Sudden Aspen Decline: A Review of Pattern and Process in a Changing Climate
by
Jack A. Singer
1,*, 
Rob Turnbull
1,
Mark Foster
1,
Charles Bettigole
1,4,
Brent R. Frey
2,
Michelle C. Downey
1,
Kristofer R. Covey
1,3 and
Mark S. Ashton
1 1
Yale University, School of Forestry and Environmental Studies, New Haven, CT 06511, USA
2
Department of Forestry, Mississippi State University, Starkville, MS 39762, USA
3
Program in Environmental Studies and Sciences, Skidmore College, Saratoga Springs, NY 12866, USA
4
GIS Center, Skidmore College, Saratoga Springs, NY 12866, USA
*
Author to whom correspondence should be addressed.
Forests 2019, 10(8), 671; https://doi.org/10.3390/f10080671
Submission received: 28 June 2019
/
Revised: 2 August 2019
/
Accepted: 2 August 2019
/
Published: 9 August 2019
(This article belongs to the Special Issue Dieback on Drought-Prone Forest Ecosystems)
Abstract
:The American quaking aspen (Populus tremuloides Michx.) and its close relative, the Eurasian quaking aspen (Populus tremula L.), cover a realm that is perhaps the most expansive of all tree species in the world. In North America, sudden aspen decline (SAD) is a growing concern that marks the rapid decline of quaking aspen trees leading to mortality at the stand and landscape scale. Research suggests that drought and water stress are the primary causes of SAD. Predisposing factors (age, structure, and landscape position), as well as associated stressors (i.e., pests and pathogens), have been linked to mortality in affected stands. The conflation of multiple interacting factors across the aspen’s broad geographic range in North America has produced significant debate over the classification of SAD as a disease and the proper management of affected stands. Interestingly, no such effects have been reported for the Eurasian aspen. We here review and synthesize the growing body of literature for North America and suggest that SAD is a novel decline disease resulting from multiple inciting and interacting factors related to climate, land-use history, and successional dynamics. We suggest that the range of aspen observed at the onset of the 21st Century was bolstered by a wet period in western North America that coincided with widespread regional cutting and clearing of late-successional forests for timber and grazing. No comparable land-use history, successional status, or age-class structure is apparent or linked for Eurasian forests. Eurasian aspen is either absent or young in managed forests, or old and decadent in parks in Fenno-Scandinavia, or it grows more intimately with a more diverse mixture of tree species that have arisen from a longer period of frequent timber cutting in Russia. Based on these insights we provide recommendations for practical management techniques that can promote stand resilience and recovery across a range of stand conditions in North America. Managers should attempt to identify SAD-prone stands using the presence of predisposing conditions and focus treatments such as coppice or prescribed fire on stands with suitable topographies, elevations, and climates. We conclude that SAD will persist throughout the coming decades, given the enormity of past cutting history, fire exclusion, and current changes in climate until a more active restoration agenda is implemented.
Keywords:
climate warming; disturbance; drought; forest fires; forest dieback; Populus tremula; P. tremuloides; SAD1. Introduction
Quaking aspen (Populus tremuloides Michx., hereinafter aspen) is the most widely distributed tree species in North America [1]. Its close relative, the Eurasian aspen (Populus tremula L.), has a similar distribution throughout all of northern and central Eurasia [2]. Taken together they create their own unique monodominant forests and can be considered the most widespread forest type in the world. In a changing climate, it is therefore of critical importance to understand their response to current and future stresses.
Aspen forests support high understory vegetation diversity [3], provide unique wildlife habitat [4], serve as efficient carbon sinks [5], and have aesthetic value, especially during the fall when the leaves change to golden-yellow [6]. Aspen stands throughout the American West and Eurasia are seral and fundamentally short-lived in nature (typically < 140 years) [5]. Concerns about stand dieback and regeneration have been raised throughout western North America for several decades [7]. However, over the last fifteen years, forest managers in North America have increasingly observed the rapid decline of otherwise healthy aspen stands throughout large parts of its expansive range [8,9]. Interestingly, no such decline has been reported in Eurasia, where forests are much more intensively managed [10].
This increasingly common phenomenon of rapid landscape-scale mortality was first noted in a study [11] and then comprehensively reviewed [7]. With further evidence of widespread aspen dieback throughout its range, this phenomenon has now been termed sudden aspen decline (SAD) [8,9]. Since defining the disease, multiple studies have characterized SAD’s etiology [12], physiology [13,14,15,16,17,18,19], predisposing factors [20,21], secondary pathogens [22], and restoration silviculture [23]. These advancements have been used in conjunction with climate predictions to model SAD’s future impact [9,24].
The existing literature suggests that SAD is best described as a novel decline disease, but like other forest processes, it reflects a variety of factors (i.e., autecology, climate, and land-use) and manifests in complex ways on the landscape as a result [12]. The objective of our paper is to clarify the presence and cause of SAD in North America by synthesizing pre-existing research on the topic, to make management recommendations, and suggest why this appears not to be of importance in Eurasia. This topic has implications for continental-scale landscape changes to forest cover that will continue to rapidly change North American forests in the coming decades. Our review has utility for understanding and examining other decline diseases and forest type changes. We structure our review first with an explanation of SAD, then discuss the main predisposing and contributing factors of SAD, assess future conditions through remote sensing and models, and finally make management recommendations.
2. Explaining Normal Patterns of Aspen Mortality versus SAD
SAD is distinguished from normal age-related decline primarily by how quickly it affects stands. Affected stands exhibit total stand-level collapse and mortality within a short period of time, typically two-to-five years. For instance, Worrall et al. [8] attributed a five-fold increase in aspen mortality over a three-to-four-year period in Southwestern Colorado to SAD. Other authors also describe “rapid” (i.e., 5-year) declines as much faster than the age-related mortality recognized as the successional progression of aspen stand development. SAD can be identified with a much larger suite of exogenous mortality agents primarily caused by drought with secondary interactions from insects and pathogens, and its rapid mortality from these interacting factors [7,25]. Because many aspen stands in western North America are between 60 and 100 years old, the recent increase in the successional decline of aspen and SAD are sometimes conflated to stand age and forest succession but their differences in the rate of aspen mortality and circumstance are actually different (Figure 1; [21]).
Frey et al. [7] discussed the general patterns of age-related decline as compared to rapid stand dieback. In cases of age-related stand decline associated with forest succession, the mortality of individual aspen stems, and the clonal stand as a whole occurs over the course of decades as the aspen gradually break up and are replaced by later successional species. Rapid dieback, in contrast, is characterized by the sudden progressive death of branches in the upper crown and descends downward, resulting in rapid loss of leaf area, reduced vigor, and, unless the inciting event is short-term, widespread mortality within a stand [7]. However, the age of aspen stands varies regionally and is largely driven by the natural fire return interval, averaging 60 years in the Great Lakes, and about 80 years in Northeast Ontario across to Saskatchewan [7]. In the Rocky Mountains, fire return intervals depend upon elevation, aspect, and soil, and can range up to 100 years on the more fire-prone areas and up to several hundred years or more at higher, cooler, and moister elevations [7].
SAD thus represents a rapid acceleration of the autogenic process consistent with a decline disease paradigm. The decline disease paradigm, introduced by Manion [27], posits that multiple interacting factors contribute to forest decline, and these factors may be characterized by their role as inciting, contributing or predisposing factors. Frey et al. [7] first applied this framework in aspen to assess factors involved in rapid dieback of stands in western North America. Subsequent work by Worrall et al. [8,12], and Anderegg et al. [13] extended these assessments, such that a clearer picture of factors driving this process is now evident. SAD is largely understood to be driven by a two-step process: first, acute drought, often accompanied by high temperatures during the growing season, operates as the inciting factor, with secondary factors, such as successional status (older stands), topography (southern aspects), and low site fertility, either singly or in combination, predisposing a stand to decline. Second, contributing factors (usually insects or other pathogens) infiltrate the weakened stand and cause rapid mortality [22,27] (Table 1). Although inciting and secondary factors affect stands in complex ways that vary from stand-to-stand and place-to-place, the result is the same: the rapid and pervasive decline and eventual death of aspen trees across a landscape.
