
Calcium Biogeochemical Cycle in a Typical Karst Forest: Evidence from Calcium Isotope Compositions


1
Institute of Earth Sciences, China University of Geosciences (Beijing), Beijing 100083, China
2
GEOMAR Helmholtz-Zentrum für Ozeanforschung Kiel, Wischhofstr. 1-3, 24148 Kiel, Germany
*
Author to whom correspondence should be addressed.
Forests 2021, 12(6), 666; https://doi.org/10.3390/f12060666
Submission received: 16 April 2021
/
Revised: 20 May 2021
/
Accepted: 22 May 2021
/
Published: 25 May 2021
(This article belongs to the Special Issue Stable Isotope Applications in Forest Ecosystems)
Abstract
:In order to better constrain calcium cycling in natural soil and in soil used for agriculture, we present the δ44/40Ca values measured in rainwater, groundwater, plants, soil, and bedrock samples from a representative karst forest in SW China. The δ44/40Ca values are found to differ by ≈3.0‰ in the karst forest ecosystem. The Ca isotope compositions and Ca contents of groundwater, rainwater, and bedrock suggest that the Ca of groundwater primarily originates from rainwater and bedrock. The δ44/40Ca values of plants are lower than that of soils, indicating the preferential uptake of light Ca isotopes by plants. The distribution of δ44/40Ca values in the soil profiles (increasing with soil depth) suggests that the recycling of crop-litter abundant with lighter Ca isotope has potential effects on soil Ca isotope composition. The soil Mg/Ca content ratio probably reflects the preferential plant uptake of Ca over Mg and the difference in soil maturity. Light Ca isotopes are more abundant in mature soils than nutrient-depleted soils. The relative abundance in the light Ca isotope (40Ca) is in the following order: farmland > burnt grassland > forests > grassland > shrubland. Our results further indicate that biological fractionation in a soil–plant system is a vital factor for Ca–geochemical transformations in soil surface systems.
1. Introduction
As an essential nutrient for plants, calcium (Ca) in organics-rich forest ecosystems (including mineral soils) is generally absorbed via the finer feeder roots of trees [1]. In particular, a major threat to forest ecosystem health is linked to the loss of environmental Ca [2]; therefore, many studies have focused on Ca sources and sinks in soils, plants, and the atmosphere across forest ecosystems [3,4,5,6]. Previous work has reported the use of 87Sr/86Sr (radiogenic Sr isotopic systems) and elemental ratios of Ba/Sr and Ca/Sr as typical tools for exploring Ca sources in forest ecosystems [7,8,9,10], while the application of Ca isotopes has been relatively limited due to high-precision measurements being difficult to perform. Radiogenic strontium isotope ratios, commonly used for distinguishing the sources of alkaline earth elements [8,11,12,13,14] due to its similar behavior to Ca in plant uptake processes and in ionic radius (rSr/rCa = 1.13), are now more often applied in studies of the biogeochemical cycle in forest ecosystem [7,9,10,12,15]. Although Ca and Sr are both alkaline earth elements, previous studies have suggested that these two elements are different [16,17]. In addition, the ionic behaviors of Ca2+ and Sr2+ are different in the long-term development of a forest ecosystem [18,19,20,21].
In recent studies, Ca isotope fractionation has been used in marine carbonate studies [22,23,24,25,26] and stable Ca isotopes have been applied directly to determine the biogeochemical cycle of elements in different ecosystems on Earth [3,4,21,27,28,29,30,31,32,33,34,35,36,37,38]. The results highlight the complex effects of biologically mediated trace metal cycling in forest ecosystems [3,4,21,29,31,38,39,40].
Many studies have confirmed that large amounts of Ca are available in the processes that internally circulate organic matter, in vegetation, and in the soil of forest ecosystems [41]. The litter-decomposition rates determine the Ca mineralization rates in vegetation, which varies significantly among different plant species [42,43]. For example, previous studies indicated that the base cation (including Ca and Mg) demands of American basswood and sugar maple are relatively high [44], resulting in different element turnover rates. Currently, whether Ca and Mg vary significantly in the bottom soil and surface soil under different vegetation covers is still unclear. In the present study, the δ44/40Ca isotope compositions and Mg/Ca ratios of different forest ecosystem samples (rainwater, groundwater, soil, vegetation, and rock) were determined to understand the biogeochemical cycle of elements in Maolan areas, a representative subtropical karst forest in southwest China. In parallel, Ca in different pools were determined directly to further explore cycling processes involving Ca in karst forests. The main objectives are (1) to identify whether the stable isotope of Ca can be used to obtain quantified information about Ca uptake by plants during their life span and (2) to determine whether the Ca isotope composition of vegetation and associated soils can be used to better constrain Ca budgets in forests.
2. Materials and Methods
2.1. Geological Setting
In order to explore Ca biogeochemistry in a karst soil–plant system, a representative karst forest (Maolan National Natural Reserve Park, MNNRP) located in the southeast region of Guizhou Province (China) with a cover area of ≈213 km2 and an elevation of ≈500 m (Figure 1) was selected. The terrain is high in the northwest and low in the southeast. Well known for its good vegetation and rich fauna, the study area has a hot and humid tropical monsoon climate with an annual mean air temperature of 15.3 ℃. The wet season lasts from April to October, while the dry season lasts from November to March. The annual precipitation is ≈1750 mm [45]. MNNRP is primarily covered by an evergreen broad-leaved forest (≈88%), with plants such as incense cedar and Taxus chinensis, that effectively reduces water loss and soil erosion. Moreover, MNNRP has a well-developed karst landform with jagged carbonate rock (shallow marine carbonate) as well as sporadic sandstones (Figure 1). Black calcareous soil (shallow layer) dominates the study region due to lithological control (poor soil-forming capacity). Bare rocks on the ground lead to surface discontinuity, depleted water-holding capacity of the soil, and Ca-rich soil (salt base). Nevertheless, forest vegetation plays a key role in improving soil quality in MNNRP and in shaping the normal landscape and zonal vegetation.
