

Mycorrhizal Associations between Helvella bachu and Its Host Plants


1
Xinjiang Production & Construction Corps Key Laboratory of Protection and Utilization of Biological Resources in Tarim Basin, College of Life Science and Technology, Tarim University, Alar 843300, China
2
State Key Laboratory of Mycology, Institute of Microbiology, Chinese Academy of Sciences, Beijing 100101, China
*
Authors to whom correspondence should be addressed.
Forests 2024, 15(4), 721; https://doi.org/10.3390/f15040721
Submission received: 4 March 2024
/
Revised: 8 April 2024
/
Accepted: 12 April 2024
/
Published: 19 April 2024
(This article belongs to the Special Issue Fungal Interactions with Host Trees and Forest Sustainability)
Abstract
:Helvella bachu, a prized edible and medicinal fungus, is primarily found in the forests of Populus euphratica, an ancient and endangered species crucial to desert riparian ecosystems. Despite extensive efforts, the isolation of pure cultures and cultivation of fruiting bodies of H. bachu have remained elusive. While some species within the Helvella genus have been confirmed as ectomycorrhizal fungi, others have been considered either saprotrophic or mycorrhizal. By integrating field observations of H. bachu habitat, macro- and micro-anatomical examination of plant root tips, and molecular data from fruiting bodies, mycorrhizae, and host plants, it has been confirmed that H. bachu forms ectomycorrhizal associations with Populus trees. The mycorrhiza of H. bachu displays a light earth color with a curved smooth cylindrical shape. It features a thick mantle and the presence of a Hartig net, accompanied by a small amount of epitaxy mycelia. Morphological observation of the root tips requires meticulous handling, and the paraffin section technique has yielded noteworthy results. Host plants encompass four Populus species, including P. euphratica, P. pruinosa, P. nigra, and P. alba var. pyramidalis (synonym Populus bolleana). A conservation area was established within the young P. euphratica forest at Tarim University, resulting in a 14.75% increase in the quantity of fruiting bodies during the second year. Establishing a conservation area and in situ propagation of H. bachu holds economic and ecological implications. This study will contribute to the conservation of resources related to H. bachu and P. euphratica.
1. Introduction
Bachu mushrooms are commonly enjoyed as delicious edible fungi in the Xinjiang Autonomous Region, China [1], as well as in Pakistan [2] and Iraq [3]. These mushrooms have received considerable attention in recent years due to their high economic, nutritional, and medicinal values [4]. They have been reported to efficiently enhance the phagocytic ability of leukocytes, lymphocyte conversion ratio, and antibody titer [5]. The polysaccharides from Bachu mushrooms exhibit strong antioxidant activities and demonstrate a relatively high inhibition rate on HepG2 (hepatocellular carcinoma cell) cells [6]. Additionally, the polysaccharides can attenuate high-fat-diet-induced high blood cholesterol and regulate the expression of lipid metabolism genes in vivo. This makes them a promising novel complementary alternative strategy for managing hyperlipidemia [7].
However, Bachu mushrooms have proven challenging to cultivate successfully, and obtaining a pure culture has also been elusive after years of effort. The annual yield of dried fruiting bodies of the Bachu mushrooms in China is up to 3000–5000 kg, averaging USD 150 per kilogram. In addition, Bachu mushrooms are distributed in forests of Populus euphratica, which is a rare, ancient, and endangered species and an essential component of desert riparian ecosystems [8]. Hence, the artificial domestication of Bachu mushrooms holds both economic and ecological significance. Understanding the fungal nutritional mode is essential for the successful artificial domestication and cultivation of this valuable species.
Bachu mushrooms were confirmed to comprise two distinct new species, Helvella bachu and H. subspadicea [1]. H. bachu is a common and widespread species in the southern Xinjiang Autonomous Region, with “bachu” proposed as the trade name for Bachu mushroom. Pezizales species constitute a considerable proportion of the mycobionts in ectomycorrhiza (ECM) fungal communities in mature boreal deciduous and coniferous forests across several soil types. Two species of Helvella, Helvella aff. Cupuliformis and H. crispa, were detected on the root tips of oak and Pinus armandii [9,10]. Previous studies showed that H. aestivalis and two unidentified Helvella spp. formed ECM with Dryas sp. And Salix sp. [11]. The study by Hwang et al. [10] revealed that mycorrhizal lineages were distributed across the Helvella phylogeny. They confirmed that certain species of Helvella—such as H. dovrensis, H. lacunosa, H. zhongtiaoensis, H. reflexa, H. cf. crispa, H. macropus and Helvella cf. Lacunosa [12]—were ectomycorrhizal fungi. Meanwhile, H. elastica and H. atra were highly likely to be saprophytic fungi. H. bachu was reported to be scattered or gregarious on the sandy ground under P. euphratica or P. bolleana [1]. Whether it can form ECM with Populus is important for achieving the pure culture and artificial cultivation of this fungus and for the protection of P. euphratica forests.
Here, through investigating the field habitat of H. bachu, examining the macro- and micro-anatomical characteristics of the host root tip, and conducting molecular biological identification of the fruiting body and mycorrhiza, this study has initially demonstrated that H. bachu is an ectomycorrhiza. This finding helps to understand the nutritional mode of H. bachu and offers guidance for its artificial cultivation. Additionally, these results hold great significance for the restoration of degraded poplar forests and desert ecosystem restoration in Xinjiang.
2. Materials and Methods
2.1. Sampling, Processing, and Observation
Fungal samples were collected in Aral City, Bachu, and Maigaiti Counties of Kashi Prefecture, Xinjiang Uygur Autonomous Region of China, in April 2022 and 2023. Sample details, including fruiting bodies of H. bachu and soils and leaves of host plants, are listed in Table 1. The growth state of H. bachu in each location was photographed and observed. Additionally, the occurrence quantity of H. bachu and the distance between H. bachu and plants were measured. The soil pH, nitrogen (N), phosphorus (P), and potassium (K) contents were tested using a seven-in-one soil parameter speedometer (Prsens, Jinan, China).
