1. Introduction
Rice (
Oryza sativa) holds significant economic importance in Africa, with Nigeria being one of the leading contributors to the continent’s global rice production share (4.2%), accounting for 24% [
1]. Despite this, Nigeria remains the second-largest importer of rice worldwide, trailing only behind China. In 2018 alone, Nigeria imported approximately 3 million metric tons of milled rice, struggling to meet its demand deficit for the past decade [
2]. Mali, a landlocked country in West Africa, ranks fifth among African nations in terms of rice production. Its rice production is being managed through irrigated systems connected to the Niger River. In Rwanda, the demand for rice is estimated at 145,000 tons per year, while national supply accounts for about 40%, creating a 60% deficit that is met through imports [
3]. While the rice cultivation area is expanding in Africa, the average yields (2.35 tons/ha in 2021) are low when compared to Asia (4.95 tons/ha in 2021) [
1]. Rice production is constrained by various factors, including biotic stresses such as pests and diseases. Africa has recorded a steady and substantial increase in the incidence of rice diseases such as rice yellow mottle virus (RYMV), rice blast (
Pyricularia oryzae), bacterial leaf blight (
Xanthomonas oryzae pv oryzae), bacterial leaf streak (
Xanthomonas oryzae pv oryzicola), and rice stripe necrosis virus (RSNV) [
4,
5,
6,
7,
8].
Rice sheath rot is an emerging disease worldwide [
9].
Sarocladium oryzae [
10], formerly
Acrocylindrium oryzae, was the first organism to be associated with rice sheath rot symptoms in Taiwan in 1922 [
11].
Sarocladium attenuatum was originally described as a distinct species causing rice sheath rot and was then considered a synonym of
Sarocladium oryzae [
12], but has recently been reestablished as a separate species causing rice sheath rot in Taiwan [
13]. These authors also described a third species that causes sheath rot symptoms on rice called
S. sparsum, which is closely related to
S. oryzae and
S. attenuatum. Sarocladium has been associated with rice sheath rot in thirty-eight countries [