2.2. Sampling Procedure
The calcium isotopic compositions of rainwater, groundwater, soils, forest tree leaves, crop leaves, and bedrock samples were measured. As shown in Figure 1, four rainwater samples were collected at the R site from June to September in 2008. Nine groundwater samples from springs and wells (≈5 m depth) were collected using a pre-cleaned sampler and bottles in July 2007 within the MNNRP (more details on the groundwater sites can be found in Reference [46]). Fifteen soil samples were collected from five typical soil profiles at different depths in the summer of 2007 [47], including farmland (LBP; 0, 40, and 110 cm), burnt grassland (LBM; 0, 40, and 70 cm), shrubland (LBS; 0, 40, and 90 cm), virgin forests (LBF; 0, 40, and 120 cm), and grassland (LBG; 0, 40, and 110 cm). Forest tree leaves, including the three most representative tree leaves from Nandina domestica, Folium Platycaryae, and Handliodendron bodinier, were collected during the summer of 2007 near the virgin forest soil site (LBF). Two crop leaves from soybean and corn were also collected at the farmland soil site (LBP) in 2007. A typical bedrock (dolomite rock) sample was collected near the virgin forest soil site (LBF) in 2007. The fresh plant samples were cleaned via ultrapure water, freeze dried, then put into a crushing device immersed in liquid nitrogen, and smashed into powder (<150 µm). All of the soil and bedrock samples were air-dried at ambient conditions and ground to less than 150 µm. The soils of five land use types had different pH and SOC values [47]. The cation concentrations of rainwater and groundwater were derived from our previous work [46,48].
2.3. Chemical Analysis
For digestion of the plant, soil, and bedrock samples, (i) 100 mg of the sample powder was digested by 4 mL of 1:3 (v/v) HF and HNO3 in a PFA sample jar (Savillex co., Eden Prairie, MN, USA) and then heated at 140 °C on a hotplate (≈7 d) to ensure complete digestion of the fluorides, carbon, and silicates [49,50,51]. The samples were dried subsequently. (ii) The dried samples were dissolved again in 4 mL of 1:3 (v/v) HF and HNO3 using the same digestion procedure repeatedly until the solution became limpid. (iii) After complete digestion and drying, pure HNO3 (2 mL) was applied two times to remove the fluorin and then vaporized using a hotplate. (iv) The remaining substance after digestion and after drying was dissolved in 100 mL volumetric flasks with 2% HNO3. All of the digestion processes were performed in an ultra-clean laboratory at the Institute of Geochemistry, Chinese Academy of Sciences (IGCAS). The concentrations of major cations were measured using ICP-OES in IGCAS. The measurement of a procedural blank, the standard reference material (GSB071192 and GSB071193), and sample replicates were performed during whole analyzation processes to ensure quality control. The accuracy of cations was better than ±5% for the quality control standards. The measurements of rainwater and groundwater samples were the same as those in our previous work [46,48].
2.4. Calcium Isotope Analysis
The double spike method was applied to measure Ca isotope compositions following the previous studies [30,52]. The 43Ca and 48Ca double spikes were added into all samples before the purification procedure. In the mixed solution of the sample and spike, the 40Ca and 44Ca were all from the sample Ca while most of the 43Ca and 48Ca came from the spike Ca. Thus, according to the measured ratios for 40Ca/48Ca, 44Ca/48Ca, and 43Ca/48Ca (spike), all of the non-naturally derived Ca isotope fractionations during purification and measurements could be corrected. Calcium was separated from other elements on cation exchange resin columns (BioRad AGW 50x8 resin) using 1.5N HCl as an eluent, concluding a Ca blank of <2 ng (<0.7%) [52]. After drying, the mixed solution of the purified sample (about 300 ng Ca) and the spike were re-dissolved using 2.2 N HCl (1 µL). All of the purification procedures were conducted at GEOMAR [52]. For the thermal ionization mass spectrometer (TIMS) measurements of the Ca isotopes, the “sandwich technique (TaCl5-activator solution)” and the out-gassed single filament (rhenium) were applied to preload the re-dissolved solution. Briefly, about 0.5 μL of TaCl5-activator solution was first added onto the filament and heated to near dryness. Then, the re-dissolved sample (1 µL) was added on top of the activator solution and dried again. Another 0.5 μL of the activator solution was then added onto the filament and dried [52]. Finally, the Ca isotopes were measured via Triton T1 TIMS in GEOMAR following the method described in a previous work [52]. Two data-acquisition steps were performed in measurement processes since the cup configuration of Triton T1 TIMS does not cover the Ca mass range (40 to 48 amu) in a single step. In other words, the data for masses 40, 42, 43, and 44 were collected in the first step while that for 43 and 48 were detected simultaneously in the second step. Moreover, 41K was also measured to evaluate the potential influence on mass 40K isobaric interferences. The details for the double spike correction of the Ca isotope data can be found in a previous study [52]. Typically, the Triton T1 TIMS presented a 40Ca signal intensity of 7–9 V during the measurement. The Ca isotope composition was expressed as δ44/40Ca = [(44Ca/40Ca)sample/(44Ca/40Ca)standard − 1] × 1000. NIST SRM 915a was applied as the standard reference material for the calculation of δ44/40Ca. The average value for repeat measurements was defined as the external precision (2SE). NIST SRM 915a presented a mean external precision of ±0.09‰ (2SD of the δ44/40Ca values, reproducibility between runs, n = 4) and showed a mean 44Ca/40Ca value of 0.021182 ± 0.000006 (2σ, n = 36) during the entire measurement process.
3. Results
The Ca isotope compositions of the samples collected from MNNRP are summarized in Table 1 and Figure 2. The values measured from the forest ecosystem samples vary between −1.44‰ and 1.48‰, corresponding to a ≈3.0‰ natural variation in δ44/40Ca.
3.1. δ44/40Ca Ratios of Rainwater
3.2. δ44/40Ca Ratios of Groundwater
The δ44/40Ca values of groundwater varied from 0.40 ± 0.14‰ to 1.08 ± 0.21‰ and are significantly heavier than the δ44/40Ca values of dissolved Ca reported in rivers worldwide (−1.71 ± 0.23‰ to −0.63 ± 0.07‰) [16]. The δ44/40Ca values of groundwater in MNNRP are also higher than those of the Yangtze River draining into carbonate-dominated catchments (−0.63 ± 0.07‰) [26]. This interval overlaps with the δ44/40Ca value of bedrock (0.36 ± 0.09‰) and rainwater (0.55 ± 0.24‰ to 1.01 ± 0.15‰).
3.3. δ44/40Ca Ratios of Bedrock and Soils
The carbonate bedrock sample yielded a δ44/40Ca value of 0.36 ± 0.09‰ (Table 1). The Ca concentrations of the surface soil pools in different soil profiles varied from 20 to 106 mmol/kg. Among the soil profiles, only the Ca concentration of grasslands increased with depth and relatively high Ca concentrations were found in the surface soil of farmland and virgin forests, while no obvious patterns were observed in the other profiles (Table 1). As shown in Figure 4, the δ44/40Ca values of surface soil varied from −0.51 ± 0.25‰ to 0.29 ± 0.15‰, lower than that of rainwater, groundwater, and bedrock. However, the pools of Ca in bottom soil varied from 0.91 ± 0.18‰ to 1.48 ± 0.21‰, higher than that of bedrock and surface soils. The δ44/40Ca values of soil increased with depth in the soil profiles but were different for each soil profile. Simultaneously, it can be seen that the δ44/40Ca values of surface farmland is the lowest (Figure 5), indicating the depletion of heavier isotopes in surface farmland relative to other soils. In contrast, grassland and shrubland show the highest δ44/40Ca values, demonstrating 44Ca enrichment compared with the virgin forests.
