A Study of the Effects of Stimulants on Resin Yield, Resin Duct and Turpentine Chemical Composition in Pinus kesiya var. langbianensis

Abstract
:1. Introduction
2. Materials and Methods
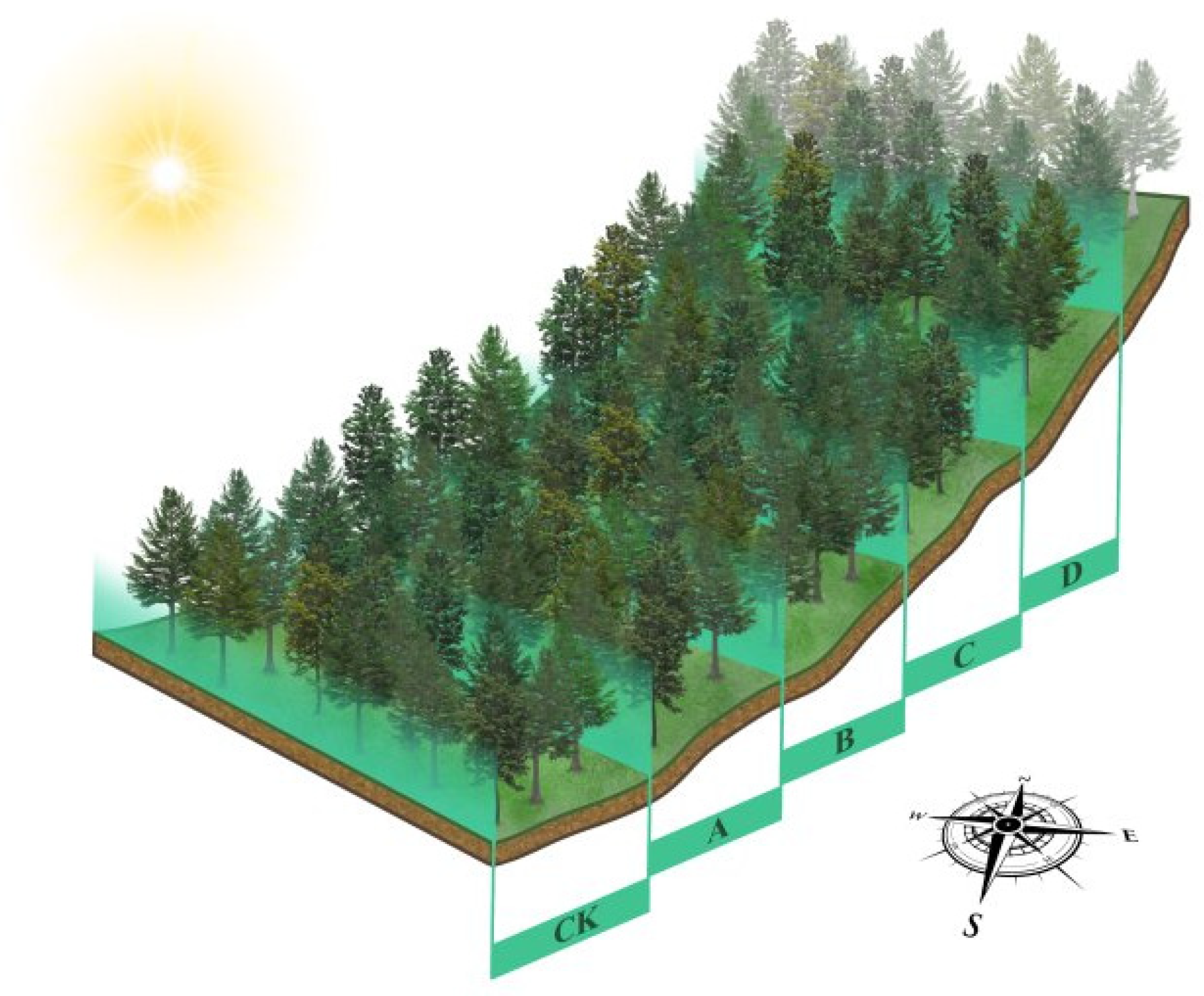
2.1. Study Area and Selection of Sample Trees
2.2. Stimulant Application Methods
2.3. Tapping Method
2.4. Analysis of Resin Yield Growth Rate
2.5. Analysis of Resin Duct Anatomy
- (1)
- After the final resin collection, anatomical samples were taken from each tree. The sampling location was approximately 2 cm below the wound, where a 2 × 2 × 2 cm sample block was obtained and immediately placed in FAA (10 mL of 37% formaldehyde, 5 mL of glacial acetic acid, and 85 mL of a 50% ethanol–water solution) solution for fixation.
