
Determination of Genetic Variations of Toll-Like Receptor (TLR) 2, 4, and 6 with Next-Generation Sequencing in Native Cattle Breeds of Anatolia and Holstein Friesian †


Abstract
:1. Introduction
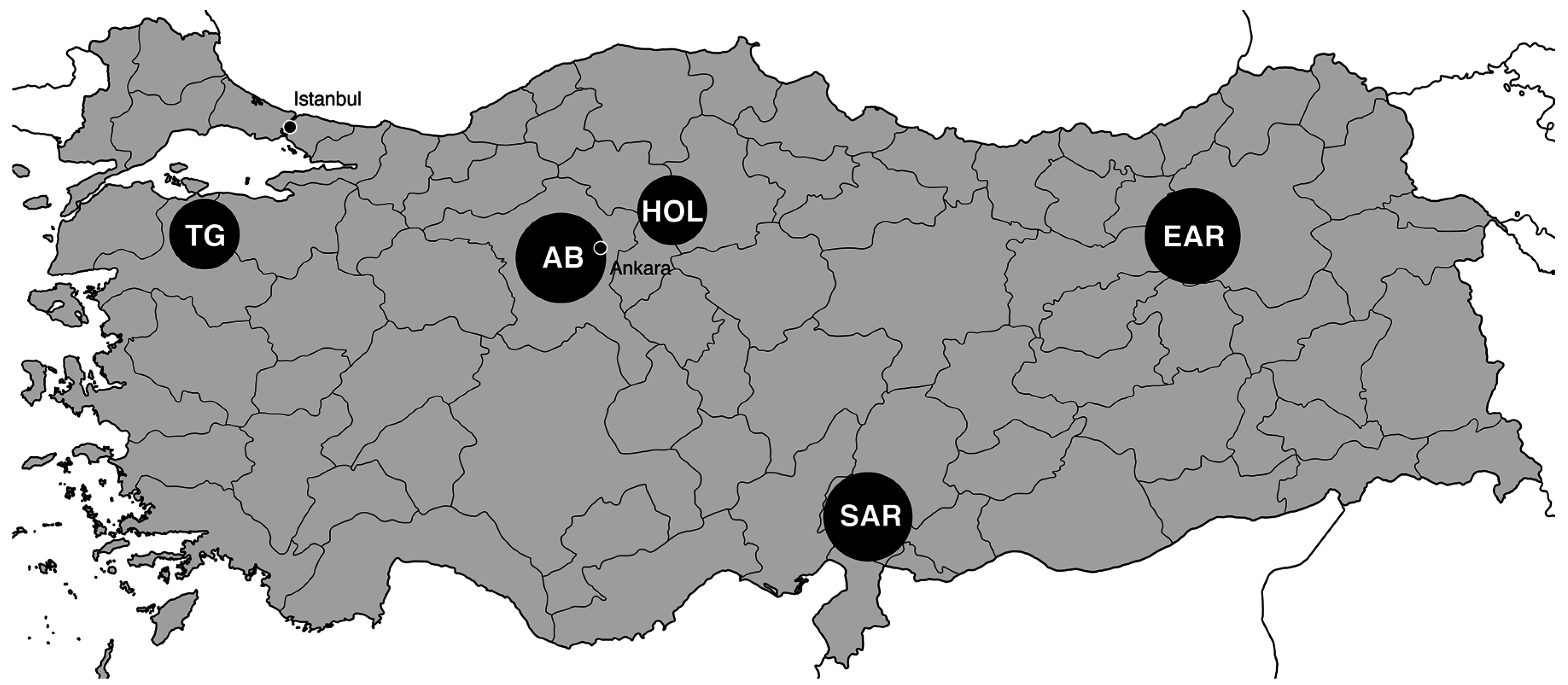
2. Materials and Methods
2.1.DNA Isolation and Amplicon Sequencing
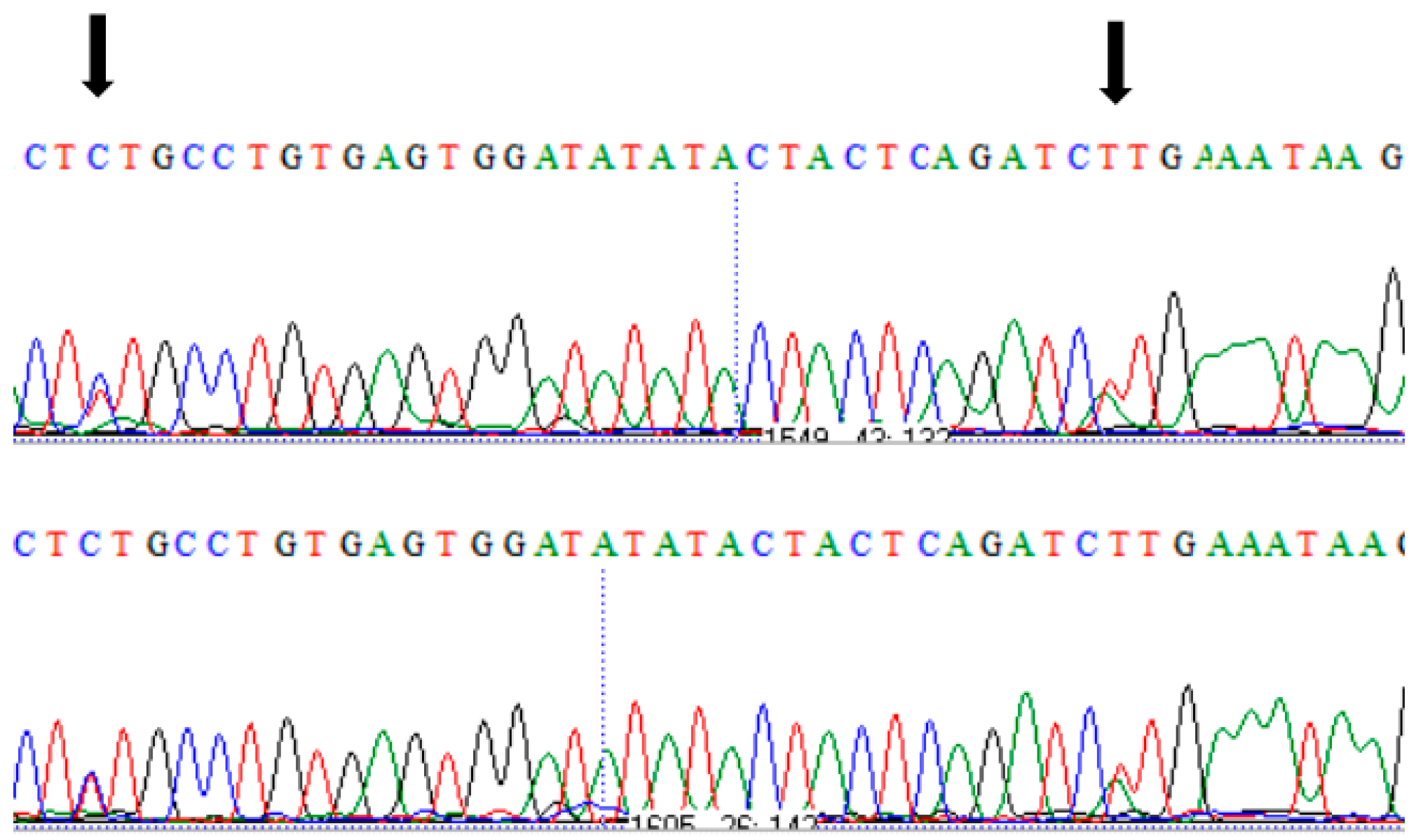
2.2. Sequence Analysis, Variant Verification, and Protein Modelling
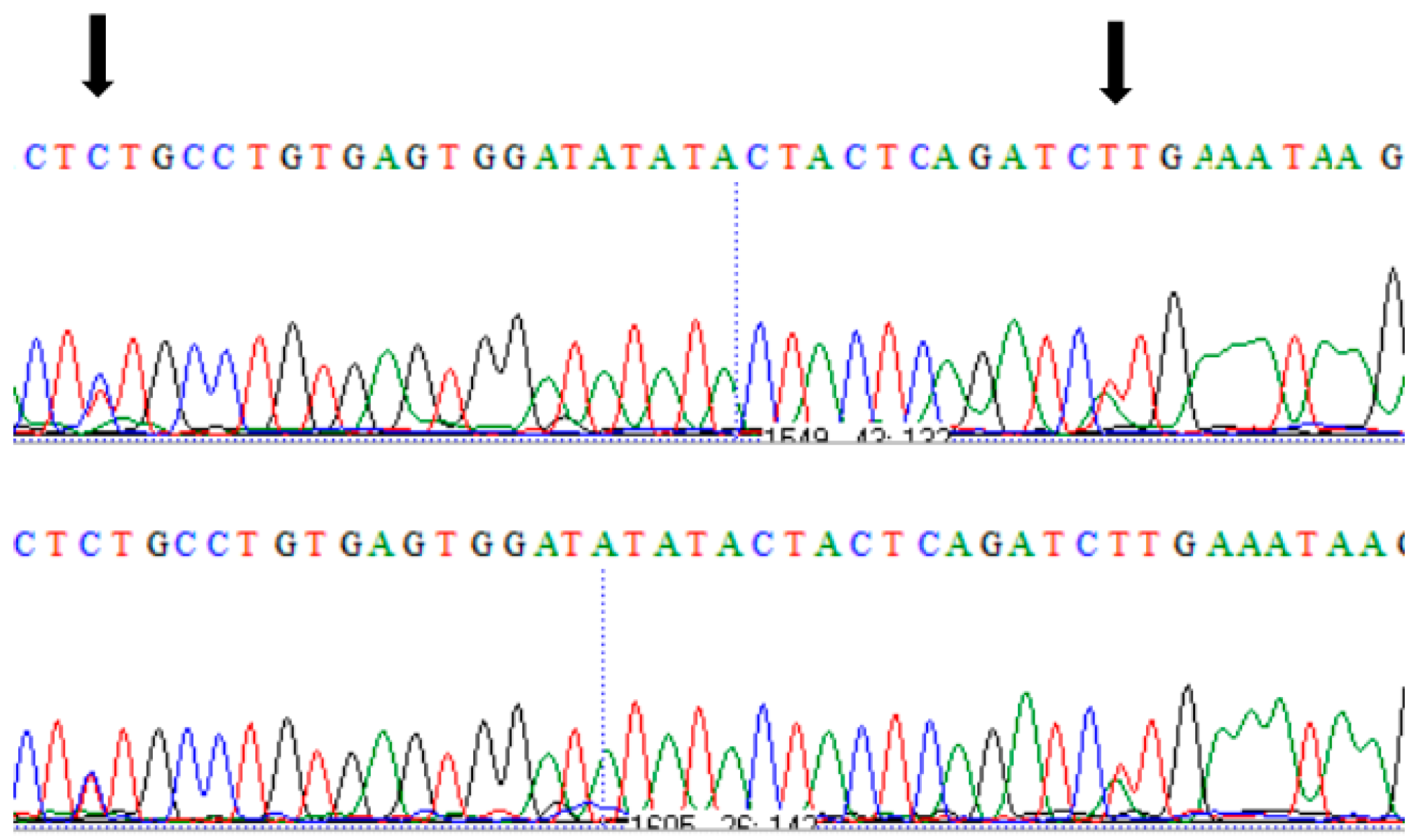
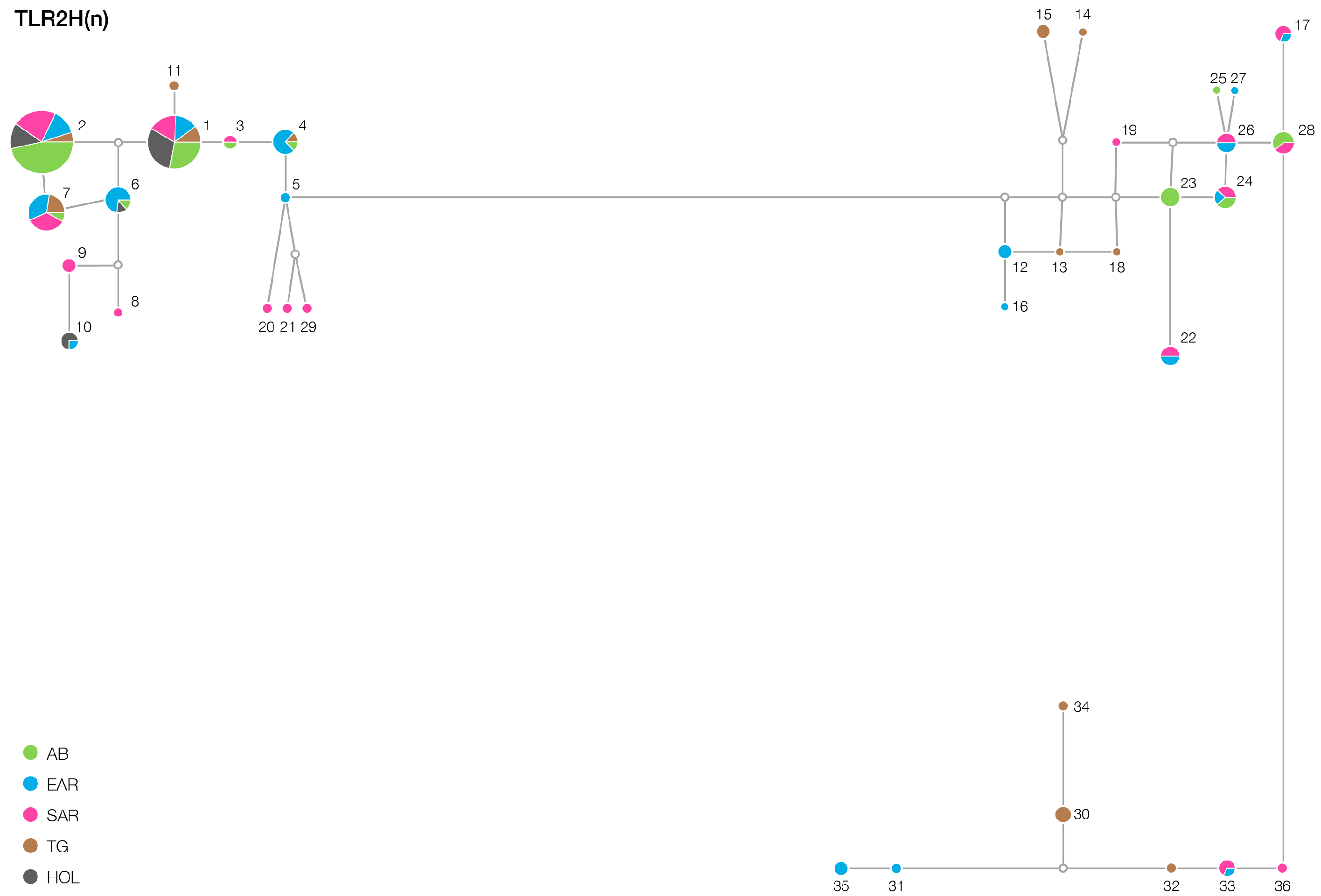
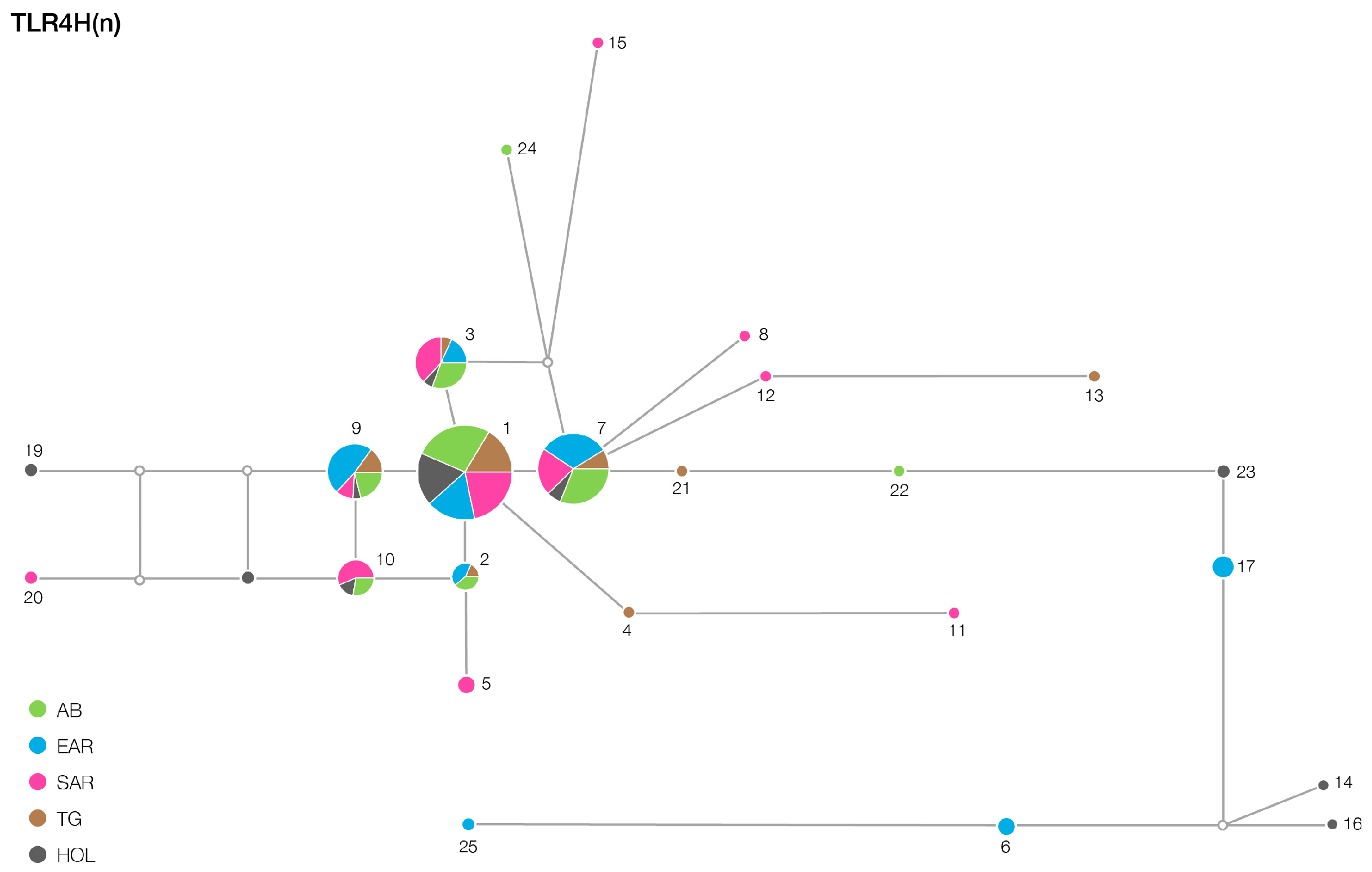
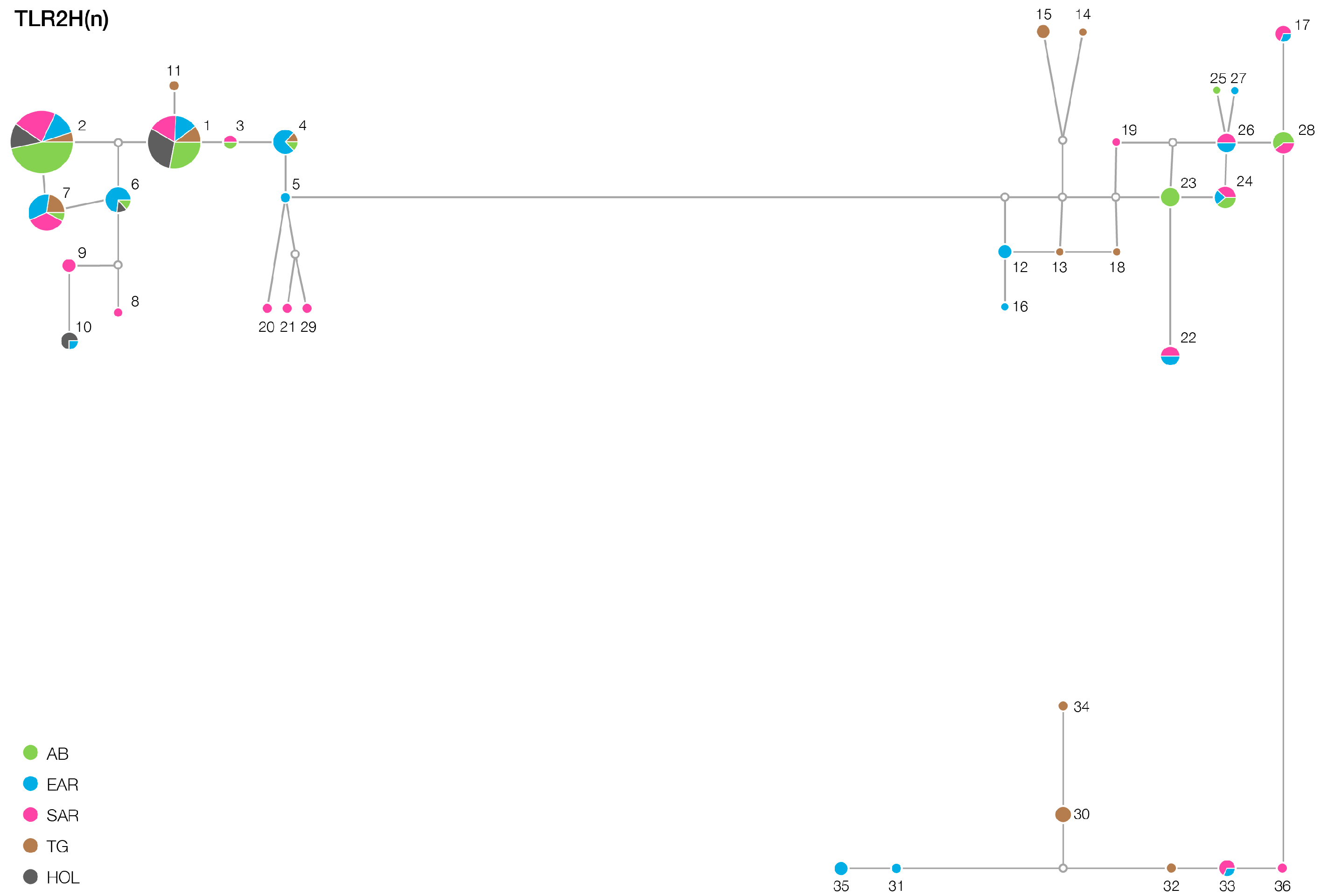
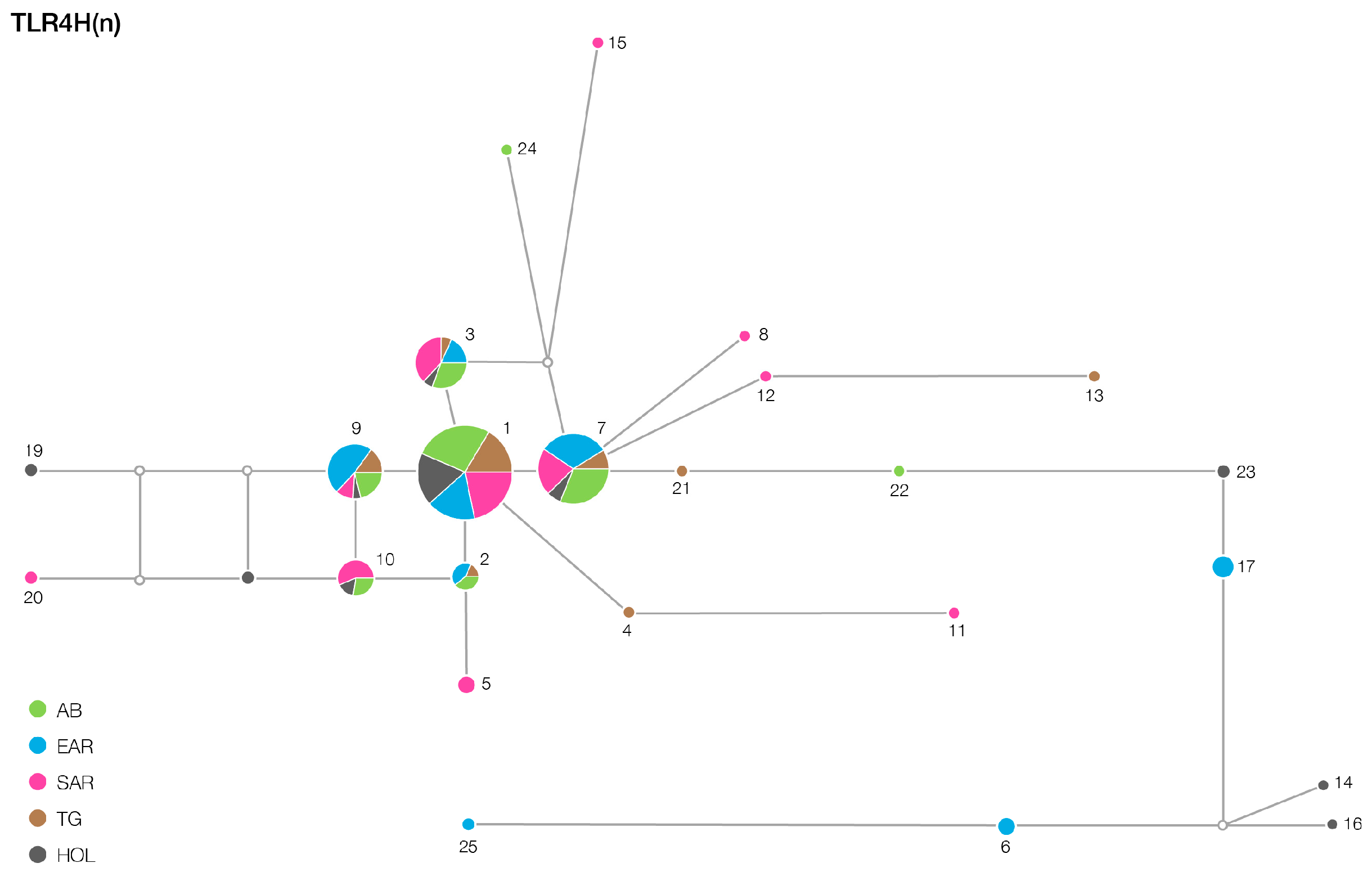
3. Results
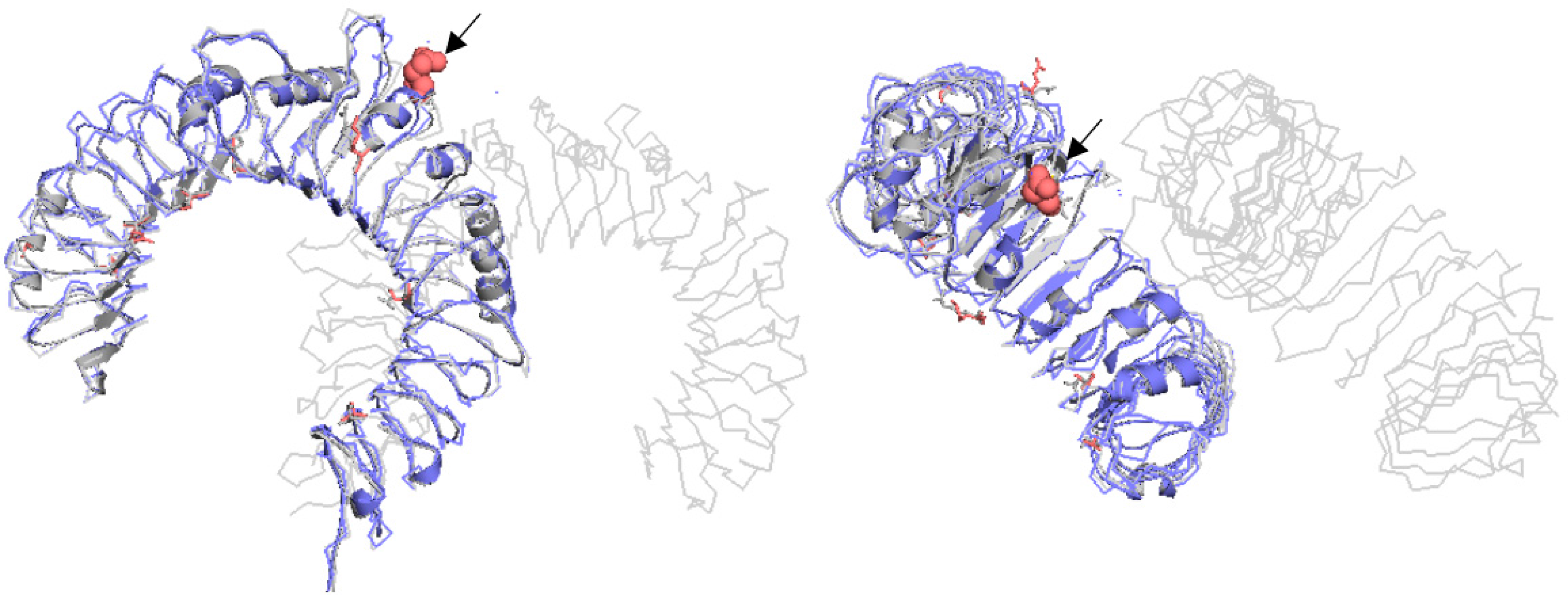
Impact of Identified SNPs on Protein Model
