
Temperature and Precipitation Significantly Affect Resource Allocation in Dioecious Plants: A Meta-Analysis

1
The Key Laboratory for Silviculture and Conservation of Ministry of Education, Key Laboratory for Silviculture and Forest Ecosystem of State Forestry and Grassland Administration, Research Center for Urban Forestry, Beijing Forestry University, Beijing 100083, China
2
Department of Soil, Plant and Food Sciences University of Bari Aldo Moro, 70126 Bari, Italy
*
Author to whom correspondence should be addressed.
Forests 2024, 15(4), 720; https://doi.org/10.3390/f15040720
Submission received: 29 March 2024
/
Revised: 12 April 2024
/
Accepted: 13 April 2024
/
Published: 19 April 2024
(This article belongs to the Special Issue Urban Forestry and Sustainable Cities)
Abstract
:Dioecious plants are often used in landscaping because of sex differences in individual appearance, resistance and ornamental value. Although a large number of studies have investigated the overall differences in resource allocation between different sexes in dioecious plants, the effects of environmental factors such as temperature and precipitation on resource allocation in sex-dimorphic plants are not fully understood. In order to explore the influencing factors, 39 works of literature on dioecious plant resource allocation published in English databases from 1992 to 2023 were selected, and the biomass data of dioecious plant stems, leaves, flowers and fruits from these pieces of literature were extracted. A total of 545 independent experimental groups were obtained, which were divided into four groups for analysis, and the data were analyzed using METAWIN 2.1 software. Four sets of data were used to quantitatively study the effects of different temperatures, precipitations and life forms on the resource allocation of dioecious plants of different sexes in large-scale space. The results showed that female plants invested more resources in reproductive growth and less resources in vegetative growth. In terms of total biomass, the average biomass of female plants was 3.09% higher than that of male plants, indicating that female plants reduced nutrient investment to compensate for reproductive investment in the process of resource allocation. Temperature and precipitation significantly affect the adaptability of male and female plants to environmental changes and the cooperative relationships among the stressed components. The vegetative biomass investment of female plants showed a positive correlation with the increase in temperature, while the reproductive biomass showed a negative correlation with the decrease. The average annual precipitation had little effect on the vegetative biomass of dioecious plants, but had a significant effect on reproductive biomass. The study of this trade-off relationship is helpful in revealing the relationship between vegetative growth and reproductive growth of plants, exploring the countermeasures of plant life history, and providing a scientific basis for urban landscaping and urban forest management.
1. Introduction
According to the theory of plant life history, plants can obtain limited resources through physiological processes such as photosynthesis, and allocate resources among various organs to meet the needs of their different growth and reproductive stages [1]. All the growth and developmental processes of plants including defense and reproduction are accompanied by competition for resources [2]. The response measures of resource allocation differ among plants. Correspondingly, different allocation measures will diversify the growth process of plants, and the resource allocation mode, whether the allocation is reasonable or not, will also have an impact on plant growth and even plant life [3,4,5]. Under disparate external conditions, growth and developmental stages, plants of different sexes will adjust the allocation of resources appropriately to promote enhanced development [6,7].
A variety of resources are invested in reproductive activities by individual plants of different sexes, and those of female individuals are usually more numerous than those of males. This contrast in reproductive investment leads to the separation of plant sex functions [8,9]. Therefore, it is assumed that a sort of trade-off relationship exists between different life-history traits of dioecious plants in the process of resource allocation [10,11]. There are differences in survival rate, functional traits, physiological characteristics and vegetative growth of individual plants of differing sexes [12]. Contrary to the supporters of the trade-off theory, some scholars do not support the existence of a trade-off relationship between plant life history traits, arguing that although the amount of resources devoted to the reproductive process of female individuals is much higher than that of male individuals, they still have comparable or even greater vegetative growth [13,14]. Wang Lifen et al. (2020) studied the sex difference in reproductive consumption of rhamnus species in Northeast China. Females did not reduce the resource input for vegetative growth due to higher reproductive resource input, but compensated for their higher reproductive resource input through higher photosynthetic capacity, indicating that there was a sex difference in the ability of female females to obtain resources [15]. However, it was also believed that the vegetative growth of female individuals was significantly greater than that of male individuals [16]. Some scholars believe that due to higher reproductive pressure, female individuals have gradually formed a series of potential compensation mechanisms in the evolutionary process to cope with higher reproductive investment, thus improving the ability of plants to obtain available resources from the external environment [17,18]. These different forms of compensation mechanisms affect the total amount of available resources obtained by individuals of different sexes in dioecious plants, thus masking the trade-off between resource allocation and vegetative growth [19]. A trade-off is observed between male and female functions when a plant increases the allocation of one sex and the input of the other sex inevitably decreases; this trade-off is also consistent with relevant theories of resource allocation [20,21,22]. The actual function of this feedback mechanism has a certain impact on the selection and development of the plant mating system [23].
The characteristic of plant resource allocation for reproductive growth and vegetative growth is one of the core contents of plant life history strategies. Dioecious plants are important model species for studying plant resource allocation patterns due to their independence in sex allocation. Meanwhile, dioecious plants, as an important part of the ecosystem, play a pivotal role in maintaining and protecting biodiversity [24]. Current research mainly focuses on sex determination, reproductive cost and differences in response to the environmental factors of a dioecious plant, due to a lack of systematic inquiry into different varieties and living conditions of such plants [25].
This paper aims to clarify the role of dioecious plants in resource allocation under different conditions. Based on the results of literature studies on dioecious plants since 1992, the data of effective biomass allocation in this paper were extracted, and the specific effects of different life types, temperature and precipitation on resource allocation of dioecious plants were analyzed using the meta-analysis method. The biomass distribution structure of organs of dioecious plants with different temperatures, precipitations and different plant life types was analyzed, and the relationship between vegetative growth and reproductive growth of dioecious plants was deduced. Based on this study, we can analyze the ability of male and female plants to adapt to environmental stress and measure the cooperative relationship between various components in this situation. It is convenient to study the functional traits, physiological characteristics and vegetative growth of dioecious plants of different genders, which is conducive to providing a scientific basis for urban landscaping and forestry construction management.
2. Materials and Methods
2.1. Literature Retrieval
The literature on dioecy resource allocation was published early in 1992; therefore, our study covers the period from 1992 to 2023. The Chinese literature database employed for the meta-analysis was derived from China HowNet (https://www.cnki.net, accessed on 1 March 2023), VIP (https://cqvip.com, accessed on 3 March 2023), Wanfang (https://www.wanfangdata.com.cn/index.html, accessed on 3 March, 2023), Baidu Academic (https://xueshu.baidu.com, accessed on 4 March 2023), Google Academic (https://xs.scqylaw.com, accessed on 10 March 2023), among others, and was searched using the terms “dioecious plants/resource allocation/biomass allocation”. The English database was extracted from Web of Science (Thompson-ISI, Philadelphia, PA, USA, http://apps.webofknowledge.com, accessed on 15 March 2023) and was searched using the keywords “dioecious plants/resource allocation/biomass allocation”. Among the types of retrieved data are scientific publications, books, patents, conference papers or abstracts and dissertations.
The process of data extraction is as follows (Figure 1): read all the pieces of literature found, extract the biomass data of each organ of dioecy in the literature, eliminate the biomass data without organs and the incomplete biomass data of organs in the literature, retain the documents that meet the requirements, and carry out the unified unit conversion of the data in the literature. A database of valid data was established, and subgroup analysis was conducted according to different temperatures, different life types and different rainfall to clarify the effect of sex on resource allocation of dioecious plants.
2.2. Inclusion and Exclusion Criteria
According to the actual research on dioecious resource allocation technology at this stage, the criteria for data screening were obtained. It is considered a valid document if it meets the following criteria: (1) The object of research of the document is dioecious plants; (2) The literature mainly focuses on the research/experiment of the distribution of dioecious plant resources according to male–female sex. The literature must contain actual research data, including the average and standard deviations of vegetative biomass (leaves and branches) and reproductive biomass (male-flower, female-flower+fruit), and be described using words, tables or images.