- (2)
- Sectioning procedure: Utilizing the sliding microtome (Leica 2000R, Nussloch, Germany) cut cross sections (XS) and tangential sections (TS) of the adequately softened samples to a thickness of approximately 8–10 µm. Prepare at least 3 slices for each section type as spares.
- (3)
- Safranin and fast green staining: Place the sections in 1% safranin O solution for staining for 12 h or more. Afterward, sequentially transfer the stained sections into 50% ethanol solution, 75% ethanol solution, 95% ethanol solution, 1% fast green solution, 95% ethanol solution, absolute ethanol, absolute ethanol, a mixture of ethanol and xylene in a 1:1 ratio, xylene, xylene. Immerse the sections in 50% ethanol solution for 10 s, followed by 15 s in 1% fast green solution, and 3 min in all other solutions. Finally, place the processed sections on glass slides, add a suitable amount of neutral gum, gently place a coverslip on top while pushing out air bubbles from underneath, and let it dry at room temperature to complete the permanent section-making process.
- (4)
- Image J software (ImageJ-win64) was used to measure the number of resin ducts within a 6 mm2 area in 30 different fields of view for each sample. Additionally, the area of 30 axial resin ducts was measured; there were 15 trees in each group, so we measured 450 data per group, and their average values were calculated.
2.6. Resin Composition Analysis
2.7. Data Analysis
3. Results
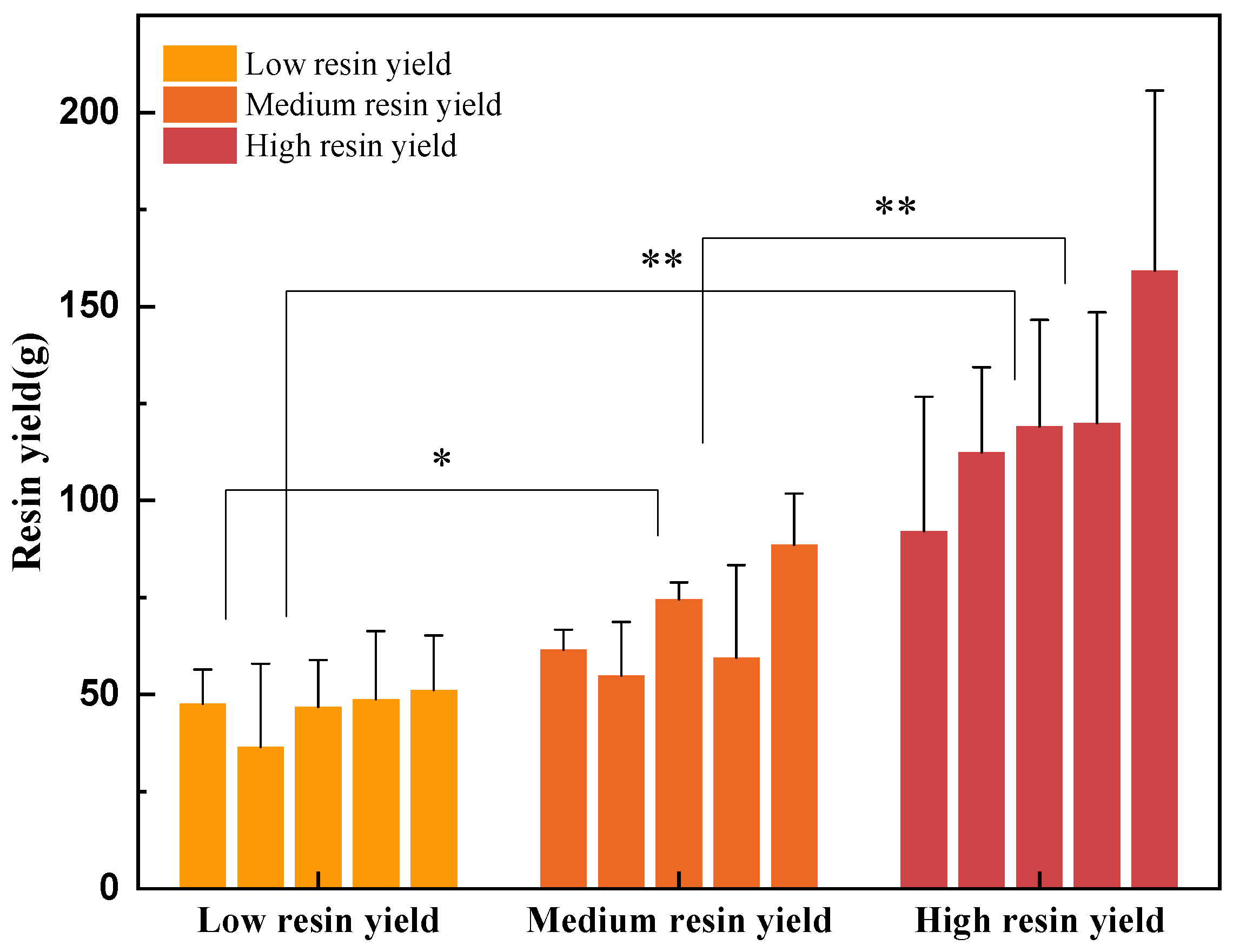
3.1. The Impact of Different Concentrations of Stimulants on Resin Yield
3.2. Anatomical Structure Analysis
3.2.1. Effects of Different Concentration Stimulant on Anatomical Structure
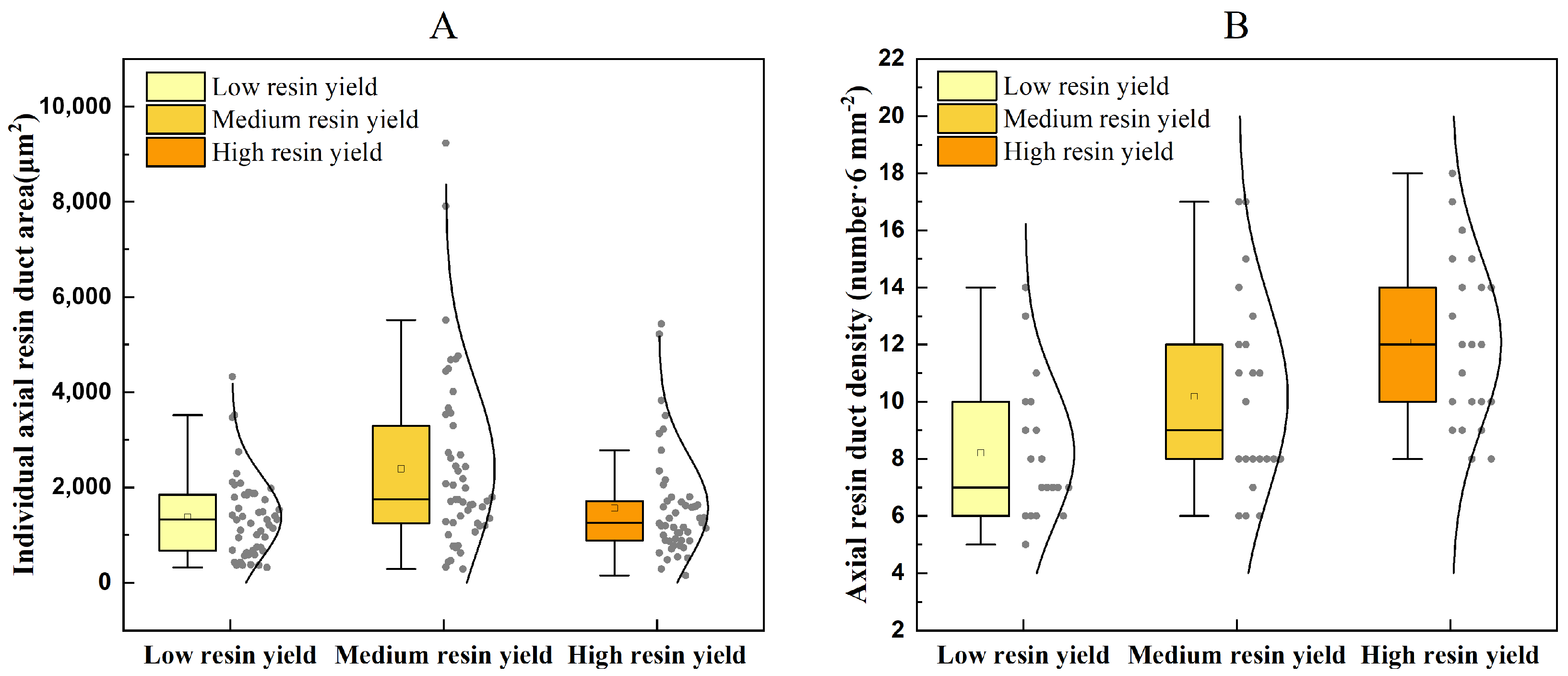
3.2.2. Analysis of Resin Duct Variations in Different Resin Yield Levels of P. kesiya var. langbianensis
3.2.3. Analysis of the Correlation between Resin Yield and Anatomical Structure