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kumar, H.; Kawai, T.; Akira, S. Pathogen recognition by the innate immune system. Int. Rev. Immunol. 2011, 30, 16–34. [Google Scholar] [CrossRef] [PubMed]
- Medzhitov, R.; Janeway, C.A. Innate immunity: The virtues of a nonclonal system of recognition. Cell 1997, 91, 295–298. [Google Scholar] [CrossRef]
- Tizard, I.R. Veterinary Immunology, 3rd ed.; Saunders Elsevier: Saint Louis, MO, USA, 2009; pp. 13–28. [Google Scholar]
- Takeda, K.; Akira, S. TLR signaling pathways. Semin. Immunol. 2004, 16, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Ogorevc, J.; Kunej, T.; Razpet, A.; Dovc, P. Database of cattle candidate genes and genetic markers for milk production and mastitis. Anim. Genet. 2009, 40, 832–851. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.H.; Uthe, J.J.; Bearson, S.M.; Demirkale, C.Y.; Nettleton, D.; Knetter, S.; Tuggle, C.K. Distinct peripheral blood RNA responses to Salmonella in pigs differing in Salmonella shedding levels: İntersection of IFNG, TLR and miRNA pathways. PLoS ONE. 2011, 6, e28768. [Google Scholar] [CrossRef] [PubMed]
- McGuire, K.; Jones, M.; Werling, D.; Williams, J.L.; Glass, E.J.; Jann, O. Radiation hybrid mapping of all 10 characterized bovine Toll-like receptors. Anim. Genet. 2006, 37, 47–50. [Google Scholar] [CrossRef] [PubMed]
- Kent, W.J.; Sugnet, C.W.; Furey, T.S.; Roskin, K.M.; Pringle, T.H.; Zahler, A.M.; Haussler, D. The human genome browser at UCSC. Genome Res. 2002, 12, 996–1006. