
Ecological Impacts of Introduced European Rabbits (Oryctolagus cuniculus) on Island Ecosystems in the Mediterranean
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
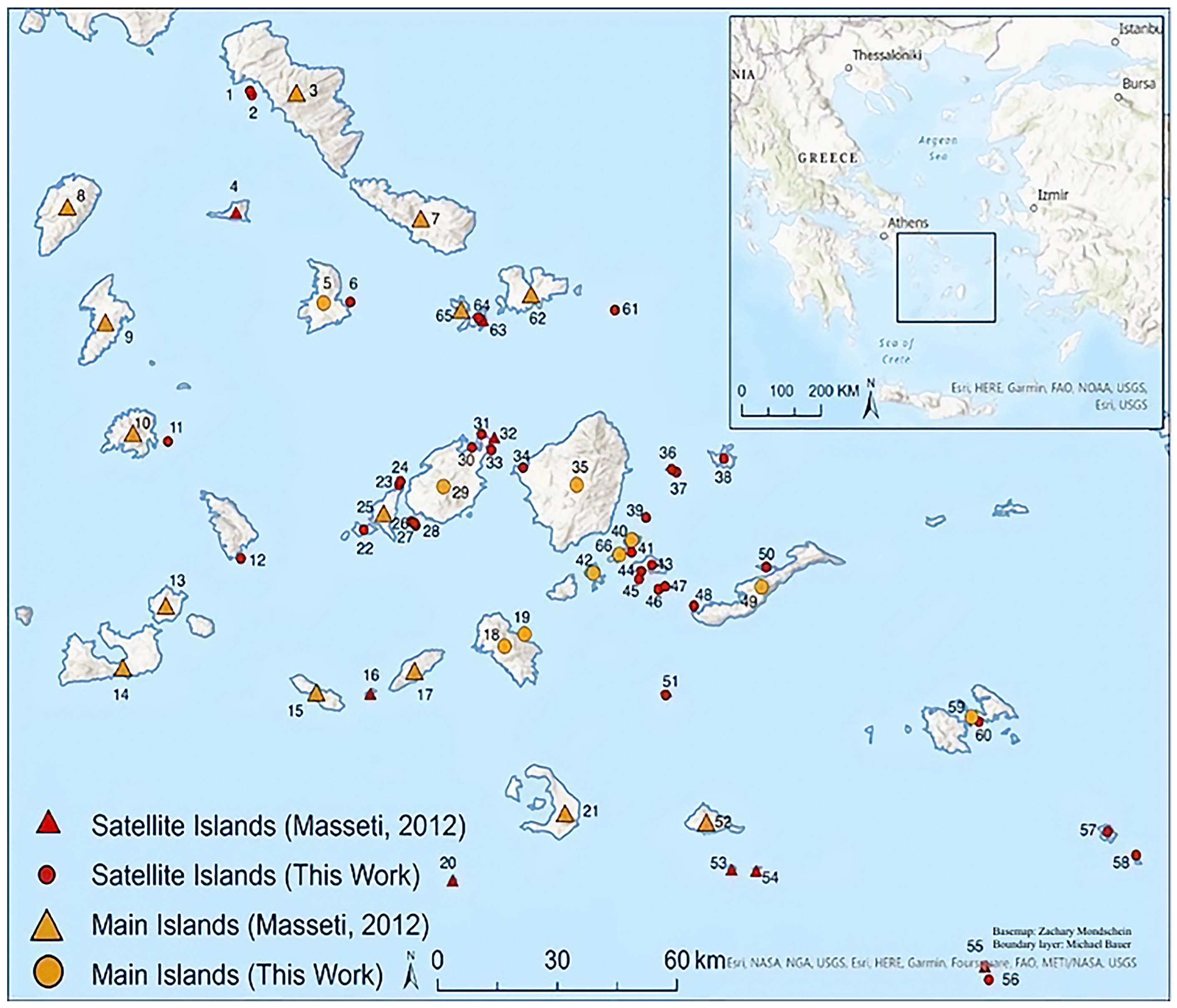
2.1. Study Area
2.2. Grazing Intensity
2.3. Vegetation Assessment
2.4. Arthropod Community Assessment
2.5. Soil Characteristics
2.6. Statistical Analyses
3. Results
3.1. Survey
3.2. Grazing Intensity
3.3. Vegetation
3.3.1. Ground Cover
3.3.2. Vegetation Height
3.3.3. Vegetation Biomass
3.3.4. Vegetation Species Richness
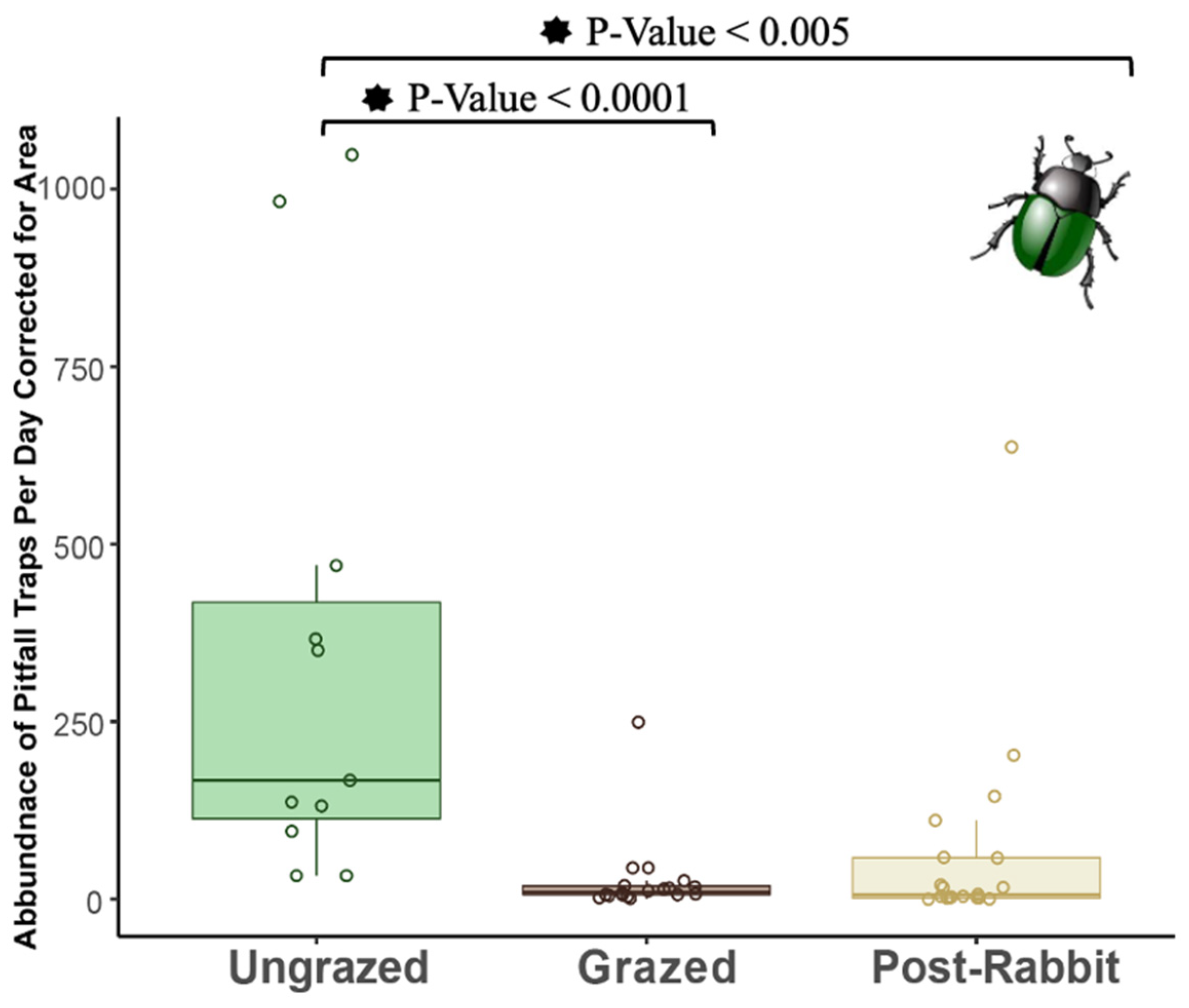
3.4. Arthropods
3.5. Soil
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Agia Kali | Grambonisi | Tourlos | Glarombi | Lower Fira | Panterionisi | Filitzi | Galiastos | Tigani | Gramvousa | Upper Fira | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Grazing Status | Ungrazed | Ungrazed | Ungrazed | Grazed | Grazed | Grazed | Grazed | Post-rabbit | Post-rabbit | Post-rabbit | Post-rabbit |
| Area (km2) | 0.01 | 0.15 | 0.03 | 0.21 | 0.46 | 0.48 | 0.04 | 0.01 | 0.08 | 0.79 | 0.26 |
| Vegetation Height (cm) | 15.26 ± 4.22 | 35.5 ± 5.21 | 13.35 ± 1.62 | 12.81 ± 3.40 | 29.72 ± 5.66 | 13.46 ± 5.28 | 13.74 ± 4.88 | 3.74 ± 2.85 | 6.02 ± 1.05 | 9.40 ± 5.28 | 24.12 ± 9.84 |
| Observed Plant Species | 12.00 | 15.00 | 8.00 | 9.00 | 8.00 | 11.00 | 9.00 | 8.00 | 10.00 | 17.00 | 8.00 |
| Chao1 Bias-Corrected Estimate (plant) | 14.55 | 16.99 | 9.76 | 9.00 | 6.00 | 17.50 | 14.75 | 14.82 | 13.62 | 75.14 | 7.47 |
| Average Aboveground Biomass (g) | 183.2 ± 98.24 | 417.99 ± 226.35 | 443.60 ± 112.45 | 63.06 ± 29.39 | 216.20 ± 145.96 | 120.80 ± 57.7 | 314.98 ± 151.52 | 111.6 ± 50.64 | 175.38 ± 106.22 | 278.89 ± 188.08 | 218 ± 44.56 |
| Average Soil Depth (cm) | 31.98 | 25.64 | 10.03 | 8.40 | N/A | 13.12 | 10.68 | 9.24 | 4.78 | 4.09 | N/A |
| Observed Pitfall Trap Species | 9.00 | 4.00 | 11.00 | 8.00 | 11.00 | 10.00 | 7.00 | 3.00 | 5.00 | 7.00 | 6.00 |
| Average Arthropod Biomass per day (g) | 0.036 | 0.244 | 0.085 | 0.020 | 0.024 | 0.199 | 0.008 | 0.022 | 0.040 | 0.159 | 0.003 |
| Arthropod Abundance per day (pitfall) | 24.79 | 20.50 | 46.97 | 4.75 | 37.33 | 21.00 | 13.14 | 10.81 | 14.40 | 7.33 | 11.20 |