3.4. δ44/40Ca Ratios of Vegetation
Vegetation yields the lowest δ44/40Ca values and the highest Ca concentrations in the forest ecosystem (Table 1). Nandian domestica, a typical shrub in Maolan, generates leaves characterized by a δ44/40Ca value of −0.26 ± 0.18‰. Folium platycaryae and Handliodendron bodinier are two typical arbors widely distributed in Maolan with δ44/40Ca values of −0.22 ± 0.08‰ and −0.26 ± 0.00‰ in their leaves, respectively. All of the δ44/40Ca values from the three natural leave samples are similar.
Crops yield the lowest δ44/40Ca values and relatively high Ca concentrations (Table 1) in comparison to other plants. Soybean is a typical crop in Maolan characterized by a δ44/40Ca value of −1.44 ± 0.14‰. The δ44/40Ca value of Maize is −1.35 ± 0.20‰.
4. Discussion
4.1. Atmospheric Inputs of Calcium
In general, two distinct sources are suggested to be the main contributors of Ca in groundwater: one is atmospheric input, and the other is bedrock weathering. Although the lowest Ca concentrations were observed in rainwater (12.9–24.0 μmol/L) and the Ca concentration in groundwater (1163–2538 μmol/L) is tens of times greater than that in rainwater, the range of δ44/40Ca values in rainwater (0.55‰ to 1.01‰) is completely within the range of δ44/40Ca values in groundwater (0.40‰ to 1.08‰) (Table 1). A previous study observed similar results in rainwater, snow, and throughfall samples in the Strengbach catchment [27]. Therefore, δ44/40Ca values in groundwater reflect atmospheric input to a certain extent. Carbonate rocks generally have high Ca concentrations and relatively low δ44/40Ca values [53,54,55,56]. In MNNRP, bedrock has the highest Ca concentration (6066 mmol/kg) and a δ44/40Ca value (0.36‰) close to that for groundwater. Thus, both local carbonate rock weathering and atmospheric inputs tend to be responsible for (a potential source of) the Ca isotope compositions of groundwater in MNNRP, which also indicates that carbonate dissolution does not give rise to significant Ca isotopic fractionation. Our 87Sr/86Sr data also support this finding (more details will be published elsewhere).
4.2. Plant and Soil Calcium
Generally, Ca is absorbed by plants via fine lateral roots, then transported, and redistributed to different organs through the xylem sap. After autumn, mineralized Ca in the litter of leaves and fruits is migrated to soil solutions, where it is partially reabsorbed by plants [41,57]. During pedogenesis, the contents of Ca and Mg in the soil decrease due to the combined influence of plant uptake (as important micronutrients for plants) and the rock weathering process (yielding elements/soluble components in the soil and further being potentially removed by groundwater [58,59]). The increasing Mg/Ca ratio shows a higher plant utilization rate for Ca than for Mg (or a potential higher groundwater-removal rate of Ca than for Mg), probably indicating soil maturation and that pedogenesis started. Considering the comparable effects of the weathering process and the subsequent groundwater-removal process on soil Ca and Mg contents in a local region, it is reasonably assumed that the longer the pedogenic time, the higher the Mg/Ca ratio in soils may become under the impact of plants. The δ44/40Ca value of bottom virgin forests (120 cm) is as high as ≈1.48‰, and the δ44/40Ca value of soybean is as low as −1.44‰ (Table 1 and Figure 4). Therefore, the total range of δ44/40Ca values in the Maolan karst forest ecosystem is about 3.0‰. This observation is in line with the reported results. In other words, the Ca uptake by plant (as nutrient) is an important reason for Ca isotope fractionation on the Earth’s surface [16,27,31,38]. The light Ca isotopes are favored by plant absorption and results in the enrichment of heavy Ca isotopes in soil pools. This can be demonstrated particularly from the soybean (δ44/40Ca = −1.44‰) and corn (δ44/40Ca = −1.35‰) samples, which are both related to farmland (surface soil δ44/40Ca = −0.51‰). It also can be seen from Table 2 that δ44/40Ca values of surface soil tend to decrease with the increase in Mg/Ca ratios (reflecting the increase in extraction and usage of Ca from the soil), except for shrubland; the δ44/40Ca values of surface soil was negatively correlated with the Mg/Ca ratios (r = −0.89, p < 0.05). The increment of vegetation coverage and soil maturation lead to elemental discrimination and isotope fractionation in soils.
A previous study also suggested that the total amount and the available pools of C, N, and P are the highest for forests and are less for shrublands and grasslands in the study area [60]. Moreover, the increasing δ44/40Ca values were observed to be accompanied by more soil development in a longer pedogenic process in Ohia tree-dominated Hawaiian rainforests [21]. Although the δ44/40Ca values in five soil profiles varied to different degrees (Figure 4), our data imply that the soil δ44/40Ca values always increased with depth in the soil profiles with different vegetation covers. Surface pristine grassland and shrubland show the highest soil δ44/40Ca values, whereas surface farmland shows the lowest soil δ44/40Ca value (Table 1 and Figure 4). The lowest soil δ44/40Ca value observed in surface farmland may reflect the recycling of crop-litter-rich organic matter, which tends to be isotopically lighter than the underlying soil. This also reflects the result of the Ca nutrient demand in crops: the light Ca isotopes were preferentially extracted by the roots so that the soil was relatively enriched with heavy Ca isotopes. The biological fraction effects can be a reasonable explanation for the fractionation of Ca isotopes. This can also be supported by the results of a previous study which concluded that the isotope compositions of roots are dominated by the source variations and fractionation mechanisms, whereas the translocation mechanism is governed by fractionation processes [61]. Moreover, considering the different depths of plants roots, the δ44/40Ca of net fractionation processes would be higher if the roots spread within high δ44/40Ca value soil, which is an important direction of further research.
