Norway Spruce (Picea abies L.) Provenances Use Different Physiological Strategies to Cope with Water Deficit

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil Water Content
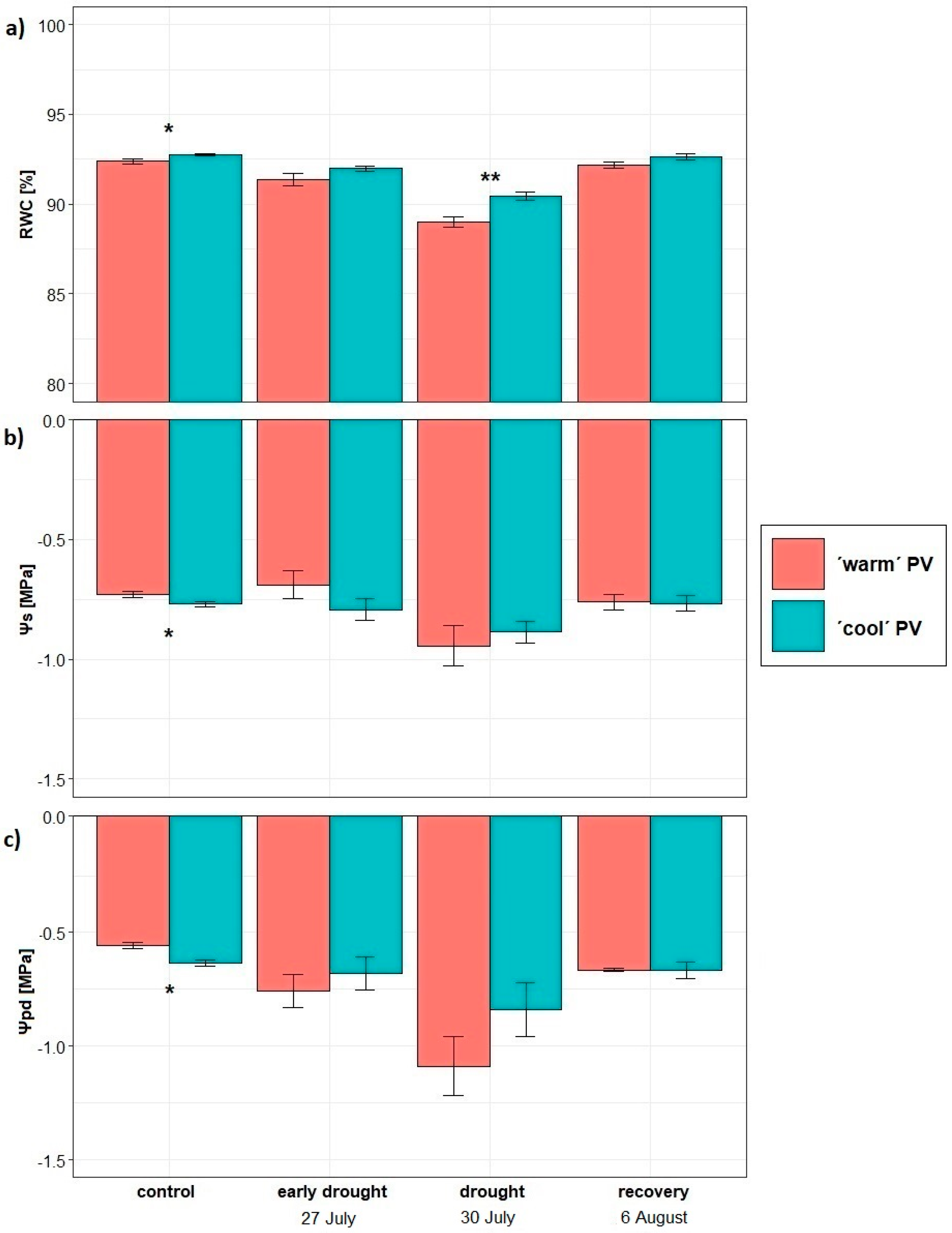
2.2. Predawn Water Potential and Osmotic Potential of Needles
2.3. Relative Water Content of Needles
2.4. Free Proline Content
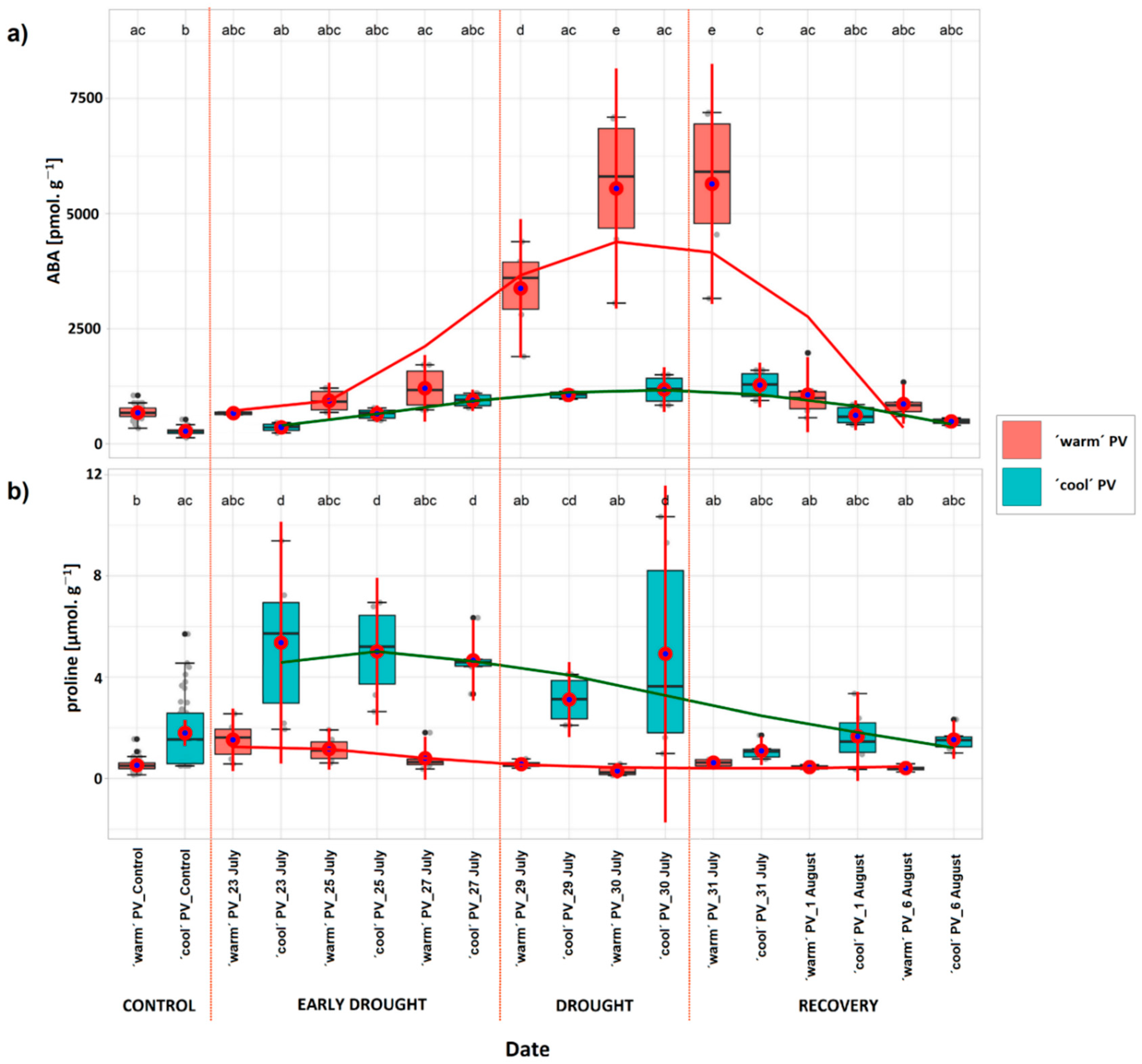
2.5. Concentration of Abscisic Acid
2.6. Gas Exchange
2.7. Data Analysis
3. Results
3.1. Course of Drought Stress and Recovery
3.2. Water Status
3.3. Gas Exchange Parameters
3.4. Biochemical Parameters
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Aitken, S.N.; Yeaman, S.; Holliday, J.; Wang, T.; Curtis-McLane, S. Adaptation, migration or extirpation: climate change outcomes for tree populations. Evol. Appl. 2008, 1, 95–111. [Google Scholar] [CrossRef] [PubMed]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef] [Green Version]
- Lindner, M.; Maroschek, M.; Netherer, S.; Kremer, A.; Barbati, A.; Garcia-Gonzalo, J.; Seidl, R.; Delzon, S.; Corona, P.; Kolström, M.; et al. Climate change impacts, adaptive capacity, and vulnerability of European forest ecosystems. For. Ecol. Manag. 2010, 259, 698–709. [Google Scholar] [CrossRef]
- Gömöry, D.; Longauer, R.; Hlásny, T.; Pacalaj, M.; Strmeň, S.; Krajmerová, D. Adaptation to common optimum in different populations of Norway spruce (Picea abies Karst.). Eur. J. For. Res. 2012, 131, 401–411. [Google Scholar] [CrossRef]
- Kremer, A.; Ronce, O.; Robledo-Arnuncio, J.J.; Guillaume, F.; Bohrer, G.; Nathan, R.; Bridle, J.R.; Gomulkiewicz, R.; Klein, E.K.; Ritland, K.; et al. Long-Long-distance gene flow and adaptation of forest trees to rapid climate change. Ecol. Lett. 2012, 15, 378–392. [Google Scholar] [CrossRef] [PubMed]
- Bolte, A.; Ammer, C.; Löf, M.; Madsen, P.; Nabuurs, G.J.; Schall, P.; Spathelf, P.; Rock, J. Adaptive forest management in central Europe: Climate change impacts, strategies and integrative concept. Scand. J. For. Res. 2009, 24, 473–482. [Google Scholar] [CrossRef]