| Arthropod Abundance per day (sticky) | 14.14 ± 2.28 | 39.1 ± 11.90 | 11.51 ± 3.32 | 6.75 ± 3.93 | 11.84 ± 2.36 | 26.9 ± 4.70 | 6.63 ± 1.88 | 23.73 ± 3.70 | 6.7 ± 0.93 | 30.53 ± 6.45 | 37.01 ± 14.35 |
| Percent Vegetation Cover | 53.33% ± 3.6 | 79.57% ± 5.01 | 55% ± 3.97 | 41.67% ± 7.39 | 56.67% ± 8.05 | 35% ± 5.53 | 41.67% ± 6.16 | 24.17% ± 1.6 | 45.83% ± 6.29 | 44.27% ± 4.85 | 54.17% ± 16.52 |
| Number | Island | Documented By | Latitude | Longitude |
|---|---|---|---|---|
| 1 | Akamatis | JF | 37.856210 | 24.746671 |
| 2 | Megalo | JF | 37.848055 | 24.750482 |
| 3 | Andros | Masseti | 37.854974 | 24.851252 |
| 4 | Giaros | Masseti | 37.611585 | 24.715466 |
| 5 | Syros | JF | 37.424795 | 24.912170 |
| 6 | Dydimi | JF | 37.426787 | 24.973094 |
| 7 | Tinos | Masseti | 37.599734 | 25.130841 |
| 8 | Kea | Masseti | 37.623806 | 24.336106 |
| 9 | Kithnos | Masseti | 37.387624 | 24.421376 |
| 10 | Serifos | Masseti | 37.162023 | 24.483325 |
| 11 | Vous | JF | 37.142318 | 24.561666 |
| 12 | Kitriani | JF | 36.904085 | 24.726438 |
| 13 | Kimolos | Masseti | 36.809761 | 24.557400 |
| 14 | Milos | Masseti | 36.683792 | 24.460690 |
| 15 | Folegandros | Masseti | 36.633207 | 24.896052 |
| 16 | Kardiotisa | Masseti | 36.629623 | 25.017722 |
| 17 | Sikinos | Masseti | 36.676971 | 25.116797 |
| 18 | Ios | JF | 36.724885 | 25.319705 |
| 19 | Psathonisi | JF | 36.749432 | 25.364105 |
| 20 | Cristiana | Masseti | 36.249606 | 25.202587 |
| 21 | Santorini | Masseti | 36.387112 | 25.455643 |
| 22 | Despotiko | JF | 36.962468 | 25.002743 |
| 23 | Lower Fira | JF | 37.054512 | 25.082179 |
| 24 | Upper Fira | JF | 37.061153 | 25.085618 |
| 25 | Antiparos | Masseti | 36.998277 | 25.047446 |
| 26 | Glarombi | JF | 36.979221 | 25.109781 |
| 27 | Tigani | JF | 36.976707 | 25.116035 |
| 28 | Panterionisi | JF | 36.971061 | 25.119186 |
| 29 | Paros | JF | 37.050244 | 25.181891 |
| 30 | Galiatsos | JF | 37.130813 | 25.245973 |
| 31 | Gaidouronisi | JF | 37.157282 | 25.268061 |
| 32 | Ovriokastro | Masseti | 37.152427 | 25.296432 |
| 33 | Filitzi | JF | 37.124964 | 25.289978 |
| 34 | Mando | JF | 37.089253 | 25.361565 |
| 35 | Naxos | JF | 37.054048 | 25.482152 |
| 36 | Macheres | JF | 37.085339 | 25.695641 |
| 37 | Agia Paraskevi | JF | 37.079723 | 25.70582 |
| 38 | Donousa | JF | 37.107447 | 25.812942 |