4.3. Local-Scale Ca Cycling Effects in Maolan Karst Forest Ecosystem
Our data confirm that the plant uptake of Ca (as nutrient) is an important source of Ca isotope fractionation on the Earth’s surface [21,38,57]. Although we did not calculate the Ca budgets in the forest ecosystem, Ca–geochemical transformations occur in a surface system. The vegetation leaves are enriched with 40Ca compared with soil: the light Ca isotopes are preferentially absorbed by plants, resulting in heavy Ca isotopes remaining in the soil. Soil mineral weathering-derived calcium presented a higher δ44/40Ca value (0.91‰ to 1.48‰) than bedrock (0.36‰). Therefore, mineral weathering cannot explain the heavy δ44/40Ca compositions of soil pools.
The Maolan forest catchment may only be representative of a typical karst ecosystem, but the significant vegetation-related biological uptake of Ca flux is observed. Therefore, its consideration in the future is necessary. As a function of ambient variations with time, the processes of Ca isotopic enrichment in different environmental systems (e.g., different plant–soil systems) is of great significance in correcting the Ca biogeochemical cycle model [57].
5. Conclusions
We observed that rainwater and groundwater are more enriched with heavy 44Ca than bedrock and that soils and plants are more abundant in light 40Ca. The greater abundance of 40Ca in nutrient-rich soils than in nutrient-depleted soils reflects the decomposition and recycling of plant litter. The sequence of abundance in 40Ca is as follows: farmland > burnt grassland > forests > grassland > shrubland. The evolution of Ca isotopes in karst forest ecosystems implies that intensely weathered soils have increased δ44/40Ca values and that the internal circulation of a nutrient pool controls the Ca cycle of forest ecosystems. The Ca isotope compositions of plants might record the fractionation produced via the dynamic equilibrium between soils and plants. Moreover, the soil Mg/Ca content ratio and the Ca isotope compositions probably reflect the preferential plants uptake of Ca over Mg and the different soil maturities. This study highlights that Ca isotopes may be useful in exploring (or quantifying in the future) the origins and sinks of Ca in forest ecosystems.
Author Contributions
Conceptualization, G.H.; data curation, G.H.; formal analysis, G.H.; funding acquisition, G.H.; investigation, G.H.; methodology, G.H. and A.E.; project administration, G.H.; resources, G.H.; software, G.H. and M.L.; supervision, G.H.; validation, G.H.; visualization, G.H.; writing—original draft, G.H., J.Z. and M.L.; writing—review and editing, G.H. and J.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded jointly by National Natural Science Foundation of China, grant numbers 41661144029 and 41325010.
Data Availability Statement
The data presented in this study are available in Table 1.
Acknowledgments
The authors gratefully acknowledge Ana Kolevica and Florian Böhm for laboratory assistance and technical support. We also thank Qiu Tan and Yang Tang for sampling assistance and thank Shitong Zhang for the English polishing.
Conflicts of Interest
The authors declare no conflict of interests.
References
- Momoshima, N.; Bondietti, E.A. Cation binding in wood: Applications to understanding historical changes in divalent cation availability to red spruce. Can. J. For. Res. 1990, 20, 1840–1849. [Google Scholar] [CrossRef]
- Likens, G.E.; Driscoll, C.T.; Buso, D.C. Long-term effects of acid rain: Response and recovery of a forest ecosystem. Science 1996, 272, 244. [Google Scholar] [CrossRef]
- Hindshaw, R.S.; Bourdon, B.; Pogge von Strandmann, P.A.E.; Vigier, N.; Burton, K.W. The stable calcium isotopic composition of rivers draining basaltic catchments in Iceland. Earth Planet. Sci. Lett. 2013, 374, 173–184. [Google Scholar] [CrossRef]
- Hindshaw, R.S.; Reynolds, B.C.; Wiederhold, J.G.; Kiczka, M.; Kretzschmar, R.; Bourdon, B. Calcium isotope fractionation in alpine plants. Biogeochemistry 2013, 112, 373–388. [Google Scholar] [CrossRef] [Green Version]
- Huntington, T.G. Assessment of calcium status in Maine forests: Review and future projection. Can. J. For. Res. 2005, 35, 1109–1121. [Google Scholar] [CrossRef]
- Lawrence, A.D.; Bu, J.; Gokulakrishnan, P. The interactions between SO2, NOx, HCl and Ca in a bench-scale fluidized combustor. J. Inst. Energy 1999, 72, 34–40. [Google Scholar]
- Blum, J.D.; Klaue, A.; Nezat, C.A.; Driscoll, C.T.; Johnson, C.E.; Siccama, T.G.; Eagar, C.; Fahey, T.J.; Likens, G.E. Mycorrhizal weathering of apatite as an important calcium source in base-poor forest ecosystems. Nature 2002, 417, 729–731. [Google Scholar] [CrossRef] [Green Version]
- Bullen, T.D.; Bailey, S.W. Identifying calcium sources at an acid deposition-impacted spruce forest: A strontium isotope, alkaline earth element multi-tracer approach. Biogeochemistry 2005, 74, 63–99. [Google Scholar] [CrossRef]
- Kennedy, M.J.; Hedin, L.O.; Derry, L.A. Decoupling of unpolluted temperate forests from rock nutrient sources revealed by natural 87Sr/86Sr and 84Sr tracer addition. Proc. Natl. Acad. Sci. USA 2002, 99, 9639. [Google Scholar] [CrossRef] [Green Version]
- Miller, E.K.; Blum, J.D.; Friedland, A.J. Determination of soil exchangeable-cation loss and weathering rates using Sr isotopes. Nature 1993, 362, 438–441. [Google Scholar] [CrossRef]
- Åberg, G.; Jacks, G.; Hamilton, P.J. Weathering rates and 87Sr/86Sr ratios: An isotopic approach. J. Hydrol. 1989, 109, 65–78. [Google Scholar] [CrossRef]
- Bailey, S.W.; Hornbeck, J.W.; Driscoll, C.T.; Gaudette, H.E. Calcium inputs and transport in a base-poor forest ecosystem as interpreted by Sr isotopes. Water Resour. Res. 1996, 32, 707–719. [Google Scholar] [CrossRef]