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, F.; Amode, M.R.; Barrell, D.; Beal, K.; Billis, K.; Brent, S.; Gil, L. Ensembl 2015. Nucleic Acids Res. 2015, 43, D662–D669. [Google Scholar] [CrossRef] [PubMed]
- Jann, O.C.; Werling, D.; Chang, J.S.; Haig, D.; Glass, E.J. Molecular evolution of bovine Toll-like receptor 2 suggests substitutions of functional relevance. BMC Evol. Biol. 2008, 8, 288. [Google Scholar] [CrossRef] [PubMed]
- Werling, D.; Jann, O.C.; Offord, V.; Glass, E.J.; Coffey, T.J. Variation matters: TLR structure and species-specific pathogen recognition. Trends Immunol. 2009, 30, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Schröder, N.W.; Schumann, R.R. Single nucleotide polymorphisms of Toll-like receptors and susceptibility to infectious disease. Lancet Infect. Dis. 2005, 5, 156–164. [Google Scholar] [CrossRef]
- Alpan, O.; Arpacik, R. Sigir Yetiştiriciliği, 2nd ed.; Sahin Matbaası: Ankara, Turkey, 1996; pp. 39–46. [Google Scholar]
- Yilmaz, O.; Akin, O.; Yener, S.M.; Ertugrul, M.; Wilson, R.T. The domestic livestock resources of Turkey: Cattle local breeds and types and their conservation status. Anim. Genet. Resour. 2012, 50, 65–73. [Google Scholar] [CrossRef]
- Özbeyaz, C. Sigir Yetiştiriciliği Ders Notlari; Ankara University, Faculty of Veterinary Medicine: Ankara, Turkey, 2015; pp. 10–26. [Google Scholar]
- Rozen, S.; Skaletsky, H. Primer3 on the WWW for general users and for biologist programmers. Methods Mol. Biol. 1999, 132, 365–386. [Google Scholar]
- Institue B. Picard. Available online: http://broadinstitute.github.io/picard (accessed on 11 May 2015).