| 39 | Prasoura | JF | 36.986712 | 25.638222 |
| 40 | Ano Koufonisi | JF | 36.941124 | 25.606195 |
| 41 | Glaronisi | JF | 36.916409 | 25.605192 |
| 42 | Schoinoussa | JF | 36.873949 | 25.519387 |
| 43 | Keros | JF | 36.890691 | 25.651398 |
| 44 | Megali Plaka | JF | 36.877677 | 25.626787 |
| 45 | Andreas | JF | 36.861629 | 25.621933 |
| 46 | Kato Antikeri | JF | 36.841104 | 25.665571 |
| 47 | Pano Antikeri | JF | 36.846570 | 25.680731 |
| 48 | Gramvousa | JF | 36.807258 | 25.745579 |
| 49 | Amorgos | JF | 36.846272 | 25.898340 |
| 50 | Nikouria | JF | 36.886292 | 25.908540 |
| 51 | Anydros | JF | 36.625212 | 25.682358 |
| 52 | Anafi | Masseti | 36.368622 | 25.773609 |
| 53 | Pachia | Masseti | 36.271722 | 25.830563 |
| 54 | Makria | Masseti | 36.269564 | 25.886052 |
| 55 | Megalo Sofrano | JF | 36.075218 | 26.400744 |
| 56 | Mikro Sofrano | JF | 36.046704 | 26.409475 |
| 57 | Syrna | JF | 36.347422 | 26.676584 |
| 58 | Mesonisi | JF | 36.299744 | 26.740516 |
| 59 | Astypalea | JF | 36.580607 | 26.370291 |
| 60 | Diapori | JF | 36.570960 | 26.387153 |
| 61 | Chtapodia | JF | 37.410302 | 25.567967 |
| 62 | Mykonos | Masseti | 37.444814 | 25.379182 |
| 63 | Delos | Masseti | 37.391613 | 25.271329 |
| 64 | Megalos Revmatiaris | JF | 37.394672 | 25.260716 |
| 65 | Rhinia | Masseti | 37.413464 | 25.222172 |
| 66 | Kato Koufonisi | JF | 36.912094 | 25.577397 |

References
- Sayre, R.; Noble, S.; Hamann, S.; Smith, R.; Wright, D.; Breyer, S.; Butler, K.; Van Graafeiland, K.; Frye, C.; Karagulle, D. A new 30 meter resolution global shoreline vector and associated global islands database for the development of standardized ecological coastal units. J. Oper. Oceanogr. 2019, 12 (Suppl. S2), S47–S56. [Google Scholar] [CrossRef]
- Fernández-Palacios, J.M.; Kreft, H.; Irl, S.D.; Norder, S.; Ah-Peng, C.; Borges, P.A.; Burns, K.C.; de Nascimento, L.; Meyer, J.-Y.; Montes, E. Scientists’ warning–The outstanding biodiversity of islands is in peril. Glob. Ecol. Conserv. 2021, 31, e01847. [Google Scholar] [CrossRef] [PubMed]
- Kier, G.; Kreft, H.; Lee, T.M.; Jetz, W.; Ibisch, P.L.; Nowicki, C.; Mutke, J.; Barthlott, W. A global assessment of endemism and species richness across island and mainland regions. Proc. Natl. Acad. Sci. USA 2009, 106, 9322–9327. [Google Scholar] [CrossRef] [PubMed]