- Montwé, D.; Spiecker, H.; Hamann, A. An experimentally controlled extreme drought in a Norway spruce forest reveals fast hydraulic response and subsequent re-covery of growth rates. Trees 2014, 28, 891–900. [Google Scholar] [CrossRef]
- Skrøppa, T. EUFORGEN Technical Guidelines for Genetic Conservation and Use for Norway Spruce (Picea Abies); International Plant Genetic Resources Institute: Rome, Italy, 2003; p. 6. [Google Scholar]
- Jyske, T.; Mäkinen, H.; Kalliokoski, T.; Nöjd, P. Intraannual tracheid production of Norway spruce and Scots pine across a latitudinal gradient in Finland. Agric. For. Meteorol. 2014, 194, 241–254. [Google Scholar] [CrossRef]
- Tužinský, L.; Bublinec, E.; Tužinský, M. Development of soil water regime under spruce stands. Folia Oecol. 2017, 44, 46–53. [Google Scholar] [CrossRef] [Green Version]
- Štroch, M.; Vrábl, D.; Podolinská, J.; Kalina, J.; Urban, O.; Špunda, V. Acclimation of Norway spruce photosynthetic apparatus to the combined effect of high irradiance and temperature. J. Plant Physiol. 2010, 167, 597–605. [Google Scholar] [CrossRef]
- Olsen, J.E.; Lee, Y.K. Trees and Boreal Forests. In Temperature Adaptation in a Changing Climate: Nature at Risk; Storey, K.B., Tanino, K.K., Eds.; CAB International: Wallingford, England, 2011; pp. 160–178. [Google Scholar]
- Pashkovskiy, P.P.; Vankova, R.; Zlobin, I.E.; Dobrev, P.; Ivanov, Y.V.; Kartashov, A.V.; Kuznetsov, V.V. Comparative analysis of abscisic acid levels and expression of abscisic acid-related genes in Scots pine and Norway spruce seedlings under water deficit. Plant Physiol. Biochem. 2019, 140, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Riechmann, J.L.; Ratcliffe, O.J. A genomic perspective on plant transcription factors. Curr. Opin. Plant Biol. 2000, 3, 423–434. [Google Scholar] [CrossRef]
- Centritto, M.; Togneti, R.; Leitgeb, E.; Střelcová, K.; Cohen, S. Above ground processes: Anticipating climate change influences. In Forest Management and the Water Cycle; Bredemeier, M., Cohen, S., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 31–64. [Google Scholar]