- Elias, R.W.; Hirao, Y.; Patterson, C.C. The circumvention of the natural biopurification of calcium along nutrient pathways by atmospheric inputs of industrial lead. Geochim. Cosmochim. Acta 1982, 46, 2561–2580. [Google Scholar] [CrossRef]
- Poszwa, A.; Dambrine, E.; Pollier, B.; Atteia, O. A comparison between Ca and Sr cycling in forest ecosystems. Plant Soil 2000, 225, 299–310. [Google Scholar] [CrossRef]
- Åberg, G.; Jacks, G.; Wickman, T.; Hamilton, P.J. Strontium isotopes in trees as an indicator for calcium availability. Catena 1990, 17, 1–11. [Google Scholar] [CrossRef]
- Schmitt, A.-D.; Chabaux, F.; Stille, P. The calcium riverine and hydrothermal isotopic fluxes and the oceanic calcium mass balance. Earth Planet. Sci. Lett. 2003, 213, 503–518. [Google Scholar] [CrossRef]
- Schmitt, A.-D.; Stille, P.; Vennemann, T. Variations of the 44Ca/40Ca ratio in seawater during the past 24 million years: Evidence from δ44Ca and δ18O values of Miocene phosphates. Geochim. Cosmochim. Acta 2003, 67, 2607–2614. [Google Scholar] [CrossRef]
- Capo, R.C.; Stewart, B.W.; Chadwick, O.A. Strontium isotopes as tracers of ecosystem processes: Theory and methods. Geoderma 1998, 82, 197–225. [Google Scholar] [CrossRef]
- Pearce, C.R.; Parkinson, I.J.; Gaillardet, J.; Charlier, B.L.A.; Mokadem, F.; Burton, K.W. Reassessing the stable (δ88/86Sr) and radiogenic (87Sr/86Sr) strontium isotopic composition of marine inputs. Geochim. Cosmochim. Acta 2015, 157, 125–146. [Google Scholar] [CrossRef]
- Pearce, C.R.; Parkinson, I.J.; Gaillardet, J.; Chetelat, B.; Burton, K.W. Characterising the stable (δ88/86Sr) and radiogenic (87Sr/86Sr) isotopic composition of strontium in rainwater. Chem. Geol. 2015, 409, 54–60. [Google Scholar] [CrossRef] [Green Version]
- Wiegand, B.A.; Chadwick, O.A.; Vitousek, P.M.; Wooden, J.L. Ca cycling and isotopic fluxes in forested ecosystems in Hawaii. Geophys. Res. Lett. 2005, 32. [Google Scholar] [CrossRef] [Green Version]
- Fantle, M.S.; DePaolo, D.J. Ca isotopes in carbonate sediment and pore fluid from ODP Site 807A: The Ca2+(aq)–calcite equilibrium fractionation factor and calcite recrystallization rates in Pleistocene sediments. Geochim. Cosmochim. Acta 2007, 71, 2524–2546. [Google Scholar] [CrossRef]
- Farkaš, J.; Buhl, D.; Blenkinsop, J.; Veizer, J. Evolution of the oceanic calcium cycle during the late Mesozoic: Evidence from δ44/40Ca of marine skeletal carbonates. Earth Planet. Sci. Lett. 2007, 253, 96–111. [Google Scholar] [CrossRef]
- Nägler, T.F.; Eisenhauer, A.; Müller, A.; Hemleben, C.; Kramers, J. The δ44Ca-temperature calibration on fossil and cultured Globigerinoides sacculifer: New tool for reconstruction of past sea surface temperatures. Geochem. Geophys. Geosyst. 2000, 1. [Google Scholar] [CrossRef] [Green Version]
- Skulan, J.; DePaolo, D.J.; Owens, T.L. Biological control of calcium isotopic abundances in the global calcium cycle. Geochim. Cosmochim. Acta 1997, 61, 2505–2510. [Google Scholar] [CrossRef]
- Zhu, P.; Macdougall, J.D. Calcium isotopes in the marine environment and the oceanic calcium cycle. Geochim. Cosmochim. Acta 1998, 62, 1691–1698. [Google Scholar] [CrossRef]
- Cenki-Tok, B.; Chabaux, F.; Lemarchand, D.; Schmitt, A.D.; Pierret, M.C.; Viville, D.; Bagard, M.L.; Stille, P. The impact of water–rock interaction and vegetation on calcium isotope fractionation in soil- and stream waters of a small, forested catchment (the Strengbach case). Geochim. Cosmochim. Acta 2009, 73, 2215–2228. [Google Scholar] [CrossRef]
- Cobert, F.; Schmitt, A.-D.; Bourgeade, P.; Labolle, F.; Badot, P.-M.; Chabaux, F.; Stille, P. Experimental identification of Ca isotopic fractionations in higher plants. Geochim. Cosmochim. Acta 2011, 75, 5467–5482. [Google Scholar] [CrossRef]
- Farkaš, J.; Déjeant, A.; Novák, M.; Jacobsen, S.B. Calcium isotope constraints on the uptake and sources of Ca2+ in a base-poor forest: A new concept of combining stable (δ44/42Ca) and radiogenic (εCa) signals. Geochim. Cosmochim. Acta 2011, 75, 7031–7046. [Google Scholar] [CrossRef]
- Han, G.; Song, Z.; Tang, Y.; Wu, Q.; Wang, Z. Ca and Sr isotope compositions of rainwater from Guiyang city, Southwest China: Implication for the sources of atmospheric aerosols and their seasonal variations. Atmos. Environ. 2019, 214, 116854. [Google Scholar] [CrossRef]
- Page, B.D.; Bullen, T.D.; Mitchell, M.J. Influences of calcium availability and tree species on Ca isotope fractionation in soil and vegetation. Biogeochemistry 2008, 88, 1–13. [Google Scholar] [CrossRef]
- Schmitt, A.-D.; Stille, P. The source of calcium in wet atmospheric deposits: Ca-Sr isotope evidence. Geochim. Cosmochim. Acta 2005, 69, 3463–3468. [Google Scholar] [CrossRef]
- Turchyn, A.V.; DePaolo, D.J. Calcium isotope evidence for suppression of carbonate dissolution in carbonate-bearing organic-rich sediments. Geochim. Cosmochim. Acta 2011, 75, 7081–7098. [Google Scholar] [CrossRef]
- Wiegand, B.A.; Schwendenmann, L. Determination of Sr and Ca sources in small tropical catchments (La Selva, Costa Rica)—A comparison of Sr and Ca isotopes. J. Hydrol. 2013, 488, 110–117. [Google Scholar] [CrossRef]
- van der Heijden, G.; Dambrine, E.; Pollier, B.; Zeller, B.; Ranger, J.; Legout, A. Mg and Ca uptake by roots in relation to depth and allocation to aboveground tissues: Results from an isotopic labeling study in a beech forest on base-poor soil. Biogeochemistry 2015, 122, 375–393. [Google Scholar] [CrossRef]
- Hindshaw, R.S.; Reynolds, B.C.; Wiederhold, J.G.; Kretzschmar, R.; Bourdon, B. Calcium isotopes in a proglacial weathering environment: Damma glacier, Switzerland. Geochim. Cosmochim. Acta 2011, 75, 106–118. [Google Scholar] [CrossRef]