- Vié, J.-C.; Hilton-Taylor, C.; Stuart, S.N. Wildlife in a Changing World: An Analysis of the 2008 IUCN Red List of Threatened Species; IUCN: Fontainebleau, France, 2009. [Google Scholar]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Leclerc, C.; Courchamp, F.; Bellard, C. Insular threat associations within taxa worldwide. Sci. Rep. 2018, 8, 6393. [Google Scholar] [CrossRef] [PubMed]
- Cuttelod, A.; García, N.; Malak, D.A.; Temple, H.J.; Katariya, V. The Mediterranean: A biodiversity hotspot under threat. In The 2008 Review of The IUCN Red List of Threatened Species; IUCN: Gland, Switzerland, 2009; pp. 89–101. [Google Scholar]
- Gratsia, I. Citizen and monument. The case of the Greek island of Naxos. Conserv. Manag. Archaeol. Sites 2010, 12, 75–86. [Google Scholar] [CrossRef]
- Salladay, R.A.; Ramirez, A.R. Reduced defenses and increased herbivore preference of island chaparral shrubs compared to mainland relatives. West. North Am. Nat. 2018, 78, 768–776. [Google Scholar] [CrossRef]
- Bergmeier, E.; Dimopoulos, P. The vegetation of islets in the Aegean and the relation between the occurrence of islet specialists, island size, and grazing. Phytocoenologia 2003, 33, 447–474. [Google Scholar] [CrossRef]
- Marín-García, P.J.; Llobat, L. What are the keys to the adaptive success of European wild rabbit (Oryctolagus cuniculus) in the Iberian Peninsula? Animals 2021, 11, 2453. [Google Scholar] [CrossRef]
- Courchamp, F.; Langlais, M.; Sugihara, G. Control of rabbits to protect island birds from cat predation. Biol. Conserv. 1999, 89, 219–225. [Google Scholar] [CrossRef]
- Carlberg, R.A.; Karris, G.; Verma, M.; Foufopoulos, J. Food versus Disturbance: Contradictory Effects of Human Activities on an Opportunistic Seabird Breeding in an Oligotrophic Marine System. Diversity 2022, 14, 421. [Google Scholar] [CrossRef]
- Cubas, J.; Irl, S.D.; Villafuerte, R.; Bello-Rodríguez, V.; Rodríguez-Luengo, J.L.; Del Arco, M.; Martín-Esquivel, J.L.; González-Mancebo, J.M. Endemic plant species are more palatable to introduced herbivores than non-endemics. Proc. R. Soc. B 2019, 286, 20190136. [Google Scholar] [CrossRef] [PubMed]
