol. Ecol. 2007, 344, 88–100. [Google Scholar] [CrossRef]
- Breau, L. Etude de la Bioaccumulation des Métaux dans Quelques Espèces Marines Tropicales: Recherche de Bioindicateurs de Contamination et Application à la Surveillance de L’environnement Côtier dans le Lagon Sud-Ouest de la Nouvelle-Calédonie. Ph.D. Thesis, Université de La Rochelle, La Rochelle, France, 2003. [Google Scholar]
- Harrison, P.L.; Wallace, C.C. Reproduction, dispersal and recruitment of scleractinian corals. In Coral Reef Ecosystems, Ecosystems of the World; Dubinsky, Z., Ed.; Elsevier Science Publishers: Amsterdam, The Netherlands, 1990; Volume 25, Chapter 7; pp. 133–207. [Google Scholar]
- Loya, Y. Plotless and transect methods. In Coral Reefs: Research Methods; Stoddart, D.R., Johannes, R.E., Eds.; UNESCO: Paris, France, 1978; pp. 197–217. [Google Scholar]
- Cleary, D.F.R.; Suharsono; Hoeksema, B.W. Coral diversity across a disturbance gradient in the Pulau Seribu reef complex off Jakarta, Indonesia. Biodivers. Conserv. 2006, 15, 3653–3674. [Google Scholar] [CrossRef]
- Grimaud, J.; Kulbicki, M. Influence de la distance à l’océan sur les peuplements ichtyologiques des récifs frangeants de Nouvelle-Calédonie. Comptes Rendus Acad. Sci. Ser. III-Sci. Vie-Life Sci. 1998, 321, 923–931. [Google Scholar] [CrossRef]
- Moustaka, M.; Mohring, M.B.; Holmes, T.; Evans, R.D.; Thomson, D.; Nutt, C.; Stoddart, J.; Wilson, S.K. Cross-shelf heterogeneity of coral assemblages in Northwest Australia. Diversity 2019, 11, 15. [Google Scholar] [CrossRef]
- Edmunds, P.J. Patterns in the distribution of juvenile corals and coral reef community structure in St John, US Virgin Islands. Mar. Ecol. Prog. Ser. 2000, 202, 113–124. [Google Scholar] [CrossRef]
- Rogers, C.S.; Fitz, H.C.; Gilnack, M.; Beets, J.; Hardin, J. Scleractinian recruitment patterns at Salt River Canyon, St. Croix, U.S. Virgin Islands. Coral Reefs 1984, 3, 69–76. [Google Scholar] [CrossRef]
- Gleason, M.G. Coral recruitment in Moorea, French Polynesia: The importance of patch type and temporal variation. J. Exp. Mar. Biol. Ecol. 1996, 207, 79–101. [Google Scholar] [CrossRef]
- Carlon, D.B. Depth-related patterns of coral recruitment and cryptic suspention-feeding invertebrates on Guana Island, British Virgin Islands. Bull. Mar. Sci. 2001, 68, 525–541. [Google Scholar]
- Miller, M.W.; Weil, E.; Szmant, A.M. Coral recruitment and juvenile mortality as structuring factors for reef benthic communities in Biscayne National Park, USA. Coral Reefs 2000, 19, 115–123. [Google Scholar] [CrossRef]
- Mundy, C.N.; Babcock, R.C. Are vertical distribution patterns of scleractinian corals maintained by pre- or post-settlement processes? A case study of three contrasting species. Mar. Ecol. Prog. Ser. 2000, 198, 109–119. [Google Scholar] [CrossRef] [Green Version]
- Penin, L.; Adjeroud, M. Relative importance of recruitment and post-settlement processes in the maintenance of coral assemblages in an insular, fragmented reef system. Mar. Ecol. Prog. Ser. 2013, 473, 149–162. [Google Scholar] [CrossRef]
- Baird, A.H.; Babcock, R.C.; Mundy, C.P. Habitat selection by larvae influences the depth distribution of six common coral species. Mar. Ecol. Prog. Ser. 2003, 252, 289–293. [Google Scholar] [CrossRef] [Green Version]