- Holmden, C.; Bélanger, N. Calcium isotope fractionation in a boreal forest ecosystem. Geochim. Cosmochim. Acta 2006, 70, A261. [Google Scholar] [CrossRef]
- Holmden, C.; Bélanger, N. Ca isotope cycling in a forested ecosystem. Geochim. Cosmochim. Acta 2010, 74, 995–1015. [Google Scholar] [CrossRef] [Green Version]
- Fantle, M.S.; Tipper, E.T. Calcium isotopes in the global biogeochemical Ca cycle: Implications for development of a Ca isotope proxy. Earth Sci. Rev. 2014, 129, 148–177. [Google Scholar] [CrossRef]
- Zeng, J.; Han, G.; Wu, Q.; Tang, Y. Effects of agricultural alkaline substances on reducing the rainwater acidification: Insight from chemical compositions and calcium isotopes in a karst forests area. Agric. Ecosyst. Environ. 2020, 290, 106782. [Google Scholar] [CrossRef]
- Likens, G.E.; Driscoll, C.T.; Buso, D.C.; Siccama, T.G.; Johnson, C.E.; Lovett, G.M.; Fahey, T.J.; Reiners, W.A.; Ryan, D.F.; Martin, C.W.; et al. The biogeochemistry of calcium at Hubbard Brook. Biogeochemistry 1998, 41, 89–173. [Google Scholar] [CrossRef]
- Dijkstra, F.A. Calcium mineralization in the forest floor and surface soil beneath different tree species in the northeastern US. For. Ecol. Manag. 2003, 175, 185–194. [Google Scholar] [CrossRef]
- Dijkstra, F.A.; Van Breemen, N.; Jongmans, A.G.; Davies, G.R.; Likens, G.E. Calcium weathering in forested soils and the effect of different tree species. Biogeochemistry 2003, 62, 253–275. [Google Scholar] [CrossRef]
- Fujinuma, R.; Bockheim, J.; Balster, N. Base-cation cycling by individual tree species in old-growth forests of upper michigan, USA. Biogeochemistry 2005, 74, 357–376. [Google Scholar] [CrossRef]
- Zeng, J.; Han, G. Rainwater chemistry reveals air pollution in a karst forest: Temporal variations, source apportionment, and implications for the forest. Atmosphere 2020, 11, 1315. [Google Scholar] [CrossRef]
- Han, G.; Tang, Y.; Wu, Q. Hydrogeochemistry and dissolved inorganic carbon isotopic composition on karst groundwater in Maolan, southwest China. Environ. Earth Sci. 2010, 60, 893–899. [Google Scholar] [CrossRef]
- Han, G.; Li, F.; Tang, Y. Variations in soil organic carbon contents and isotopic compositions under different land uses in a typical karst area in Southwest China. Geochem. J. 2015, 49, 63–71. [Google Scholar] [CrossRef] [Green Version]
- Han, G.; Tang, Y.; Wu, Q.; Tan, Q. Chemical and strontium isotope characterization of rainwater in karst virgin forest, Southwest China. Atmos. Environ. 2010, 44, 174–181. [Google Scholar] [CrossRef]
- Zeng, J.; Han, G.; Yang, K. Assessment and sources of heavy metals in suspended particulate matter in a tropical catchment, northeast Thailand. J. Clean. Prod. 2020, 265, 121898. [Google Scholar] [CrossRef]
- Li, X.; Han, G. One-step chromatographic purification of K, Ca, and Sr from geological samples for high precision stable and radiogenic isotope analysis by MC-ICP-MS. J. Anal. At. Spectrom. 2021, 36, 676–684. [Google Scholar] [CrossRef]
- Wang, J.; Wang, L.; Wang, Y.; Tsang, D.C.W.; Yang, X.; Beiyuan, J.; Yin, M.; Xiao, T.; Jiang, Y.; Lin, W.; et al. Emerging risks of toxic metal(loid)s in soil-vegetables influenced by steel-making activities and isotopic source apportionment. Environ. Int. 2021, 146, 106207. [Google Scholar] [CrossRef] [PubMed]
- Heuser, A.; Eisenhauer, A.; Gussone, N.; Bock, B.; Hansen, B.T.; Nägler, T.F. Measurement of calcium isotopes (δ44Ca) using a multicollector TIMS technique. Int. J. Mass Spectrom. 2002, 220, 385–397. [Google Scholar] [CrossRef]
- De La Rocha, C.L.; DePaolo, D.J. Isotopic evidence for variations in the marine calcium cycle over the Cenozoic. Science 2000, 289, 1176–1178. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.; Han, G. Preliminary copper isotope study on particulate matter in Zhujiang River, southwest China: Application for source identification. Ecotoxicol. Environ. Saf. 2020, 198, 110663. [Google Scholar] [CrossRef]
- Liu, M.; Han, G.; Zhang, Q. Effects of agricultural abandonment on soil aggregation, soil organic carbon storage and stabilization: Results from observation in a small karst catchment, Southwest China. Agric. Ecosyst. Environ. 2020, 288, 106719. [Google Scholar] [CrossRef]
- Liu, M.; Han, G.; Li, X. Comparative analysis of soil nutrients under different land-use types in the Mun River basin of Northeast Thailand. J. Soils Sediments 2021. [Google Scholar] [CrossRef]
- Schmitt, A.-D.; Cobert, F.; Bourgeade, P.; Ertlen, D.; Labolle, F.; Gangloff, S.; Badot, P.-M.; Chabaux, F.; Stille, P. Calcium isotope fractionation during plant growth under a limited nutrient supply. Geochim. Cosmochim. Acta 2013, 110, 70–83. [Google Scholar] [CrossRef]
- Liu, J.; Han, G. Tracing riverine particulate black carbon sources in Xijiang River basin: Insight from stable isotopic composition and Bayesian mixing model. Water Res. 2021, 194, 116932. [Google Scholar] [CrossRef]
- Liu, J.; Han, G. Major ions and δ34SSO4 in Jiulongjiang River water: Investigating the relationships between natural chemical weathering and human perturbations. Sci. Total. Environ. 2020, 724, 138208. [Google Scholar] [CrossRef]
- Hu, Z.-L.; Pan, G.-X.; Li, L.-Q.; Du, Y.-X.; Wang, X.-Z. Changes in pools and heterogeneity of soil organic carbon, nitrogen and phosphorus under different vegetation types in Karst mountainous area of central Guizhou Province, China. Acta Ecol. Sin. 2009, 29, 4187–4195, (In Chinese with English abstract). [Google Scholar]
- Schmitt, A.-D.; Gangloff, S.; Labolle, F.; Chabaux, F.; Stille, P. Calcium biogeochemical cycle at the beech tree-soil solution interface from the Strengbach CZO (NE France): Insights from stable Ca and radiogenic Sr isotopes. Geochim. Cosmochim. Acta 2017, 213, 91–109. [Google Scholar] [CrossRef]
Figure 1.