Root samples in the soil with a diameter of 20 cm and depth of 20 cm were collected at the location where the fruiting bodies of H. bachu occurred. These samples were carefully transported back to the laboratory in a sampling box within 2 h and were subsequently preserved at 4 °C for no more than one week. The root tips were handled very carefully following the method of Wang et al. [13]. First, the loose soil outside was removed by gentle shaking. Then, the roots were delicately washed in tap water to eliminate most of the soil and organic debris, minimizing any damage to ECM roots. Tightly adhering materials were removed carefully using forceps. Subsequently, the cleaned roots were cut into 2 cm long segments and placed in a Petri dish filled with tap water. The enlarged thickened root tips were meticulously selected and examined under a stereo microscope (SMZ1500, Nikon, Minato-ku, Tokyo, Japan) to observe the color, shape, size, texture, branching, emanating elements, and other taxonomically relevant morphological features [14,15].
The microstructures of mycorrhiza were identified using the paraffin section technique. The cleaned root tips were immersed in FAA fixed liquid (formaldehyde/glacial acetic acid/alcohol/distilled water at a ratio of 6:1:20:40), subjected to vacuum pumping, and then placed into 4 °C refrigerators. After fixing for more than 24 h, the root was treated with 10% KOH in a water bath at 90 °C until it became translucent. Subsequently, it was dehydrated using various alcohol gradients, followed by transparency treatment with xylene, wax soaking, embedding in molds, solidification in a freezer, and subsequent slicing. After cutting into different wax strips, they were placed in a constant temperature water bath at 40 °C to adhere to slides coated with adhesive. The slides were then put in an incubator overnight to dry, followed by dewaxing with xylene. Staining was performed using trypan blue, and then a decolorizing solution was applied [16,17]. Following microscopic examination of the lateral distribution of the mantle and the Hartig net in the intercellular space using a Nikon Eclipse 80i microscope (Nikon, Minato-ku, Tokyo, Japan), a permanent sheet was created using neutral resin for sealing.
2.2. DNA Analyses of H. bachu Ectomycorrhizae and Host Plants
The mycorrhizal samples were carefully selected and then frozen in a 2 mL centrifugal tube using liquid nitrogen. Subsequently, they were placed in a −80 °C refrigerator in preparation for DNA extraction. The improved CTAB method was utilized to extract mycorrhizal DNA, followed by the use of ITS1-F and ITS4 primers (ITS1-F: CTTGGTCATTTAGAGGAAGTAA; ITS4: TCCTCCGCTTATTGATATGC) [18] for PCR amplification. The system comprised 25 μL of green mix (Cat No. P222-w1, Vazyme Biotech Co., Ltd., Nanjing, China), 2 μL of ITS1-F and 2 μL of ITS4, 2 μL of template DNA, and 19 μL of distilled water. The PCR amplification reaction procedure involved pre-denaturation at 94 °C for 5 min, followed by 35 cycles of denaturation at 94 °C for 30 s, annealing at 55 °C for 15 s, extension at 72 °C for 15 s, and final extension at 72 °C for 10 min. The PCR products were analyzed using 1% agarose gel electrophoresis at a setting voltage of 120 V and current of 100 mA for 40~50 min. The 780 bp target band was then purified and recovered using a Vazyme kit, and the PCR products were sequenced by Sangon Biotech Co., Ltd. (Shanghai, China).
Leaves of the host plant were collected from 10 locations where H. bachu with different hosts were found (Table 1). The ITS2 sequence of Salicaceae nuclear gene was amplified using S2F and S3R primers (S2F: 5 ‘ATGCGATACTTGGTGTGAAT 3’: S3R 5 ‘GACGCTTCTCCAGACTACAAT 3’) [19]. The PCR reaction conditions for 25 μL comprised 12.5 μL of mix, 1 μL of S2F, 1 μL of S3R, 1 μL of template DNA, and 9.5 μL of distilled water. The PCR amplification procedure involved initial denaturation at 94 °C for 5 min, followed by 40 cycles of denaturation at 94 °C for 30 s, annealing at 56 °C for 30 s, extension at 72 °C for 45 s, and final extension at 72 °C for 10 min. The PCR products were then sent to Sangon Biotech Co., Ltd. for sequencing.
2.3. Phylogenetic Analyses
Relevant nucleotide sequences were downloaded from the GenBank database to determine the taxonomic status of mycorrhizae and host plants. Homologous ITS sequences obtained from GenBank were aligned using the MultAlin program version 5.4.1 [20] from the INRA server with a DNA-5-0 score matrix. Alignments were thoroughly reviewed and manually edited as needed. For phylogenetic analyses, sequence distances were calculated using Kimura’s two-parameter model in the MEGA7 program [21], with pairwise deletion of gaps. Phylogenetic trees were constructed via maximum likelihood in the MEGA7 program and evaluated through bootstrapping with 1000 replicates. The resulting phylogenetic trees were visualized and edited using the Tree Explorer tool within MEGA7 version 7.0.26 [21].
3. Results
3.1. Occurrence of Helvella bachu Mushroom
We collected Bachu mushrooms from two regions of Xinjiang Uygur Autonomous Region—Alar City and Kashi Prefecture—from 2022 to 2023. A comprehensive investigation was conducted in twelve main production locations (Table 1). The fruiting bodies were identified as H. bachu (Figure 1A,B) using universal primers for fungi (ITS4 and ITS5), and the sequence of samples from Bachu county was deposited in GenBank under accession number PP082042. It was found that H. bachu occurs only in April and May each year around the Yeerqiang River, Tarim River, and Hotan River basin, growing under Populus trees or aspen trees (Figure 1C). The area is situated in a warm temperate zone with an extreme continental arid desert climate characterized by an annual average temperature ranging from 9.8 to 11.4 °C. The temperature during the occurrence of the mushroom varied from 15 to 31 °C. Additionally, the soil composition in the region was predominantly sandy. The Aral area experienced an average annual rainfall of 40.1~82.5 mm.
H. bachu was found scattered or clustered on the sandy ground near Populus (Figure 1D,E), with some even found beneath the Populus (Figure 1F). Additionally, we observed the mushroom in the vicinity of Medicago sativa L., but several meters away there were also Populus trees (Figure 1G). The soil pH was recorded between 6.5 and 7.2, with N, P, and K content measured at 18.89, 25.67, and 64.22 mg/kg, respectively, and a conductivity of about 267 μs/cm. Fresh H. bachu mushrooms are commonly traded in the market from April to May each year and have gained popularity (Figure 1H,I).