14].
Besides
Sarocladium species,
Fusarium spp. has been associated with the rice sheath rot complex. These mainly comprise isolates in the
F. fujikuroi species complex (FFSC), including
F. proliferatum,
F. verticillioides,
F. incarnatum, and
F. fujikuroi [
15,
16,
17,
18,
19]. In addition, various bacterial species cause rice sheath rot symptoms. The most important one is
Pseudomonas fuscovaginae, which is known to cause sheath brown rot of rice at high altitudes. In Africa, this bacterium has been reported in Burundi [
9,
20] and Madagascar.
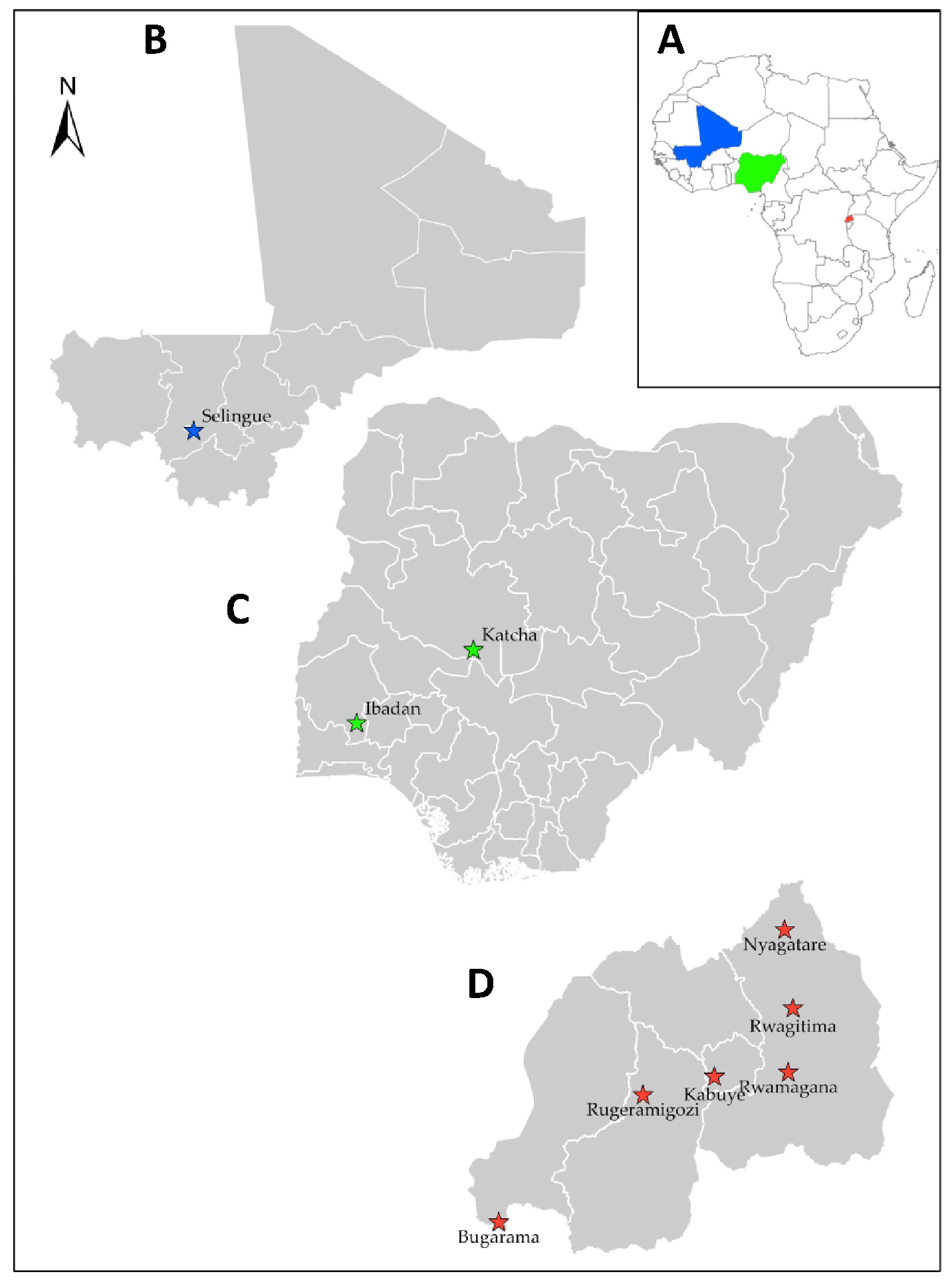
In West Africa, rice sheath rot has been reported in Cote D’ivoire, Gambia, Niger, Nigeria, and Senegal [
14]. However, no causative fungal strains were isolated, and no detailed scientific information was provided except for Nigeria [
21], where
S. attenuatum was first reported as one of the causes of grain discoloration on rice [
21]. In addition, an inhibitory effect of
S. oryzae on seed germination was later observed [
22]. Most information pertaining to the occurrence of
S. oryzae in Africa relating to stored, marketed, and field seeds, especially with respect to mycotoxigenic potentials, was enumerated by [
23].
Rice sheath rot can cause high yield losses of 20–80% [
24,
25,
26]. Furthermore, an extensive survey of rice fields across West Africa enabled the identification of sheath rot symptoms in Mali and Nigeria (AfricaRice disease database). Although yield losses due to the sheath rot disease have not been estimated in Mali and Nigeria, a field survey conducted in 2011 and 2013 revealed the high incidence and severity of the disease in Rwanda [
27].
Mycotoxin contamination of cereal products poses a serious concern for animal and human health. Several studies have reported
Fusarium species as the major producers of mycotoxins contaminating cereals, including rice [
28,
29,
30,
31,
32,
33,
34]. In the African region, previous studies have reported several mycotoxins being synthesized by
Fusarium species isolated from rice as a serious health threat to producers and consumers [
35,
36,
37,
38,
39,
40].