3.3. Analysis of Pine Turpentine Chemical Components
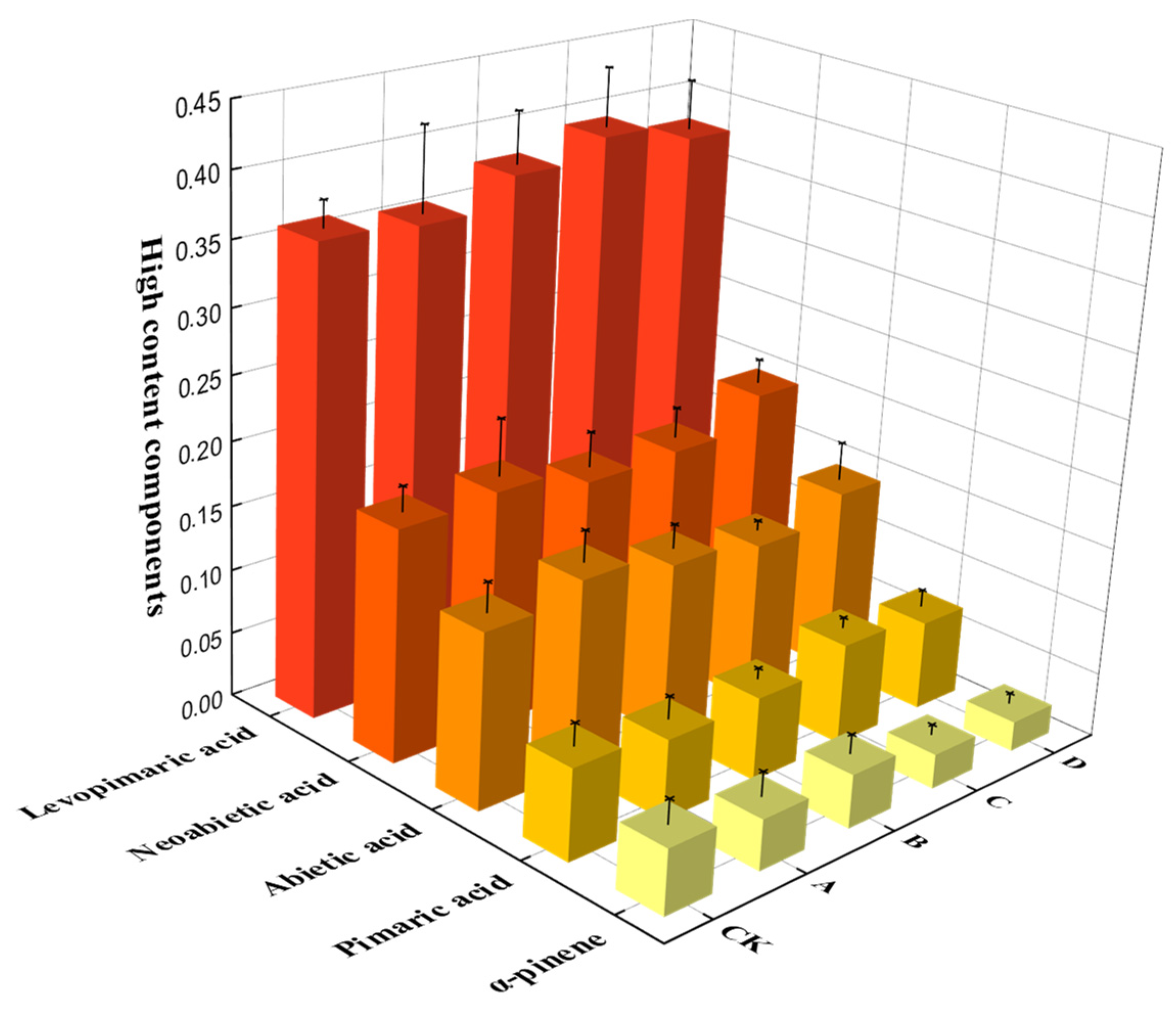
3.3.1. Analysis of the Chemical Components of Turpentine Stimulated by Different Concentrations of Stimulant
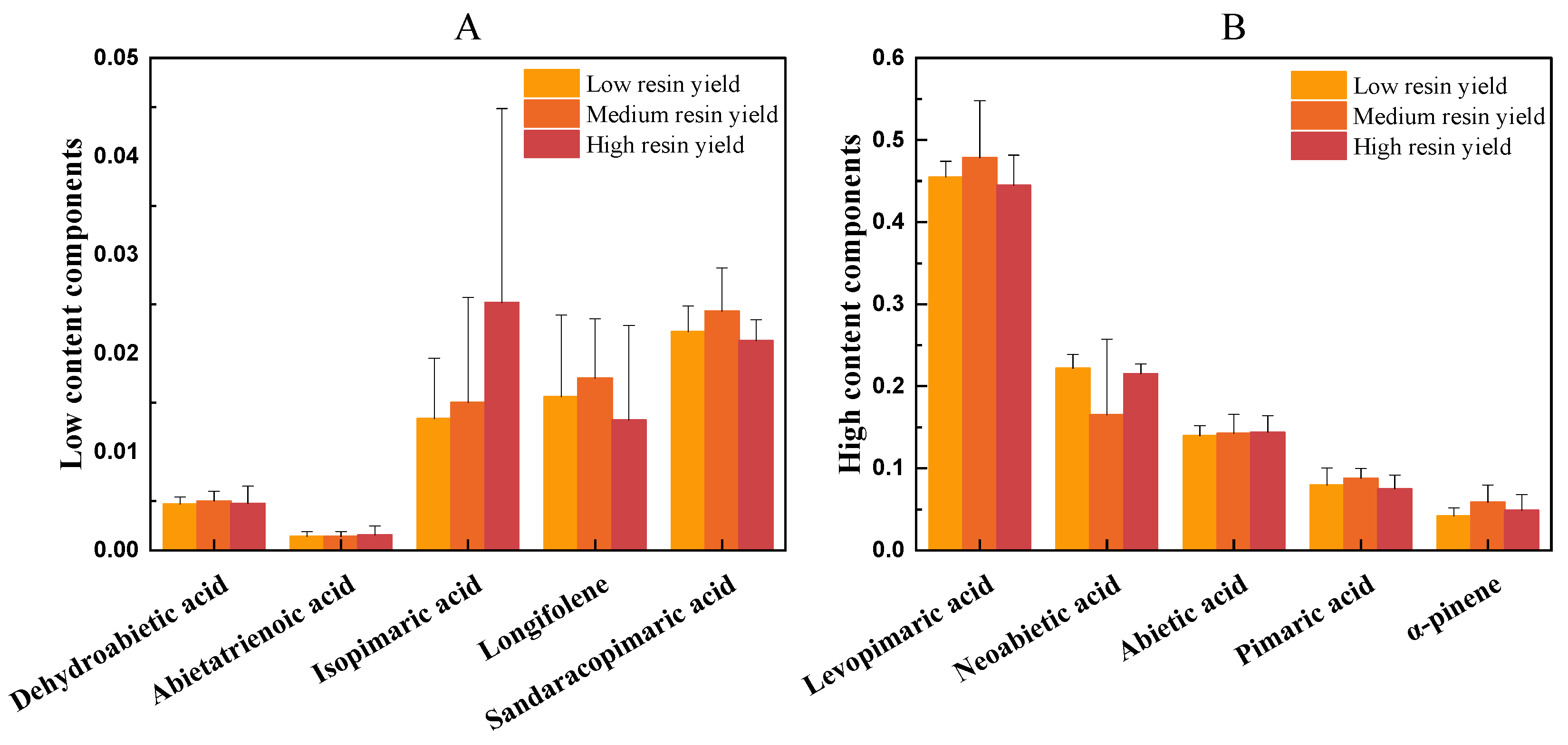
3.3.2. Analysis of Differences in Chemical Composition of P. kesiya var. langbianensis with Different Resin Yield
3.4. Correlation Analysis of Resin Yield, Anatomical Structure and Turpentine Chemical Composition
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thomte, L.; Shah, S.K.; Mehrotra, N.; Saikia, A.; Bhagabati, A.K. Dendrochronology in the tropics using tree-rings of Pinus kesiya. Dendrochronologia 2023, 78, 126070. [Google Scholar] [CrossRef]
- Weng, H.; Chen, H.; Duan, A. Selection of high-yielding fat superior trees of Simao pine. J. Northwest For. Coll. 2010, 25, 71–74+79. (In Chinese) [Google Scholar]
- Xu, M.; Dang, G.; Ling, W. Exploration of the utilisation method of Simao pine. Seed 2012, 31, 95–96, 101. (In Chinese) [Google Scholar]
- Chen, W.; Chen, S.; Luo, T.; Li, J. Principal component analysis of ecological factors related to lipid production of Simao pine. West. For. Sci. 2023, 52, 27–33. (In Chinese) [Google Scholar]
- Dong, J.; Guo, H.; Zhang, Z.; Wu, X.; Huang, F. Dynamic changes of China’s turpentine resources in the last 10 years—A dynamic assessment based on “Lianqing” data. For. Sci. 2016, 52, 112–122. (In Chinese) [Google Scholar]
- Cabrita, P. A model for resin flow. In Plant Cell and Tissue Differentiation and Secondary Metabolites: Fundamentals and Applications; Springer: Cham, Switzerland, 2021; pp. 117–144. [Google Scholar] [CrossRef]
- Franceschi, V.R.; Krokene, P.; Christiansen, E.; Krekling, T. Anatomical and chemical defenses of conifer bark against bark beetles and other pests. New Phytol. 2005, 167, 353–376. [Google Scholar] [CrossRef] [PubMed]