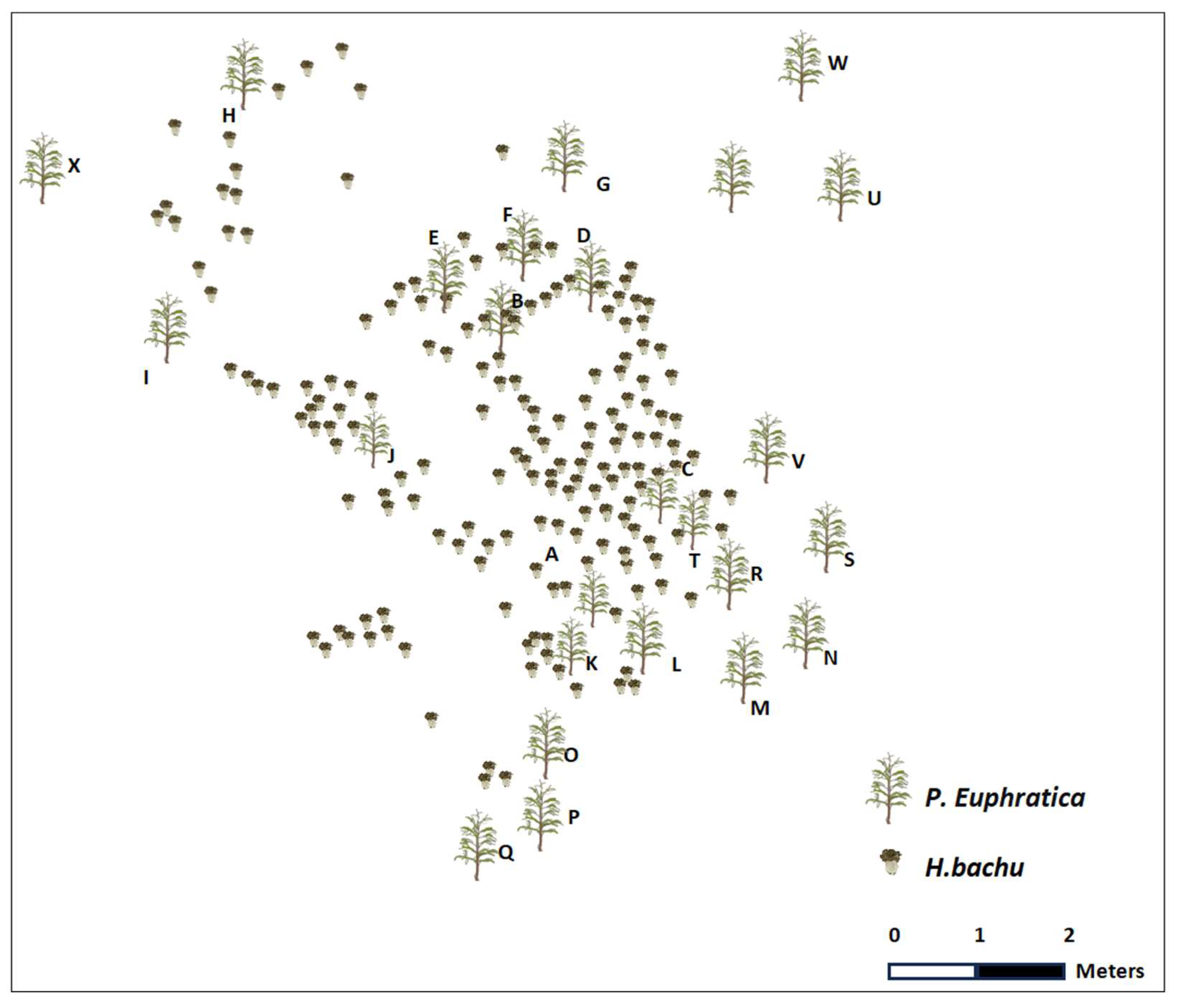
In order to observe the occurrence of H. bachu and its surrounding environments, a quadrat measuring about 300 square meters was selected in Tarim University, Alar City (Figure 2). There were 24 P. euphratica trees approximately five to ten years old within the selected quadrat. In 2022, the diameter of Populus ranged from 5.23 cm to 23 cm, with an average diameter of 11.13 cm. The average distance between Populus trees was measured at 107.48 cm, with the farthest distance recorded as 451 cm and the closest as 1 cm. During April and May in both 2022 and 2023, a detailed observation of the mushroom H. bachu in this quadrat was conducted. A total of 122 mushrooms was observed in 2022, while 140 were observed in 2023. The distance between the mushrooms varied from 0 cm to 395 cm, with an average distance of 47.98 cm. Additionally, the distance between the mushrooms and the trees ranged from 0 cm to 274.5 cm, with an average distance of 113.55 cm. Interestingly, although there were no H. bachu mushrooms around some Populus trees in 2022 (tree W and U in Figure 2), a significant harvest of mushrooms occurred in 2023. This highlights the fluctuating nature of fungal distribution in relation to host trees. This experiment conducted at Tarim University provided valuable insights into the occurrence of H. bachu mushrooms and their relationship with the surrounding environment.