Comprehensive information regarding the incidence and distribution of sheath rot disease is notably lacking. Additionally, there has been a notable absence of research examining the genetic, pathogenic, and toxigenic variability of pathogens associated with this disease in East and West Africa. Acquiring this crucial information will offer valuable insights into disease control and enhance management strategies for breeding programs. Therefore, this study aimed to identify, characterize, as well as assess the genetic, pathogenic, and toxigenic diversity of the pathogens associated with rice sheath rot disease in Mali, Nigeria, and Rwanda.
4. Discussion
This study shows that both
Fusarium and
Sarocladium species are associated with rice sheath rot disease in Nigeria, Rwanda, and Mali and provides insight into their genetic, pathogenic, and toxigenic diversity. Molecular characterization using the
EF-1α gene enabled the delineation of
Fusarium isolates into four distinct
Fusarium species complexes, whereas concatenation of
ITS and
ACT sequences delineated
Sarocladium into three species. Phylogenetic analysis showed that isolates grouped differently according to their geographical location (
Figure 3 and
Figure 4).
Four
Fusarium species complexes (FIESC, FFSC, FOSC, and FSAMSC) were found to be associated with rice sheath rot in SSA. Previous research has resolved FIESC species as a mere complex indicated by numbers, but current studies further updated them according to the new nomenclature, well elucidated from numbers to names [
50,
51,
52]. Additionally, they were characterized using the recently updated
Fusarium ID database (Fusarioid-ID). Dominant in our findings were members of the FIESC previously classified from rice, cereals, insects, and human samples [
29,
33,
34,
52,
53,
54,
55], including
F. sulawesiense (FIESC 16),
F. pernambucatum (FIESC 17),
F. tanahbumbuense (FIESC 24),
F. hainanense (FIESC 26), and
F. incarnatum (FIESC 38). This is similar to those found to be associated with rice sheath rot in Indonesia [
19], India [
17], and the USA [
18]. A larger part of the isolates clustered with
F. sulawesiense, which supports the findings in Brazil and China [
29,
56], while the abundance of
F. hainanense and a few others, such as
F. pernambucatum and
F. tanahbumbuense, were among the FIESC reported on Brazilian rice. There is a wide variation among the species obtained within the FIESC complex, and the observed variation could be correlated with variation in agro-ecological zones. Notable is the fact that some of the
F. sulawesiense isolates from Ibadan (Nigeria) are closely related to strains NTB 1 (rice sheath rot, Indonesia), LC6936 (rice, China), F1 (Sweet potato, US), BT48, and PRT6 (oil palm, Indonesia), and ITEM7547 (
Musa, Bahamas), all of which originated from climates characterized by high temperature and humidity. Isolate F1 originated from Louisiana, USA, which is known for its humid subtropical climate with long, hot, and humid summer, similar to the climate of ITEM7547 from the Bahamas [
34,
57]. Based on our findings, we can hypothesize that the environmental factors could be the driving forces to be considered in the distribution of the FIESC isolates. Notable differences in climate and farming practices could not be underestimated as the three
F. incarnatum isolates from a higher altitude in Rwanda formed part of recently classified FIESC 38 isolates from a similar altitude and climate in Brazil [
29]. Within the FIESC clades, none of the studied isolates was found among the
F. equiseti clade, which disagrees with the previous studies of [
54,
55,
58], whereby variable percentages of both
F. equiseti and
F. incarnatum were reported from rice samples. Members of the
F. equiseti clade are frequent in cereals grown in Western Europe, Turkey, and North America [
34].
FFSC species have been implicated as another causal agent of various rice diseases, including sheath rot [
51,
59].
F. andiyazi and
F. madaense represent the principal species recovered in Rwanda;
F. nygamai was only isolated from Mali, while
F. marum was recovered from Nigeria. Isolates of
F. casha (Rwanda) and
F. annulatum (Mali) were also found among our FFSC. The peculiarity in the clustering of
F. andiyazi,
F. madaense, and
F. marum, clades observed in our studies is comparable to the typical phylogenetic pattern observed in [
60].