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Durbin, R. The sequence alignment/map format and SAM tools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; DePristo, M.A. The Genome Analysis Toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef] [PubMed]
- Danecek, P.; Auton, A.; Abecasis, G.; Albers, C.A.; Banks, E.; DePristo, M.A.; McVean, G. The variant call format and VCFtools. Bioinformatics 2011, 27, 2156–2158. [Google Scholar] [CrossRef] [PubMed]
- Cingolani, P.; Patel, V.M.; Coon, M.; Nguyen, T.; Land, S.J.; Ruden, D.M.; Lu, X. Using Drosophila Melanogaster as a Model for Genotoxic Chemical Mutational Studies with a New Program, SnpSift. In Toxicogenomics in Non-Mammalian Species; Frontiers E-books: Lausanne, Switzerland, 2012; p. 35. [Google Scholar]
- Cingolani, P.; Platts, A.; Wang, L.L.; Coon, M.; Nguyen, T.; Wang, L.; Ruden, D.M. A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff: SNPs in the genome of Drosophila melanogaster strain w1118; iso-2; iso-3. Fly 2012, 6, 80–92. [Google Scholar] [CrossRef] [PubMed]
- McLaren, W.; Pritchard, B.; Rios, D.; Chen, Y.; Flicek, P.; Cunningham, F. Deriving the consequences of genomic variants with the Ensembl API and SNP Effect Predictor. Bioinformatics 2010, 26, 2069–2070. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Webb, B.; Sali, A. Comparative Protein Structure Modeling Using Modeller. Curr. Protoc. Bioinform. 2014, 47, 1–32. [Google Scholar]
- Laskowski, R.A.; MacArthur, M.W.; Moss, D.S.; Thornton, J.M. PROCHECK: A program to check the stereochemical quality of protein structures. J. Appl. Crystallogr. 1993, 26, 283–291. [Google Scholar] [CrossRef]
- Eisenberg, D.; Lüthy, R.; Bowie, J.U. VERIFY3D: Assessment of protein models with three-dimensional profiles. Methods Enzymol. 1997, 277, 396–404. [Google Scholar] [PubMed]
- Colovos, C.; Yeates, T.O. ERRAT: An empirical atom-based method for validating protein structures. Protein Sci. 1993, 2, 1511–1519. [Google Scholar] [CrossRef] [PubMed]
- Wallner, B.; Elofsson, A. Can correct protein models be identified? Protein Sci. 2003, 12, 1073–1086. [Google Scholar] [CrossRef] [PubMed]
- Thorvaldsdottir, H.; Robinson, J.T.; Mesirov, J.P. Integrative Genomics Viewer (IGV): High-performance genomics data visualization and exploration. Brief. Bioinform. 2013, 14, 178–192. [Google Scholar] [CrossRef] [PubMed]
- Delaneau, O.; Howie, B.; Cox, A.; Zagury, J.F.; Marchini, J. Haplotype estimation using sequence reads. Am. J. Hum. Biol. 2013, 93, 787–696. [Google Scholar]
- Bandelt, H.; Forster, P.; Rohl, A. Median joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef] [PubMed]
- DeLano, W.L. PyMOL; DeLano Scientific: San Carlos, CA, USA, 2002; p. 700. [Google Scholar]
- Mariotti, M.; Williams, J.L.; Dunner, S.; Valentini, A.; Pariset, L. Polymorphisms within the toll-like receptor (TLR)-2, -4, and-6 genes in cattle. Diversity 2009, 1, 7–18. [Google Scholar] [CrossRef]
- Garza-Gonzalez, E.; Bosques-Padilla, F.J.; Mendoza-Ibarra, S.I.; Flores-Gutierrez, J.P.; Maldonado-Garza, H.J.; Perez-Perez, G.I. Assessment of the toll-like receptor 4 Asp299Gly, Thr399Ile and interleukin-8-251 polymorphisms in the risk for the development of distal gastric cancer. BMC Cancer 2007, 7, 70. [Google Scholar] [CrossRef] [PubMed]
- Mucha, R.; Bhide, M.R.; Chakurkar, E.B.; Novak, M.; Mikula, I. Toll-like receptors TLR1, TLR2 and TLR4 gene mutations and natural resistance to Mycobacterium avium subsp paratuberculosis infection in cattle. Vet. Immunol. Immunopathol. 2009, 128, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Larrañaga, O.; Manzano, C.; Iriondo, M.; Garrido, J.M.; Molina, E.; Vazquez, P.; Estonba, A. Genetic variation of toll-like receptor genes and infection by Mycobacterium avium ssp. paratuberculosis in Holstein-Friesian cattle. J. Dairy Sci. 2011, 94, 3635–3641. [Google Scholar] [CrossRef] [PubMed]
- Jungi, T.W.; Farhat, K.; Burgener, I.A.; Werling, D. Toll-like receptors in domestic animals. Cell Tissue Res. 2011, 343, 107–120. [Google Scholar] [CrossRef] [PubMed]
- Osvaldova, A.; Woodman, S.; Patterson, N.; Offord, V.; Mwangi, D.; Gibson, A.J.; Matiasovic, J.; Werling, D. Replacement of two aminoacids in the bovine Toll-like receptor 5 TIR domain with their human counterparts partially restores functional response to flagellin. Dev. Comp. Immunol. 2014, 47, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, P. The Immune System. In Breeding for Disease Resistance in Farm Animals, 3rd ed.; Bishop, S.C., Axford, R.F.E., Nicholas, F.W., Owen, J.B., Eds.; Cabi Publishing: Cambridge, MA, USA, 2010; pp. 15–38. [Google Scholar]
- Ropert, C.; Gazzinelli, R.T. Regulatory role of Toll-like receptor 2 during infection with Trypanosoma cruzi. J. Endotoxin Res. 2004, 10, 425–430. [Google Scholar] [CrossRef] [PubMed]
- Deniz, A.; Oncel, T.; Icen, H.; Simsek, A. Detection of Theileria annulata and T. buffeli in Cattle by Multiplex PCR in Diyarbakir Area. Kafkas Univ. Vet. Fak. Derg. 2012, 18, A111–A114. [Google Scholar] [CrossRef]
- Duzlu, O.; Yıldırım, A.; Inci, A.; Avcioglu, H.; Balkaya, I. Investigation of Babesia bovis and Babesia bigemina in cattle by Real Time PCR and molecular characterization of the isolates. Ankara Univ. Vet. Fak. Derg. 2015, 62, 27–35. [Google Scholar]