3.2. Ectomycorrhiza Identification through Analysis of ITS Sequences
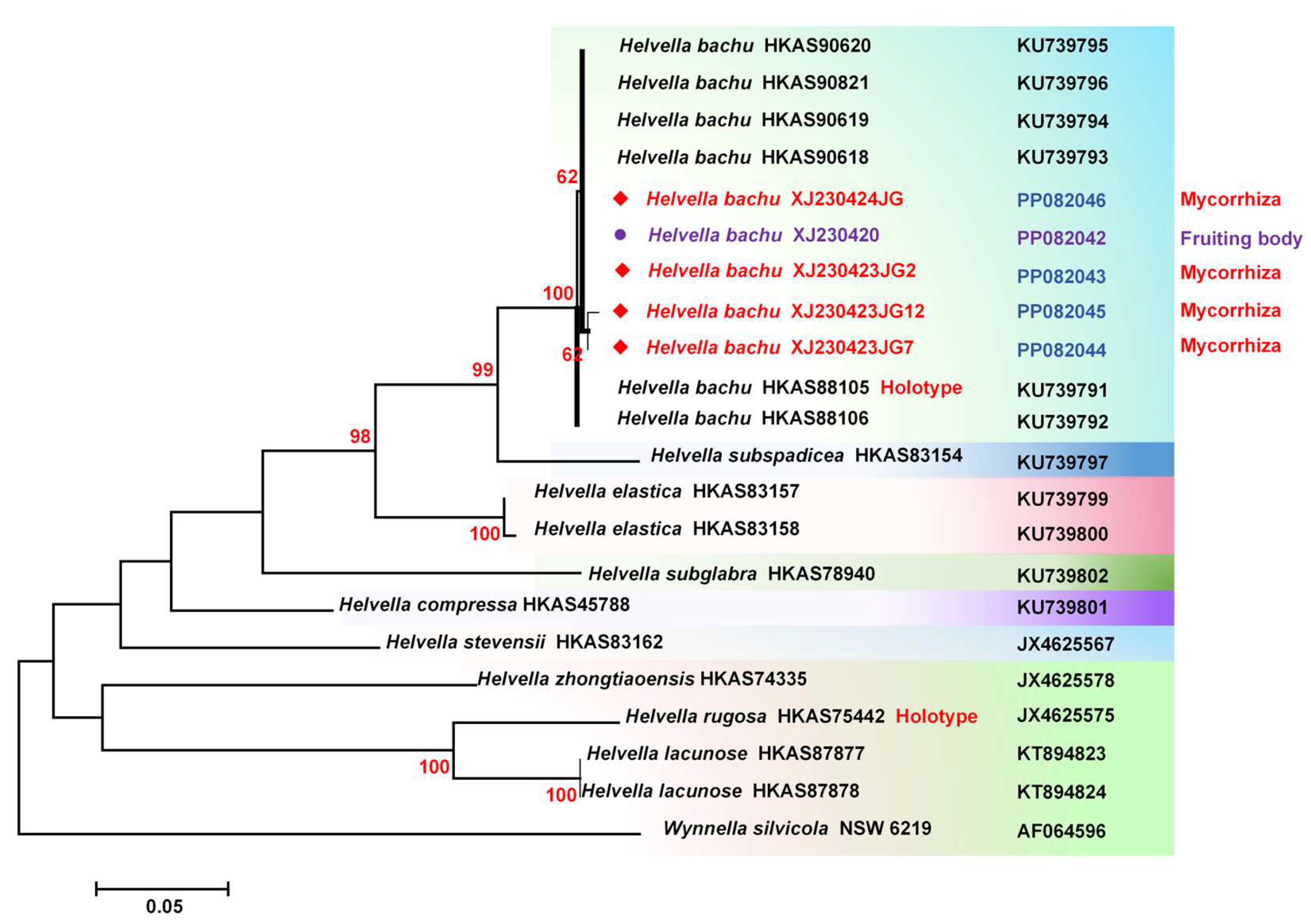
Following the harvest of fruiting body of H. bachu, several mycorrhizal samples were carefully selected under a stereo microscope. A total of 20 mycorrhizal samples were collected from the location where the fruiting bodies occurred in the aforementioned P. euphratica forest quadrat and were blended for DNA extraction. PCR amplification was performed using the specific primers ITS1F and ITS4 for ectomycorrhiza [18], which resulted in a target band of approximately 780 bp. The amplified DNA was subsequently sequenced and the sequence has been deposited in GenBank under the accession number PP082046. Phylogenetic analysis conclusively identified the mycorrhizal samples as H. bachu (Figure 3).
Additionally, mycorrhizal samples were collected from windbreak poplars in Yahumudan village, Bazhaziemi town, Makti county, and Kashi City, where the fruiting bodies of H. bachu were found. Twenty mycorrhizal samples with different morphologies were selected, and three of them displaying similar morphology were identified as H. bachu (Figure 3) with GenBank entry numbers PP082043, PP082044, and PP082045. Based on these findings, it is highly probable that H. bachu is a type of ectomycorrhizal fungi.
3.3. Macro- and Micro-Morphological Characteristics of Mycorrhiza of Helvella bachu
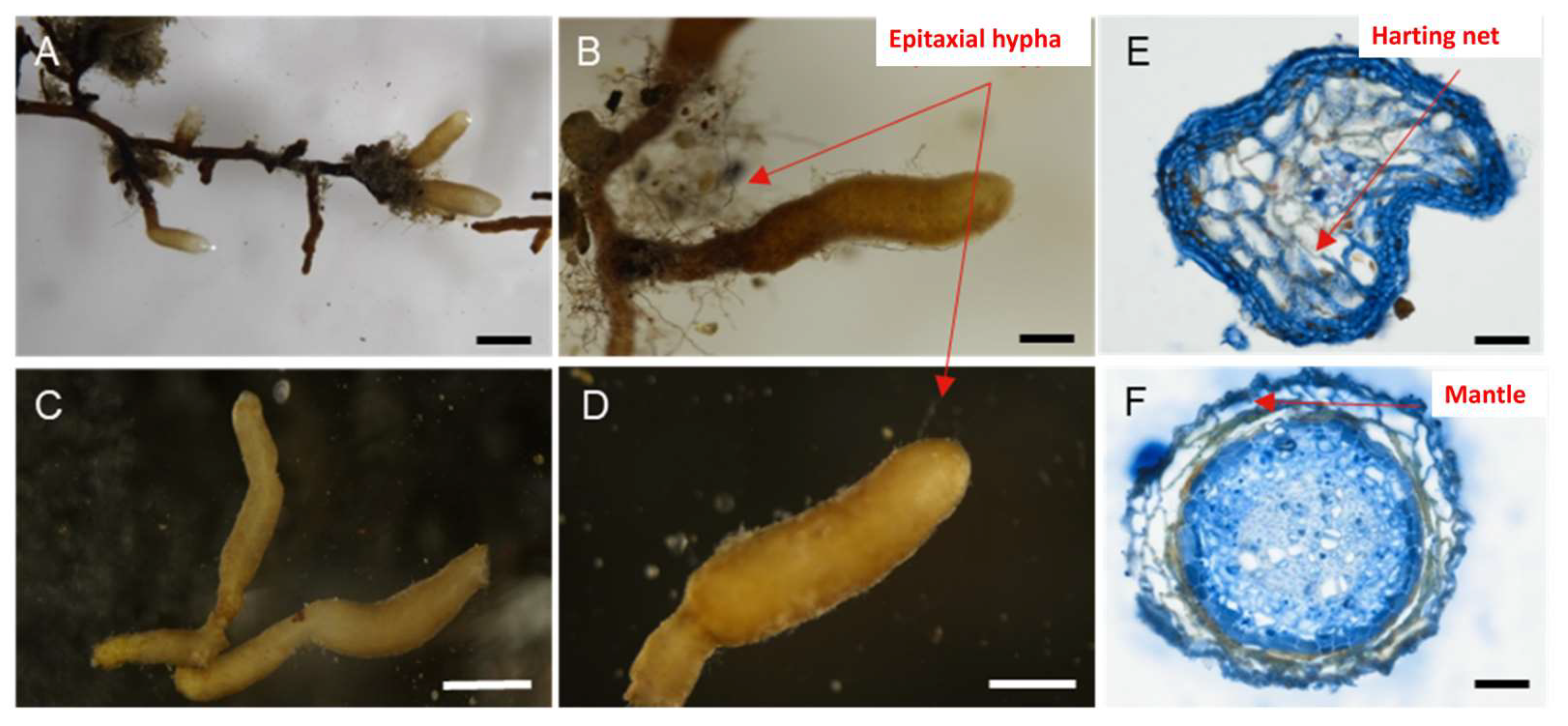
The root tips displayed a light earth color (Figure 4A–D). The mycorrhiza formed a coral-like structure without branches. The diameter of the mycorrhiza ranged from 200 to 500 μm, while the length varied from 1 to 2 mm. A thin sheath was observed wrapping around the root tip, with some hyphae present surrounding the root system (Figure 4B). The mycorrhizae appeared with a light earth color and had a curved smooth cylindrical shape. Paraffin sectioning and trypan blue staining revealed that the intercellular space in the transverse section was stained blue, indicating that the mycelia had penetrated the intercellular space and formed the characteristic structure known as the Hartig net (Figure 4E,F). Epitaxial hyphae were observed around the mycorrhizal structure (Figure 4D). Additionally, the mycorrhizae were encircled by a dense network of white hyphae on the surface of the root system, referred to as the mantle (Figure 4F).