- Renninger, H.J.; Carlo, N.J.; Clark, K.L.; Schäfer, K.V.R. Resource use and efficiency, and stomatal responses to environmental drivers of oak and pine species in an Atlantic Coastal Plain forest. Front. Plant Sci. 2015, 6, 297. [Google Scholar] [CrossRef] [PubMed]
- Heuer, B. Role of Proline in Plant Response to Drought and Salinity. In Handbook of Plant and Crop Stress, 3rd ed.; Pessarakli, A., Ed.; CRC Press: Boca Raton, Florida, 2010; pp. 213–238. [Google Scholar]
- Ben Hassine, A.; Ghanem, M.E.; Bouzid, S.; Lutts, S. An inland and a coastal population of the Mediterranean xero-halophyte species Atriplex halimus L. differ in their ability to accumulate prolineand glycinebetaine in response to salinity and water stress. J. Exp. Bot. 2008, 59, 1315–1326. [Google Scholar] [CrossRef] [PubMed]
- Parida, A.K.; Dagaonkar, V.S.; Phalak, M.S.; Aurangabadkar, L.P. Differential responses of the enzymes involved in proline biosynthesis and degradation in drought tolerant and sensitive cotton genotypes during drought stress and recovery. Acta Physiol. Plant. 2008, 30, 619–627. [Google Scholar] [CrossRef]
- Evers, D.; Lefevre, I.; Legay, S.; Lamoureux, D.; Hausman, J.-F.; Gutierrez Rosales, R.O.; Tincopa Marca, L.R.; Hoffmann, L.; Bonierbale, M.; Schafleitner, R. Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metabolite approach. J. Exp. Bot. 2010, 61, 2327–2343. [Google Scholar] [CrossRef] [Green Version]
- Pšidová, E.; Živčák, M.; Stojnić, S.; Orlović, S.; Gömöry, D.; Kučerová, J.; Ditmarová, Ľ.; Střelcová, K.; Brestič, M.; Kalaji, H.M. Altitude of origin influences the responses of PSII photochemistry to heat waves in European beech (Fagus sylvatica L.). Environ. Exp. Bot. 2018, 152, 97–106. [Google Scholar]
- Czajkowski, T.; Bolte, A. Unteschiedliche Reaktionen deutscher und polnischer Herkunfte der Buche (Fagus sylvatica L.) auf Trockenheit. Allg. Forst-u. J.-Ztg. 2006, 177, 30–40. [Google Scholar]
- Rose, L.; Leuschner, C.; Köckemann, B.; Buschmann, H. Are marginal beech (Fagus sylvatica L.) provenances a source for drought tolerant ecotypes? Eur. J. For. Res. 2009, 128, 335–343. [Google Scholar] [CrossRef]