- Loftus, R.T.; Ertuğrul, O.; Harba, A.H.; El-Barody, M.A.; Machugh, D.E.; Park, S.D.; Bradley, D.G. A microsatellite survey of cattle from a center of origin: The Near East. Mol. Ecol. 1999, 8, 2015–2022. [Google Scholar] [CrossRef] [PubMed]
- Decker, J.E.; McKay, S.D.; Rolf, M.M.; Kim, J.; Alcalá, A.M.; Sonstegard, T.S.; Hanotte, O.; Götherström, A.; Seabury, C.M.; Praharani, L.; et al. Worldwide patterns of ancestry, divergence, and admixture in domesticated cattle. PLoS Genet. 2014, 10, e1004254. [Google Scholar] [CrossRef] [PubMed]
- Seabury, C.M.; Seabury, P.M.; Decker, J.E.; Schnabel, R.D.; Taylor, J.F.; Womack, J.E. Diversity and evolution of 11 innate immune genes in Bos taurus taurus and Bos taurus indicus cattle. Proc. Natl. Acad. Sci. USA 2010, 107, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Fisher, C.A.; Bhattarai, E.K.; Osterstock, J.B.; Dowd, S.E.; Seabury, P.M.; Vikram, M.; Womack, J.E. Evolution of the bovine TLR gene family and member associations with Mycobacterium avium subspecies paratuberculosis infection. PLoS ONE 2011, 6, e27744. [Google Scholar] [CrossRef] [PubMed]
- Ingvartsen, K.L.; Dewhurst, R.J.; Friggens, N.C. On the relationship between lactational performance and health: Is it yield or metabolic imbalance that cause production diseases in dairy cattle? A position paper. Livest. Prod. Sci. 2003, 83, 277–308. [Google Scholar] [CrossRef]
- Oltenacu, P.A.; Broom, D.M. The impact of genetic selection for increased milk yield on the welfare of dairy cows. Anim. Welf. 2010, 19, 39–49. [Google Scholar]
- Haldane, J.B.S. Disease and evolution. Curr. Sci. 1992, 63, 599–604. [Google Scholar]
- Bermingham, M.L.; Bishop, S.C.; Woolliams, J.A.; Pong-Wong, R.; Allen, A.R.; McBride, S.H.; Glass, E.J. Genome-wide association study identifies novel loci associated with resistance to bovine tuberculosis. Heredity 2014, 112, 543–551. [Google Scholar] [CrossRef] [PubMed]
- De Mesquita, A.Q.; Rezende, C.S.M.; de Mesquita, A.J.; Jardim, E.A.G.; Kipnis, A.P.J. Association of TLR4 polymorphisms with subclinical mastitis in Brazilian holsteins. Braz. J. Microbiol. 2012, 43, 692–697. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Breed | ||||
|---|---|---|---|---|---|
| AB | EAR | SAR | TG | HOL | |
| TLR2 | 133/11 | 153/17 | 155/13 | 187/16 | 45/3 |
| Exon | 23 | 27 | 28 | 26 | 6 |
| missense | 13 | 16 | 17/1 | 15/1 | 4 |
| synonymous | 10 | 11/1 | 11/1 | 11 | 2 |
| Intron | 93/10 | 111/15 | 110/10 | 146/15 | 34/3 |
| 3′UTR | 15/1 | 13/1 | 15/1 | 14 | 4 |
| 5′UTR | 2 | 2 | 2 | 1 | 1 |
| TLR6 | 209/14 | 230/5 | 285/52 | 179/5 | 155 |
| Exon | 26 | 23 | 26 | 24 | 9 |
| missense | 10/2 | 9/1 | 9/1 | 8/1 | 3 |
| synonymous | 16/1 | 14 | 17/2 | 16/1 | 6 |
| Intron | 177/8 | 203/4 | 252/46 | 153/2 | 103 |
| 3′UTR | 2 | 1 | 3/2 | 2 | 1 |
| 5′UTR | 4/3 | 4 | 5/1 | 1/1 | 2 |
| TLR4 | 18/1 | 80/1 | 80/3 | 57/1 | 74/1 |
| Exon | 2 | 16 | 13 | 10 | 15 |
| missense | 1 | 6 | 2 | 3/1 | 5 |
| synonymous | 1 | 10 | 11 | 7 | 10 |
| Intron | 14 | 59/1 | 62/3 | 45 | 55/1 |
| 3′UTR | 1 | 2 | 2 | 1 | 1 |
| 5′UTR | 1 | 3 | 5 | 1 | 5 |
| Gene | Breed | ||||
|---|---|---|---|---|---|
| AB | EAR | SAR | TG | HOL | |
| TLR2 | 12/12 | 16/13 | 15/12 | 16/13 | 6/6 |
| TLR6 | 23/9 | 28/14 | 38/24 | 21/13 | 17/3 |
| TLR4 | 3/1 | 5/3 | 8/3 | 6/4 | 5/1 |
| Domain | TLR2 (aa) | AB | EAR | SAR | TG | HOL | SNP ID |
|---|---|---|---|---|---|---|---|
| LRR1 | 54..77 | 62 N/N | 62 N/N | 62 N/N | 62 N/N | rs68268249 | |
| 63 E/D | 63 E/D | 63 E/D | 63 E/D | 63 E/D | rs55617172 | ||
| 68 G/S | 68 G/S | 68 G/S | 68 G/S | rs68268250 | |||
| LRR3 | 102..125 | 119 W/L | 119 W/L | 119 W/L | rs211243949 | ||
| LRR5 | 151..175 | 152 R/Q | 152 R/Q | 152 R/Q | rs43706434 | ||
| LRR7 | 200..223 | 201 S/N | 201 S/N | rs110491977 | |||
| 211 I/V | 211 I/V | 211 I/V | 211 I/V | 211 I/V | rs43706433 | ||
| LRR8 | 224..250 | 227 F/L | 227 F/L | 227 F/L | 227 F/L | rs68268251 | |
| LRR9-10 | 251..308 | ||||||
| LRR11 | 309..337 | 315 R/R | rs68268253 | ||||
| 326 H/Q | 326 H/Q | 326 H/Q | 326 H/Q | rs68343167 | |||
| LRR12 | 338..361 | 337 R/Q | 337 R/Q | 337 R/Q | 337 R/Q | rs68343168 | |
| LRR13 | 362..388 | ||||||
| LRR14 | 389..414 | 405 T/M | 405 T/M | 405 T/M | 405 T/M | rs68268255 | |
| LRR15 | 415..437 | 417 N/S | 417 N/S | 417 N/S | 417 N/S | rs68268256 | |
| 436 G/G | 436 G/G | 436 G/G | 436 G/G | rs68268257 | |||
| LRR16-18 | 438..500 | ||||||
| LRR19 | 501..524 | 502 S/A | 502 S/A | 502 S/A | 502 S/A | rs68268258 | |
| 530 A/A | novel | ||||||
| 531 G/S | novel | ||||||
| LRR20 | 533..586 | 544 F/F | 545 F/F | 546 F/F | 547 F/F | 544 F/F | rs68268259 |
| 563 R/H | 563 R/H | 563 R/H | 563 R/H | rs68268260 | |||
| 569 H/H | 569 H/H | 569 H/H | 569 H/H | 569 H/H | rs41830058 | ||
| Trans Membrane | 588..608 | 574 R/W | novel | ||||
| 593 A/A | 593 A/A | 593 A/A | 593 A/A | rs68268261 | |||
| 594 A/A | 594 A/A | 594 A/A | 594 A/A | rs68343169 | |||
| 605 T/M | 605 T/M | 605 T/M | 605 T/M | rs68343170 | |||
| 634 A/V | novel | ||||||
| TIR | 640..784 | 644 F/F | novel | ||||
| 655 V/A | |||||||
| 665 H/Q | 665 H/Q | 665 H/Q | 665 H/Q | rs68268263 | |||