Considering the molecular identification, morphology, and micro-anatomical structure, it is evident that H. bachu is closely associated with the host plant and functions as an ectomycorrhiza through symbiosis.
3.4. Identification of the Host Plant of Helvella bachu
We collected leaves from the 10 locations where the H. bachu mushrooms were harvested (Table 1) to determine the specific species of the Populus trees that serve as hosts for H. bachu. DNA was extracted, amplified, and sequenced with the nuclear gene fragment ITS2, targeting a nuclear gene specific to Poplus (S2F and S3R) for identification. The sequences have been deposited in NCBI with accession numbers PP188354 to PP188366. Phylogenetic analyses revealed the presence of four kinds of Populus trees, namely P. euphratica, P. pruinosa, P. nigra, and P. alba var. pyramidalis (synonym Populus bolleana) (Figure 5). Notably, P. alba var. pyramidalis and P. tremula (Specimen H1, 2, 8, and 10) were grouped together and could not be distinguished. Upon detailed comparison and distinction of specimens from the China Digital Herbarium (CVH) (https://www.cvh.ac.cn/, accessed on 1 March 2024), it was observed that the morphological characteristics of P. tremula leaves differed from those of our Leuce tree. The margin of P. tremula leaves exhibited shallow and dense serrations, whereas the margin of our Leuce samples’ leaves showed deeply and sparsely serrated features, mostly resembling a palmate shape. Consequently, the Leuce samples were consistent with the characteristics of P. alba var. pyramidalis, leading to the classification of the host as P. alba var. pyramidalis.
This information suggests that H. bachu is associated with specific species of Populus trees, providing further insight into its ecological preferences and distribution.
The species in bold indicate the sequences were obtained from this study, while the others were downloaded from GenBank. The right column lists the GenBank accession, and the middle column shows the specimen number as indicated in Table 1 or obtained from GenBank. The species in blue is the outgroup.
4. Discussion
While certain species within the Helvella genus have been confirmed to be ectomycorrhizal fungi, the ecological roles of many other species have become a subject of controversy. These particular species have alternatively been considered as either saprotrophic or mycorrhizal [10]. Here, by combining the field habitat of H. bachu, along with macro- and micro-anatomical characteristics of the plant root tip, and molecular information of the fruiting body, mycorrhiza, and host plant, we have confirmed that H. bachu forms an ectomycorrhizal relationship with Populus.
4.1. Characterization of Helvella bachu Mycorrhiza
It has been reported that Pezizalean ECMs are primarily characterized by a pseudoparenchymatous mantle, a well-developed Hartig net, and infrequent, thick, stout, and thick-walled emanating hyphae [9]. In this study, mycorrhizal samples of H. bachu collected in Maigeti County were analyzed, with 20 mycorrhizal samples representing approximately 10 different forms selected for molecular identification. This process confirmed that the H. bachu mycorrhiza exhibited a light earth color and featured a curved smooth cylindrical shape. Additionally, a thick mycelial mantle was observed, with the presence of a Hartig net and a small amount of epitaxy mycelia. Apart from the mycelium of ascomycetes H. bachu lacking a clamp connection, the mycorrhiza of H. bachu lacked rhizomorphs and sclerotia. These findings were consistent with the morphological structure of ectomycorrhiza from the Pezizomycetes of Ascomycotina, as reported by Tedersoo et al. [9] and the insights presented by Brundrett et al. [22]. It was reported that the mycorrhiza of Pezizaceae has no branching, dichotomous, or coralliform branches [9], which is consistent with our results. However, mycorrhizae of some Lactarius species predominantly exhibited a bifurcated structure [23,24].
4.2. Morphologic Observation of Mycorrhiza
When observing the morphology of mycorrhiza, sample pretreatment and sectioning techniques play a crucial role. The mycorrhiza was found to detach easily from the root tips, making detection challenging. Therefore, it is essential to handle the root tips with great care and gentleness. Various sectioning techniques were attempted, including frozen, paraffin, and freehand methods. The freehand section proved to be the most unstable, while frozen sectioning for plant tissue required exploration of corresponding fixatives due to the tissue’s tendency to break. Ultimately, the paraffin section technique yielded remarkable results. During the slicing process, efforts were made to cut the mycorrhizal slice as thinly as possible, targeting slices within 4 μm.
Out of twenty mycorrhizal samples collected from windbreak poplars of Maigeti county and Kashi City, only three were identified as H. bachu through ITS sequence analysis. Despite the scarcity of other mushrooms during the H. bachu season, diverse mycorrhizae, including Inocybe vulpinella and species of Pyronemataceae, were found while identifying those root tips. Undoubtedly, this situation significantly complicates the task of identifying the specific mycorrhiza associated with H. bachu.