- Fuentealba, A.; Alfaro, R.; Bauce, É. Theoretical framework for assessment of risks posed to Canadian forests by invasive insect species. For. Ecol. Manag. 2013, 302, 97–106. [Google Scholar] [CrossRef]
- Esteban, L.G.; Martín, J.A.; de Palacios, P.; Fernández, F.G. Influence of region of provenance and climate factors on wood anatomical traits of Pinus nigra Arn. subsp. salzmannii. Eur. J. For. Res. 2012, 131, 633–645. [Google Scholar] [CrossRef]
- Lai, M.; Dong, L.; Yi, M.; Sun, S.; Zhang, Y.; Fu, L.; Xu, Z.; Lei, L.; Leng, C.; Zhang, L. Genetic variation, heritability and genotype × environment interactions of resin yield, growth traits and morphologic traits for Pinus elliottii at three progeny trials. Forests 2017, 8, 409. [Google Scholar] [CrossRef]
- Lombardero, M.J.; Ayres, M.P.; Lorio, P.L., Jr.; Ruel, J. Environmental effects on constitutive and inducible resin defences of Pinus taeda. Ecol. Lett. 2000, 3, 329–339. [Google Scholar] [CrossRef]
- Garcia-Forner, N.; Campelo, F.; Carvalho, A.; Vieira, J.; Rodríguez-Pereiras, A.; Ribeiro, M.; Salgueiro, A.; Silva, M.E.; Louzada, J.L. Growth-defence trade-offs in tapped pines on anatomical and resin production. For. Ecol. Manag. 2021, 496, 119406. [Google Scholar] [CrossRef]
- Rodríguez-García, A.; López, R.; Martín, J.A.; Pinillos, F.; Gil, L. Resin yield in Pinus pinaster is related to tree dendrometry, stand density and tapping-induced systemic changes in xylem anatomy. For. Ecol. Manag. 2014, 313, 47–54. [Google Scholar] [CrossRef]
- Neis, F.A.; de Costa, F.; de Almeida, M.R.; Colling, L.C.; Junkes, C.F.d.O.; Fett, J.P.; Fett-Neto, A.G. Resin exudation profile, chemical composition, and secretory canal characterization in contrasting yield phenotypes of Pinus elliottii Engelm. Ind. Crops Prod. 2019, 132, 76–83. [Google Scholar] [CrossRef]