3. Predisposing Factors to Sudden Aspen Decline
3.1. Succession, Forest Structure, and Conifer Competition
Stand-level factors resulting from land-use history and successional status strongly control the extent of aspen mortality during drought [21]. Across the Intermountain West, young stands with a relatively high stem density are at increased risk of mortality following low annual precipitation [21,25]. Aspen mortality, therefore, increases in stands with dense, monodominant even-aged structures characteristic of stem exclusion and/or growth stagnation on drought-sensitive sites. Even-aged, monodominant stands associated with moister sites can exhibit greater vertical structure and heterogeneity and experience less mortality (Figure 1) [21].
Other studies show that understory shrub and conifer recruitment beneath aspen canopies are driven by a combination of (1) an absence of periodic lethal disturbance, and (2) the presence of supra-annual hotter and drier climatic periods. Both in combination promote the rapid transition of quaking aspen stands to more shade and drought-tolerant conifer-dominated stands [5,29,30,31]. Conifers and shrub species limit water resources to the aspen overstory promoting its death and inhibit the establishment of the more shade-intolerant aspen regeneration thereby causing its local extermination [5,29,32] (Figure 1). In the absence of lethal disturbance, aspen may therefore be replaced by slower-growing, more shade-tolerant conifer species, such as white spruce (Picea glauca (Moench) Voss) [33,34], subalpine fir (Abies lasiocarpa (Hooker) Nuttall), and Engelmann spruce (Picea engelmannii Parry x Engelm.) [35,36,37] (Figure 1). Fire suppression has decreased the frequency of stand-replacing disturbances that favor aspen establishment [38,39,40]. During drought, moisture competition from an encroaching understory of shrubs, and conifers renders the aspen canopy especially susceptible to SAD-induced mortality [41,42,43]. Interestingly, one contradictory study by Worrall et al. [12] found no significant relationship between conifer density and overstory aspen crown loss. But we suggest that this is perhaps related to other differences in the site, soil, and climatic range (see the section on edaphic and topographic conditions below).
3.2. Edaphic and Topographic Conditions
Faster-growing, shorter-lived species with high water demand, such as aspen are more vulnerable to mortality during drought events [20,44]. Aspen trees depend on shallow sub-surface water conditions in the soil and show little plasticity in water use; xylem water content in the stem is usually similar to that at the top 5–10 cm of soil moisture [18]. Predisposition to near-surface soil conditions makes aspen particularly vulnerable to soil water loss due to surface evaporation. Areas that are droughty and of low site productivity are, therefore, particularly susceptible to SAD-induced mortality.
The 2002 Colorado Plateau drought combined with poor snowpack and high temperatures in the spring created some of the harshest moisture stress in a century, and the sudden decline of aspen clones occurred in the wake of this drought [18]. SAD mortality was abnormally high in some bottom slopes and flat benches, where topography would not typically cause high moisture stress but low soil moisture associated with shallow soils overlying bedrock was identified as the cause of mortality [18].
In Grand Mesa, Colorado, the high moisture-stressed conditions across a soil gradient helped explain high aspen mortality rates at otherwise suitable elevations [8]. The basaltic soils (a remnant of prehistoric volcanic flows) extant on Grand Mesa form a landslide bench of particularly coarse soils that are excessively well-drained and prone to drought, and therefore, unsuitable for stable, long-term aspen stands. Similar patterns of mortality occurred in Cococino National Forest, northern Arizona, where nearly 95% of living aspen stems died on low elevation xeric sites, as opposed to 61% of living stems at higher elevation mesic sites [45]. SAD-related mortality is, therefore, highest on soils that are moisture-limited, and on desiccation-prone hotter topographies at the xeric margins of aspen’s range, on southern and southwestern aspects, and lower elevations [8,9,12].
4. Contributing Factors to Sudden Aspen Decline
4.1. Primary and Secondary Insects
Secondary fungal pathogens and insects are pervasive SAD mortality agents (Table 2; [9,46,47]). Primary defoliating insects (insects that attack healthy trees, in this case, pre-drought), such as forest tent caterpillar (Malacosoma disstria Hübner, hereinafter FTC) can coexist and interact with SAD. For example, in the Great Lakes region, the boreal woodlands of Saskatchewan, Manitoba, and Alberta, northeastern Ontario, and in the Gila Mountains of Arizona, aspen is subject to massive outbreaks of FTC that greatly decrease live crown area [48,49,50,51]. These areas experienced extreme drought prior to the FTC outbreak that caused larger aspen dieback than attributable solely to the primary defoliators. In areas where primary defoliators were absent (e.g., southwest Colorado, aspen parkland), drought alone accounted for aspen mortality [9,52].
4.2. Primary and Secondary Pathogens
Secondary fungal pathogen and insect attacks have been present in every documented episode of SAD [9]. Pathogens and insects frequently occur in concert (with multiple present on drought-stressed trees) and often differ from those that typically cause a long-term decline in healthy aspen stands [8,22]. Generally, these pathogens attack the vascular system (fungal blights and cankers—Hypoxylon spp., Cryptosphaeria spp.) and cause further crown loss [22,62]. Certain insects, such as bronze poplar borer (Agrilus liragus), can only invade stands when they reach a critical population threshold, and then once SAD affects trees, the invasion of other fungal pathogens (leaf diseases—Melampsoria spp., Venturia spp.) and insects is likely to occur (including in adjacent healthy trees) [22]. The presence of such pathogens, either prior to, during, or after extreme drought conditions, exacerbate typical SAD symptoms and induce mortality [9,22,63].
5. Physiology of SAD
5.1. Proneness to Cavitation and Drought
Landscape-level SAD events have provided a unique opportunity to study the physiological mechanisms of drought-induced tree decline. Research has now investigated the physiological mechanisms of SAD, which can be characterized as a systems-level cascade of failures brought on by initial drought stress [7,19]. Aspen trees rely heavily on current and recent xylem growth for water transport and continued growth [59]; the accumulation of hydraulic damage and reduced growth over multiple drought seasons limits their ability to tolerate secondary pathogens [13,14,15,16,17,19]. Even after a season of normal to above-normal precipitation, SAD-affected trees exhibited a five-fold decrease in hydraulic conductance and lowered levels of gas exchange than healthy trees following major drought events [19]. This type of hydraulic damage may not lead to complete mortality of SAD-affected stands during or immediately after the drought, but losses can be substantial. For example, SAD stricken forests in southwestern Colorado lost approximately 28% of their basal area after the drought in the early 2000s [64].
5.2. Photosynthate Allocation and Carbon Storage
Carbon starvation frequently occurs in drought-stricken trees as an indirect consequence of hydraulic damage to the vascular system. After hydraulic damage, non-structural carbohydrates are allocated from long-term storage to higher priority tasks, particularly maintaining respiration and to compensate for the decline in photosynthesis from reduced water uptake. Because root systems are a more distal sink, root carbon stores are not replenished [65] and root biomass growth decreases—while carbohydrate concentration in stems increases significantly as carbohydrates are mobilized for other higher priority, particularly maintenance respiration [14,15]. The eventual loss of fine root biomass because of carbohydrate starvation is a major consequence of, and a key factor leading to, canopy decline and tree mortality [13,14,15,16,23]. Nonetheless, baseline data of pre-SAD fine root biomass has not been documented, precluding any comparison of SAD effects upon the fine root system. This is an important area of future study. The loss of stored carbohydrates makes trees susceptible to future chronic stressors because no further reserves can allow for their growth and survival [16]. Aspen trees under carbon-starved conditions exhibit decreased chemical defense capacities in the presence of primary defoliators [66], and primary defoliators and SAD-mortality, therefore, can typically co-occur [7].
6. Landscape Level Change through Models and Remote Sensing
6.1. Climate Models and Changes in Aspen Range
The recent rapid decline of aspen has economic, aesthetic, and biodiversity consequences [24]. As a result, projecting the range and distribution of aspen over the coming decades is of significant interest. Climate change is widely predicted to alter the geographic distribution of aspen forests in North America and across Eurasia, as aspen is largely limited in its range by water availability [7,9,31,67].
General Circulation Models (GCMs) have been used to model the aspen’s future distribution in response to climate change. Research predicts a strong and heterogeneous decline in aspen’s range over the coming decades [31]. However, global climate models used to infer micro-site predictions associated with SAD are uncertain [8,68]. Projections indicate that in drier low-elevation sites and higher elevation sites where conifers and shrubs outcompete aspen, increased drought and heat stress will make these locations uninhabitable for aspen [5,8,68,69].