- Aguirre-Muñoz, A.; Croll, D.A.; Donlan, C.J.; Henry, R.W.; Hermosillo, M.A.; Howald, G.R.; Keitt, B.S.; Luna-Mendoza, L.; Rodríguez-Malagón, M.; Salas-Flores, L.M. High-impact conservation: Invasive mammal eradications from the islands of western Mexico. AMBIO A J. Hum. Environ. 2008, 37, 101–107. [Google Scholar] [CrossRef]
- Marchant, R.; Kefford, B.; Wasley, J.; King, C.; Doube, J.; Nugegoda, D. Response of stream invertebrate communities to vegetation damage from overgrazing by exotic rabbits on subantarctic Macquarie Island. Mar. Freshw. Res. 2011, 62, 404–413. [Google Scholar] [CrossRef]
- North, S.; Bullock, D.; Dulloo, M. Changes in the vegetation and reptile populations on Round Island, Mauritius, following eradication of rabbits. Biol. Conserv. 1994, 67, 21–28. [Google Scholar] [CrossRef]
- Eldridge, D.J.; Myers, C.A. The impact of warrens of the European rabbit (Oryctolagus cuniculus L.) on soil and ecological processes in a semi-arid Australian woodland. J. Arid. Environ. 2001, 47, 325–337. [Google Scholar] [CrossRef]
- Lees, A.C.; Bell, D.J. A conservation paradox for the 21st century: The European wild rabbit Oryctolagus cuniculus, an invasive alien and an endangered native species. Mammal Rev. 2008, 38, 304–320. [Google Scholar] [CrossRef]
- De Tournefort, J.P. A Voyage into the Levant; Cambridge University Press: Cambridge, UK, 2014; Volume 2. [Google Scholar]
- Erhard, T. Fauna Der Cykladen; Voigt & Günther: Leipzig, Germany, 1858. [Google Scholar]
- Kontsiotis, V.J.; Bakaloudis, D.; Merou, T.; Xofis, P. Trophic ecology of the European wild rabbit Oryctolagus cuniculus on the Mediterranean island of Lemnos, Greece. Ecol. Res. 2015, 30, 683–691. [Google Scholar] [CrossRef]
- Kontsiotis, V.; Bakaloudis, D.; Xofis, P.; Konstantaras, N.; Petrakis, N.; Tsiompanoudis, A. Modeling the distribution of wild rabbits (Oryctolagus cuniculus) on a Mediterranean island. Ecol. Res. 2013, 28, 317–325. [Google Scholar] [CrossRef]
- Gizicki, Z.S.; Tamez, V.; Galanopoulou, A.P.; Avramidis, P.; Foufopoulos, J. Long-term effects of feral goats (Capra hircus) on Mediterranean island communities: Results from whole island manipulations. Biol. Invasions 2018, 20, 1537–1552. [Google Scholar] [CrossRef]