- Fabián-Plesníková, I.; Sáenz-Romero, C.; Terrazas, T.; Reyes-Ramos, A.; Martínez-Trujillo, M.; Cruz-De-León, J.; Sánchez-Vargas, N.M. Traumatic ducts size varies genetically and is positively associated to resin yield of open-pollinated progenies. Silvae Genet. 2022, 71, 10–19. [Google Scholar] [CrossRef]
- Neis, F.A.; de Costa, F.; de Araujo, A.T., Jr.; Fett, J.P.; Fett-Neto, A.G. Multiple industrial uses of non-wood pine products. Ind. Crops Prod. 2019, 130, 248–258. [Google Scholar] [CrossRef]
- Yadav, B.K.; Gidwani, B.; Vyas, A. Rosin: Recent advances and potential applications in novel drug delivery system. J. Bioact. Compat. Polym. 2016, 31, 111–126. [Google Scholar] [CrossRef]
- Liang, J.; Chen, X.; Wang, L.; Wei, X.; Mo, K.; Xiong, Z.; Liang, J. Influence of a novel resin tapping method on the resin yield and turpentine chemical composition of Masson pine (Pinus massoniana). Ind. Crops Prod. 2023, 204, 117377. [Google Scholar] [CrossRef]
- Kelkar, V.M.; Geils, B.W.; Becker, D.R.; Overby, S.T.; Neary, D.G. How to recover more value from small pine trees: Essential oils and resins. Biomass Bioenergy 2006, 30, 316–320. [Google Scholar] [CrossRef]
- Soliño, M.; Yu, T.; Alía, R.; Auñón, F.; Bravo-Oviedo, A.; Chambel, M.R.; de Miguel, J.; del Río, M.; Justes, A.; Martínez-Jauregui, M.; et al. Resin-tapped pine forests in Spain: Ecological diversity and economic valuation. Sci. Total Environ. 2018, 625, 1146–1155. [Google Scholar] [CrossRef]
- Hu, W.J.; Chen, J.; Liu, T.W.; Wu, Q.; Wang, W.-H.; Liu, X.; Shen, Z.-J.; Simon, M.; Wu, F.-H.; Pei, Z.-M.; et al. Proteome and calcium-related gene expression in Pinus massoniana needles in response to acid rain under different calcium levels. Plant Soil 2014, 380, 285–303. [Google Scholar] [CrossRef]
- Yi, M.; Jia, T.; Dong, L.; Zhang, L.; Leng, C.; Liu, S.; Lai, M. Resin yield in Pinus elliottii Engelm. is related to the resin flow rate, resin components and resin duct characteristics at three locations in southern China. Ind. Crops Prod. 2021, 160, 113141. [Google Scholar] [CrossRef]