At the same time, aspen is likely to expand into new areas (e.g., higher elevation or northerly aspect sites) as these sites become more suitable for aspen establishment [70]. Fires, bark beetles, and other lethal disturbances in conifer forests will also promote the temporary successional colonization of these newly suitable sites by aspen [24,31]. For example, in the aspen parkland of Alberta, aspen has actually expanded southward in latitude due in part to fire suppression, cropland irrigation, and the elimination of bison [30]. Upslope shifts of aspen forests are already occurring and have been attributed to climate shifts and colonization following fires [32,56]. Nonetheless, many climate models, including IPCC AR-5 projections, conclude that SAD will significantly reduce the footprint of aspen across much of its current range [9,31]. In these scenarios, precipitation events in the intermountain West and boreal region of North America will become less frequent and more intense, while summer temperatures will increase; SAD and other forms of decline will continue to impact aspen across the continent [8,68].
6.2. Detecting Changes in SAD through Remote Sensing
Remotely sensed data supports empirical studies by demonstrating that regional SAD episodes follow prolonged regional drought [9,55]. Paired remotely-sensed data and field measurements across 11.5 million acres of southern boreal parkland forest of western Canada produced strong correlations between biomass loss and drought with estimates that up to 20% of total aboveground biomass died following a SAD event during 2001–2002 [55]. Remotely sensed data showed that aspen trees had the lowest survival on south-facing aspects [55] and at lower elevations in southern Utah where decadent and dying stands were usually monodominant and open relative to healthy stands [54].
Worrall et al. [9] incorporated remote sensing data and IPCC AR-4 projections into a bioclimatic model to determine that exceptional drought often preceded SAD events, which occurred at the species’ margins of habitat suitability. Their study concluded that regeneration potential in most aspen stands has declined significantly.
7. Management Implications: Strategies for Mitigating SAD and Restoring Aspen Forests
Most past studies of SAD have focused on observational studies that rely on detecting large landscape-scale disturbance as the primary mechanism to assess stand dynamics of aspen [71,72,73,74]. More recently, studies have begun to consider silvicultural options for mitigating SAD impacts, including the application of coppice systems, true clearcuts, prescribed fire treatments, and other preventative site treatments. Researchers have also considered the exclusion of large browsing mammals, which threaten aspen regeneration, by using fences and other natural barriers such as logging debris and natural landscape barriers [75].
Aspen’s current range may reflect land-use patterns and climate regimes that no longer exist across the Intermountain West of North America. For example, intensive logging, large prescribed fires, and sheep (Ovis aris L.) grazing in the early part of the twentieth century all led to favorable conditions after these disturbances stopped for aspen establishment and growth as even-aged monodominant stands. But fire suppression and over-browsing by elk (Cervus canadaensis Erxleben) led to a 40%–60% aerial decline in its extent by the late 1990s [76,77,78]. Aspen is therefore unstable across much of its current range in the Rocky Mountains, especially at the margins of its southern range, at a relatively lower elevation, and on southern aspects with more xeric hotter sites [8,24,31]. Aspen stands are most likely to be maintained on mesic sites, at mid-elevation, and on northern aspects.
From our review of the literature on SAD, we conclude this is very much a North American phenomenon. We found an absence of reported studies on SAD or SAD-like symptoms in the Eurasian literature. We believe this is partly due to the fact that both Canada and the United States have research communities and monitoring programs that are better funded than those in countries like Russia and the Baltics (Lithuania, Latvia, Estonia); and when research is reported there it is in a language and publication form (grey literature from government research institutions) that is often inaccessible to the larger global research community We were not able to access this information though it might well exist. However, Scandinavia and Finland have very strong research communities and monitoring programs, but no SAD has been reported on any widespread scale [79,80]. This is perhaps largely because of the more intensive forest management practices in these countries where extensive aspen stands do not exist outside of protected areas, and within protected areas, they are often old and decadent [10]. As such, there is a call for forest restoration that includes a large component of deciduous trees, including aspen, particularly in intensely managed forests of the region [81].
There are climatic differences between Eurasia and North America, particularly throughout the aspen’s range in the Rocky Mountains. There is no such equivalent mountain range in Eurasia other than the much older and lower elevation Urals. The Rocky Mountains provide an avenue for aspens much more fragmented southern extension in latitude into a more strongly arid and seasonal climate—well below its more traditional boreal range; but as noted above it is restricted to the moist, cool high elevation areas that comprise large snowpacks. This is contrary to the North American aspen’s range throughout boreal Canada, which we believe to be similar in climate to the boreal of Eurasia. SAD is prevalent in the Canadian boreal as well where aspen can dominate in extensive monodominant stands potentially regulated by fire [7].
There are interesting observational anomalies. Like, Scandinavia and Finland, the Maritime provinces of Canada, northern and northeastern U.S. and the coastal Pacific Northwest and Alaska appear unaffected by SAD. We conclude, again that this is perhaps a combination of changes in climate (moist year-round), disturbance regime (absence of extensive periodic fires), land-use history (extensive timbering and/or grazing) and richer competing assemblages of other tree species (northern hardwoods), that make SAD inconsequential. We suggest more observation and study is necessary to understand these differences. We suspect that if SAD is to be found in Eurasia, it is likely in the least populous and underreported region of northern-central Eurasia where the climate, disturbance type (fire), and land-use history (heavy extensive cutting), is similar to the northern interior of North America; as well as in a more fragmented and site-sensitive way to the Rocky Mountains.
7.1. Coppice Systems
Clear felling and its impact on aspen regeneration have evolved over time, especially in the recent past with respect to SAD. Foresters historically relied on coppice cuttings, which have light-to-moderate impacts on soil and herbaceous groundstory. This facilitates aspen vegetative regeneration by triggering the formation and release of dormant and adventitious root buds by increasing soil temperature, nutrient and light availability, and decreasing competition [82,83,84,85]. Coppice cutting in stands experiencing low to moderate mortality in the initial stages of SAD can promote natural sprout origin regeneration, mitigate the loss of root carbohydrate reserves, and limit hydraulic stress following intense drought [12,17,18,23,83]. Understory trees and areas subject to recent coppice do not exhibit the rapid mortality seen in the overstory of uncut aspen dominant stands. This corresponds to the autecology of vegetative regeneration in aspen: small-sized clonally regenerating trees are better supported by the large residual root systems than adult trees. Treating stands that have already experienced severe decline is less effective; hydraulic root failure and impeded sucker growth were observed in such stands following treatments [12,23]. Implementing this strategy effectively can be challenging because it requires assessing stand health and timing a cut. Researchers have developed an index that may lessen this challenge. The index classifies stands as low-moderate-high in SAD condition based on the percentage of basal area mortality, crown loss, or a combination thereof [5,12,23].
Encouraging a wider range of tree size classes within an aspen stand may also promote resiliency [12]. This approach would require intensive variable density and free-form thinning and the conversion to multiple age-classes though patch and group coppice selection systems—with openings large enough to allow direct sunlight and to encourage sprouting [86]. This can be done in conjunction with groundstory prescribed burning that limits conifer encroachment and decreases the likelihood of a stand to experience SAD or successional decline [86,87].
7.2. Prescribed Fire
The reintroduction of fire may be a useful SAD management tool, although managers should carefully consider stand structure before burning [23,88]. Aspen stands can be classified into five distinct types on the basis of mean fire severity and annual fire probability: (1) fire-independent, stable aspen; (2) fire-influenced, stable aspen; (3) fire-dependent, seral, conifer-aspen; (4) fire-dependent, seral, montane conifer-aspen; and (5) fire-dependent, seral, subalpine, conifer-aspen (81). These fire regimes, as well as other stand factors (structural composition, presence of invasive plants, and desired management goals), can influence forest management decisions in regards to timing and nature of the regeneration harvest, site preparation treatments, and type of thinning that can be done [23,88].
Aspen regenerates vigorously following fire. Smith et al. [56] found that recruitment was strong following a severe fire, despite high herbivory density. They recommend prescribed fire wherever landscape-scale aspen maintenance is desired and either (1) the percentage of overstory conifer stems exceeds 80% or (2) overstory aspen density is less than 200 overstory stems per hectare. Conversely, in stands experiencing conifer encroachment, lower rates of aspen regeneration may occur post-fire, though this response could be attributable to site conditions and the presence of serotinous conifer seeds in the seed bank [58].