- Bello-Rodríguez, V.; Mateo, R.G.; Pellissier, L.; Cubas, J.; Cooke, B.; González-Mancebo, J.M. Forecast increase in invasive rabbit spread into ecosystems of an oceanic island (Tenerife) under climate change. Ecol. Appl. 2021, 31, e02206. [Google Scholar] [CrossRef]
- Moemken, J.; Reyers, M.; Feldmann, H.; Pinto, J.G. Future changes of wind speed and wind energy potentials in EURO-CORDEX ensemble simulations. J. Geophys. Res. Atmos. 2018, 123, 6373–6389. [Google Scholar] [CrossRef]
- Hao, L.; Pan, C.; Fang, D.; Zhang, X.; Zhou, D.; Liu, P.; Liu, Y.; Sun, G. Quantifying the effects of overgrazing on mountainous watershed vegetation dynamics under a changing climate. Sci. Total Environ. 2018, 639, 1408–1420. [Google Scholar] [CrossRef]
- Haarsma, R.J.; Selten, F.; Hurk, B.v.; Hazeleger, W.; Wang, X. Drier Mediterranean soils due to greenhouse warming bring easterly winds over summertime central Europe. Geophys. Res. Lett. 2009, 36. [Google Scholar] [CrossRef]
- Ulbrich, U.; May, W.; Li, L.; Lionello, P.; Pinto, J.G.; Somot, S. The Mediterranean climate change under global warming. In Developments in Earth and Environmental Sciences; Elsevier: Amsterdam, The Netherlands, 2006; Volume 4, pp. 399–415. [Google Scholar]
- Bloom, A.; Kotroni, V.; Lagouvardos, K. Climate change impact of wind energy availability in the Eastern Mediterranean using the regional climate model PRECIS. Nat. Hazards Earth Syst. Sci. 2008, 8, 1249–1257. [Google Scholar] [CrossRef]
- Cheng, W.-C. Impact of Livestock Grazing on Ecosystem Services in a Mediterranean Ecosystem. Ph.D. Thesis, University of Michigan, Ann Arbor, Michigan, 2015. [Google Scholar]
- Masseti, M. Atlas of Terrestrial Mammals of the Ionian and Aegean Islands; Walter de Gruyter: Berlin, Germany, 2012. [Google Scholar]
- Médail, F. Plant biogeography and vegetation patterns of the Mediterranean islands. Bot. Rev. 2022, 88, 63–129. [Google Scholar] [CrossRef]
- Lafranchis, T.; Sfikas, G. Flowers of Greece. 2 vols. Paris (Dia Theo) 2009, 431, 447. [Google Scholar]
- Fielding, J.; Turland, N.J.; Mathew, B. Flowers of Crete; Royal Botanic Gardens, Kew: Richmond, UK, 2005. [Google Scholar]
- Snogerup, S.; Snogerup, B. Repeated floristical observations on islets in the Aegean. Plant Syst. Evol. 1987, 155, 143–164. [Google Scholar] [CrossRef]