The study area and the soil sampling location: (a) location of MNNRP, (b) elevation of the study area, (c) lithology of the study area, and (d) land use of the study area. LBP, farmland soil site; LBM, burnt grassland soil site; LBS, shrubland soil site; LBF, virgin forest soil site; LBG, grassland soil site; R, rainwater site.
Figure 1.
The study area and the soil sampling location: (a) location of MNNRP, (b) elevation of the study area, (c) lithology of the study area, and (d) land use of the study area. LBP, farmland soil site; LBM, burnt grassland soil site; LBS, shrubland soil site; LBF, virgin forest soil site; LBG, grassland soil site; R, rainwater site.

Figure 2.
Ca isotope pool of the forest ecosystem in MNNRP.

Figure 3.
δ44/40Ca (‰) vs. 1/Ca of rainwater samples in the MNNRP (this study) and other studies [27,32,36].
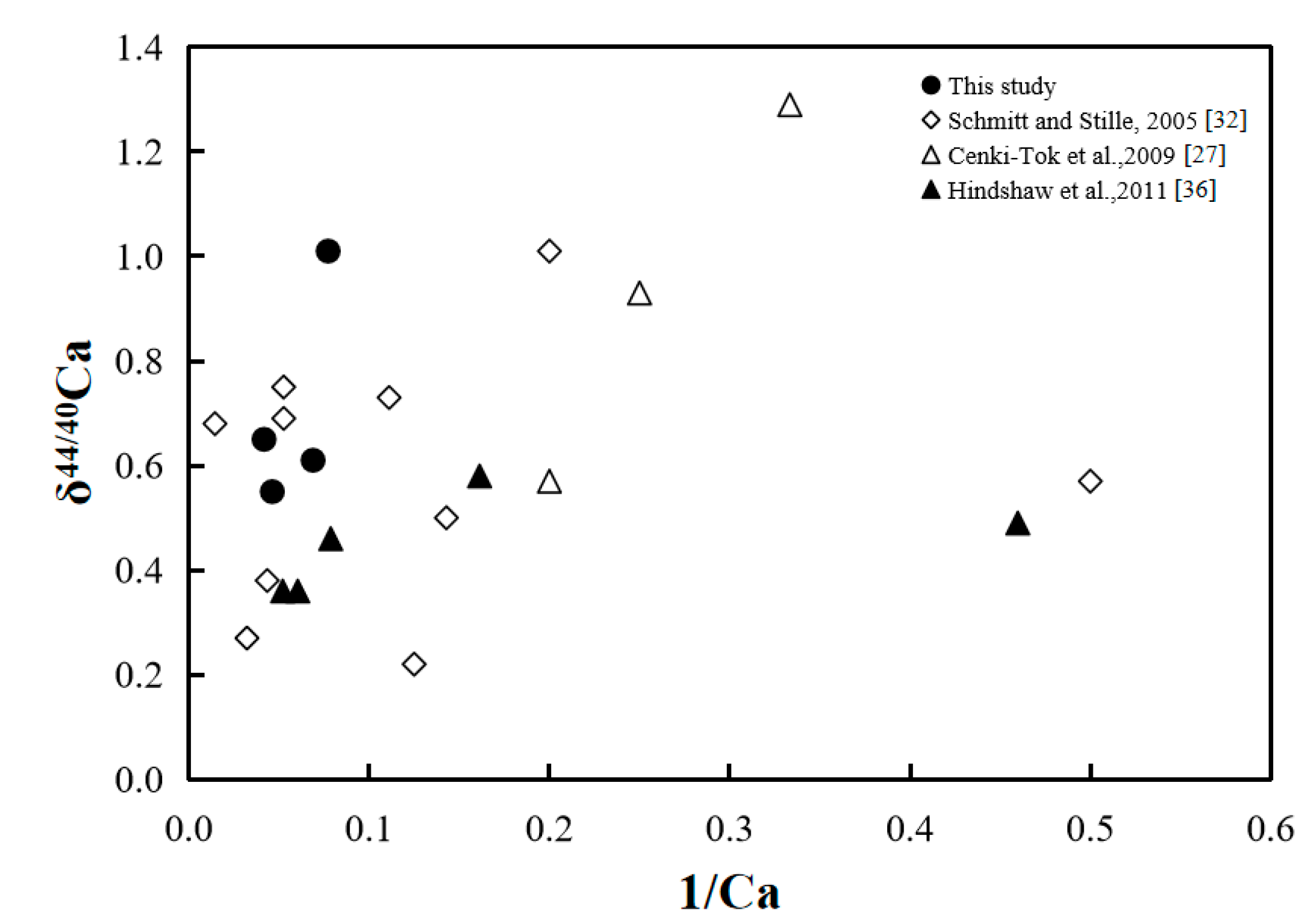
Figure 4.
The δ44/40Ca (‰) values from soil samples collected at different depths. Error bars are 2SD.
Figure 4.
The δ44/40Ca (‰) values from soil samples collected at different depths. Error bars are 2SD.
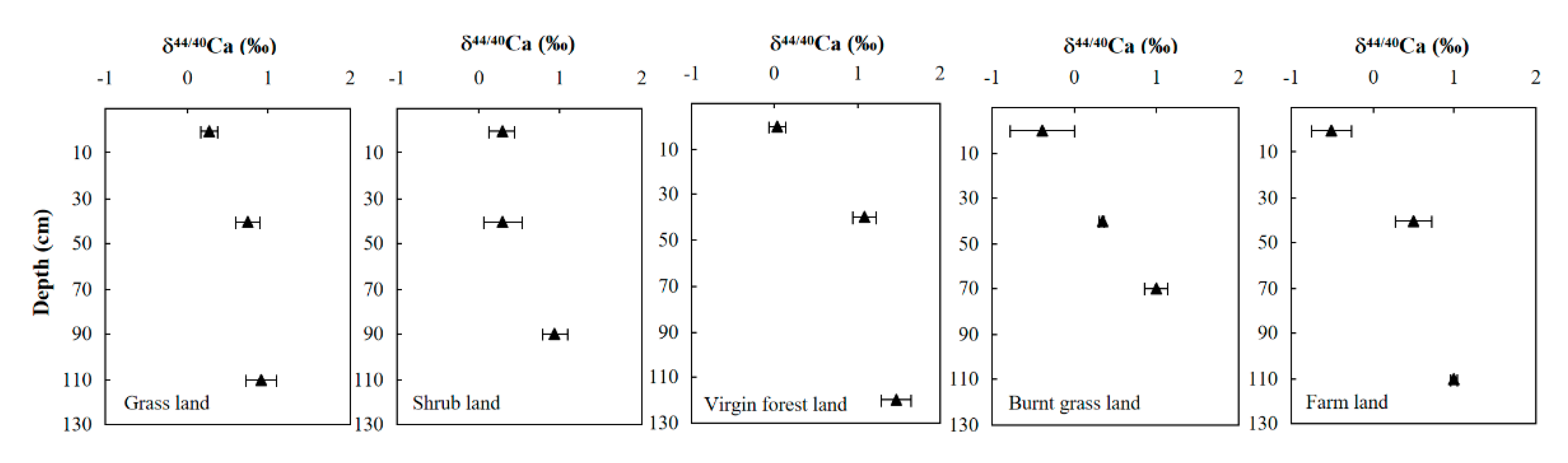
Figure 5.