2.3. Data Extraction
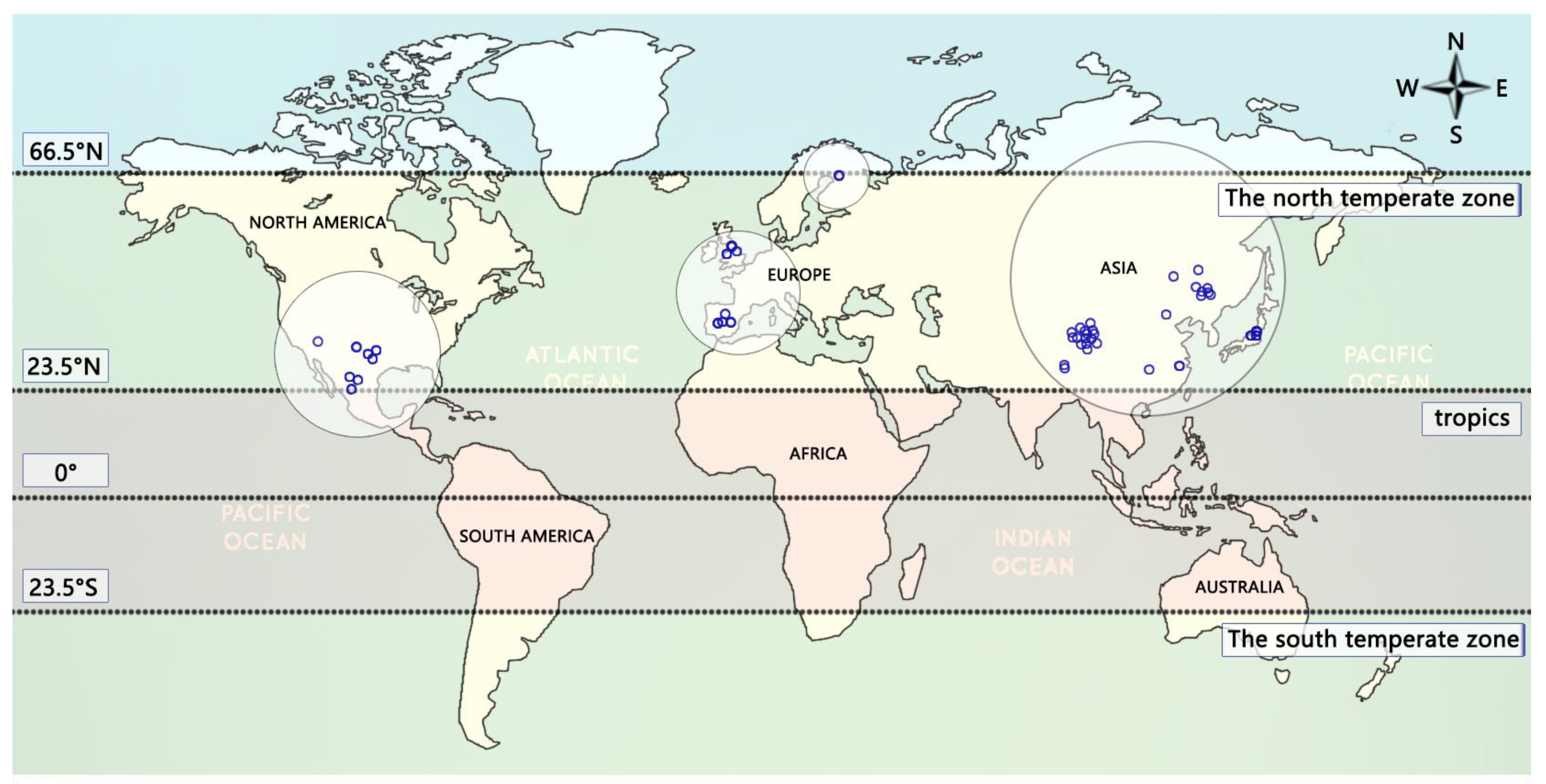
Data extraction and database establishment were carried out for the selected literature. The data mainly extracted the research results of dioecious plant resource allocation in each experimental group, selecting dioecious plants with different annual average temperatures, annual average precipitation (i.e., rainfall) and life forms as reference, and using the resource allocation results of female and male plants as the extraction amount. The screening criteria for our research included: (1) selecting only dioecious plants (either herbs, shrubs and woody plants); (2) an independent and complete set of experimental contents in a document taken as a group of independent samples for experimental data extraction; and (3) independent experiments of any group of treatment methods with more than two documents in scientific publications in order to be considered for the statistical analysis process. According to the screening criteria, a total of 131 pieces of literature were found. After 40 duplicate literature pieces were removed, 91 literature pieces were read in detail, among which 61 did not meet the screening criteria. The meta of biomass, temperature, precipitation and life form in the meta-analysis are from supplementary references. Finally, 39 usable literature were entered (Table 1); the biomass of each organ of male and female plants was extracted from the literature, and a total of 545 sets of independent sample data were collected (Figure 2). There were 91 groups of stem biomass, 107 groups of leaf biomass, 100 groups of flower biomass, 132 groups of vegetative biomass and 115 groups of reproductive biomass. Most studies that met these criteria were conducted in the Northern temperate zone (23.5–66.5 N) with data from the same biome, reducing differences in temperature, rainfall and plant responses; this reduces excessive data variation. These were conducted in Asia (67%), some in North America (20%) and Europe (13%) (Figure 3).
2.4. Meta-Analysis
The literature data extracted in this study included the mean (m), standard deviation (SDs) and sample size (n) of the biomass of each organ of dioecy. When the data are not directly tagged in the paper data graph, the data of each point in the map were read using the software GetData Graph Digitizer (http://getdata-graph-digitizer.com/registration.php, Developers in the United States, accessed on 1 June 2023). Standard errors (SEs), when provided, were converted using the following formula:
When SDs and SEs were not provided, 10% of the mean value was selected as SD by default [63]. The two most common effect value indicators for the application of integrated analysis in ecology are response ratio (LnR) and hedges’ D value. According to the statistical description of Hedges et al. [64], the response ratio was selected as the effect value in this study:
The variance (V) corresponding to the effect value is as follows:
where m1 and m2, sd1 and sd2, n1 and n2 are the mean, standard deviation and sample number of the experimental group and control group, respectively.
METAWIN 2.1 (www.metawinsoft.com, Developers in the United States, accessed on 12 June 2023) software was employed to calculate the 95% confidence intervals (CIs) of each average effect value R++ using the random effect model (REM). If all the 95% CIs were >0, the resource allocation of female plants was considered greater than that of male plants. If the CI was <0, then the male plant resource allocation was greater than that of the female plants. A value of 0 indicated no difference between the male plant resource allocation and the female plant resource allocation [65].
2.5. Heterogeneity Test
The results of the heterogeneity test can be used to select the fixed-effect model or random-effects model. By using Q-statistics to construct the heterogeneity test, when pQ < 0.05, the random-effects model was used; on the contrary, the fixed-effect model was applied [66].
2.6. Publication Bias Test
Publication bias is a common phenomenon of small sample data effects. Small samples usually have a greater processing effect than large samples, which will produce publication bias [67]. In this study, the histogram with a normal distribution curve was used to test publication bias.
2.7. Subgroup Analysis
According to the different temperatures, precipitations and life forms of dioecious plants in the literature, the biomass of the four groups of organs was divided into three subgroups. Subgroup analysis was used to find more detailed sources of heterogeneity [68].
2.8. Data Processing
Microsoft Excel 2016 data analysis software was used to process data and draw charts; METAWIN 2.1 was employed for meta-analysis [69]; and SigmaPlot 14.0 (https://systat-sigmaplot.com, Developers in the United States, accessed on 25 June 2023) was employed to draw frequency distribution histograms and for normal fitting.