F. andiyazi and
F. madaense are typically associated with tropical grasses, including sorghum, maize, millet, and rice, in various parts of the world [
60]. The two virulent
F. marum isolates from Nigeria clearly resolved into a separate clade and clustered with two
F. marum isolates obtained from sorghum in Cameroon [
60]. Isolates clustering with
F. nygamai, the most dominant species in Mali, are closely related to isolates obtained from cereals from regions with similar warm and dry climates (
Figure 4), such as Australia [
61], Italy [
28], Mexico [
62], and Tunisia [
36]. These FIESC and FFSC findings are consistent with the recent report of rice sheath rot disease in Indonesia [
19] and rice disease in China [
56,
63].
Furthermore, isolates clustering with
F. callistephi were found in Rwanda, while
F. triseptatum was found in Mali. Both are members of the FOSC.
F. callistephi is mainly known as a wilt pathogen on ornamentals from the Asteraceae family [
64]. Lastly, members of FSAMSC, which includes two isolates clustering with
F. acasiae-mearnsii of Rwanda origin, were also identified (
Figure 4).
F. acasiae-mearnsii isolates have previously been found in Australia and South Africa and can cause head blight on wheat [
65].
Reports on mycotoxins produced by
Fusarium species isolated from rice sheath rot disease in sub-Saharan Africa are very limited. In consequence, this is the first study to investigate the toxigenic potentials of
Fusarium isolates from rice sheath rot disease in Mali, Nigeria, and Rwanda. The predominant mycotoxins found in FIESC isolates from Nigeria were trichothecenes, while the three
F. incarnatum isolates from Rwanda did not produce mycotoxins. Among the 12 FIESC isolates collected from Ibadan (derived savannah region) in Nigeria, type A (DAS, NEO) and B (NIV, FUS-X) trichothecenes were detected in seven samples. This confirms the previous mycotoxins findings on cereals [
29,
58,
66]. In addition, a huge ZEN production of 32,529 and 26,173 µg/kg was detected from the two
F. hainanense isolates collected from Ibadan. Similar results for
F. hainanense were obtained in Brazilian rice [
29]. This also corroborates the study of [
67], which demonstrated that ZEN production by
Fusarium species is greater in moldy samples, which is favored by wet climates with high rainfall and high humidity. Within the FFSC,
F. andiyazi isolates did not produce mycotoxins, while the
F. annulatum isolate and three out of five
F. nygamai isolates from the dry and hot Sudan Guinea Savannah of Mali produced trichothecenes and fumonisins (FB1, FB2, and FB3), respectively. This is consistent with the findings of [
68] reporting high levels of toxins for
F. nygamai, while
F. andiyazi isolates produced little or no mycotoxins. Moreover, fumonisin producers were not detected among the isolates collected from Nigeria and Rwanda. Thus, fumonisin contamination may be expected to be higher in samples collected in the Sahel with a warm and dry climate. The development of fumonisins in cereal crops prior to harvest might increase due to heat and water stress that characterized the environmental drought [
69]. Only two of the 23
Fusarium isolates from Rwanda produced mycotoxins: trichothecenes (NIV and Fus-X) and ZEN were detected in a
F. madaense isolate and a
F. acaciae-mearnsii isolate, both obtained from the Rwamagana district. It has been shown before that
F. acacia-mearnsii isolates can produce NIV [
70] and ZEN [
71].
According to the research of [
72], which elucidated the presence and absence of biosynthetic gene clusters responsible for the synthesis of mycotoxins and secondary metabolites in FIESC, further studies are necessary to investigate if the mycotoxin production potentials of our isolates agree with their genetic profile or assess if there are differences in expression level. In conclusion, mycotoxin production is common in rice-derived
Fusarium isolates from Nigeria (12 out of 15) and Mali (5 out of 7) but rare in Rwanda (2 out of 25).