- Padilla, F.M.; Miranda, J.D.; Pugnaire, F.I. Early root growth plasticity in seedlings of three Mediterranean woody species. Plant Soil. 2007, 296, 103–113. [Google Scholar] [CrossRef]
- Niinemets, ü. Responses of forest trees to single and multiple environmental stresses from seedlings to mature plants: Past stress history, stress interactions, tolerance and acclimation. Forest Ecol. Manag. 2010, 260, 1623–1639. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- González, L.; González-Vilar, M. Determination of Relative Water Content. In Handbook of Plant Ecophysiology Techniques, 1st ed.; Reigosa, R.M.J., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2003; pp. 207–212. [Google Scholar]
- Bates, L.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Dobrev, P.I.; Havlíček, L.; Vágner, M.; Malbeck, J.; Kamínek, M. Purification and determination of plant hormones auxin and abscisic acid using solid phase extraction and two-dimensional high performance liquid chromatography. J. Chromatogr. A. 2005, 1075, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014. [Google Scholar]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R Package for Multivariate Analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef]
- Holmström, E.; Gålnander, H.; Petersson, M. Within-site variation in seedling survival in Norway spruce plantations. Forests 2019, 10, 181. [Google Scholar] [CrossRef]
- Albert, M.; Nagel, R.-V.; Nuske, R.S.; Sutmöller, J.; Spellmann, H. Tree Species Selection in the Face of Drought Risk—Uncertainty in Forest Planning. Forests 2017, 8, 363. [Google Scholar] [CrossRef]
- Slugeňová, K.; Ditmarová, Ľ.; Kurjak, D.; Váľka, J. Drought and aluminium as stress factors in Norway spruce (Picea abies [L.] Karst) seedlings. J. For. Sci. 2011, 57, 547–554. [Google Scholar] [CrossRef]
- Haas, J.C.; Vergara, A.; Hurry, V.; Street, N.R. Candidate Regulators and Target Genes of Drought Stress in Needles and Roots of Norway Spruce. Available online: https://doi.org/10.1101/517151https://t.co/uq9BXGZXXV (accessed on 1 April 2019).