Fire regimes will certainly change under the influence of climate change, and the interaction with aspen’s (and SAD’s) shifting range. This creates uncertainty in stand dynamics where aspen currently dominates [56,89,90,91]. Yang et al. [90] found that an increase in mean annual temperature by 2 °C–5 °C shifts the upper elevation limit of aspen trees following fires in conifer forest, suggesting selective use of fire may be appropriate to encourage the establishment of aspen on suitable sites at higher elevations (e.g., by implementing a true clear cut and prescribed burn in overstocked conifer stands at higher elevations, with more mesic soils, and appropriate aspect). On these fire-treated sites, aspen may establish through wind-dispersed seed, if available, as opposed to the more prevalent clonal reproduction—potentially increasing the genetic diversity of aspen trees and leading to more resilient progeny [56,58,92].
Fire severity also plays a role in the extent of aspen regeneration. In at least one study, regeneration stem densities of aspen were between 60% and 135% greater in plots subjected to high severity burns versus either unburned or low-severity plots [28]. While prescribed fire increases stem density, it will cause significant stem mortality to the residual overstory, which should be expected given aspen’s susceptibility to fire damage because of its thin bark [89]. Care is sometimes needed to limit mortality and reduce stress following prescribed burns. Options to accomplish these goals include irrigation and protection from grazing and browse [89]. The limited number of studies conducted on this topic indicate that a moderate fire disturbance regime may prevent conifer encroachment and promote suckering in certain stands, with more severe fire disturbance potentially allowing for aspen seeds to colonize a site [25,89,92].
8. Conclusions
Sudden aspen decline is a novel decline disease complex induced by drought and exacerbated by a variety of factors—including stand structure, land-use history, secondary pathogen attack, topography, and inter- and intra-specific competition. The uncertain future of aspen in North America will be dependent upon the continuing prevalence of SAD. We expect instances of SAD to continue, and be largely incited by extreme drought events associated with climate change but that this will be dependent upon site, the condition and history of the stand, and landscape position. SAD will be most prevalent on drier and hotter sites and soils, with southern aspects, where stands, because of cutting history and fire exclusion, have created even-age aggregates of aspen that now compete with conifers. Such areas and circumstances are more prone to various pests and pathogens making them even more susceptible to SAD. We believe that focusing the promotion of aspen on colder, moister sites, with the reintroduction of periodic fires, and the creation of smaller even-aged stands across a landscape mosaic that comprises a more heterogeneous age class distribution can all build resilience to SAD. It is therefore important to be mindful of regional climate projections that predict more frequent droughts in the context of the land-use history that originally facilitated the recruitment and persistence of existing stands. While early management intervention might build more resilient stands on some sites, others may be best managed to promote other forest types.
Author Contributions
K.R.C. provided the initial idea. J.A.S., R.T., and M.F. wrote the initial draft with guidance from K.R.C., C.B., and M.C.D. provided figures, edits, and comments on further drafts. M.S.A. provided substantial edits and reworked the manuscript with substantial new writing working with J.A.S., B.R.F. provided further edits and comments. J.A.S. coordinated the final completion.
Acknowledgments
Funding and support for this research were provided by the Yale School of Forestry and Environmental Studies, in conjunction with the Ucross High Plains Stewardship Initiative. We would like to thank Ian Carney of Two Shoes Ranch and Jason Jones of Aspen Tree Services in Carbondale, Colorado, for sharing their expertise and ranch. We also must thank William Lauenroth for his comments on reviewing an early draft. We received no direct funds to support this study, but J.A.S., R.T., and M.F. were supported by partial Scholarship from the School of Forestry and Environmental Studies. No funds were secured to cover the costs of publishing in open access.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Perala, D.A. Populus Tremuloides. In Silvics of North America: Volume 2. Hardwoods; Burns, R.M., Honkala, B.H., Eds.; US Department of Agriculture: Washington, DC, USA, 1990; pp. 555–569. [Google Scholar]
- De Carvalho, D.; Ingvarsson, P.K.; Joseph, J.; Suter, L.; Sedivy, C.; Macaya-Sanz, D.; Cottrell, J.; Heinze, B.; Schanzer, I.; Lexer, C. Admixture facilitates adaptation from standing variation in the European aspen (Populus tremula L.), a widespread forest tree. Mol. Ecol. 2010, 19, 1638–1650. [Google Scholar] [CrossRef] [PubMed]
- Kay, C.E. The condition and trend of aspen, Populus tremuloides, in Kootenay and Yoho National Parks: Implications for ecological integrity. Can. Field-Nat. 1997, 111, 607–616. [Google Scholar]
- Lindroth, R.L.; Clair, S.B.S. Adaptations of quaking aspen (Populus tremuloides Michx.) for defense against herbivores. For. Ecol. Manag. 2008, 299, 14–21. [Google Scholar] [CrossRef]
- Morelli, T.L.; Carr, S.C. Review of the Potential Effects of Climate Change on Quaking Aspen (Populus tremuloides) in the Western United States and a New Tool for Surveying Aspen Decline; USDA Forest Service Pacific Southwest Research Station, General Technical Report PSW-GTR-235; USDA Forest Service: Washington, DC, USA, 2011. [CrossRef]
- McCool, S.F. Quaking aspen and the human experience: Dimensions, issues, and challenges. In Sustaining Aspen in Western Landscapes: Symposium Proceedings, Grand Junction, CO, USA, 13–15 June 2000; Shepperd, W.D., Binkley, D., Bartos, D.L., Stohlgren, T.J., Eskew, L.G., Eds.; Proceedings RMRS-P-18; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2001; pp. 147–162. [Google Scholar]
- Frey, B.R.; Lieffers, V.J.; Hogg, E.H.; Landhausser, S.M. Predicting landscape patterns of aspen dieback: Mechanisms and knowledge gaps. Can. J. For. Res. 2004, 34, 1379–1390. [Google Scholar] [CrossRef]
- Worrall, J.J.; Egeland, L.; Eager, T.; Mask, R.A.; Johnson, E.W.; Kemp, P.A.; Shepperd, W.D. Rapid mortality of Populus Tremuloides in southwestern Colorado, USA. For. Ecol. Manag. 2008, 255, 686–696. [Google Scholar] [CrossRef]
- Worrall, J.J.; Rehfeldt, G.E.; Hamann, A.; Hogg, E.H.; Marchetti, S.B.; Michaelian, M.; Gray, L.K. Recent declines of Populus tremuloides in North America linked to climate. For. Ecol. Manag. 2013, 299, 35–51. [Google Scholar] [CrossRef]
- Latva-Karjanmaa, T.; Penttilä, R.; Siitonen, J. The demographic structure of European aspen (Populus tremula) populations in managed and old-growth boreal forests in eastern Finland. Can. J. For. Res. 2007, 37, 1070–1081. [Google Scholar] [CrossRef]
- Hogg, E.H.; Brandt, J.P.; Kochtubaida, B. Growth and dieback of aspen forests in northwestern Alberta, Canada, in relation to climate and insects. Can. J. For. Res. 2002, 32, 823–832. [Google Scholar] [CrossRef]
- Worrall, J.J.; Marchetti, S.B.; Egeland, L.; Mask, R.A.; Eager, T.; Howell, B. Effects and etiology of sudden aspen decline in southwestern Colorado, USA. For. Ecol. Manag. 2010, 260, 638–648. [Google Scholar] [CrossRef]
- Anderegg, W.R.L.; Anderegg, L.D.L.; Sherman, C.; Karp, D.S. Effects of widespread drought-induced aspen mortality of understory plants. Conserv. Biol. 2012, 26, 1082–1090. [Google Scholar] [CrossRef]
- Anderegg, W.R.L.; Berry, J.A.; Field, C.B. Linking definitions, mechanisms, and modeling of drought-induced tree death. Trends Plant Sci. 2012, 17, 1360–1385. [Google Scholar] [CrossRef] [PubMed]
- Anderegg, W.R.L.; Berry, J.A.; Smith, D.D.; Sperry, J.S.; Anderegg, L.D.L.; Field, C.B. The roles of hydraulic and carbon stress in a widespread climate-induced forest die-off. Proc. Natl. Acad. Sci. USA 2012, 109, 233–237. [Google Scholar] [CrossRef] [PubMed]
- Anderegg, W.R.L. Complex aspen forest carbon and root dynamics during drought. Clim. Chang. 2012, 111, 983–991. [Google Scholar] [CrossRef]