3. Results
3.1. Heterogeneity and Publication Bias Test
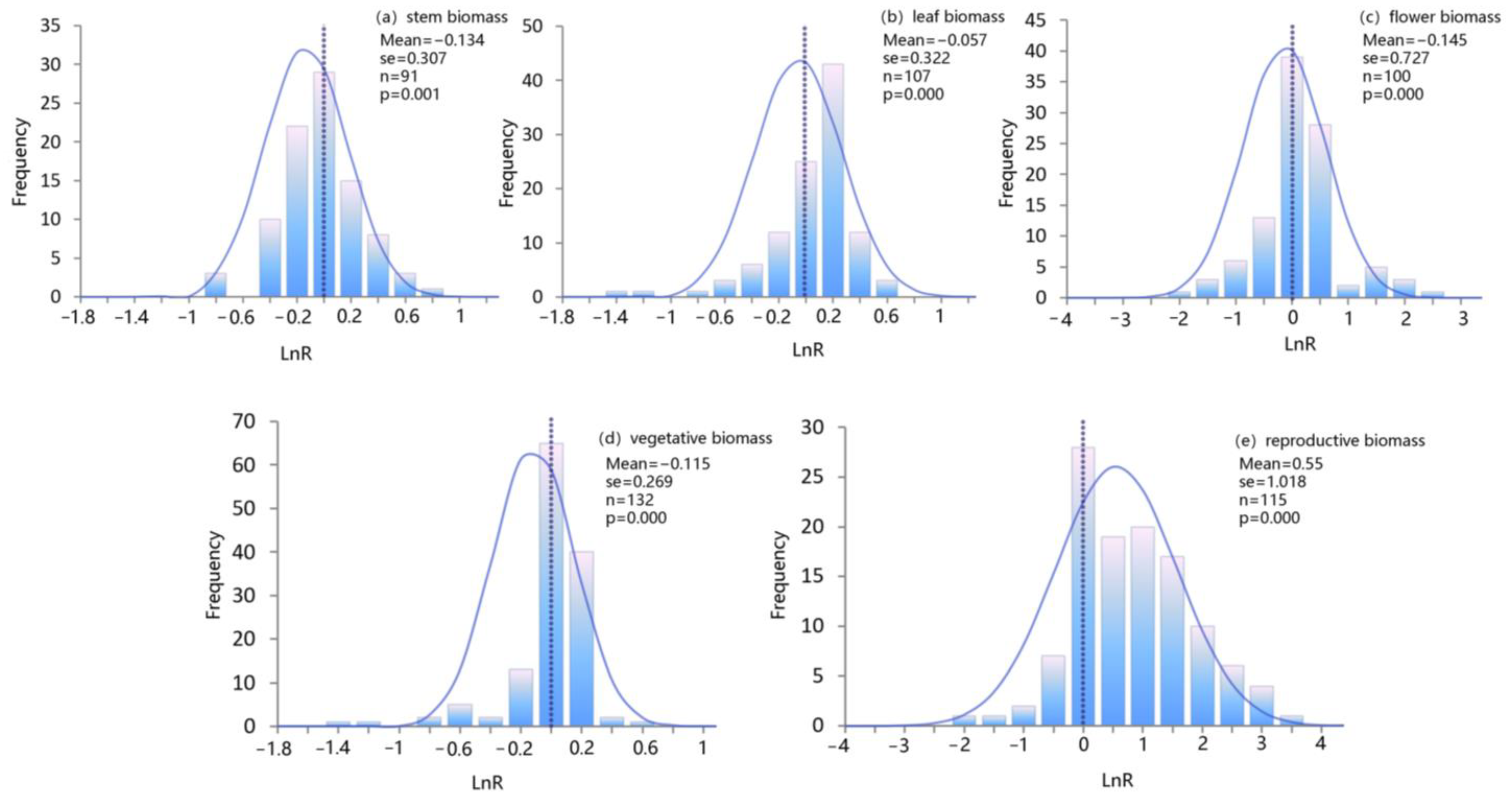
The chi-square test was used to examine the heterogeneity of the biomass ratio of organs of 454 groups of female and male plants extracted from the literature (Table 2). The results showed that the heterogeneity test of biomass proportion in the organs of female and male plants was extremely significant (pQ < 0.05). Therefore, the REM was adopted. The results of the publication bias test revealed no bias between different organ biomass ratio combinations for female and male plants (pB > 0.05).
To verify whether the responses of different organ biomass ratios of female and male dioecious plants follow a normal distribution, a Gaussian (normal) distribution was used for fitting (Figure 4). The results showed that the responses of different organ biomass ratios of female and male plants complied with normal distribution (p < 0.001).
3.2. Overall Response to Dioecious Plant Resource Allocation
According to the analysis of the experimental results, some differences were observed in the distribution of dioecious plant resources, and the reproductive biomass of female individuals was significantly greater than that of male individuals (p < 0.001). Figure 5 shows that the overall average biomass of female plants is 3.09% greater than that of male plants. For the reproductive and vegetative biomass of dioecious plants, the increase in the rate of reproductive biomass of female plants was positively correlated, at 73.66%; the resources invested were significantly more numerous than those of male plants. A negative correlation was found between the rate of increase of resource biomass invested in vegetative growth. The rate of increase of vegetative biomass was −9.67%, and the resources invested were significantly fewer than those of the male plants.
The results of specific organ biomass distribution of male and female plants showed that the increase in the rate of biomass in male plants was higher than that in female plants; namely, the rate increase of plant stems was −14.42%, of plant leaf biomass −2.97%, and of plant flower biomass −14.19%. The rate increase of organ biomass was consistent with the results of the rate increase of vegetative biomass and reproductive biomass. Due to the small sample size of root biomass in the experimental data, root biomass was not compared separately. The percentage of root biomass was included in the proportion of nutrient biomass in the experiment with root biomass. Since the male plant had no fruit, the reproductive biomass of the male plant was the flower biomass, and the reproductive biomass of the female plant was the total biomass of flowers and fruits.
3.3. Subgroup Analysis of Dioecious Plant Resource Allocation
3.3.1. Effect of Different Temperatures on Resource Allocation of Dioecious Plants
The data sources included in this study involve three temperature gradients. As depicted in Figure 6, differences in the allocation of resources can be observed in the organs of dioecious plants. The CI of the average reproductive biomass of female individuals did not include 0 and was >0, indicating that the average reproductive biomass of female plants is greater than that of male plants; the promotion rate was 77.59% (p < 0.008). However, the CIs of the average flower biomass, vegetative biomass and stem biomass did not include 0 and were <0, indicating that the average biomass of male plants is greater than that of female plants at different temperatures; the average rates of increase were −14.8%, −8.02% and −14.26%, respectively (p < 0.001).
Concerning the organ biomass of dioecious plants, the reproductive biomass of female plants was positively correlated at three different temperature gradients; the promotion rate was the highest when the temperature ranged from 0 to 10 °C. A significant positive correlation between flower biomass and stem biomass was found from 10 to 15 °C, but no significant difference was observed between vegetative biomass and stem biomass; both were negatively correlated. When the temperature was >15 °C, the leaf biomass of female plants showed a significant positive correlation.
3.3.2. Influence of Precipitation on Dioecious Plant Resource Allocation
Figure 7 illustrates differences in the allocation of resources in the organs of dioecious plants under various rainfall conditions. The CI of the average reproductive biomass of female individuals did not include 0 and was >0, indicating that the average reproductive biomass of female plants is greater than that of male plants; the promotion rate was 77.36% (p < 0.001). However, the CIs of the average flower biomass, vegetative biomass and stem biomass did not include 0 and were <0, indicating that the average male plant biomass is greater than the average female plant reproductive biomass under different rainfall conditions; the average rates of increase were −14.76%, −8.01% and −14.24% (p < 0.001), respectively.
The reproductive biomass of female plants was positively correlated with the three rainfall gradients, and the rate of increase was the highest when the average annual rainfall recorded was 500 to 800 mm. Flower biomass was positively correlated only when the average annual rainfall was <500 mm. However, there was no significant difference in nutrient biomass and stem biomass among the three precipitation gradients, and a significant negative correlation was observed. The leaf biomass of female plants showed a significant positive correlation when the annual rainfall recorded was >800 mm and a significant negative correlation when it was <800 mm.