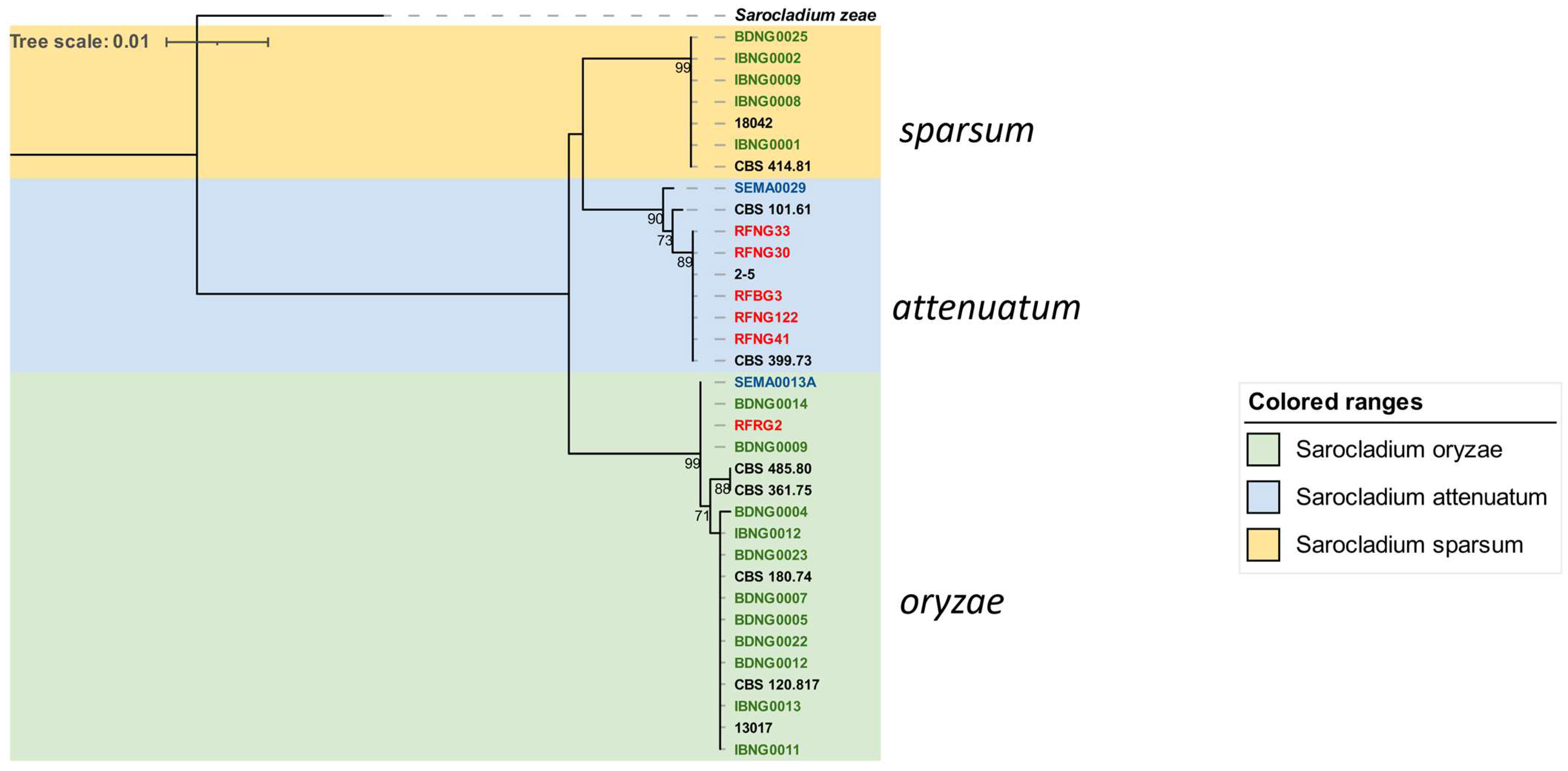
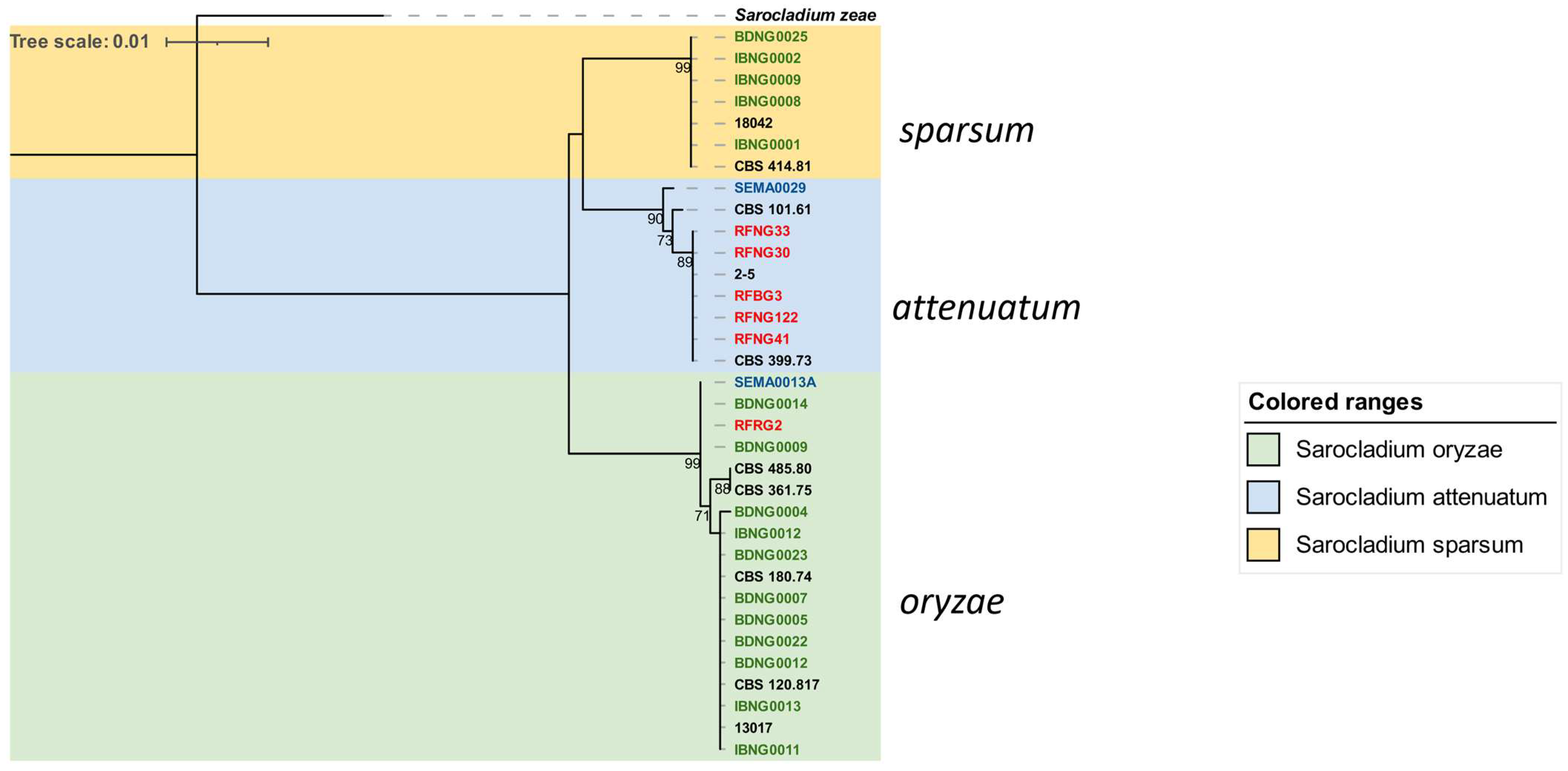
In contrast to the heterogeneity observed among
Fusarium species, three clearly delineated
Sarocladium species were recovered from the three countries of study, but with a lower frequency of occurrence in Mali and Rwanda. Following the characterization of
Sarocladium species causing rice sheath rot in Taiwan by [
13] and using a concatenation of two genes, we were able to resolve our isolates into three species, namely,
attenuatum,
oryzae, and
sparsum (
Figure 4). Isolates belonging to
S. sparsum were only found in Nigeria and mostly originated from Ibadan (DS), except for an isolate (BDNG0025) from an infected seed in Katcha (SGS). Finding a larger part of this group in Ibadan was not unexpected because it clusters with an Ibadan-origin reference isolate CBS 414.81, collected and reported as
S. attenuatum [
21], and later deposited into the Genbank [
44]. Our results clearly show, however, that isolate CBS 414.81 belongs to