4.3. Identification of Helvella bachu Host Plants
H. bachu mushrooms were confirmed to form ectomycorrhizal associations with species of Populus, a genus of trees with economic and ecological importance. Traditionally, species in the genus Populus have been classified into the following six sections based on their morphological traits and crossability: Abaso, Leuce (Populus), Aigeiros, Tacamahaca, Leucoides, and Turanga [25]. In the present study, the hosts of H. bachu were initially identified as three factions, namely section Turanga (Populus euphratica and P. pruinosa), section Aigeiros (P. nigra), and section Leuce (Populus alba var. pyramidalis) using nuclear gene ITS2 sequence (Figure 5).
Populus alba, commonly known as the white poplar, is an ecologically and economically important species widely distributed and cultivated in Xinjiang. Populus alba var. pyramidalis represents one variety of this species. Moreover, natural populations of the P. alba species frequently hybridize with other closely related species, resulting in numerous natural hybrids [26]. It remains to be determined whether P. alba, its other varieties, and the natural hybrids serve as hosts for H. bachu. During our field investigation, a local farmer informed us that H. bachu was observed in a grafting forest of P. alba on P. euphratica, although we did not find any mushrooms. While we have confirmed four kinds of host plants, it is plausible that the host of H. bachu includes more than just these four species, necessitating broader sampling for a comprehensive understanding.
In 2018, Cristina has synthesized H. cf. lacunosa mycorrhizae on Pinus ayacahuite [12]. Based on this results, we can conduct in-depth research on mycorrhizal seedling synthesis of H. bachu on Populus. However, this process involves infection using pure culture mycelia or sterile spore solution, which could take from 3-5 months to as long as 2-5 years.
4.4. Establishment of a Conservation Area and In Situ Propagation of Helvella bachu
The life cycle of H. bachu depends on the involvement of plants. Due to the challenging nature of pure culture and artificial cultivation, establishing a conservation area and in situ propagation of H. bachu appears to be a practical strategy.
We have established a conservation area within the young P. euphratica forest at Tarim University. After two years of management, there was a 14.75% increase in the number of fruiting bodies during the second year. Other researchers attempted to inoculate with soil containing the mycelia, ascospore suspension, and tissues of Helvella in the field. Their efforts resulted in a 100% survival rate, with the fruiting body density reaching 3.4 times higher than that observed in the natural area [27].
Furthermore, promising outcomes have been achieved in the semi-artificial simulated cultivation of highly desirable ectomycorrhizal fungi such as Tuber, Tricholoma, Lactarius, Chanterellus, and Rhizopogon [28,29,30,31,32,33]. These successes provide a strong foundation for similar technologies for H. bachu.
The Food and Agriculture Organization of the United Nations’ Forest Tree Genetic Resources Panel confirmed P. euphratica as a core protected tree in arid and semi-arid areas [8]. Regenerating and protecting the damaged desert riparian ecosystem, which features P. euphratica as a significant component, has become a paramount concern for the Chinese government. Given that H. bachu mushrooms form ECM and mutually beneficial symbioses with Populus tree species, they are crucial organisms in nutrient and carbon cycles in forest ecosystems, particularly in the P. euphratica forest. H. bachu may potentially promote the growth of Populus, enhance the drought tolerance of Populus, and aid Populus in withstanding the arid desert climate, akin to other ectomycorrhizal fungi. Implementing appropriate mycorrhizal propagation technology to expand Populus planting in desert hinterlands could not only reduce desertification but also increase the occurrence of H. bachu, thereby fostering economic development and ecological environment protection in southern Xinjiang under mutually beneficial conditions.
5. Conclusions
In summary, we have identified the mycorrhiza of H. bachu and confirmed it to be an ectomycorrhizal fungus. This determination was based on research into the occurrence, ITS sequences, and macro- and micro-anatomical characteristics of both the fruiting bodies and mycorrhizae of H. bachu. The host plants consisted of four Populus species, including P. euphratica, P. pruinosa, P. nigra, and P. alba var. pyramidalis (synonym P. bolleana). The establishment of a conservation area and in situ propagation of H. bachu hold significant economic and ecological importance.
Author Contributions
Investigation, C.W., M.L., L.Z., J.H. and C.D.; resources, C.D. and L.Z.; data curation, C.W.; writing—original draft preparation, C.W.; writing—review and editing, C.W. and C.D.; visualization, C.W., M.L. and C.D.; supervision, C.D. and L.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Ministry of Science and Technology of the People’s Republic of China under grant number 2022xjkk020604.
Data Availability Statement
Data are contained within the article.
Acknowledgments
Special thanks to Guo, L.D.; Yu, F.Q.; Ren, A.; and Shi, L. for technical support, and Jin, Z.S.; Kong, J.X.; Feng, B.B; and Zhang, D.D. for materials and experiment support.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Zhao, Q.; Sulayman, M.; Zhu, X.T.; Zhao, Y.C.; Yang, Z.L.; Hyde, K.D. Species clarification of the culinary Bachu mushroom in western China. Mycologia 2016, 108, 828–836. [Google Scholar] [CrossRef]
- Badshah, H.; Ali, B.; Khan, S.M.; Chaudhari, M.I.; Nisa, S.U.; Badshah, L.; Mumtaz, A.S. The in vitro antibacterial and antioxidant potentials of Helvetia bachu (Ascomycetes): A newly recorded species from Pakistan. Int. J. Med. Mushrooms 2020, 22, 1011–1020. [Google Scholar] [CrossRef]
- Al-Rawi, J.M.; Abdul-Hadi, S.Y. First new record and molecular identification of Helvella bachu (Ascomycetes) isolated from Iraq/Mosul. Int. J. Res. Appl. Sci. Biotechnol. 2022, 9, 143–148. [Google Scholar]
- Xu, J.Y.; Hu, G.H.; Gao, X.H. Resources and research of wild Helvella leucopus in Xinjiang. Chin. Wild Plant Resour. 2003, 22, 25–26. (In Chinese) [Google Scholar]