3.3.3. Influence of Different Life Forms on the Resource Allocation of Dioecious Plants
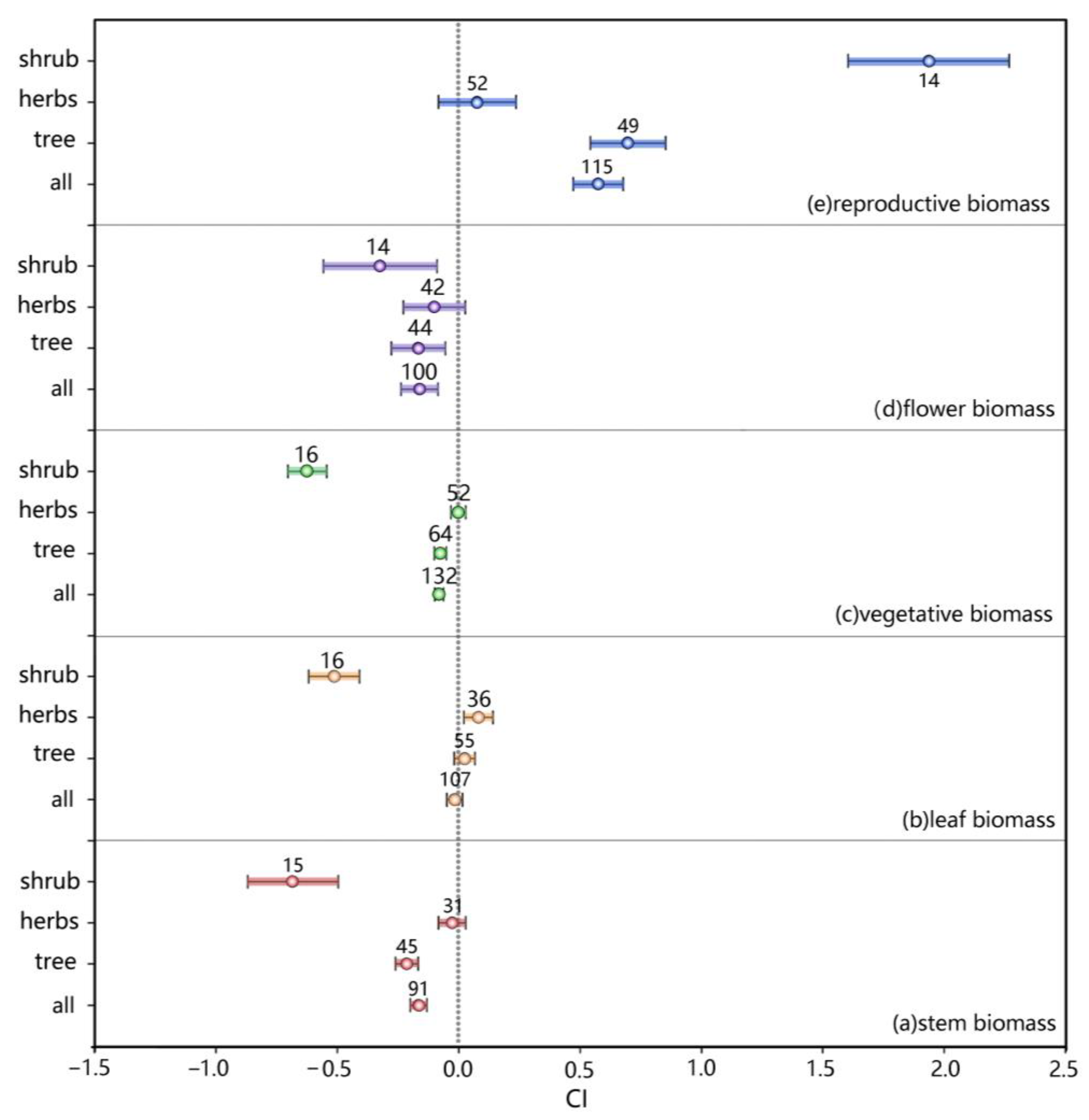
The comparison and analysis of data according to the life forms of plants revealed differences in the organ biomass distribution of dioecious individuals (p < 0.003) (Figure 8). The CI of the average reproductive biomass of female plants with different life forms did not include 0 and was >0, indicating that the average reproductive biomass of female plants is greater than that of male plants; the promotion rate was 77.61% (p < 0.001). However, the CIs of flower biomass, vegetative biomass and stem biomass were <0, which indicated that the average biomass of male plants is greater than that of female plants at different temperatures; the average rates of increase were −14.95%, −7.8% and −15.24%, respectively (p < 0.001).
A positive correlation was found in the reproductive biomass of different life forms of plants. When the life form was a shrub, the reproductive biomass of female plants was the highest. The differences in flower biomass, nutrient biomass and stem biomass were all negatively correlated, and the promotion rate of shrubs was the lowest. No noticeable difference was found between herbs and woody plants. Except for shrubs, leaf biomass was positively correlated, but the promotion rate was low.
4. Discussion
4.1. Characteristics of Resource Allocation of Dioecious Plants
The sex difference of dioecious plants enables them to meet the energy requirements for reproduction; hence, they are considered an evolved group [70]. However, dioecious plants are highly sensitive to changes in external environmental factors, and as a result, their reproductive pattern is closely related to specific microenvironmental factors [71]. The biomass allocation of dioecious plants is easily disturbed by external environmental factors as well. Consequently, it is possible to analyze the ability of plants to adapt to environmental changes and measure the cooperative relationship between various components when their biomass allocation structure is subject to stress [72]. EHRLEN J, SAKAI A K and other authors believe there is no significant difference between the nutritional growth of male and female individuals, and that even the nutritional growth of female individuals is significantly higher than that of male individuals. According to Zhang Dayong and Li Yanlian, a trade-off relationship exists between different life history traits in the process of resource allocation. The results of this work are similar to those of Zhang Dayong, where sex differences were observed in the biomass of different organs of dioecious plants [73]. At the plant level, the rate increase of the flower biomass and stem biomass of female plants was −14.19% and −14.42%, respectively, which was significantly lower than that of male plants. The rate increase of the leaf biomass of female plants was −2.97%, which was not significantly different from that of male plants. The total amount of environmental resources that plants can exploit is limited and there is little difference between male and female plants. The rate increase of reproductive biomass of female plants was 73.66%, while the rate increase of nutritional biomass was −9.67%, indicating that individuals with high reproductive investment reduce their own nutritional growth [74]. Compared to males, female individuals must undergo two processes of flowering and fruiting, and the reproductive cost is relatively high. The amount of available resources that female individuals can allocate to vegetative growth is lesser than that of males [75]. The analysis of reproductive and vegetative growth patterns of plants of different sexes shows that there is a significant difference between female and male plants. Female plants invest more resources in reproductive growth than male plants, but invest fewer resources in vegetative growth; in other words, sex has a significant impact on the resource allocation pattern of dioecious plants [76].
4.2. Temperature
Most physiological and biochemical changes of plant growth occur in suitable temperature environments. If the temperature is lower or higher than the suitable temperature, it will affect plant growth; a rising temperature induces plants to absorb water and nutrients at a faster pace. When the temperature is too high, it reduces the area absorbed by the roots, while the plants at suitable temperatures can accumulate more nutrients [77]. At the same time, temperature and photoperiod are the key factors affecting the development of flower buds. A temperature increase will lead to more bud maturation in male plants than in female plants [78]. Moreover, temperature treatment can increase the activities of catalase, peroxidase and superoxide dismutase in female flower buds, while the increase of malondialdehyde content in male flower buds is significantly delayed [79]. All the physiological changes and biochemical reactions of plants need to be carried out under suitable temperature conditions. In the temperature range where dioecious plants grow, different temperature values exert different effects. The value of the stem biomass of male plants was higher than that of female plants at different temperatures, which indicated that stem biomass was less affected by temperature. The leaf biomass of female plants was greater than that of male plants when the annual average temperature was higher than 15 °C. Leaf biomass is greatly affected by temperature, and the increase of leaf biomass in female plants is directly proportional to temperature in a suitable temperature range. The average flower biomass value of female plants was significantly lower than that of male plants, but significantly higher than that of male plants from 10 to 15 °C, indicating that the flower biomass of dioecious plants is affected by temperature. The results of our study show that some differences exist in the resource allocation of dioecious plants under different temperature conditions, and that there is a positive correlation between the investment in vegetative biomass of female plants and the decrease in reproductive biomass with an increase in temperature [80]. In accordance with the overall characteristics of dioecious plant resource allocation, the reproductive biomass of female plants is greater than that of male plants under different temperature conditions, and the nutritional biomass value is lower than that of male plants.