- Anderegg, L.D.L.; Anderegg, W.R.L.; Abatzoglou, J.; Hausladen, A.M.; Berry, J.A. Drought characteristics’ role in widespread Aspen forest mortality across Colorado, USA. Glob. Chang. Biol. 2013, 19, 1526–1537. [Google Scholar] [CrossRef] [PubMed]
- Anderegg, W.R.L.; Plavcova, L.; Anderegg, L.D.L.; Hacke, U.G.; Berry, J.A.; Field, C.B. Drought’s legacy: Multiyear hydraulic deterioration underlies widespread aspen forest die-off and portends increased future risk. Glob. Chang. Biol. 2013, 19, 1188–1196. [Google Scholar] [CrossRef] [PubMed]
- Anderegg, W.R.L.; Anderegg, L.D.L.; Berry, J.A.; Field, C.B. Loss of whole-tree hydraulic conductance during severe drought and multi-Year forest die-Off. Oecologia 2014, 175, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Ireland, K.B.; Moore, M.M.; Fule, P.Z. Slow lifelong growth predisposes Populus tremuloides trees to mortality. Oecologia 2014, 175, 847–859. [Google Scholar] [CrossRef]
- Bell, D.M.; Bradford, J.B.; Lauenroth, W.K. Forest stand structure, productivity, and age mediate climatic effects on aspen decline. Ecology 2014, 95, 2040–2046. [Google Scholar] [CrossRef]
- Marchetti, S.B.; Worrall, J.J.; Eager, T. Secondary insects and diseases contribute to sudden aspen decline in southwestern Colorado, USA. Can. J. For. Res. 2011, 41, 2315–2325. [Google Scholar] [CrossRef]
- Shepperd, W.D.; Smith, F.W.; Pelz, K.A. Group clearfell harvest can promote regeneration of aspen forests affected by sudden aspen decline in western Colorado. For. Sci. 2015, 61, 932–937. [Google Scholar] [CrossRef]
- Kulakowski, D.; Kaye, M.W.; Kashian, D.M. Long-term aspen cover change in the western US. For. Ecol. Manag. 2013, 299, 52–59. [Google Scholar] [CrossRef]
- Hogg, E.H.; Brandt, J.P.; Michaelian, M. Impacts of a regional drought on the productivity, dieback, and biomass of western Canadian aspen forests. Can. J. For. Res. 2008, 38, 1373–1384. [Google Scholar] [CrossRef] [Green Version]
- Oliver, C.D.; Larson, B.C. Forest Stand Dynamics; McGraw Hill Publishing Company: New York, NY, USA, 1990. [Google Scholar]
- Manion, P.D. Tree Disease Concepts, 2nd ed.; Prentice-Hall Inc.: Upper Saddle River, NJ, USA, 1991. [Google Scholar]
- Petty, J.L. Bionomics of two aspen bark beetles, Trypophloeus populi and Procryphalus mucronatus (Coleoptera: Scolytidae). Great Basin Nat. 1977, 37, 105–127. [Google Scholar]
- Zegler, T.J.; Moore, M.M.; Fairweather, M.L.; Ireland, K.B.; Fulé, P.Z. Populus Tremuloides mortality near the southwestern edge of its range. For. Ecol. Manag. 2012, 282, 196–207. [Google Scholar] [CrossRef]
- Rogers, P.C.; Landhausser, S.M.; Pinno, B.D.; Ryel, R.J. A functional framework for improved management of Western North American aspen (Populus tremuloides Michx.). For. Sci. 2014, 60, 345–359. [Google Scholar] [CrossRef]
- Rehfeldt, G.E.; Worrall, J.J.; Marchetti, S.B.; Crookston, N.L. Adapting forest management to climate change using bioclimate models with topographic drivers. For. Int. J. For. Res. 2015, 88, 528–539. [Google Scholar] [CrossRef] [Green Version]
- Bretfeld, M.; Franklin, S.B.; Peet, R.K. A multiple-scale assessment of long-term aspen Persistence and elevational range shifts in the Colorado Front Range. Ecol. Monogr. 2016, 86, 244–260. [Google Scholar] [CrossRef]
- Milakovsky, B.; Frey, B.R.; Ashton, M.S.; Larson, B.C.; Schmitz, O.J. Influences of gap position, vegetation management and herbivore control on survival and growth of white spruce (Picea glauca (Moench) Voss) seedlings. For. Ecol. Manag. 2011, 261, 440–446. [Google Scholar] [CrossRef]
- Rutenbeck, N.E.; Frey, B.R.; Covey, K.R.; Berlyn, G.P.; Schmitz, O.J.; Larson, B.C.; Ashton, M.S. Influence of gap position and competition control on the leaf physiology of planted Picea glauca and natural regeneration of Populus tremuloides. For. Ecol. Manag. 2018, 424, 228–235. [Google Scholar] [CrossRef]
- Heinselman, M.L. Fire and succession in the conifer forests of northern North America. In Forest Succession; West, D.C., Shugart, H.H., Botkin, D.B., Eds.; Springer: New York, NY, USA, 1981. [Google Scholar] [CrossRef]
- Romme, W.H.; Knight, D.H. Fire frequency and subalpine forest succession along a topographic gradient in Wyoming. Ecology 1981, 62, 319–326. [Google Scholar] [CrossRef]
- Van Cleve, K.; Viereck, L.A. Forest succession in relation to nutrient cycling in the boreal forest of Alaska. In Forest Succession; West, D.C., Shugart, H.H., Botkin, D.B., Eds.; Springer: New York, NY, USA, 1981. [Google Scholar]
- Frelich, L.E.; Reich, P.B. Spatial patterns and succession in a Minnesota southern-boreal forest. Ecol. Monogr. 1995, 65, 325–346. [Google Scholar] [CrossRef]
- Wadleigh, L.; Jenkins, M.J. Fire frequency and the vegetative mosaic of a spruce-fir forest in northern Utah. Great Basin Nat. 1996, 56, 28–37. [Google Scholar]
- Bergeron, Y.; Cyr, D.; Drever, C.R.; Flannigan, M.; Gauthier, S.; Kneeshaw, D.; Lauzon, È.; Leduc, A.; Goff, H.L.; Lesieur, D.; et al. Past, current, and future fire frequencies in Quebec’s commercial forests: Implications for the cumulative effects of harvesting and fire on age-class structure and natural disturbance-based management. Can. J. For. Res. 2006, 36, 2737–2744. [Google Scholar] [CrossRef]
- Choat, B.; Brodribb, T.J.; Brodersen, C.R.; Duursma, R.A.; López, R.; Medlyn, B.E. Triggers of tree mortality under drought. Nature 2018, 558, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.E.; Smith, F.W. Twenty-year change in aspen dominance in pure aspen and mixed aspen/conifer stands on the Uncompahgre Plateau, Colorado, USA. For. Ecol. Manag. 2005, 213, 338–348. [Google Scholar] [CrossRef]
- Binkley, D.; Alsanousi, A.; Romme, W.H. Age Structure of aspen forests on the Uncompahgre Plateau, Colorado. Can. J. For. Res. 2014, 44, 836–841. [Google Scholar] [CrossRef]
- Hanna, P.; Kulakowski, D. The influences of climate on aspen dieback. For. Ecol. Manag. 2012, 274, 91–98. [Google Scholar] [CrossRef]
- Fairweather, M.L.; Geils, B.W.; Manthei, M. Aspen decline on the Coconino National Forest. In Proceedings of the 55th Annual Western International Forest Disease Work Conference, Sedona, AZ, USA, 15–19 October 2007; McWilliams, M., Palacios, P., Eds.; Oregon Department of Forestry: Salem, OR, USA, 2008; pp. 53–62. [Google Scholar]
- Dudley, M.M.; Burns, K.S.; Jacobi, W.R. Aspen Mortality in the Colorado and southern Wyoming Rocky Mountains: Extent, severity, and causal factors. For. Ecol. Manag. 2015, 353, 240–259. [Google Scholar] [CrossRef]
- Dudley, M.M.; Negron, J.; Tisserat, N.A.; Shepperd, W.D.; Jacobi, W.R. Influence of climate on the growth of quaking aspen (Populus Tremuloides) in Colorado and Southern Wyoming. Can. J. For. Res. 2015, 45, 1546–1563. [Google Scholar] [CrossRef]
- Hildahl, V.; Reeks, W.A. Outbreaks of the forest tent caterpillar, Malacosoma disstria Hbn, and their effects on stands of trembling aspen in Manitoba and Saskatchewan. Can. Entomol. 1960, 92, 199–209. [Google Scholar] [CrossRef]
- Roland, J. Large-scale forest fragmentation increases the duration of tent caterpillar outbreak. Oecologia 1993, 93, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Roland, J.; Mackey, B.G.; Cooke, B. Effects of climate and forest structure on duration of forest tent caterpillar outbreaks across central Ontario, Canada. Can. Entomol. 1998, 130, 703–714. [Google Scholar] [CrossRef]
- Cooke, B.J.; Lorenzetti, F. The dynamics of forest tent caterpillar outbreaks in Quebec, Canada. For. Ecol. Manag. 2006, 226, 110–121. [Google Scholar] [CrossRef]
- Michaelian, M.; EHogg, H.; Hall, R.J.; Arsenault, E. Massive mortality of aspen following severe drought along the southern edge of the Canadian boreal forest. Glob. Chang. Biol. 2011, 17, 2084–2094. [Google Scholar] [CrossRef]
- Worrall, J.L.; Egeland, T.; Eager, R.; Mask, E.; Johnson, P.K.; Shepherd, W. Sudden aspen decline in southwest Colorado: Site and stand factors and a hypothesis on etiology. In Proceedings of the 55th Western International Forest Disease Work Conference, Sedona, AZ, USA, 15–19 October 2007; Oregon Department of Forestry: Salem, OR, USA, 2007; Volume 55, pp. 15–19. [Google Scholar]
- Evans, D.M. A Spatiotemporal analysis of aspen decline in southern Utah’s Cedar Mountain, using remote sensing and Geographic Information Systems. Master’s Thesis, Utah State University, Logan, UT, USA, 2010. Available online: https://digitalcommons.usu.edu/etd/734 (accessed on 20 December 2017).