- Snogerup, S.; Snogerup, B. Changes in the flora of some Aegean islets 1968–2000. Plant Syst. Evol. 2004, 245, 169–213. [Google Scholar] [CrossRef]
- Schmidt, M.H.; Clough, Y.; Schulz, W.; Westphalen, A.; Tscharntke, T. Capture efficiency and preservation attributes of different fluids in pitfall traps. J. Arachnol. 2006, 34, 159–162. [Google Scholar] [CrossRef]
- Avramidis, P.; Nikolaou, K.; Bekiari, V. Total organic carbon and total nitrogen in sediments and soils: A comparison of the wet oxidation–titration method with the combustion-infrared method. Agric. Agric. Sci. Procedia 2015, 4, 425–430. [Google Scholar] [CrossRef]
- Avramidis, P.; Bekiari, V. Application of a catalytic oxidation method for the simultaneous determination of total organic carbon and total nitrogen in marine sediments and soils. PLoS ONE 2021, 16, e0252308. [Google Scholar] [CrossRef] [PubMed]
- Bekiari, V.; Avramidis, P. Data quality in water analysis: Validation of combustion-infrared and combustion-chemiluminescence methods for the simultaneous determination of Total Organic Carbon (TOC) and Total Nitrogen (TN). Int. J. Environ. Anal. Chem. 2014, 94, 65–76. [Google Scholar] [CrossRef]
- Rstudio Team. RStudio: Integrated Development for R; PBC: Boston, MA, USA, 2022. [Google Scholar]
- Chao, A.; Ma, K.; Hsieh, T.; Chiu, C.-H. User’s guide for online program SpadeR (Species-richness prediction and diversity estimation in R). Natl. Tsing Hua Univ. Hsinchu Taiwan 2016, 88. [Google Scholar]
- Chao, A.; Colwell, R.K.; Lin, C.-W.; Gotelli, N.J. Sufficient sampling for asymptotic minimum species richness estimators. Ecology 2009, 90, 1125–1133. [Google Scholar] [CrossRef] [PubMed]
- Macarthur, R.H.; Wilson, E.O. The Theory of Island Biogeography; Princeton University Press: Princeton, NJ, USA, 1967. [Google Scholar]
- Rocha, R.; Sequeira, M.M.; Douglas, L.R.; Gouveia, M.; Jardim, R.; Jesus, J.; Jones, H.P.; Russo, D. Extinctions of introduced game species on oceanic islands: Curse for hunters or conservation opportunities? Biodivers. Conserv. 2017, 26, 2517–2520. [Google Scholar] [CrossRef]
- Agnew, A.; Flux, J.E. Plant dispersal by hares (Lepus capensis L.) in Kenya. Ecology 1970, 51, 735–737. [Google Scholar] [CrossRef]
- Tzanoudakis, D.; Panitsa, M. The flora of the Greek islands. Ecol. Mediterr. 1995, 21, 195–212. [Google Scholar] [CrossRef]