4.3. Precipitation
Precipitation influences the growth of plants by affecting soil water content, transpiration and air humidity. Water scarcity affects the division and elongation of plant cells and the growth of plants [81]. Water use efficiency is an important indicator of the plant’s water physiological characteristics [82]. The higher the water use efficiency, the more fully plants use water resources, and the stronger the ability of plants to cope with drought stress [48]. Numerous research results reveal that once male and female individuals are subjected to water stress, significant differences can be observed in physiological indices, such as dry matter accumulation, net photosynthetic rate, transpiration rate and water use efficiency, as well as in some morphological indices. Female plants in wet habitats are stronger than those in dry habitats in their growth, reproduction, seed number, germination and embryo formation potential [83]. The physiological contrasts between different sexes depend on the environment, and sex-specific water use efficiency plays a decisive role in explaining the interaction between different sexes in growth and ecology [84]. When there is an overabundance of water, the root system remains underdeveloped and the stems and leaves are thin and young, which affects yield. The growth of most plants is directly proportional to water supply, and growth is limited by extremely low or high levels of soil humidity [85]. Under different conditions of precipitation, the biomass of organs of dioecious plants is affected differently; among them, stem biomass is the least affected. The leaf biomass (value) of female plants is higher than that of male plants when annual precipitation is greater than 800 mm, which indicates that female plants invest more resources in leaves when there is an abundance of water. With the increase in annual rainfall, the flower biomass of female plants in our study showed a negative correlation trend, and when the annual rainfall was less than 500 mm, the female plants were significantly taller than the male plants. The overall results show that average annual precipitation had an insignificant effect on the vegetative biomass of dioecious plants, but a significant effect on reproductive biomass. When the precipitation level ranged from 500 to 800 mm, the reproductive biomass of female plants reached the highest value.
4.4. Life Forms of Dioecious Plants
Since the dioecious plants studied in this study include herbs, shrubs and trees, the plant organs of these different life forms have multiple characteristics; in addition, their allocation of resources varies among different life forms [86]. Comparing the stem, leaf and flower biomass of the three life forms of dioecious plants, the biomass of female shrubs was significantly lower than that of male shrubs, showing a negative correlation. There was no significant difference in plant biomass between male and female herbaceous plants. The reproductive biomass of trees was significantly higher than that of males. The stem biomass and flower biomass of male and female plants were not significantly different, but the leaf biomass was significantly higher than that of male and female plants. The stem biomass and flower biomass of female trees were significantly lower than that of male trees, and the two showed a negative correlation, while the leaf biomass was not significantly different from that of male trees. The results indicate a reproductive biomass of female plants; in other words, the nutrient biomass of dioecious shrubs was much higher than that of male plants, and the nutrient biomass of male plants was also higher than that of female plants, indicating that female plants invested more resources in reproductive distribution. There was no significant difference in reproductive biomass and vegetative biomass between male and female plants, and no significant difference in resource allocation between male and female plants. Trees are similar to shrubs and therefore invest more resources in their reproductive distribution [50]. In summary, the reproductive distribution of dioecious plants with different life forms is slightly different, but overall, the reproductive biomass of female plants is larger than that of male plants. In addition, the nutrient biomass of female plants is smaller than that of male plants, which is in line with the characteristics of the overall resource allocation of dioecious plants [87].
5. Conclusions
The resource allocation of dioecious plants is a very complex process. At the same time, due to the complexity of the growth of different dioecious plants in each independent test, the effects of soil texture and fertilization on dioecious plant biomass were not considered in this study [88]. In addition to the influence of objective conditions (such as sampling time at different growth stages of plants), some experimental results have specific temporal applicability. To understand it, we need to not only analyze the biomass of these plants, but also discuss the effects of multiple factors and consider the combined effects caused by various environmental changes to provide more databases for future meta-analyses. The quantitative study on the resource allocation of dioecious plants in this study is consistent with the empirical results. By searching published experimental data, the authors concluded that resource allocation was significantly greater in female plants than in male plants. The results showed that the female plant invested more in reproductive growth and less in vegetative growth. In terms of total biomass, the average biomass of female plants was 3.09% higher than that of male plants, indicating that female plants reduced nutrient input in resource allocation to compensate for reproductive input and that there was a trade-off between male and female functions. When plants increase the allocation of one sex, the input of the other sex will inevitably decrease, which is also in line with the relevant theory of resource allocation. The practical function of this feedback mechanism has a certain influence on the selection and development of plant breeding systems. At the same time, temperature and precipitation significantly affected the adaptability of male and female plants to environmental changes and the synergistic relationship between stress components. The vegetative biomass input of female plants was positively correlated with the increase in temperature, while the reproductive biomass input was negatively correlated with the decrease in temperature. The average annual precipitation had little effect on the vegetative biomass of dioecious plants but a significant effect on reproductive biomass. The study of this trade-off relationship is helpful to reveal the relationship between plant vegetative growth and reproductive growth, explore the countermeasures of plant life history and further understand the life history strategy of dioecious plants and its application and scientific layout in urban landscaping from a macro perspective, which will affect the planning and design of urban forest management.
Funding
This research was funded by “National Natural Science Foundation Project, grant number 32271832 and National Key R&D Program of China, grant number 2021YFE0193200”.
Data Availability Statement
The datasets analyzed during the current study are available from the corresponding author upon reasonable request.
Acknowledgments
The author sincerely thanks all the researchers for their work and for effectively providing the data of this meta-analysis, as well as the tutor for his guidance. The author also wishes to thank the anonymous reviewers for their valuable comments and suggestions on this manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Zhang, D.; Bai, W.; Ren, W.; Zhong, Y.; Shi, S.; Chen, X. Plant Life History Evolution and Reproductive Ecology; Science Press: Beijing, China, 2004. [Google Scholar]
- Xu, X.; Yang, F.; Yin, C.; Li, C. Progress in the research on the sex difference of dioecious plants in response to environmental stress. J. Appl. Biol. 2007, 18, 2626–2631. [Google Scholar]
- Lloyd, D.G. Selection of combined versus separate sexes in seed plants. Am. Nat. 1982, 120, 571–585. [Google Scholar] [CrossRef]
- Richards, A.J. Plant Breeding Systems; George Allen and Unwin: London, UK, 1986; pp. 67–71. [Google Scholar]
- Obeso, J.R. The costs of reproduction in plants. New Phytol. 2002, 155, 321–348. [Google Scholar] [CrossRef]