- Wolter, K.E.; Peters, W.J.; Roberts, D.R.; McReynolds, R.D.; Broomfield, J.; Crews, E.R. Process for Increasing Oleoresin Synthesis in Pinus Species. U.S. Patent 4,203,253, 20 May 1980. [Google Scholar]
- Parham, M.R. Stimulation of oleoresin yield in conifers. Outlook Agric. 1976, 9, 76–81. [Google Scholar] [CrossRef]
- Füller, T.N.; de Lima, J.C.; de Costa, F.; Rodrigues-Corrêa, K.C.; Fett-Neto, A.G. Stimulant paste preparation and bark streak tapping technique for pine oleoresin extraction. In Biotechnology of Plant Secondary Metabolism: Methods and Protocols; Humana Press: New York, NY, USA, 2016; pp. 19–26. [Google Scholar] [CrossRef]
- Neis, F.A.; de Costa, F.; Füller, T.N.; de Lima, J.C.; Rodrigues-Corrêa, K.C.d.S.; Fett, J.P.; Fett-Neto, A.G. Biomass yield of resin in adult Pinus elliottii Engelm. trees is differentially regulated by environmental factors and biochemical effectors. Ind. Crops Prod. 2018, 118, 20–25. [Google Scholar] [CrossRef]
- Rodrigues-Corrêa, K.C.d.S.; Fett-Neto, A.G. Seasonality and chemical elicitation of defense oleoresin production in field-grown slash pine under subtropical climate. Theor. Exp. Plant Physiol. 2013, 25, 56–61. [Google Scholar] [CrossRef]
- Silverman, F.P.; Petracek, P.D.; Fledderman, C.M.; Ju, Z.; Heiman, D.F.; Warrior, P. Salicylate activity. 1. Protection of plants from paraquat injury. J. Agric. Food Chem. 2005, 53, 9764–9768. [Google Scholar] [CrossRef] [PubMed]
- López-Álvarez, Ó.; Zas, R.; Marey-Perez, M. Resin tapping: A review of the main factors modulating pine resin yield. Ind. Crops Prod. 2023, 202, 117105. [Google Scholar] [CrossRef]
- Méijome, A.G.; Chamorro, E.M.; Blanco, E.F.; García, E.G. Análisis de producciones y rendimientos del sistema de resinación de pica de corteza con estimulación química de doble cara ancha en masas de Pinus pinaster Ait. cuyo objetivo principal es la producción de madera. Recur. Rurais 2020, 16, 5–10. [Google Scholar] [CrossRef]