- Meng, Q.L.; Zhang, P.P.; Hu, J.W. Effects of immunoregulatory activity of polysaccharide from Bachu-mushroom. China Poult. 2005, 9, 118–120. (In Chinese) [Google Scholar]
- Zeng, D.; Zhu, S. Purification, characterization, antioxidant and anticancer activities of novel polysaccharides extracted from Bachu mushroom. Int. J. Biol. Macromol. 2018, 107, 1086–1092. [Google Scholar] [CrossRef]
- Ge, Y.; Qiu, H.; Zheng, J. Physicochemical characteristics and anti-hyperlipidemic effect of polysaccharide from Bachu mushroom (Helvella leucopus). Food Chem. X. 2022, 15, 100443. [Google Scholar] [CrossRef]
- Ling, H.; Zhang, P.; Xu, H.; Zhao, X.F. How to regenerate and protect desert riparian Populus euphratica forest in arid areas. Sci. Rep. 2015, 5, 15418. [Google Scholar] [CrossRef]
- Tedersoo, L.; Hansen, K.; Perry, B.A.; Kjøller, R. Molecular and morphological diversity of Pezizalean ectomycorrhiza. New Phytol. 2006, 170, 581–596. [Google Scholar] [CrossRef]
- Hwang, J.; Zhao, Q.; Yang, Z.L.; Wang, Z.; Townsend, J.P. Solving the ecological puzzle of mycorrhizal associations using data from annotated collections and environmental samples-an example of saddle fungi. Environ. Microbiol Rep. 2015, 7, 658–667. [Google Scholar] [CrossRef]
- Weidemann, H.N. Påvisning av Helvella Ektomykorrhiza hos Dryas og Salix ved Hjelp av Taxonselektive nrDNA Baserte Helvella Primere. Master’s Thesis, University of Oslo, Oslo, Norway, 1998. [Google Scholar]
- Cristina, A.L.; Jesús, P.M.; David, E.V.; Juan, J.A.S.; Hilda, S.R.; Adriana, D.A. Ectomycorrhizal inoculation with edible fungi increases plant growth and nutrient contents of Pinus ayacahuite. Rev. Mex. Biodivers. 2018, 89, 1089–1099. [Google Scholar] [CrossRef]
- Wang, Q.; He, X.H.; Guo, L.D. Ectomycorrhizal fungus communities of Quercus liaotungensis Koidz of different ages in a northern China temperate forest. Mycorrhiza 2012, 22, 461–470. [Google Scholar] [CrossRef]
- Dell, B.; Malajczuk, N.; Grove, T.S.; Thomson, G. Ectomycorrhiza formation in Eucalyptus.: IV. Ectomycorrhizas in the sporocarps of the hypogeous fungi Mesophellia and Castorium in Eucalypt forests of western Australia. New Phytol. 1990, 114, 449–456. [Google Scholar] [CrossRef]
- Norris, J.R.; Read, D.J.; Varma, A.K. Techniques for the Study of Mycorrhiza, 1st ed.; Academic Press: London, UK, 1991; pp. 25–73. [Google Scholar]
- Agerer, R. Exploration types of ectomycorrhizae. Mycorrhiza 2001, 11, 107–114. [Google Scholar] [CrossRef]
- Meier, S. Quality versus quantity: Optimizing evaluation of ectomycorrhizae for plants under stress. Env. Pollut. 1991, 73, 205–216. [Google Scholar] [CrossRef]
- Healy, R.A.; Zurier, H.; Bonito, G.; Smith, M.E.; Pfister, D.H. Mycorrhizal detection of native and non-native truffles in a historic arboretum and the discovery of a new North American species, Tuber arnoldianum sp. nov. Mycorrhiza 2016, 26, 781–792. [Google Scholar] [CrossRef]
- Ren, W.C.; Xu, J.; Sun, W.; Liu, M.Q.; Yu, X.X.; Wang, S.J.; Ma, W. Phylogenetic analysis of DNA barcoding Sequences in the genus Salix. Bull. Bot. Res. 2021, 41, 896–903. [Google Scholar]
- Corpet, F. Multiple sequence alignment with hierarchical clustering. Nucl. Acids Res. 1988, 16, 10881–10890. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol Biol Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Brundrett, M.; Bougher, N.; Dell, B.; Grove, T.; Malajczuk, N. Working with mycorrhizas in forestry and agriculture, 1st ed.; Pirie Printers: Canberra, Australia, 1996; Volume 32, pp. 34–35. [Google Scholar]
- Wang, R.; Guerin-Laguette, A.; Huang, L.L.; Wang, X.H.; Butler, R.; Wang, Y.; Yu, F.Q. Mycorrhizal syntheses between Lactarius spp. section Deliciosi and Pinus spp. and effects of grazing insects in Yunnan, China. Can. J. For. Res. 2019, 49, 616–627. [Google Scholar] [CrossRef]
- Wang, D.; Hallian, R.; He, X.L.; Xiong, W.P.; Peng, W.H. Morphological observation of synthesized ectomycorrhizae betwecen Lactarius deliciosus and three Pinus species and molecular identification of contaminating species. Acta Edulis Fungi 2021, 28, 129–134. (In Chinese) [Google Scholar]
- Stettler, R.F.; Bradshaw, H.D.; Heilman, P.E.; Hinckler, T.M. Biology of Populus and Its Implications for Management and Conservation, 1st ed.; NRC Research Press: Ottawa, ON, Canada, 1996; pp. 7–30. [Google Scholar]
- Lexer, C.; Fay, M.F.; Joseph, J.A.; Nica, M.; Heinze, B. Barrier to gene flow between two ecologically divergent Populus species, P. alba (white poplar) and P. tremula (European aspen): The role of ecology and life history in gene introgression. Mol. Ecol. 2005, 14, 1045–1157. [Google Scholar] [CrossRef]
- Lu, T.P.; Xu, J.Y.; Qiao, K.L.K.N. Propagation rule and effect of artificial propagation of Helvella leucopus. Xinjiang Agric. Sci. 1995, 6, 270–272. (In Chinese) [Google Scholar]
- Danell, E.; Camacho, F.J. Successful cultivation of the golden chanterelle. Nature 1997, 385, 303. [Google Scholar] [CrossRef]
- Wang, Y.; Cummings, N.; Guerin-Laguette, A. Cultivation of basidiomycete edible ectomycorrhizal mushrooms: Tricholoma, Lactarius and Rhizopogon. In Edible Ectomycorrhizal Mushrooms; Zambonelli, A., Bonito, G., Eds.; Springer: Berlin, Germany, 2012; Volume 34, pp. 281–304. [Google Scholar] [CrossRef]
- Jesús, P.M.; Alexis, G.L.; Andrea, C.R.; Fuqiang, Y.; Annemieke, V.; Faustino, H.S.; Magdalena, M.R. Edible mycorrhizal fungi of the world, forest sustainability, food security, biocultural conservation and climate change. Planets People Planet 2021, 1–20. [Google Scholar]
- Guerin-Laguette, A. Successes and challenges in the sustainable cultivation of edible mycorrhizal fungi-furthering the dream. Mycoscience 2021, 62, 10–28. [Google Scholar] [CrossRef]
- Wang, R.; Wang, Y.; Guerin-Laguette, A.; Zhang, P.; Colinas, C.; Yu, F. Factors influencing successful establishment of exotic Pinus radiata seedlings with co-introduced Lactarius deliciosus or local ectomycorrhizal fungal communities. Front Microbiol. 2022, 13, 973483. [Google Scholar] [CrossRef]
- Yamada, A. Cultivation studies of edible ectomycorrhizal mushrooms: Successful establishment of ectomycorrhizal associations in vitro and efficient production of fruiting bodies. Mycoscience 2022, 63, 235–246. [Google Scholar] [CrossRef]
Figure 1.