- Case, A.L.; Ashman, T.L. Sex-Specific Physiology and Its Implications for the Cost of Reproduction. In Reproductive Allocation in Plants; Reekie, E.G., Bazzaz, F.A., Eds.; Academic Press: New York, NY, USA, 2005; pp. 129–157. [Google Scholar]
- Iglesias, M.C.; Bell, G. The small-scale spatial distribution of male and female plants. Oecologia 1989, 80, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Jing, S.W.; Coley, P.D. Dioecy and herbivory: The effect of growth rate on plant defense in Acer negundo. Oikos 1990, 58, 369–377. [Google Scholar] [CrossRef]
- Jing, J.; Hong, J.; Shuquan, Y.; Guomo, Z. Sex-linked photosynthetic physiologic research and the evolutionary ecological analysis in living fossil plant, Ginkgo biloba L. Acta Ecol. Sin. 2008, 28, 1128–1136. [Google Scholar] [CrossRef]
- Shi, D.W.; Wei, X.D.; Chen, G.X.; Xu, Y. Changes in photosynthetic characteristics and antioxidative protection in male and female ginkgo during natural senescence. J. Am. Erican Soc. Hortic. Sci. 2012, 137, 349–360. [Google Scholar] [CrossRef]
- Cao, Q.; Jiang, H.; Zeng, B.; Yu, S.; Wang, B.; Jiang, F. Preliminary study on water physiological characteristics of male and female plants of Ginkgo biloba L. Anhui Agric. Sci. 2009, 37, 8732–8735. [Google Scholar]
- Xu, X.; Peng, G.; Wu, C.; Korpelainen, H.; Li, C. Drought inhibits photosynthetic capacity more in females than in males of Populus cathayana. Tree Physiol. 2008, 28, 1751–1759. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Chen, L.; Duan, B.; Korpelainen, H.; Li, C. Populus cathayana males exhibit more efficient protective mechanisms than females under drought stress. For. Ecol. Manag. 2012, 275, 68–78. [Google Scholar] [CrossRef]
- Xing, X.; Han, X.; Chen, L. Review on Plant Nutrient Use Efficiency. J. Appl. Ecol. 2000, 785–790. [Google Scholar] [CrossRef]
- Zhang, D. Theoretical Ecological Research; Higher Education Press: Beijing, China, 2000. [Google Scholar]
- Ren, Y. Resource Allocation and Ecological Service Function Evaluation of Dioecious Fraxinus tomentosa. Master’s Thesis, Beijing Forestry University, Beijing, China, 2020. [Google Scholar]
- Silvertown, J.; Dodd, M. The demagraphic cost of reproduction andits consequences in balsam fir (Abies balsamea). Am. Nat. 1999, 154, 321–332. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Chen, J.M.; Wang, Q.F. Trade-offs between sexual andasexual reproduction in a monoecious species Sagitfaria pygmaed (Alismataceae): The effect of different nutrient levels. Plant Syst. Evol. 2009, 277, 61–65. [Google Scholar] [CrossRef]
- Teitel, Z.; Pickup, M.; Field, D.L. The dynamics of resource allocation and costs of reproduction in a sexually dimorphic, wind-pollinated dioecious plant. Plant Biol. 2016, 18, 98–103. [Google Scholar] [CrossRef]
- Cornelissen, T.; Stiling, P. Sex-biased herbivory: A meta-analysis ofthe effects of gender on plant-herbivore interactions. Oikos 2005, 111, 488–500. [Google Scholar] [CrossRef]
- Rocheleau, A.F.; Houle, G. Different cost of reproduction for themales and females of the rare dioecious shrub Coremacorradii (Empetraceae). Am. J. Bot. 2001, 88, 659–666. [Google Scholar] [CrossRef]
- Leigh, A.; Cosgrove, M.J.; Nicotra, A.B. Reproductive allocation in agender dimorphic shurb: Anomalous females investment in Gynatrix pulchella. J. Ecol. 2006, 94, 1261–1271. [Google Scholar] [CrossRef]
- Yin, C.; Li, C. Research status and prospect of sex differences related to sex ratio in dioecious plants. J. Appl. Environ. Biol. 2007, 13, 419–425. [Google Scholar]
- Chen Xi Wei, H.; Lin, M. Research progress on the impact of climate change on dioecious plants. J. Ecol. 2014, 33, 3144–3149. [Google Scholar] [CrossRef]
- Yan, H.; Xiao, J.; Yang, Y. Research progress of dioecious plants based on bibliometrics. Guangxi Plants 2017, 37, 661–674. [Google Scholar]
- Zhu, J. Effects of Roots of Neighboring Plants of Different Sexes on the Growth of Populus cathayana. Master’s Thesis, China West Normal University, Nanchong, China, 2016. [Google Scholar]
- Liu, J.; You, M.; Duan, J.; Zhang, L. Plasticity regulation of reproductive strategy of dioecious plant Humulus scandens under water stress. Acta Prataculture 2015, 24, 226–232. [Google Scholar]
- Liu, H.; Zong, R.; Liu, J. Effects of grassland community types on population characteristics, morphology and resistance of female and male plants of Eupatorium odoratum. Acta Prataculture 2018, 27, 113–124. [Google Scholar]
- Wu, L. Physiological and Biochemical Responses of Male and Female Populus yunnanensis Plants to Enhanced UV-B Radiation and Heavy Metal Lead and Copper Stress. Master’s Thesis, Chengdu University of Technology, Chengdu, China, 2017. [Google Scholar]
- Liu, H.; Zong, R.; Liu, J. Effect of slope position on population characteristics and morphological plasticity of female and male plants of Eupatorium odoratum. Grassl. Sci. 2018, 35, 2105–2114. [Google Scholar]
- Wen, X.; Xian, T.; Liu, J.; Xu, X.; Dong, Y. Effects of defoliation on the growth and nonstructural carbohydrates of Populus cathayana. J. Ecol. 2021, 40, 41–48. [Google Scholar] [CrossRef]
- Guo, H.; Liu, H.; Liu, J. Gender differences in the effects of shading on the component characters and biomass structure of Humulus scandens. Grassl. Sci. 2018, 35, 2481–2488. [Google Scholar]
- Zhang, L. Effects of Drought Stress and Root Exudates Exchange on Nitrogen Transfer between Male and Female Plants of Populus cathayana. Master’s Thesis, China West Normal University, Nanchong, China, 2020. [Google Scholar]
- Fan, X.; Wang, D.; Liu, J. Gender differences in the effects of supports on the morphological plasticity and reproductive strategies of climbing Humulus scandens. J. Ecol. 2015, 34, 1559–1564. [Google Scholar] [CrossRef]
- Zhu, Z. Effects of Competition Modes on the Growth and Physiological Characteristics of Populus yunnanensis in Different Light. Environments. Thesis, Sichuan Agricultural University, Yaan, China, 2018. [Google Scholar]
- Li, B. Responses of Physiological and Ecological Characteristics of Populus deltoides Seedlings of Different Genders to Drought Stress. Master’s Thesis, Zhejiang A&F University, Hangzhou, China, 2021. [Google Scholar] [CrossRef]
- Guo, H.; Duan, J.; Liu, J. Gender differences in the effects of temperature on flower bud differentiation, pigment content and photosynthesis of male and female Humulus scandens. Acta Pratensis 2017, 26, 104–112. [Google Scholar]
- Zhang, L.; Zhang, D. Study on gender differences in growth and physiological response of Phellodendron amurense under shading conditions. Plant Res. 2020, 40, 735–742. [Google Scholar]
- Zhang, W. Study on Reproductive Ecology of Broussonetia papyrifera. Master’s Thesis, Sichuan Agricultural University, Yaan, China, 2008. [Google Scholar]
- Zhu, S. Effects of Root Exudates of Homosexual or Heterosexual Plants on the Growth of Populus cathayana. Master’s Thesis, West China Normal University, Chengdu, China, 2016. [Google Scholar]
- He, L.; Huang, X.; Liu, J. Analysis on the difference of physiological response and regeneration strategy between male and female plants of Humulus scandens during overwintering. Acta Prataceae 2020, 29, 49–59. [Google Scholar]
- Liu, J.; You, M.; Zhang, L.; Duan, Q. Effects of different supports on photosynthetic characteristics and biomass structure of male and female climbing plants-Humulus scandens. Acta Ecol. Sin. 2015, 35, 6032–6040. [Google Scholar]
- Liang, Y. Differences between Male and Female in Leaf Functional traits of Taxus cuspidata and Their Dynamic Changes in Growth Period. Master’s Thesis, Beijing Forestry University, Beijing, China, 2020. [Google Scholar] [CrossRef]
- Wang, L.; Huang, Y.; Fan, C.; Zhang, X.; Wang, J. Study on the sex difference of reproductive cost of Rhamnus rhamnoides in Northeast China. J. Cent. South Univ. For. Technol. 2020, 40, 69–74. [Google Scholar] [CrossRef]
- Huang, Y.; Xin, B.; Wang, J. Reproductive allocation and reproductive cost compensation mechanism of dioecious plant Rhamnus rhamnoides. J. Beijing For. Univ. 2019, 41, 31–36. [Google Scholar] [CrossRef]
- Zhao, F.; Yang, Y. Study on Reproductive Allocation of Polygonum cuspidatum Populations at Different Altitudes. Acta Bot. 2008, 830–835. [Google Scholar]
- Wang, J.; Zhang, C.; Zhao, X.; von Gadow, K. Reproductive distribution of dioecious plant Rhamnus rhamnoides. J. Ecol. 2011, 31, 6371–6377. [Google Scholar]
- Wang, J. Study on Reproductive Strategies of Dioecious Plants in Mixed Coniferous and Broad-Leaved Forests in Northeast China. Master’s Thesis, Beijing Forestry, Beijing, China, 2014. [Google Scholar]