- Huang, C.-Y.; Anderegg, W.R.L. Large drought-induced aboveground live biomass losses in southern Rocky Mountain aspen Forests. Glob. Chang. Biol. 2012, 18, 1016–1027. [Google Scholar] [CrossRef]
- Smith, E.A.; O’Loughlin, D.; Buck, J.R.; Clair, S.B.S. The influences of conifer succession, physiographic conditions and herbivory on Quaking aspen regeneration after fire. For. Ecol. Manag. 2011, 262, 325–330. [Google Scholar] [CrossRef]
- Bell, D.M.; Bradford, J.B.; Lauenroth, W.K. Scale dependence of disease impacts on quaking aspen (Populus tremuloides) mortality in the southwestern United States. Ecology 2015, 96, 1835–1845. [Google Scholar] [CrossRef] [PubMed]
- Krasnow, K.D.; Stephens, S.L. Evolving paradigms of aspen ecology and management: Impacts of stand condition and fire severity on vegetation dynamics. Ecosphere 2015, 6, 1–16. [Google Scholar] [CrossRef]
- Sperry, J.S.; Nichols, K.L.; Sullivan, J.E.M.; Eastlack, S.E. Xylem embolism in ring-porous, diffuse-porous, and coniferous trees of northern Utah and interior Alaska. Ecology 1994, 75, 1736–1752. [Google Scholar] [CrossRef]
- Blodgett, J.T.; Allen, K.K.; Schotzko, K.; Dymerski, A. Aspen health on National Forests in the Northern Rocky Mountain Region. 2017. Available online: https://digitalcommons.usu.edu/cgi/ (accessed on 8 September 2017).
- Rice, J.; Bardsley, T.; Gomben, P.; Bambrough, D.; Weems, S.; Huber, A.; Joyce, L.A. Assessment of aspen ecosystem vulnerability to climate change for the Uinta-Wasatch-Cache and Ashley National Forests; Utah. Gen. Tech. Rep. RMRS-GTR-366; U.S. Department of Agriculture, Forest Service: Fort Collins, CO, USA, 2017; 67p. Available online: https://www.fs.fed.us/rm/pubs_series/rmrs/gtr/rmrs_gtr366.pdf (accessed on 8 September 2017).
- Juzwik, J.W.; Nishijima, T.; Hinds, T.E. Survey of aspen cankers in Colorado. Plant Dis. Rep. 1978, 62, 906–910. [Google Scholar]
- Guyon, J.; Hoffman, J. Survey of Aspen Dieback in the Intermountain Region. USDA Forest Service. Available online: https://digitalcommons.usu.edu/aspen_bib (accessed on 21 November 2017).
- Worrall, J.J.; Keck, A.G.; Marchetti, S.B. Populus tremuloides stands continue to deteriorate after drought-incited sudden aspen decline. Can. J. For. Res. 2015, 45, 1768–1774. [Google Scholar] [CrossRef]
- Landhäusser, S.M.; Lieffers, V.J. Defoliation increases risk of carbon starvation in root systems of mature aspen. Trees 2012, 26, 653. [Google Scholar] [CrossRef]
- Najar, A.; Landhausser, S.M.; Whitehill, J.G.A.; Bonello, P.; Erbilgin, N. Reserves accumulated in non-photosynthetic organs during the previous growing season drive plant defenses and growth in aspen in the subsequent growing season. J. Chem. Ecol. 2014, 40, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Ulo, N.; Valladares, F. Tolerance to shade, drought, and waterlogging of temperate Northern Hemisphere trees and shrubs. Ecol. Monogr. 2006, 76, 521–547. [Google Scholar]
- Rehfeldt, G.E.; Ferguson, D.E.; Crookston, N.L. Aspen, climate, and sudden decline in Western USA. For. Ecol. Manag. 2009, 258, 2353–2364. [Google Scholar] [CrossRef]
- Sexton, J.O.; Ramsey, D.R.; Bartos, D.L. Habitone analysis of quaking aspen in the Utah Book Cliffs: Effects of site water demand and conifer Cover. Ecol. Model. 2006, 198, 301–311. [Google Scholar] [CrossRef]
- Landhäusser, S.M.; Deshaies, D.; Lieffers, V.J. Disturbance facilitates rapid range expansion of aspen into higher elevations of the Rocky Mountains under a warming climate. J. Biogeogr. 2010, 37, 68–76. [Google Scholar] [CrossRef]
- Weigle, W.G.; Frothingham, E.H. The Aspens: Their Growth and Management; United States Department of Agriculture, Forest Service: Washington, DC, USA, 1911.
- Baker, F.S. Aspen in the Central Rocky Mountain Region; United States Department of Agriculture: Washington, DC, USA, 1925; p. 1291.
- Sampson, A.W. Effect of Grazing Upon Aspen Reproduction (No. 741); US Department of Agriculture: Washington, DC, USA, 1919.
- Schier, G.A. Physiological and environmental factors controlling vegetative regeneration of aspen. Utilization and Marketing as Tools for Aspen Management in the Rocky Mountains. Hessel, D.L., Betters, D.R., Loring, T.J., Porter, L.D., Bennett, J.E., Eds.; Aspen Bibliography. Paper 5007. 1976. Available online: https://digitalcommons.usu.edu/aspen_bib/5007 (accessed on 20 December 2017).