- Greuter, W. Origin and peculiarities of Mediterranean island floras. Ecol. Mediterr. 1995, 21, 1–10. [Google Scholar] [CrossRef]
- Bowen, L.; Van Vuren, D. Insular endemic plants lack defenses against herbivores. Conserv. Biol. 1997, 11, 1249–1254. [Google Scholar] [CrossRef]
- Cubas, J.; Martín-Esquivel, J.L.; Nogales, M.; Irl, S.D.; Hernández-Hernández, R.; López-Darias, M.; Marrero-Gómez, M.; del Arco, M.J.; González-Mancebo, J.M. Contrasting effects of invasive rabbits on endemic plants driving vegetation change in a subtropical alpine insular environment. Biol. Invasions 2018, 20, 793–807. [Google Scholar] [CrossRef]
- van Klink, R.; van der Plas, F.; Van Noordwijk, C.; WallisDeVries, M.F.; Olff, H. Effects of large herbivores on grassland arthropod diversity. Biol. Rev. 2015, 90, 347–366. [Google Scholar] [CrossRef] [PubMed]
- Van Breemen, N.; Buurman, P. Soil Formation; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2002. [Google Scholar]
- Copson, G.; Whinam, J. Review of ecological restoration programme on subantarctic Macquarie Island: Pest management progress and future directions. Ecol. Manag. Restor. 2001, 2, 129–138. [Google Scholar] [CrossRef]
- Zavaleta, E.S.; Hobbs, R.J.; Mooney, H.A. Viewing invasive species removal in a whole-ecosystem context. Trends Ecol. Evol. 2001, 16, 454–459. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kossoff, A.; Schueller, S.; Nossan, H.; Slack, I.; Avramidis, P.; Foufopoulos, J. Ecological Impacts of Introduced European Rabbits (Oryctolagus cuniculus) on Island Ecosystems in the Mediterranean. Diversity 2024, 16, 244. https://doi.org/10.3390/d16040244
Kossoff A, Schueller S, Nossan H, Slack I, Avramidis P, Foufopoulos J. Ecological Impacts of Introduced European Rabbits (Oryctolagus cuniculus) on Island Ecosystems in the Mediterranean. Diversity. 2024; 16(4):244. https://doi.org/10.3390/d16040244
Chicago/Turabian StyleKossoff, Angelina, Sheila Schueller, Hannah Nossan, Ian Slack, Pavlos Avramidis, and Johannes Foufopoulos. 2024. "Ecological Impacts of Introduced European Rabbits (Oryctolagus cuniculus) on Island Ecosystems in the Mediterranean" Diversity 16, no. 4: 244. https://doi.org/10.3390/d16040244