- García-Méijome, A.; Lorenzo, M.J.R.; Blanco, E.F.; Chamorro, E.M.; Gómez-García, E. Resin-tapping production in Pinus pinaster Ait. Stands in Galicia (NW Spain): Effects of location, number of faces, wound width and production year. Forests 2023, 14, 128. [Google Scholar] [CrossRef]
- Lin, J.; Hu, Y.; He, X.; Ceulemans, R. Systematic survey of resin canals in Pinaceae. Belg. J. Bot. 2002, 135, 3–14. [Google Scholar]
- Sood, Y.; Mahajan, P.K.; Bharti; Gupta, R.K. Correlation and Regression Studies on Estimation of Resin Yield in Pinus roxburghii. Ind. J. Pure Appl. Biosci. 2019, 7, 63–66. [Google Scholar] [CrossRef]
- An, N.; Ding, G.J. Study on chemical constituents of oleoresin from Pinus msssoniana in Guangxi. J. Cent. South Univ. For. Technol. 2012, 32, 59–62. (In Chinese) [Google Scholar]
- Blanche, C.A.; Lorio, P.L., Jr.; Sommers, R.; Hodges, J.; Nebeker, T. Seasonal cambial growth and development of loblolly pine: Xylem formation, inner bark chemistry, resin ducts, and resin flow. For. Ecol. Manag. 1992, 49, 151–165. [Google Scholar] [CrossRef]
- Lorio, P.L., Jr.; Hodges, J.D. Microsite effects on oleoresin exudation pressure of large loblolly pines. Ecology 1968, 49, 1207–1210. [Google Scholar] [CrossRef]
- Zas, R.; Quiroga, R.; Touza, R.; Vázquez-González, C.; Sampedro, L.; Lema, M. Resin tapping potential of Atlantic maritime pine forests depends on tree age and timing of tapping. Ind. Crops Prod. 2020, 157, 112940. [Google Scholar] [CrossRef]
- Rodrigues-Honda, K.C.d.S.; Junkes, C.F.d.O.; de Lima, J.C.; Waldow, V.d.A.; Rocha, F.S.; Sausen, T.L.; Bayer, C.; Talamini, E.; Fett-Neto, A.G. Carbon sequestration in resin-tapped slash pine (Pinus elliottii Engelm.) subtropical plantations. Biology 2023, 12, 324. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | DBH | Stimulant | Altitude | Samples |