- Kota, A.M.; Bartos, D.L. Evaluation of techniques to protect aspen suckers from ungulate browsing in the Black Hills. West. J. Appl. For. 2010, 25, 161–168. [Google Scholar]
- White, C.A.; Olmsted, C.E.; Kay, C.E. Aspen, elk, and fire in the Rocky Mountain national parks of North America. Wildl. Soc. Bull. 1998, 26, 449–462. [Google Scholar]
- Bartos, D.L. Landscape dynamics of aspen and conifer forests. In Sustaining Aspen in Western Landscapes: Symposium Proceedings; Proceedings RMRS-P-18; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2001. [Google Scholar]
- Hessl, A.E.; Graumlich, L.J. Interactive effects of human activities, herbivory and fire on quaking aspen (Populus tremuloides) age structures in western Wyoming. J. Biogeogr. 2002, 29, 889–902. [Google Scholar] [CrossRef]
- Vanha-Majamaa, I.; Lilja, S.; Ryömä, R.; Kotiaho, J.S.; Laaka-Lindberg, S.; Lindberg, H.; Puttonen, P.; Tamminen, P.; Toivanen, T.; Kuuluvainen, T. Rehabilitating boreal forest structure and species composition in Finland through logging, dead wood creation and fire: The EVO experiment. For. Ecol. Manag. 2007, 250, 77–88. [Google Scholar] [CrossRef]
- Esseen, P.A.; Ehnström, B.; Ericson, L.; Sjöberg, K. Boreal forests. Ecol. Bull. 1997, 46, 16–47. [Google Scholar]
- Härkönen, S.; Eerikäinen, K.; Lähteenmäki, R.; Heikkilä, R. Does moose browsing threaten European aspen regeneration in Koli National Park, Finland. Alces J. Devoted Biol. Manag. Moose 2008, 44, 31–40. [Google Scholar]
- Fraser, E.C.; Lieffers, V.J.; Landhäusser, S.M.; Frey, B.R. Soil nutrition and temperature as drivers of root suckering in trembling aspen. Can. J. For. Res. 2002, 32, 1685–1691. [Google Scholar] [CrossRef]
- Frey, B.R.; Lieffers, V.J.; Landhäusser, S.M.; Comeau, P.G.; Greenway, K.J. An analysis of sucker regeneration of trembling aspen. Can. J. For. Res. 2003, 33, 1169–1179. [Google Scholar] [CrossRef]
- Landhäusser, S.M.; Wan, X.; Chow, P.S.; Lieffers, V.J. Nitrate stimulates root suckering in trembling aspen (Populus tremuloides). Can. J. For. Res. 2010, 40, 1962–1969. [Google Scholar] [CrossRef]
- Stoeckler, J.H.; Macon, J.W. Regeneration of aspen cut-over areas in Northern Wisconsin. J. For. 1956, 54, 13–16. [Google Scholar]
- Ashton, M.S.; Kelty, M.J. The Practice of Silviculture: Applied Forest Ecology, 10th ed.; John Wiley and Sons: Hoboken, NJ, USA, 2018. [Google Scholar]
- Bartos, D.L.; Campbell, R.B. Decline of quaking aspen in the interior West--examples from Utah. Rangel. Arch. 1998, 20, 17–24. [Google Scholar]
- Shinneman, D.J.; Baker, W.L.; Rogers, P.C.; Kulakowski, D. Fire regimes of quaking aspen in the Mountain West. For. Ecol. Manag. 2013, 299, 22–34. [Google Scholar] [CrossRef]
- Krasnow, K.D.; Halford, A.S.; Stephens, S.L. Aspen restoration in the eastern Sierra Nevada: Effectiveness of prescribed fire and conifer removal. Fire Ecol. 2012, 8, 104–118. [Google Scholar] [CrossRef]
- Yang, J.; Weisberg, P.J.; Shinneman, D.J.; Dilts, T.E.; Earnst, S.L.; Scheller, R.M. Fire modulates climate change response of simulated aspen distribution across topoclimatic gradients in a semi-arid montane landscape. Landsc. Ecol. 2015, 30, 1055–1073. [Google Scholar] [CrossRef]
- Fairweather, M.L.; Rokala, E.A.; Mock, K.E. Aspen seedling establishment and growth after wildfire in central Arizona: An instructive case history. For. Sci. 2014, 60, 703–712. [Google Scholar] [CrossRef]
- Keyser, T.L.; Smith, F.W.; Shepperd, W.D. Trembling aspen response to a mixed-severity wildfire in the Black Hills, South Dakota, USA. Can. J. For. Res. 2005, 35, 2679. [Google Scholar] [CrossRef]
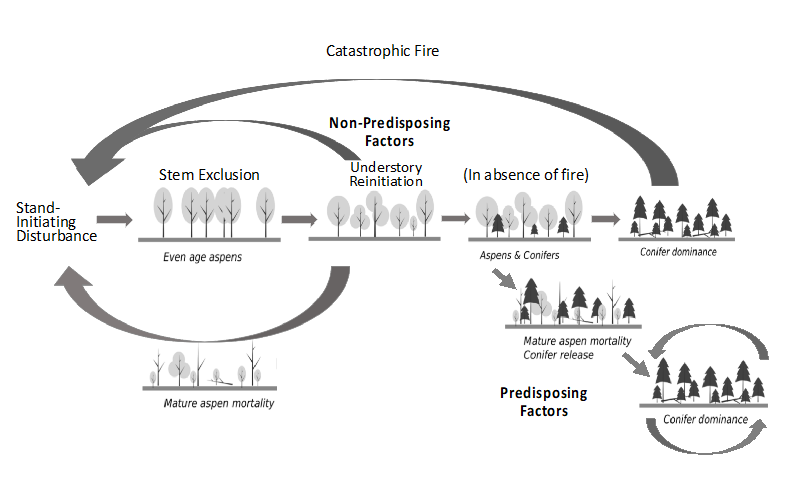
Figure 1.
Possible successional trajectories of aspen stands in the Rocky Mountains. Aspen develops as even-aged stands following a large-scale initiating disturbance. Following stem exclusion understory re-initiation will produce suppressed suckers and an herbaceous ground story [26]. In fertile sites where aspen can self-perpetuate, or in the presence of some intense disturbance, such as fire, stands return to initiation. In the absence of fire, conifer encroachment can convert the stand, either permanently (self-perpetuating conifer stand in the bottom right corner—usually facilitated by conditions suited to SAD), or temporarily, returning to seral aspen following another stand-replacing fire. Conifer species can vary across the range of aspen, including lodgepole and juniper in drier sites, and spruce and fir at higher elevations.
Figure 1.
Possible successional trajectories of aspen stands in the Rocky Mountains. Aspen develops as even-aged stands following a large-scale initiating disturbance. Following stem exclusion understory re-initiation will produce suppressed suckers and an herbaceous ground story [26]. In fertile sites where aspen can self-perpetuate, or in the presence of some intense disturbance, such as fire, stands return to initiation. In the absence of fire, conifer encroachment can convert the stand, either permanently (self-perpetuating conifer stand in the bottom right corner—usually facilitated by conditions suited to SAD), or temporarily, returning to seral aspen following another stand-replacing fire. Conifer species can vary across the range of aspen, including lodgepole and juniper in drier sites, and spruce and fir at higher elevations.

{kind=link}
Table 1.
We list the various predisposing, and contributing factors of SAD, which synergize to transform a healthy aspen stand into sudden decline and dieback. These parameters are derived primarily from Frey et al., [7]; but modified by more recent information from work by Worrall et al., [12], Anderegg et al., [13], and Morelli and Carr [5], with supporting information from Petty [28].
Table 1.
We list the various predisposing, and contributing factors of SAD, which synergize to transform a healthy aspen stand into sudden decline and dieback. These parameters are derived primarily from Frey et al., [7]; but modified by more recent information from work by Worrall et al., [12], Anderegg et al., [13], and Morelli and Carr [5], with supporting information from Petty [28].
| Characteristics of a Healthy Aspen Stand | Predisposing Factors | Inciting Factor | Contributing Factors | Characteristics of Affected Stands |
|---|---|---|---|---|
| Closed canopy Vigorous suckering Rapid growth Fine root mass growth Mixed-age stand (Values vary regionally) | Aspect (South or Southwest) Lower elevation Physiological maturity Low site index High stand density | Climate-induced drought | Bronze poplar borer (Agrilus liragus Barter and Brown 1949) Bark beetles (Trypophloeus populi, Hopkins 1915; Procryphalus mucronata, LeConte, 1879) Forest tent moth caterpillar (Malacosoma disstria Hübner, 1820) Cytospora canker (Cytospora chrysosperma, (Pers.:Fr) Fr) Armillaria root rot (Armillaria spp.) | Rapid die-off of previously healthy stands in 2–5 years Decreased photosynthesis Increased tree carbohydrate concentrations Decline in sap flow Smaller leaf area Decline in sap flow Root biomass decline Loss of hydraulic conductance Earlier leaf-shedding in autumn |
Table 2.
Key studies that studied SAD. We examine their methods, key findings—which range from characterizing the disease to implementing management strategies—and the language used to discuss aspen die-off.