| 675 H/H | 675 H/H | 675 H/H | 675 H/H | rs68343171 | |||
| 685 I/I | 685 I/I | 685 I/I | 685 I/I | rs68268264 | |||
| 738 E/Q | 738 E/Q | 738 E/Q | rs207552166 | ||||
| 738 E/E | 738 E/E | 738 E/E | 738 E/E | rs68268266 | |||
| ATG16Lmotif | 761..778 | 765 P/P | 765 P/P | 765 P/P | 765 P/P | rs68268267 |
| Domain | TLR4 (aa) | AB | EAR | SAR | TG | HOL | SNP ID |
|---|---|---|---|---|---|---|---|
| LRR1 | 55..76 | ||||||
| LRR2 | 79..100 | ||||||
| LRR3 | 103..124 | ||||||
| LRR4 | 127..148 | ||||||
| LRR5 | 151..172 | 151 N/T | 151 N/T | rs8193049 | |||
| LRR6 | 176..197 | ||||||
| LRR7 | 205..225 | ||||||
| 238 N/K | rs8193050 | ||||||
| 276 F/F | rs8193051 | ||||||
| 347 A/E | 347 A/E | rs8193053 | |||||
| LRR8 | 352..373 | ||||||
| LRR9 | 374..394 | 374 P/P | 374 P/P | 374 P/P | 374 P/P | rs8193054 | |
| 381 K/R | rs8193055 | ||||||
| 385 L/L | rs8193056 | ||||||
| 389 G/G | 389 G/G | rs8193057 | |||||
| LRR10 | 400..422 | ||||||
| LRR11 | 423..444 | ||||||
| LRR12 | 448..469 | ||||||
| LRR13 | 472..495 | 482 S/Y | novel | ||||
| LRR14 | 497..518 | 507 Q/Q | 507 Q/Q | 507 Q/Q | 507 Q/Q | rs8193059 | |
| LRR15 | 521..542 | ||||||
| LRR16 | 545..568 | 552 S/S | 552 S/S | 552 S/S | 552 S/S | 552 S/S | rs8193060 |
| LRR17 | - | ||||||
| LRR18 | - | ||||||
| LRR19 | - | ||||||
| LRRCT | 579..626 | 589 S/S | rs8193061 | ||||
| 609 C/C | 609 C/C | rs8193062 | |||||
| 622 S/S | rs8193063 | ||||||
| 625 N/N | 625 N/N | rs8193064 | |||||
| 649 G/G | 649 G/G | 649 G/G | rs8193065 | ||||
| 640 V/I | 640 V/I | rs8193066 | |||||
| Trans membrane | 633..653 | ||||||
| 664 G/G | 664 G/G | rs8193067 | |||||
| 674 T/I | 674 T/I | 674 T/I | 674 T/I | 674 T/I | rs8193069 | ||
| 676 D/D | rs8193070 | ||||||
| TIR | 677..815 |
| Domain | TLR6 (aa) | AB | EAR | SAR | TG | HOL | SNP ID |
|---|---|---|---|---|---|---|---|
| LRR_RI | <43..164 | 37 D/N | 37 D/N | 37 D/N | |||
| LRR1 | 54..77 | 61 Q/Q | 61 Q/Q | 61 Q/Q | 61 Q/Q | rs68268271 | |
| LRR2 | 78..101 | 87 R/G | 87 R/G | 87 R/G | 87 R/G | rs68268272 | |
| LRR3 | 102..122 | 116 S/P | 116 S/P | ||||
| LRR4 | 123..147 | 135 D/H | 135 D/H | 135 D/H | 135 D/H | rs520121582 | |
| LRR5-LRR6 | 148..196 | ||||||
| LRR7 | 197..219 | 214 D/N | 214 D/N | 214 D/N | 214 D/N | 214 D/N | rs43702941 |
| 217 A/A | 217 A/A | 217 A/A | 217 A/A | rs68268273 | |||
| LRR8-LRR12 | 220.354 | ||||||
| LRR13 | 355..378 | 374 D/D | 374 D/D | 374 D/D | rs68268274 | ||
| LRR14 | 379..404 | 395 T/A | 395 T/A | 395 T/A | 395 T/A | rs68268275 | |
| 400 K/K | 400 K/K | 400 K/K | 400 K/K | rs211657505 | |||
| LRR15 | 405..428 | 425 S/S | 425 S/S | 425 S/S | 425 S/S | rs68268276 | |
| LRR16 | 429..449 | ||||||
| LRR17 | 450..473 | 458 H/H | 458 H/H | rs68268277 | |||
| LRR18 | 474..495 | ||||||
| LRR19 | 496..519 | 505 N/N | 505 N/N | 505 N/N | 505 N/N | rs55617146 | |
| 526 V/A | 526 V/A | 526 V/A | 526 V/A | 526 V/A | rs68343174, rs133754378 | ||
| 526 V | 526 V | 526 V | 526 V | 526 V | rs68343175, rs136574510 | ||
| LRRCT | 529..582 | 539 D/D | 539 D/D | 539 D/D | rs68343176 | ||
| 544 V/I | 544 V/I | rs55617465, rs68268279 | |||||
| 573 K/K | 573 K/K | 573 K/K | 573 K/K | rs55617193 | |||
| Trans membrane | 585..605 | 589 V/I | 589 V/I | 589 V/I | 589 V/I | 589 V/I | rs55617317, rs207882984 |
| 605 L/L | 605 L/L | 605 L/L | 605 L/L | 605 L/L | rs68268280, rs378853146 | ||
| TIR | 641..784 | 642 F/F | 642 F/F | 642 F/F | 642 F/F | rs438448894 | |
| 669 I/V | 669 I/V | 669 I/V | 669 I/V | rs210580164 | |||
| 674 H/H | 674 H/H | 674 H/H | 674 H/H | 674 H/H | rs209572763 | ||
| 676 R/R | 676 R/R | 676 R/R | 676 R/R | rs68343178 | |||
| 680 A/A | 680 A/A | 680 A/A | novel | ||||
| 684 I/I | 684 I/I | 684 I/I | 684 I/I | rs68343179 | |||
| 700 F/F | 700 F/F | 700 F/F | 700 F/F | rs55617339, rs211454671 | |||
| 701 V/V | 701 V/V | 701 V/V | 701 V/V | 701 V/V | rs207586910 | ||
| 709 S/S | 709 S/S | rs55617335 | |||||
| E/E 710 | rs68268282 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bilgen, N.; Cinar Kul, B.; Offord, V.; Werling, D.; Ertugrul, O. Determination of Genetic Variations of Toll-Like Receptor (TLR) 2, 4, and 6 with Next-Generation Sequencing in Native Cattle Breeds of Anatolia and Holstein Friesian. Diversity 2016, 8, 23. https://doi.org/10.3390/d8040023
Bilgen N, Cinar Kul B, Offord V, Werling D, Ertugrul O. Determination of Genetic Variations of Toll-Like Receptor (TLR) 2, 4, and 6 with Next-Generation Sequencing in Native Cattle Breeds of Anatolia and Holstein Friesian. Diversity. 2016; 8(4):23. https://doi.org/10.3390/d8040023
Chicago/Turabian StyleBilgen, Nuket, Bengi Cinar Kul, Victoria Offord, Dirk Werling, and Okan Ertugrul. 2016. "Determination of Genetic Variations of Toll-Like Receptor (TLR) 2, 4, and 6 with Next-Generation Sequencing in Native Cattle Breeds of Anatolia and Holstein Friesian" Diversity 8, no. 4: 23. https://doi.org/10.3390/d8040023
APA StyleBilgen, N., Cinar Kul, B., Offord, V., Werling, D., & Ertugrul, O. (2016). Determination of Genetic Variations of Toll-Like Receptor (TLR) 2, 4, and 6 with Next-Generation Sequencing in Native Cattle Breeds of Anatolia and Holstein Friesian. Diversity, 8(4), 23. https://doi.org/10.3390/d8040023