
Distribution Dynamics of Diplopanax stachyanthus Hand.-Mazz. (Mastixiaceae) and Its Implications in Relict Mastixioid Flora Conservation
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Region
2.2. Distributional Records of Species and Bioclimatic Variables
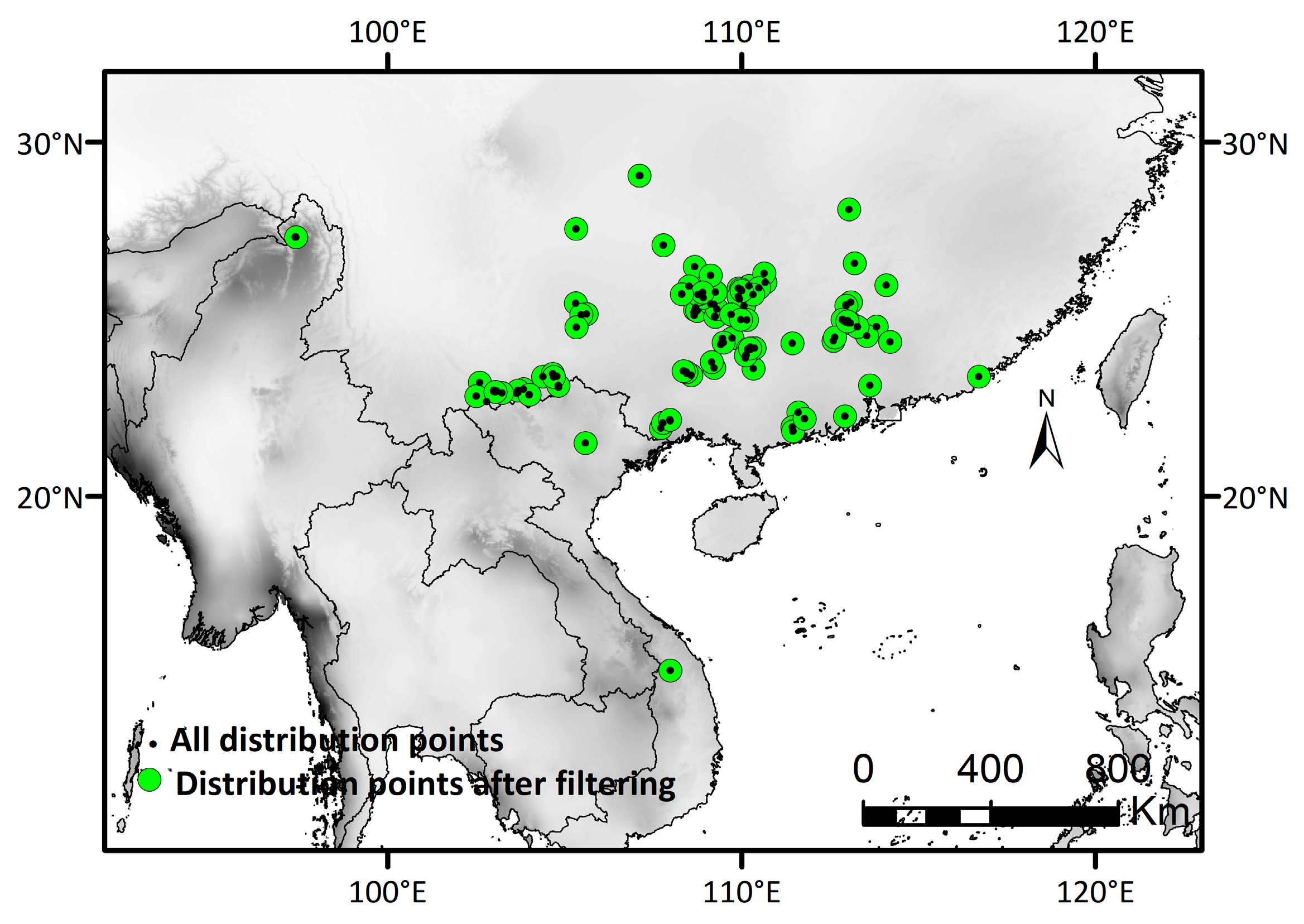
2.2.1. Occurrence Records