Table 2.
Key studies that studied SAD. We examine their methods, key findings—which range from characterizing the disease to implementing management strategies—and the language used to discuss aspen die-off.
| Author | Year | Methods | Key Findings | Terminology for SAD |
|---|---|---|---|---|
| Frey et al. | 2004 [7] | Review | Insect defoliation, drought, and thaw–freeze events appear to be the most likely factors initiating dieback in mature aspen stands. | Sudden dieback of mature plants |
| Worrall et al. | 2008 [8] | Aerial surveys, Geospatial Analysis, and field observation. | Predisposing factors include stand maturation, low density, southern aspects, and low elevations. A major inciting factor was the recent, acute drought accompanied by high temperatures. Sites with poor regeneration and weak root systems may exhibit clonal death and long-term aspen forest cover loss. | SAD |
| Worrall et al. | 2007 [53] | Used geographic information from the 2006 aerial survey on aspen damage, together with the aspen cover type. | Extreme drought with little regeneration after overstory loss incited rapid dieback of aspen in Southwest Colorado. Predisposing environmental and insect and/or pathenogenic damage also contributed. | SAD |
| Evans | 2010 [54] | Regression analyses and a topographic analysis using zonal statistics were performed to determine climatic factors and landscape positions that correlated to aspen decline prevalence. | The most significant predictor of aspen decline was elevation, which was significantly greater in the live aspen for three of the five years. Drought weakens aspen, making it susceptible to future decline. | SAD |
| Worrall et al. | 2010 [12] | To test the role of climate as an inciting factor for SAD, a landscape-scale climate model was used to compare the moisture status of declining and healthy aspen at the height of the warm drought in 2002. | Overstory age and diameter were not related to SAD severity. The severity of SAD was inversely and weakly related to the basal area, stem slenderness, and site index, and positively related to upper slope positions. | SAD |
| Anderegg | 2012 [16] | Compared potted and naturally occurring aspen to conduct a water deprivation experiment. | Increased allocation to root non-structural carbohydrates is a direct response to drought in aspen and plays an important role in the die-off. | Wide-spread aspen die-off |
| Marchetti et al. | 2011 [22] | Compared insects and diseases in 162 damaged and neighboring healthy plots to determine contributing factors and their ecological roles. | Cytospora canker, bronze poplar borer, and aspen bark beetles were the primary agents associated with crown loss and other factors related to SAD. Environmental stress may have increased host susceptibility. | SAD |
| Michaelian et al. | 2011 [55] | Used plot-based, meteorological, and remote sensing measures to examine aspen die-off following an exceptionally severe drought. | Spatial variation in the percentage of dead biomass showed a moderately strong correlation with drought severity. | aspen mortality |
| Morelli and Carr | 2011 [5] | Literature review | Complex, unpredictable future for aspen in the West, where increased drought, ozone, and insect outbreaks will vie with carbon dioxide fertilization and warmer soils, resulting in unknown cumulative effects. | SAD |
| Hanna and Kulakowski | 2012 [44] | Tested the influence of climatic variability on aspen growth and mortality in northwestern Colorado and southern Wyoming using dendroecological methods. | Aspen growth was inhibited by warm temperatures, except at the highest elevations. Mortality frequency was associated with multiple years of drought. | aspen dieback |
| Huang and Anderegg | 2012 [56] | Combined field measures, remote sensing and a digital elevation model in SAD affected areas in southwest Colorado. | SAD clustered on south-facing slopes due to relatively drier and warmer conditions, but no apparent spatial gradient was found for elevation and slope. | SAD |
| Zegler et al. | 2012 [29] | Collected data from a random sample of 48 aspen sites to determine the relationship of predisposing site and stand factors and contributing agents to tree mortality. | Relative conifer basal area and density, the incidence of canker disease and wood-boring insects, and slope were significantly associated with regeneration mortality. | aspen decline, aspen mortality |
| Anderegg et al. | 2013 [18] | Drew upon multiple sources of climate data to characterize the drought that triggered aspen mortality. | High 2002 summer temperature and low shallow soil moisture were associated with the spatial patterns of aspen mortality. | Widespread aspen forest mortality |
| Anderegg et al. | 2013 [17] | Tested whether accumulated hydraulic damage can predict the probability of tree survival over 2 years. | Hydraulic damage persisted and increased in dying trees over multiple years and exhibited few signs of repair. | SAD |
| Kulakowski et al. | 2013 [24] | Literature review | Future aspen trends will depend on the net result of direct (drought) and indirect (forest fires, bark beetle outbreaks) effects of altered climate. | Major aspen decline |
| Worrall et al. | 2013 [9] | Range-wide bioclimate model characterizing climatic controls of aspen distribution. | Researchers expect a substantial loss of suitable habitat within the current distribution. | SAD |
| Anderegg et al. | 2014 [19] | Monitored quaking aspen trees over two growing seasons, including a severe summer drought. | SAD-affected trees exhibited lower whole-tree hydraulic conductance and assimilation than healthy trees. | SAD |
| Bell et al. | 2014 [21] | Examined the relation of mortality index to forest structure and climate in the Rocky Mountains and intermountain west. | Drought mortality may be influenced by stand development, inter-species competition, and vulnerabilities of large trees to drought. | SAD |
| Ireland et al. | 2014 [20] | Tree-ring investigation of growth patterns and mortality | The initial growth rate was not associated with a longer lifespan. Younger trees with lower recent growth and more abrupt growth have an increased risk of mortality. | SAD, widespread dieback of aspen forests |
| Bell et al. | 2015 [57] | Modeled disease prevalence in live aspen stems and survival rates near the species’ southern range limit. | Mortality depends on tree size, allometry, competition, summer temperature, summer precipitation. | Mortality of diseased trees |
| Dudley et al. | 2015a [46] | Surveyed aspen stands with various mortality levels in CO and WY | Cankers, bark beetles, and wood borers were the most common damage agents. | SAD, aspen mortality |
| Dudley et al. | 2015b [47] | Analyzed a series of increment cores | Found a relationship between tree growth and annual precipitation but not summer precipitation. | SAD |
| Krasnow and Stephens | 2015 [58] | Compared regeneration dynamics of pre-fire stand composition to post-fire aspen regeneration. | Greater disturbance severity increased sprout density. Live conifer and/or dead aspen basal area prior to fire disturbance reduced sprout density. | SAD |
| Shepperd et al. | 2015 [23] | Clearfelled half of stands to compare with uncut half. | It is possible to successfully regenerate SAD-affected stands, provided that treatment occurs before the majority of the aspen are dead. | SAD |
| Worrall et al. | 2015 [59] | Diseased aspen stands were paired with a neighboring healthy aspen plot. | Diseased plots had much more recent damage than healthy plots. | SAD |
| Bretfeld et al. | 2016 [32] | Sampled 89 plots in the Colorado front range from 305 plots sampled in 1972–1973. | 22 plots no longer contained aspen. Upslope shifts suggest climate-related responses and migrations. | SAD |
| Blodgett et al. | 2017 [60] | Researchers measured and tagged mature trees and sapling density. | No significant tree mortality events have occurred. | SAD |
| Rice et al. | 2017 [61] | Literature review | A dynamic spatial and temporal response to climate change is expected. | Sudden aspen mortality |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Singer, J.A.; Turnbull, R.; Foster, M.; Bettigole, C.; Frey, B.R.; Downey, M.C.; Covey, K.R.; Ashton, M.S. Sudden Aspen Decline: A Review of Pattern and Process in a Changing Climate. Forests 2019, 10, 671. https://doi.org/10.3390/f10080671
AMA Style
Singer JA, Turnbull R, Foster M, Bettigole C, Frey BR, Downey MC, Covey KR, Ashton MS. Sudden Aspen Decline: A Review of Pattern and Process in a Changing Climate. Forests. 2019; 10(8):671. https://doi.org/10.3390/f10080671
Chicago/Turabian StyleSinger, Jack A., Rob Turnbull, Mark Foster, Charles Bettigole, Brent R. Frey, Michelle C. Downey, Kristofer R. Covey, and Mark S. Ashton. 2019. "Sudden Aspen Decline: A Review of Pattern and Process in a Changing Climate" Forests 10, no. 8: 671. https://doi.org/10.3390/f10080671
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.