sparsum species. Surprisingly, this group was not found in Mali and Rwanda. There occurs a notable correlation between this group and the collection region, which proves that geographical area and climate are the most crucial factors that influence the occurrence of these pathotypes and their virulence. This agrees with the hypothesis that isolates from different locations may also vary in their level of aggressiveness [
73,
74].
S. oryzae isolates (
Figure 4) showed a strong intra-species similarity that is not phylogeographically based. Isolates in this species were the most predominant and widely distributed. It consists of 18 similar isolates from nearly all the rice-growing regions in the world. Most isolates from Nigeria (11 of 16) belong to this group. An isolate from Mali (SEMA0013A) generated from this study, isolates from previous studies on rice, such as 13017 from Taiwan [
13], CBS 180.74 from India, African isolates CBS 361.75 from Kenya [
75], RFRG2 from Rwanda (this study), Central American CBS 120.817 from Panama, and CBS 485.80 Australia [
76], are part of this group. They were found in two agro-ecological zones of Nigeria, although more frequently in Katcha than Ibadan.
The presence of S. oryzae in all the rice-growing regions of the world is a signal of its flexibility to adjust to various agro-ecological zones. It also suggests a link between its dispersal, rice movement, and international trade. A potential quarantine threat is of great concern with the rapid distribution of this group. This might also imply that the origin of the isolate may be connected to Asia. It should be noted that African countries, including Mali, Nigeria, and Rwanda, are major importers of rice from Asia despite Nigeria being the highest producer on the continent.
A third distinct group, S. attenuatum, was dominated by Rwandan isolates; the second Mali isolate, SEMA0029, also formed part of the group.
It is important to note that the two
Sarocladium isolates obtained from Mali formed two of the three species found in this study despite the small sample size. Several reasons might be responsible, from accession variability, as local rice is mostly cultivated in Mali [
77], to toxigenic variability among the competing
Fusarium species, and multiple cropping system variations, among others. Larger-scale surveys across the country are necessary to have a wider knowledge of genetic diversity and distribution.