The δ44/40Ca of surface soil under different vegetation covers in MNNRP. Error bars are 2SD.
Figure 5.
The δ44/40Ca of surface soil under different vegetation covers in MNNRP. Error bars are 2SD.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Ca and Mg concentrations and Ca isotopic composition of rainwater, groundwater, soil, vegetation, and bedrock samples in MNNRP.
Table 1.
Ca and Mg concentrations and Ca isotopic composition of rainwater, groundwater, soil, vegetation, and bedrock samples in MNNRP.
| Sample Number | Date (Year-Month-Day) | Ca2+ | Mg2+ | δ44/40Ca | 2SD | n |
|---|---|---|---|---|---|---|
| μmol/L | μmol/L | ‰ | ‰ | |||
| Rainwater | ||||||
| LB-32 | 2008-6-08 | 14.5 | 3.2 | 0.61 | 0.29 | 3 |
| LB-41 | 2008-7-24 | 21.6 | 2.7 | 0.55 | 0.24 | 3 |
| LB-44 | 2008-8-18 | 24.0 | 2.9 | 0.65 | 0.23 | 2 |
| LB-47 | 2008-9-24 | 12.9 | 0.9 | 1.01 | 0.15 | 3 |
| Groundwater | ||||||
| 2 | 2007-7-23 | 1163 | 648 | 0.40 | 0.14 | 2 |
| 3 | 2007-7-23 | 1309 | 501 | 0.52 | 0.04 | 3 |
| 7 | 2007-7-23 | 1312 | 373 | 0.60 | - | 1 |
| 11 | 2007-7-23 | 1452 | 114 | 0.63 | - | 1 |
| 13 | 2007-7-23 | 1728 | 289 | 0.72 | - | 1 |
| 16 | 2007-7-24 | 1759 | 126 | 0.36 | 0.13 | 2 |
| 20 | 2007-7-24 | 2538 | 394 | 0.43 | 0.01 | 2 |
| 24 | 2007-7-24 | 1334 | 1353 | 1.08 | 0.21 | 2 |
| 27 | 2007-7-27 | 1394 | 609 | 0.83 | - | 1 |
| Bedrock | mmol/kg | mmol/kg | ||||
| Dolomite | 6066 | 5872 | 0.36 | 0.09 | 3 | |
| Soil | depth (cm) | |||||
| Grassland | 0 | 20 | 137 | 0.28 | 0.10 | 2 |
| 40 | 56 | 135 | 0.75 | 0.14 | 3 | |
| 110 | 2275 | 2501 | 0.91 | 0.18 | 3 | |
| Shrubland | 0 | 34 | 43 | 0.29 | 0.15 | 3 |
| 40 | 159 | 166 | 0.30 | 0.24 | 3 | |
| 90 | 23 | 74 | 0.94 | 0.15 | 2 | |
| Farmland | 0 | 106 | 464 | −0.51 | 0.25 | 3 |
| 40 | 13 | 16 | 0.50 | 0.22 | 3 | |
| 110 | 21 | 25 | 1.00 | 0.04 | 2 | |
| Burnt grassland | 0 | 31 | 97 | −0.38 | 0.19 | 2 |
| 40 | 159 | 25 | 0.34 | 0.03 | 3 | |
| 70 | 16 | 25 | 1.00 | 0.15 | 2 | |
| Virgin forests | 0 | 60 | 124 | 0.04 | 0.20 | 3 |
| 40 | 11 | 36 | 1.08 | 0.21 | 2 | |
| 120 | 21 | 155 | 1.48 | 0.23 | 3 | |
| Forest tree | ||||||
| Nandian domestica | 231 | 101 | −0.26 | 0.18 | 2 | |
| Folium platycaryae | 602 | 194 | −0.22 | 0.08 | 2 | |
| Handliodendron bodinier | 672 | 234 | −0.26 | - | 1 | |
| Crop | ||||||
| Soybean | 47 | 233 | −1.44 | 0.14 | 3 | |
| Corn | 51 | 249 | −1.35 | 0.20 | 2 |
Note: the δ44/40Ca values and uncertainties are based on two to three mass spectrometric measurements; the rainwater samples, groundwater samples, and soil samples were derived from our previous work [46,47,48]; 2SD, two times the standard deviation among the δ44/40Ca values; n, number of measurements for each sample.
Table 2.
The δ44/40Ca values and Mg/Ca ratios of surface soil in MNNRP.
| Shrubland | Virgin Forests | Grassland | Burnt Grassland | Farmland | |
|---|---|---|---|---|---|
| Mg/Ca | 1.27 | 2.08 | 2.45 | 3.15 | 4.37 |
| δ44/40Ca | 0.29 | 0.04 | 0.28 | −0.38 | −0.51 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Han, G.; Eisenhauer, A.; Zeng, J.; Liu, M. Calcium Biogeochemical Cycle in a Typical Karst Forest: Evidence from Calcium Isotope Compositions. Forests 2021, 12, 666. https://doi.org/10.3390/f12060666
AMA Style
Han G, Eisenhauer A, Zeng J, Liu M. Calcium Biogeochemical Cycle in a Typical Karst Forest: Evidence from Calcium Isotope Compositions. Forests. 2021; 12(6):666. https://doi.org/10.3390/f12060666
Chicago/Turabian StyleHan, Guilin, Anton Eisenhauer, Jie Zeng, and Man Liu. 2021. "Calcium Biogeochemical Cycle in a Typical Karst Forest: Evidence from Calcium Isotope Compositions" Forests 12, no. 6: 666. https://doi.org/10.3390/f12060666
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.