- Ditmarová, Ľ.; Kurjak, D.; Palmroth, S.; Kmeť, J.; Střelcová, K. Physiological responses of Norway spruce (Picea abies) seedlings to drought stress. Tree Physiol. 2010, 30, 205–213. [Google Scholar]
- Martínez-Vilalta, J.; Poyatos, R.; Aguadé, D.; Retana, J.; Mencuccini, M. A new look at water transport regulation in plants. New Phytol. 2014, 204, 105–115. [Google Scholar] [CrossRef] [Green Version]
- Brodribb, T.J.; McAdam, S.A.M. ABA mediates a divergence in the drought response of two conifers. Plant Physiol. 2013, 162, 1370–1377. [Google Scholar] [CrossRef] [PubMed]
- Brodribb, T.J.; McAdam, S.A.M.; Jordan, G.J.; Martins, S.C.V. Conifer species adapt to low-rainfall climates by following one of two divergent pathways. Proc. Natl. Acad. Sci. USA 2014, 111, 14489–14493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, N.Q. Anatomical, physiological and molecular responses of European beech (Fagus sylvatica L.) to drought. Ph.D. Thesis, Georg-August-University of Göttingen, Göttingen, Germany, February 2016. [Google Scholar]
- Carsjens, C.; Nguyen-Ngoc, Q.; Guzy, J.; Knutzen, F.; Meier, I.C.; Müller, M.; Finkeldey, R.; Leuschner, C.; Polle, A. Intra-specific variation in expression of stress-related genes in beech progenies are stronger than drought-induced responses. Tree Physiol. 2014, 34, 1348–1361. [Google Scholar] [CrossRef] [PubMed]
- Coupel-Ledru, A.; Tyerman, S.D.; Masclef, D.; Lebon, E.; Christophe, A.; Edwards, E.J.; Simonneau, T. Abscisic acid downregulates hydraulic conductance of grapevine leaves in isohydric genotypes only. Plant Physiol. 2017, 75, 1121–1134. [Google Scholar] [CrossRef] [PubMed]
- De Diego, N.; Pérez-Alfocea, F.; Cantero, E.; Lacuesta, M.; Moncaleán, P. Physiological response to drought in radiata pine: phytohormone implication at leaf level. Tree Physiol. 2012, 32, 435–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Diego, N.; Saiz-Fernández, I.; Rodríguez, J.L.; Pérez-Alfocea, P.; Sampedro, M.C.; Barrio, R.J.; Lacuesta, M.; Moncaleán, P. Metabolites and hormones are involved in the intraspecific variability of drought hardening in radiata pine. J. Plant Physiol. 2015, 188, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Munns, R. Genes and salt tolerance: bringing them together. New Phytol. 2005, 167, 645–663. [Google Scholar] [CrossRef]
- Voetberg, G.S.; Sharp, R.E. Growth of the maize primary root at low water potentials: III. Role of increased proline deposition in osmotic adjustment. Plant Physiol. 1991, 96, 1125–1130. [Google Scholar] [CrossRef]
- Ashraf, M.; Foolad, M.R. Roles of glycine betaine and proline in improving plant abiotic stress resistance. J. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Verslues, P.E.; Sharma, S. Proline metabolism and its implications for plant-environment interaction. In The Arabidopsis Book, 1st ed.; American Society of Plant Biologists: Rockville, MD, USA, 2010; p. 23. [Google Scholar]
- Foolad, M.R.; Subbiah, P.; Kramer, C.; Hargrave, G.; Lin, G.Y. Genetic relationships among cold, salt and drought tolerance during seed germination in an interspecific cross of tomato. Euphytica 2003, 130, 199–206. [Google Scholar] [CrossRef]