- Li, L.; Barrett, S.C.H.; Song, Z.; Chen, J. Sex-specific plasticity of reproductive allocation in response to water depth in a clonal, dioecious macrophyte. Am. J. Bot. 2019, 106, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Cansino, L.; Zunzunegui, M.; Díaz Barradas, M.C.; Esquivias, M.P. Gender-specific costs of reproduction on vegetative growth and physiological performance in the dioecious shrub Corema album. Ann. Bot. 2010, 106, 989. [Google Scholar] [CrossRef]
- Bullock, S.H. Biomass and nutrient allocation in a neotropical dioecious palm. Oecologia 1984, 63, 426–428. [Google Scholar] [CrossRef]
- Cepeda-Cornejo, V.; Dirzo, R. Sex-related differences in reproductive allocation, growth, defense and herbivory in three dioecious neotropical palms. PLoS ONE 2010, 5, e9824. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Barrett, S.C.H.; Wang, X.J.; Zhong, L.; Wang, H.; Li, D.-Z.; Zhou, W. Sexual dimorphism, temporal niche differentiation, and evidence for the Jack Sprat effect in an annual dioecious plant. J. Syst. Evol. 2022, 60, 1078–1091. [Google Scholar] [CrossRef]
- Xu, X.; Peng, G.; Wu, C.; Han, Q. Global warming induces female cuttings of Populus cathayana to allocate more biomass, C and N to aboveground organs than do male cuttings. Aust. J. Bot. 2010, 58, 519–526. [Google Scholar] [CrossRef]
- Korpelainen, H. Patterns of resource allocation in male and female plants of Rumex acetosa and R. acetosella. Oecologia 1992, 89, 133–139. [Google Scholar] [CrossRef] [PubMed]
- García, M.B.; Antor, R.J. Sex ratio and sexual dimorphism in the dioecious Borderea pyrenaica (Dioscoreaceae). Oecologia 1995, 101, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Harris, M.S.; Pannell, J.R. Roots, shoots and reproduction: Sexual dimorphism in size and costs of reproductive allocation in an annual herb. Proc. R. Soc. B Biol. Sci. 2008, 275, 2595–2602. [Google Scholar] [CrossRef] [PubMed]
- Matsuyama, S.; Sakimoto, M. Allocation to reproduction and relative reproductive costs in two species of dioecious Anacardiaceae with contrasting phenology. Ann. Bot. 2008, 101, 1391–1400. [Google Scholar] [CrossRef] [PubMed]
- Matsuyama, S.; Sakimoto, M. Sexual dimorphism of reproductive allocation at shoot and tree levels in Zanthoxylum ailanthoides, a pioneer dioecious tree. Botany 2010, 88, 867–874. [Google Scholar] [CrossRef]
- Sakai, A.; Sasa, A.; Sakai, S. Do sexual dimorphisms in reproductive allocation and new shoot biomass increase with an increase of altitude? A case of the shrub willow Salix reinii (Salicaceae). Am. J. Bot. 2006, 93, 988–992. [Google Scholar] [CrossRef] [PubMed]
- Pickering, C.M.; Arthur, J.M. Patterns of resource allocation in the dioecious alpine herb Aciphylla simplicifolia (Apiaceae). Austral Ecol. 2003, 28, 566–574. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, C.; Gadow, K.V.; Cheng, Y.; Zhao, X. Reproduction and vegetative growth in the dioecious shrub Acer barbinerve in temperate forests of Northeast China. Plant Reprod. 2015, 28, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.Q.; Hui, D.F.; Zhang, D.Q. Elevated CO2 stimulates net accumulations of carbon and nitrogen in land ecosystems: A metaanalysis. Ecology 2006, 87, 53–63. [Google Scholar] [CrossRef]
- Jiang, L.; Liu, J.; Zhao, T.; Du, L. Meta-analysis of the effect of combined application of organic and inorganic fertilizers on tomato yield and quality. J. Plant Nutr. Fertil. 2019, 25, 601–610. [Google Scholar]
- Wei, H.; Wang, S.; Yang, W.; Sun, H.; Yin, L.; Deng, X.L. Integrated analysis of the effects of no-tillage and subsoiling on the yield and water use efficiency of spring corn and winter wheat in the Loess Plateau. China Agric. Sci. 2017, 50, 461–473. [Google Scholar]
- Card, N.A. Applied Meta-Analysis for Social Science Research; Guilford Press: New York, NY, USA, 2011; pp. 198–228. [Google Scholar]
- Begg, C.B.; Berlin, J.A. Publication bias: A problem in interpreting medical data. J. R. Stat. Soc. 1988, 151, 419–463. [Google Scholar] [CrossRef]
- Yin, M.; Li, Y.; Chen, P.; Xu, L.; Sheng, S.; Wang, X. Study on yield effect of no-tillage corn in northern China based on Meta-analysis. China Agric. Sci. 2018, 51, 843–854. [Google Scholar]
- Rosenberg, M.S.; Adams, D.C.; Gurevitch, J. MetaWin: Statistical Software for Meta-Analysis; Sinauer Associates: Sunderland, MA, USA, 2000; pp. 198–228. [Google Scholar]
- Barrett, S.C.H. Gender variation in Vurmbea dioica (Liliaceae) and the evolutinonary analysis. J. Evol. Biol. 1992, 5, 423–444. [Google Scholar] [CrossRef]
- Huang, X. Differences in Response Ability and Coping Strategies of Male and Female Humulus scandens to Low Temperature stress. Master’s Thesis, China West Normal University, Nanchong, China, 2021. [Google Scholar]
- Guo, M.; Sun, S. Research progress of plant reproductive distribution and its adaptation strategies to the environment. J. North. Agric. 2016, 44, 99–103. [Google Scholar]
- Delph, L.F. Sexual Dimorphism in Live History. In Gender and Sexual Dimorphism in Flowering Plants; Geber, M.A., Dawson, T.E., Delph, L.F., Eds.; Springer: Berlin, Germany, 1999; pp. 149–173. [Google Scholar]
- Cipollini, M.L.; Whigham, D.F. Sexual dimorphism and cost of reproduction in the dioecious shrub Lindera benzoin (Lauraceae). Am. J. Bot. 1994, 81, 65–75. [Google Scholar] [CrossRef]
- Bazzazf, A.; Ackerlyd, D.; Reekie, E.G. Reproductive Allocation in Plants; CABI Publishing: New York, NY, USA, 2000. [Google Scholar]
- Yan, S. Research progress on gender response difference of poplar under environmental stress. Liaoning For. Sci. Technol. 2022, 56–60+67. [Google Scholar]
- Song, Y.; Ma, K.; Ci, D.; Zhang, Z.; Zhang, D. Biochemical, physiological and gene expression analysis reveals sex-specific differences in Populus tomentosa floral development. Physiol. Plant. 2014, 150, 18–31. [Google Scholar] [CrossRef]
- Allen, G.A.; Antos, J.A. Sex ratio variation in the dioecious shrub Oemleria cerasiformis. Am. Nat. 1993, 141, 537–553. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Zhang, X.; Wang, Z. Study on reproductive ecology of goose web: Reproductive distribution and life history strategy. J. Nanjing For. Univ. (Nat. Sci. Ed.) 2004, 71–74. [Google Scholar]
- Kevan, P.G. Sexual Differences in Temperatures of Blossoms on a Dioecious Plant, Salix arctica: Significance for Life in the Arctic. Arct. Alp. Res. 1990, 22, 283–289. [Google Scholar] [CrossRef]
- Cao, S.-K.; Feng, Q.; Si, J.-H.; Chang, Z.-Q.; Xi, H.-Y.; Zhuo, M. Summary of research methods of plant water use efficiency. China Desert 2009, 29, 853–858. [Google Scholar]
- Wardj, K.; Dawsont, E.; Ehleringerj, R. Responses of Acer negundo genders to interannual differences in wateravailability determined from carbon isotope ratios of tree ring cellulose. Tree Physiol. 2002, 22, 339–346. [Google Scholar] [CrossRef]
- Garcia-Barreda, S.; Sangüesa-Barreda, G.; García-González, M.D.; Camarero, J.J. Sex and tree rings: Females neither grow less nor are less water-use efficient than males in four dioecious tree species. Dendrochronologia 2022, 73, 125944. [Google Scholar] [CrossRef]
- Dawson, T. Patterns of water use and the tissue water relations in the dioecious shrub, Salix arctica: The physiological basis for habitat partitioning between the sexes. Oecologia 1989, 79, 332–343. [Google Scholar] [CrossRef]
- Retuerto, R.; Lema, B.F.; Obeso SR, R.R. Gender, light and water effects in carbon isotope discrimination, and growth rates in the dioecious tree Ilex aquifolium. Funct. Ecol. 2000, 14, 529–537. [Google Scholar] [CrossRef]
- Freeman, D. Differential resource utilization by the sexes of dioecious plants. Science 1976, 193, 597–599. [Google Scholar] [CrossRef]
- Delph, L.F.; Meagher, T.R. Sexual dimorphism masks life history trade-off in the dioecious plant Silene latifolia. Ecology 1995, 76, 775–785. [Google Scholar] [CrossRef]
- Li, L.; Duan, B.; Deng, D.; Tu, W.; Zhang, Y. Soil salinity alters the sexual responses to elevated CO2 and temperature in growth and leaf traits of a dioecious plant. Can. J. For. Res. 2014, 44, 1292–1301. [Google Scholar] [CrossRef]
Figure 1.
Flowchart of the analytical process.

Figure 2.
Flowchart of the data extraction process.

Figure 3.
Study site distribution map.

Figure 4.
Response ratio frequency distribution of biomass proportion in different organs of dioecious plants.
Figure 4.
Response ratio frequency distribution of biomass proportion in different organs of dioecious plants.

Figure 5.
Biomass allocation effect of dioecious plants (CI, confidence interval).

Figure 6.