- Penin, L.; Adjeroud, M.; Pratchett, M.S.; Hughes, T.P. Spatial distribution of juvenile and adult corals around Moorea (French Polynesia): Implications for population regulation. Bull. Mar. Sci. 2007, 80, 379–389. [Google Scholar]
- Pichon, M.; Morrissey, J. Benthic zonation and community structure of South Island Reef, Lizard Island (Great Barrier Reef). Bull. Mar. Sci. 1981, 31, 581–593. [Google Scholar]
- Jupiter, S.; Roff, G.; Marion, G.; Henderson, M.; Schrameyer, V.; McCulloch, M.; Hoegh-Guldberg, O. Linkages between coral assemblages and coral proxies of terrestrial exposure along a cross-shelf gradient on the southern Great Barrier Reef. Coral Reefs 2008, 27, 887–903. [Google Scholar] [CrossRef]
- Baum, G.; Januar, H.I.; Ferse, S.C.A.; Kunzmann, A. Local and regional impacts of pollution on coral reefs along the Thousand Islands North of the megacity Jakarta, Indonesia. PLoS ONE 2015, 10, e0138271. [Google Scholar] [CrossRef] [PubMed]
- Kayal, M.; Vercelloni, J.; Lison de Loma, T.; Bosserelle, P.; Chancerelle, Y.; Geoffroy, S.; Stievenart, C.; Michonneau, F.; Penin, L.; Planes, S.; et al. Predator Crown-of-Thorns Starfish (Acanthaster planci) outbreak, mass mortality of corals, and cascading effects on reef fish and benthic communities. PLoS ONE 2012, 7, e47363. [Google Scholar] [CrossRef] [PubMed]
- Darling, E.S.; Alvarez-Filip, L.; Oliver, T.A.; McClanahan, T.R.; Côté, I.M. Evaluating life-history strategies of reef corals from species traits. Ecol. Lett. 2012, 15, 1378–1386. [Google Scholar] [CrossRef]
- Kayal, M.; Vercelloni, J.; Wand, M.P.; Adjeroud, M. Searching for the best bet in life-strategy: A quantitative approach to individual performance and population dynamics in reef-building corals. Ecol. Complex. 2015, 23, 73–84. [Google Scholar] [CrossRef]
- Dollar, S.J.; Grigg, R.W. Anthropogenic and natural stresses on selected coral reefs in Hawai’i: A multidecade synthesis of impact and recovery. Pac. Sci. 2004, 58, 281–304. [Google Scholar] [CrossRef]
- Dumas, P.; Jimenez, H.; Peignon, C.; Wantiez, L.; Adjeroud, M. Small-scale habitat structure modulates the effects of no-take marine reserves for coral reef macroinvertebrates. PLoS ONE 2013, 8, e58998. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Descriptor | 2006 | 2007 | 2008 |
|---|---|---|---|
| Percent cover | 0.043 NS | 0.099 NS | 0.048 NS |
| Generic Richness | |||
| Adults | 0.289 NS | 0.343 NS | 0.321 NS |
| Juveniles | 0.387 NS | 0.518 NS | 0.548 NS |
| Abundance | |||
| Adults | 0.701 * | 0.696 * | 0.733 * |
| Juveniles | 0.815 * | 0.725 * | 0.886 * |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adjeroud, M.; Poisson, E.; Peignon, C.; Penin, L.; Kayal, M. Spatial Patterns and Short-term Changes of Coral Assemblages Along a Cross-shelf Gradient in the Southwestern Lagoon of New Caledonia. Diversity 2019, 11, 21. https://doi.org/10.3390/d11020021
Adjeroud M, Poisson E, Peignon C, Penin L, Kayal M. Spatial Patterns and Short-term Changes of Coral Assemblages Along a Cross-shelf Gradient in the Southwestern Lagoon of New Caledonia. Diversity. 2019; 11(2):21. https://doi.org/10.3390/d11020021
Chicago/Turabian StyleAdjeroud, Mehdi, Emeline Poisson, Christophe Peignon, Lucie Penin, and Mohsen Kayal. 2019. "Spatial Patterns and Short-term Changes of Coral Assemblages Along a Cross-shelf Gradient in the Southwestern Lagoon of New Caledonia" Diversity 11, no. 2: 21. https://doi.org/10.3390/d11020021