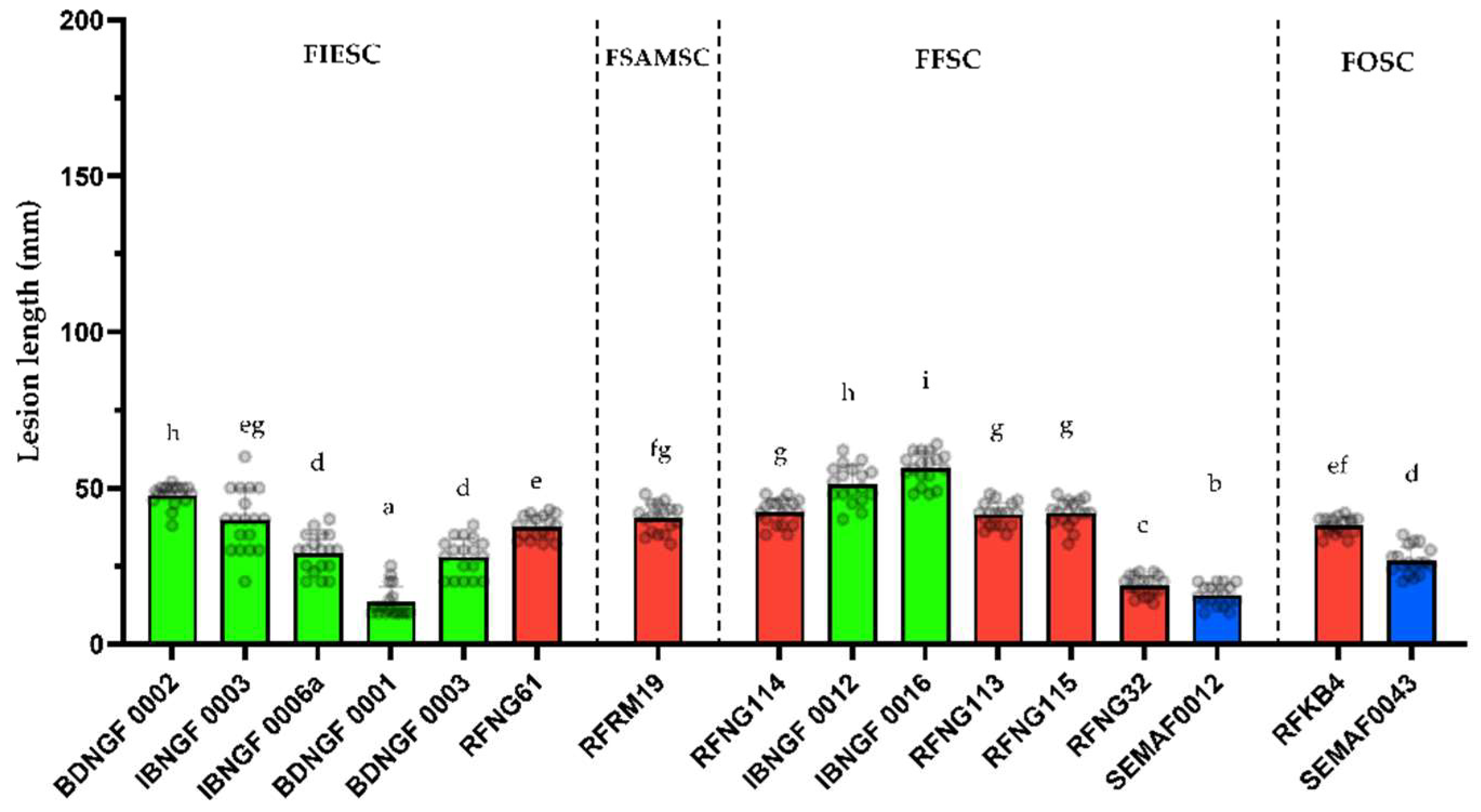
It is a well-known phenomenon that several factors such as variations in climate, topography, and farming practices, among others, are the drivers of variation in pathogen populations. To verify this claim, this study has revealed a strong ecology-driven diversification among the
Sarocladium species used. It also exposed how environmental variation was able to influence genetic and virulence relatedness. The
S. sparsum isolates that are mainly found in Ibadan (DS) are clearly more aggressive on the rice FARO44 cultivar used in this study than the
S. oryzae and
S. attenuatum strains, which are more common in the savannah region in Katcha-Badeggi and in Rwanda. The same trend of aggressiveness was reported in the study of [
21], who used four isolates (presumably
S. sparsum) collected from the southern region (DS and Humid Forest) of Nigeria. This confirms previous work showing that group 3 strains (=
S. sparsum) are more aggressive on the rice
japonica cultivar Kitaake than group 1 (=
S. oryzae) or group 2 (=
S. attenuatum) strains.
S. sparsum isolates also produce high amounts of the toxin helvolic acid
in planta which is clearly correlated with disease severity [
78].

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}