Habitats and market of Helvella bachu. Bar: 1 cm. (A,B) fruiting body of H. bachu; (C) Populus euphratica forest; (D-F) H. bachu and P. euphratica indicated by red and blue arrows, respectively; (G) Medicago sativa L. and H. bachu; (H,I) H. bachu traded in the market.
Figure 1.
Habitats and market of Helvella bachu. Bar: 1 cm. (A,B) fruiting body of H. bachu; (C) Populus euphratica forest; (D-F) H. bachu and P. euphratica indicated by red and blue arrows, respectively; (G) Medicago sativa L. and H. bachu; (H,I) H. bachu traded in the market.

Figure 2.
The distribution of Helvella bachu and Populus euphratica in the quadrat in 2022. Different letters indicated the different P. euphratica.
Figure 2.
The distribution of Helvella bachu and Populus euphratica in the quadrat in 2022. Different letters indicated the different P. euphratica.

Figure 3.
Molecular phylogenetic analysis of Helvella bachu and mycorrhiza by maximum likelihood method. The species in red and purple indicated the sequences from fruiting body and mycorrhiza respectively which were obtained from this study, while the others were downloaded from GenBank.
Figure 3.
Molecular phylogenetic analysis of Helvella bachu and mycorrhiza by maximum likelihood method. The species in red and purple indicated the sequences from fruiting body and mycorrhiza respectively which were obtained from this study, while the others were downloaded from GenBank.

Figure 4.
Macro- and micro-morphological characteristics of the mycorrhiza of Helvella bachu. (A–D) Macrograph of H. bachu mycorrhizae; (E,F) transverse structure of H. bachu mycorrhizae with Hartig net (E) and mantle (F). Scale: (A): 1 mm; (B–D): 500 μm; (E,F): 30 μm.
Figure 4.
Macro- and micro-morphological characteristics of the mycorrhiza of Helvella bachu. (A–D) Macrograph of H. bachu mycorrhizae; (E,F) transverse structure of H. bachu mycorrhizae with Hartig net (E) and mantle (F). Scale: (A): 1 mm; (B–D): 500 μm; (E,F): 30 μm.

Figure 5.
Molecular phylogenetic analysis of host plants of Helvella bachu by maximum likelihood method.
Figure 5.
Molecular phylogenetic analysis of host plants of Helvella bachu by maximum likelihood method.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Collection of fruiting bodies of H. bachu and host plants.
| ID | Location | Habitat | Host Specimens | Soil Texture | Longitude and Latitude |
|---|---|---|---|---|---|
| A | Conservation Land, Tarim Uni., Aral | Lawn and Populus | HB-ALE-TUR-H9 | Sandy loam | N: 40.323320 E: 81.18219 |
| B | North Gate, Tarim Uni., Aral | Poplar forest | HB-ALE-TUN-H10 | Sandy loam | N: 40.5488299 E: 81.2929233 |
| C | 6th Company, 11th Regiment, Aral | Mixed windbreak | HB-ALE-1106-H1 HB-ALE-1106-H5 | Sandy soil | N: 40.35179 E: 81.412649 |
| D | 4th Battalion, 11th Regiment, off State Route 217, Aral | Populus | HB-ALE-1104-H2 | Sandy soil | N: 40.35179 E: 81.412649 |
| E | Entrance to 217 Route, 11th Regiment, Aral | Populus | HB-ALE-11217-H6 | Sandy soil | N: 40.35179 E: 81.412649 |
| F | Tarim Bridge head, Next to the 12th Regiment, Aral | Populus | HB-ALE-1200-H11 | Sandy soil | N: 40.323320 E: 81.18219 |
| G | Beside the road of 5th Company, 13th Regiment, Aral | Populus | HB-ALE-1305-H3 | Sandy soil | N: 40.401973 E: 81.295430 |
| H | Kokochal Village, 14th Regiment, Aral | Populus | HB-ALE-1400-H4 | Sandy loam | N: 40.6437333 E: 81.8031466 |
| I | Alongside the road of 14th Regiment, Aral | Mixed windbreak | HB-ALE-1405-H7 HB-ALE-1405-H12 | Sandy soil | N: 40.384379 E: 81.482014 |
| J | 217 Route Junction between 14th regiment and 10th regiment 7, Aral | Mixed windbreak | HB-ALE-1410-H8 HB-ALE-1410-H13 | Sandy soil | N: 40.384379 E: 81.482014 |
| K | Xiamale Forest Farm, Bachu county, Kashi | Populus | - | Sandy soil | N: 39.5768316 E: 79.4363783 |
| L | Humudan village, Bazhajiemi Town, Maigaiti County, Kashi | Populus | - | Sandy soil | N: 38.8943850 E: 77.6115016 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wei, C.; Liu, M.; Hu, J.; Zhang, L.; Dong, C. Mycorrhizal Associations between Helvella bachu and Its Host Plants. Forests 2024, 15, 721. https://doi.org/10.3390/f15040721
AMA Style
Wei C, Liu M, Hu J, Zhang L, Dong C. Mycorrhizal Associations between Helvella bachu and Its Host Plants. Forests. 2024; 15(4):721. https://doi.org/10.3390/f15040721
Chicago/Turabian StyleWei, Caihong, Mengqian Liu, Jianwei Hu, Lili Zhang, and Caihong Dong. 2024. "Mycorrhizal Associations between Helvella bachu and Its Host Plants" Forests 15, no. 4: 721. https://doi.org/10.3390/f15040721
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.