- Sharma, S.; Verslues, P.E. Mechanisms independent of abscisic acid (ABA) or proline feedback have a predominant role in transcriptional regulation of proline metabolism during low water potential and stress recovery. Plant. Cell. Environ. 2010, 33, 1838–1851. [Google Scholar] [CrossRef] [PubMed]
- Díaz, P.; Betti, M.; Sánchez, D.H.; Udvardi, M.K.; Monza, J.; Márquez, A.J. Deficiency in plastidic glutamine synthetase alters proline metabolism and transcriptomic response in Lotus japonicus under drought stress. New Phytol. 2010, 188, 1001–1013. [Google Scholar] [CrossRef] [PubMed]
- Verbruggen, N.; Hermans, C. Proline accumulation in plants: a review. Amino Acids 2008, 35, 753–759. [Google Scholar] [CrossRef] [PubMed]
- Hare, P.D.; Cress, W.A.; van Staden, J. Dissecting the roles of osmolyte accumulation during stress. Plant Cell. Environ. 1998, 21, 535–553. [Google Scholar] [CrossRef]
- Girousse, C.; Bournoville, R.; Bonnemain, J.L. Water deficit-induced changes in concentrations in proline and some other amino acids in the phloem sap of alfalfa. Plant Physiol. 1996, 111, 109–113. [Google Scholar] [CrossRef] [PubMed]
- Kishor, P.K.; Sangam, S.; Amrutha, R.N.; Laxmi, P.S.; Naidu, K.R.; Rao, K.R.S.; Sreenivasulu, N. Regulation of proline biosynthesis, degradation, uptake and transport in higher plants: its implications in plant growth and abiotic stress tolerance. Curr. Sci. 2005, 88, 424–438. [Google Scholar]
- Lehmann, S.; Funck, D.; Szabados, L.; Rentsch, D. Proline metabolism and transport in plant development. Amino Acids. 2010, 39, 949–962. [Google Scholar] [CrossRef] [Green Version]
- Spollen, W.G.; Tao, W.; Valliyodan, B.; Chen, K.; Hejlek, L.G.; Kim, J.-J.; LeNoble, M.E.; Zhu, J.; Bohnert, H.J.; Henderson, D.; et al. Spatial distribution of transcript changes in the maize primary root elongation zone at low water potential. BMC Plant Biol. 2008, 8, 32. [Google Scholar] [CrossRef]
- Verslues, P.E.; Bray, E.A. Role of abscisic acid (ABA) and Arabidopsis thaliana ABA-insensitive loci in low water potential-induced ABA and proline accumulation. J. Exp. Bot. 2006, 57, 201–212. [Google Scholar] [CrossRef]
- Costa, D.L.C.R.; Lobato, S.D.K.A.; Silvera, D.G.A.J.; Laughinghousevi, D.H. ABA-mediated proline synthesis in cowpea leaves exposed to water deficiency and rehydration. Turk. J. Agric. For. 2011, 35, 309–317. [Google Scholar]
- Sharma, S.; Villamor, J.G.; Verslues, P.E. Essential role of tissue-specific proline synthesis and catabolism in growth and redox balance at low water potential. Plant Physiol. 2011, 157, 292–304. [Google Scholar] [CrossRef] [PubMed]
- Ain-Lhout, F.; Zunzunegui, M.; Barradas, M.C.D.; Tirado, R.; Clavijo, A.; Novo, F.G. Comparison of proline accumula-tion in two Mediterranean shrubs subjected to natural and experimental water deficit. Plant Soil 2001, 230, 175–183. [Google Scholar] [CrossRef]