Effects of different temperatures on biomass accumulation in dioecious plants. (CI, confidence interval).
Figure 6.
Effects of different temperatures on biomass accumulation in dioecious plants. (CI, confidence interval).

Figure 7.
Effects of rainfall on biomass accumulation in dioecious plants (CI, confidence interval).
Figure 7.
Effects of rainfall on biomass accumulation in dioecious plants (CI, confidence interval).

Figure 8.
Effects of different life forms on the biomass accumulation in dioecious plants (CI, confidence interval).
Figure 8.
Effects of different life forms on the biomass accumulation in dioecious plants (CI, confidence interval).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
List of scientific publications, dioecious plant species and research contents.
| Literature | Plant Habitus | Plant Families and Genera | Experimental Method | Research Contents | ||||
|---|---|---|---|---|---|---|---|---|
| Biomass Determination | Phenological Observation | Physiological Measurement | Photosynthetic Pigment | Sex Ratio | ||||
| Zhu Juan, 2016 [26] | tree | Populus | NE | ⚪ | ⚪ | \ | ⚪ | ⚪ |
| Liu Jinping et al., 2015 [27] | herbs | Humulus | NE | ⚪ | \ | ⚪ | ⚪ | \ |
| Liu Hangjiang et al., 2018 [28] | herbs | Anaphalis DC | NE | ⚪ | ⚪ | \ | \ | ⚪ |
| Ren Yaxue, 2020 [12] | tree | Bambusa | NE | ⚪ | ⚪ | ⚪ | \ | \ |
| Wu Liuran, 2017 [29] | tree | Populus | NE | ⚪ | \ | ⚪ | ⚪ | \ |
| Liu Hangjiang et al., 2018 [30] | herbs | Anaphalis DC | NE | ⚪ | ⚪ | \ | \ | ⚪ |
| Wen Xiaomei et al., 2021 [31] | tree | Populus | NE | ⚪ | \ | ⚪ | \ | \ |
| Guo Haiyan et al., 2018 [32] | herbs | Humulus | GC | ⚪ | \ | ⚪ | ⚪ | \ |
| Zhang Ling, 2020 [33] | tree | Berberis | GC | ⚪ | \ | ⚪ | ⚪ | \ |
| Fan Xing et al., 2015 [34] | herbs | Humulus | NE | ⚪ | ⚪ | \ | \ | ⚪ |
| Zhenzhen Zhu, 2018 [35] | tree | Populus | GC | ⚪ | ⚪ | ⚪ | ⚪ | \ |
| Li Bin, 2021 [36] | tree | Populus | GC | ⚪ | ⚪ | ⚪ | \ | ⚪ |
| Guo Haiyan et al., 2017 [37] | herbs | Humulus | GC | ⚪ | ⚪ | \ | ⚪ | \ |
| Zhang Linmin et al., 2020 [38] | tree | Populus | GC | ⚪ | ⚪ | ⚪ | \ | \ |
| Zhang Wang, 2008 [39] | tree | Populus | NE | ⚪ | ⚪ | \ | \ | ⚪ |
| Zhu Shihui, 2016 [40] | tree | Populus | GC | ⚪ | \ | ⚪ | ⚪ | \ |
| He Linjiang et al., 2020 [41] | herbs | Humulus | NE | ⚪ | ⚪ | \ | \ | ⚪ |
| Liu Jinping et al., 2015 [42] | herbs | Humulus | GC | ⚪ | \ | ⚪ | \ | \ |
| Liang Yanjun, 2020 [43] | tree | Taxus | NE | ⚪ | \ | ⚪ | ⚪ | \ |
| Wang Lijun et al., 2020 [44] | tree | Rhamnus | NE | ⚪ | \ | ⚪ | ⚪ | \ |
| Huang Yunhao, 2019 [45] | tree | Rhamnus | NE | ⚪ | \ | \ | ⚪ | ⚪ |
| Zhao Fang et al., 2008 [46] | herbs | Polygonum | NE | ⚪ | ⚪ | \ | \ | \ |
| Wang Juan et al., 2011 [47] | tree | Rhamnus | NE | ⚪ | ⚪ | \ | \ | ⚪ |
| Wang Juan, 2014 [48] | tree | Rhamnus | NE | ⚪ | ⚪ | \ | ⚪ | \ |
| Lei Li et al., 2019 [49] | herbs | Valleya | NE | ⚪ | ⚪ | \ | \ | \ |
| Leonor et al., 2010 [50] | shrub | Rhododendron | NE | ⚪ | \ | ⚪ | \ | \ |
| Stephen H. Bullock, 1984 [51] | tree | Trachycarpus H. Wendl. | NE | ⚪ | ⚪ | \ | \ | ⚪ |
| Vero’ nica Cepeda-Cornejo et al., 2010 [52] | tree | Trachycarpus H. Wendl. | NE | ⚪ | ⚪ | ⚪ | \ | \ |
| Qian Yu et al., 2022 [53] | herbs | Rumex | GC | ⚪ | ⚪ | \ | \ | ⚪ |
| Xiao Xu et al., 2010 [54] | tree | Populus | GC | ⚪ | ⚪ | \ | ⚪ | \ |
| Zachary Teitel et al., 2015 [19] | herbs | Rumex | GC | ⚪ | ⚪ | \ | \ | ⚪ |
| Helena Korpelainen et al., 1992 [55] | herbs | Rumex | NE | ⚪ | ⚪ | \ | \ | ⚪ |
| Maria B. Garcia Ramdn J. Antor et al., 1995 [56] | herbs | Dioscorea | NE | ⚪ | ⚪ | ⚪ | \ | \ |
| Mark S. Harris et al., 2008 [57] | herbs | Indigo | GC | ⚪ | ⚪ | ⚪ | \ | \ |
| Shuhei Matsuyama et al., 2008 [58] | tree | Sorbus | NE | ⚪ | ⚪ | \ | ⚪ | \ |
| Shuhei Matsuyama et al., 2010 [59] | tree | Zanthoxylum L. | NE | ⚪ | ⚪ | \ | ⚪ | ⚪ |
| AKIKO SAKAI et al., 2006 [60] | shrub | Salix | NE | ⚪ | ⚪ | \ | \ | ⚪ |
| CATHERINE MARINA PICKERING et al., 2003 [61] | herbs | Lotus | NE | ⚪ | ⚪ | ⚪ | \ | \ |
| Juan Wang et al., 2015 [62] | shrub | Acer | NE | ⚪ | ⚪ | ⚪ | \ | \ |
Note: ⚪, this paper has research content; \, no research content in this paper; GC, greenhouse; NE, natural growth.
Table 2.
Heterogeneity and publication bias test of the biomass ratio of different organs in female and male dioecious plants.
Table 2.
Heterogeneity and publication bias test of the biomass ratio of different organs in female and male dioecious plants.
| Index | Sample Size | Q | pQ | ZB | pB |
|---|---|---|---|---|---|
| Stem biomass | 91 | 148.28 | 0.001 | 0.689 | 0.491 |
| Leaf biomass | 107 | 257.97 | 0.0001 | −0.200 | 0.316 |
| Flower biomass | 100 | 218.32 | 0.0001 | −0.152 | 0.879 |
| Vegetative biomass | 132 | 279.76 | 0.0001 | −0.181 | 0.051 |
| Reproductive biomass | 115 | 240.4 | 0.0001 | 0.133 | 0.158 |
Note: Q represents heterogeneity test statistics; pQ represents the significance of q statistics; ZB stands for statistics of publication bias; pB represents the significance of ZB statistics.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhao, M.; Zhang, X.; Xu, C.; Li, P.; Latortezza, R. Temperature and Precipitation Significantly Affect Resource Allocation in Dioecious Plants: A Meta-Analysis. Forests 2024, 15, 720. https://doi.org/10.3390/f15040720
AMA Style
Zhao M, Zhang X, Xu C, Li P, Latortezza R. Temperature and Precipitation Significantly Affect Resource Allocation in Dioecious Plants: A Meta-Analysis. Forests. 2024; 15(4):720. https://doi.org/10.3390/f15040720
Chicago/Turabian StyleZhao, Mingjie, Xinna Zhang, Chengyang Xu, Pin Li, and Raffaele Latortezza. 2024. "Temperature and Precipitation Significantly Affect Resource Allocation in Dioecious Plants: A Meta-Analysis" Forests 15, no. 4: 720. https://doi.org/10.3390/f15040720
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.