|---|---|---|---|---|
| CK | 19.05 | 1326 | 15 | |
| A | 19.42 | 0.25% | 1328 | 15 |
| B | 17.99 | 0.5% | 1330 | 15 |
| C | 18.38 | 1% | 1332 | 15 |
| D | 18.26 | 2% | 1334 | 15 |
| SS | DF | MS | F | p Value | |
|---|---|---|---|---|---|
| Between groups | 1209.47 | 4 | 302.37 | 0.603 | >0.1 |
| Within Groups | 35,080.21 | 70 | 501.146 | ||
| Total | 36,289.68 | 74 |
| Groups | Individual Axial Resin Duct Area (μm2) | Axial Resin Duct Density (Number·6 mm−2) |
|---|---|---|
| CK | 1803.67 ± 1402.79 (c) | 10.31 ± 3.34 (a) |
| A | 3936.52 ± 2133.45 (a) | 9.80 ± 3.03 (a) |
| B | 3673.70 ± 2501.69 (ab) | 8.67 ± 3.46 (a) |
| C | 3382.70 ± 1750.26 (b) | 10.88 ± 2.71 (a) |
| D | 3665.11 ± 2457.92 (ab) | 11.65 ± 4.16 (a) |
| Groups | Individual Axial Resin Duct Area (μm2) | Axial Resin Duct Density (Number·6 mm−2) |
|---|---|---|
| Low resin yield | 1378.06 ± 861.54 (b) | 8.21 ± 2.46 (a) |
| Medium resin yield | 2390.91 ± 1836.06 (a) | 10.18 ± 3.40 (b) |
| High resin yield | 1568.54 ± 1109.08 (b) | 12.09 ± 2.95 (c) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, H.; Shi, J.; Chen, L.; Yang, C.; Li, C.; Huang, Y.; Qiu, J. A Study of the Effects of Stimulants on Resin Yield, Resin Duct and Turpentine Chemical Composition in Pinus kesiya var. langbianensis. Forests 2024, 15, 748. https://doi.org/10.3390/f15050748
Yang H, Shi J, Chen L, Yang C, Li C, Huang Y, Qiu J. A Study of the Effects of Stimulants on Resin Yield, Resin Duct and Turpentine Chemical Composition in Pinus kesiya var. langbianensis. Forests. 2024; 15(5):748. https://doi.org/10.3390/f15050748
Chicago/Turabian StyleYang, Huanxin, Junjie Shi, Lin Chen, Chunwang Yang, Changzhao Li, Yuxi Huang, and Jian Qiu. 2024. "A Study of the Effects of Stimulants on Resin Yield, Resin Duct and Turpentine Chemical Composition in Pinus kesiya var. langbianensis" Forests 15, no. 5: 748. https://doi.org/10.3390/f15050748