- Sivakumar, P.; Sharmila, P.; Pardha Saradhi, P. Proline alleviates salt-stress-induced enhancement in ribulose-1,5-bisphosphate oxygenase activity. Biochem. Biophys. Res. Commun. 2000, 279, 512–515. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Descriptors | ‘warm’ PV | ‘cool’ PV |
|---|---|---|
| Locality | Dubeň, Slovakia | Čierny Váh, Slovakia |
| Altitude | 410 m a.s.l. | 931 m a.s.l. |
| Latitude | 49°12′ | 49°00′ |
| Longitude | 18°41′ | 19°58′ |
| Climatic region | moderately humid and warm | humid and moderately cool |
| Mean annual precipitation | 824 mm | 937 mm |
| Mean precipitation of growing season | 469 mm | 557 mm |
| Mean annual air temperature | 7.7 °C | 4.9 °C |
| Mean temperature of growing season | 15.4 °C | 12.3 °C |
| Source | Sum of Squares | F-Ratio | p-Value | Differ. ‘warm’ PV_ D-‘cool’ PV_D | +/− Limits |
|---|---|---|---|---|---|
| PN w | 10.34 | 6.71 | 0.024* | −0.61 | 0.51 |
| PN e-d | 0.19 | 0.13 | 0.721ns | 0.13 | 0.79 |
| PN d | 19.67 | 11.83 | 0.005** | −1.68 | 1.06 |
| PN r | 4.25 | 6.54 | 0.025* | −0.64 | 0.54 |
| gS w | 0.01 | 12.9 | 0.004** | −0.02 | 0.01 |
| gS e-d | 0.01 | 0.17 | 0.684ns | 0 | 0.02 |
| gS d | 0.02 | 34.96 | 0.000* | −0.05 | 0.02 |
| gS r | 0 | 5.55 | 0.036* | −0.02 | 0.02 |
| ABA w | 6.08 × 107 | 40.16 | 0.000*** | 1591.73 | 559.64 |
| ABA e-d | 7 × 105 | 7.08 | 0.024* | 285.03 | 238.71 |
| ABA d | 6.70 × 107 | 42.44 | 0.000*** | 3342.49 | 1143.15 |
| ABA r | 2.70 × 107 | 37.96 | 0.000*** | 1731.25 | 626.11 |
| Proline w | 173.7 | 86.7 | 0.000*** | −2.69 | 0.64 |
| Proline e-d | 133.4 | 26.35 | 0.000*** | −3.85 | 1.67 |
| Proline d | 77.05 | 20.22 | 0.001*** | −3.58 | 1.78 |
| Proline r | 7.87 | 52.82 | 0.000*** | −0.94 | 0.29 |
| ΨS w | 2.06 | 0 | 0.967ns | 0 | 0.05 |
| ΨS e-d | 0.07 | 4.64 | 0.057ns | 0.09 | 0.09 |
| ΨS d | 0.06 | 2.17 | 0.171ns | −0.10 | 0.15 |
| ΨS r | 0 | 1.31 | 0.278ns | −0.02 | 0.04 |
| Ψpd w | 0.48 | 4.03 | 0.072ns | −0.14 | 0.16 |
| Ψpd e-d | 0.02 | 1.49 | 0.251ns | −0.05 | 0.08 |
| Ψpd d | 0.36 | 4.66 | 0.056ns | −0.24 | 0.25 |
| Ψpd r | 0.26 | 1.18 | 0.303ns | −0.17 | 0.35 |
| RWC w | 5.08 | 16.65 | 0.002** | −0.46 | 0.25 |
| RWC e-d | 2.81 | 5.17 | 0.046* | −0.56 | 0.55 |
| RWC d | 7.3 | 28.42 | 0.000*** | −1.10 | 0.46 |
| RWC r | 0.04 | 0.15 | 0.707ns | 0.07 | 0.39 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jamnická, G.; Fleischer, P., Jr.; Konôpková, A.; Pšidová, E.; Kučerová, J.; Kurjak, D.; Živčák, M.; Ditmarová, Ľ. Norway Spruce (Picea abies L.) Provenances Use Different Physiological Strategies to Cope with Water Deficit. Forests 2019, 10, 651. https://doi.org/10.3390/f10080651
Jamnická G, Fleischer P Jr., Konôpková A, Pšidová E, Kučerová J, Kurjak D, Živčák M, Ditmarová Ľ. Norway Spruce (Picea abies L.) Provenances Use Different Physiological Strategies to Cope with Water Deficit. Forests. 2019; 10(8):651. https://doi.org/10.3390/f10080651
Chicago/Turabian StyleJamnická, Gabriela, Peter Fleischer, Jr., Alena Konôpková, Eva Pšidová, Jana Kučerová, Daniel Kurjak, Marek Živčák, and Ľubica Ditmarová. 2019. "Norway Spruce (Picea abies L.) Provenances Use Different Physiological Strategies to Cope with Water Deficit" Forests 10, no. 8: 651. https://doi.org/10.3390/f10080651