2.2.2. Selection of Bioclimatic Factors
2.3. Bioclimatic Variable Selection
2.4. Model Configuration and Selection
2.5. Suitability Level Classification and Spatial Changes
2.6. Centroid Shifts
3. Results
3.1. Model Performance
3.2. Environmental Variable Contribution
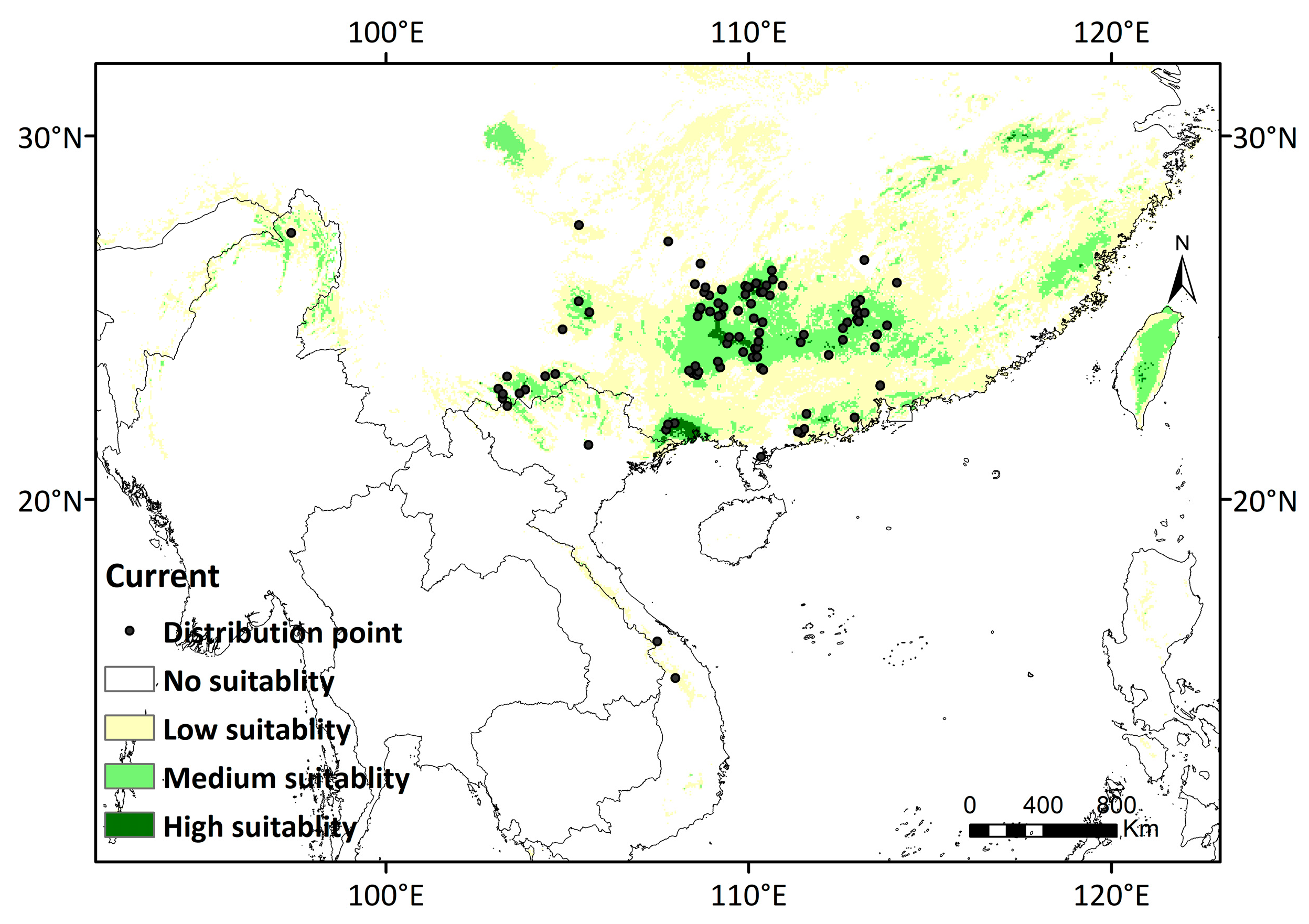
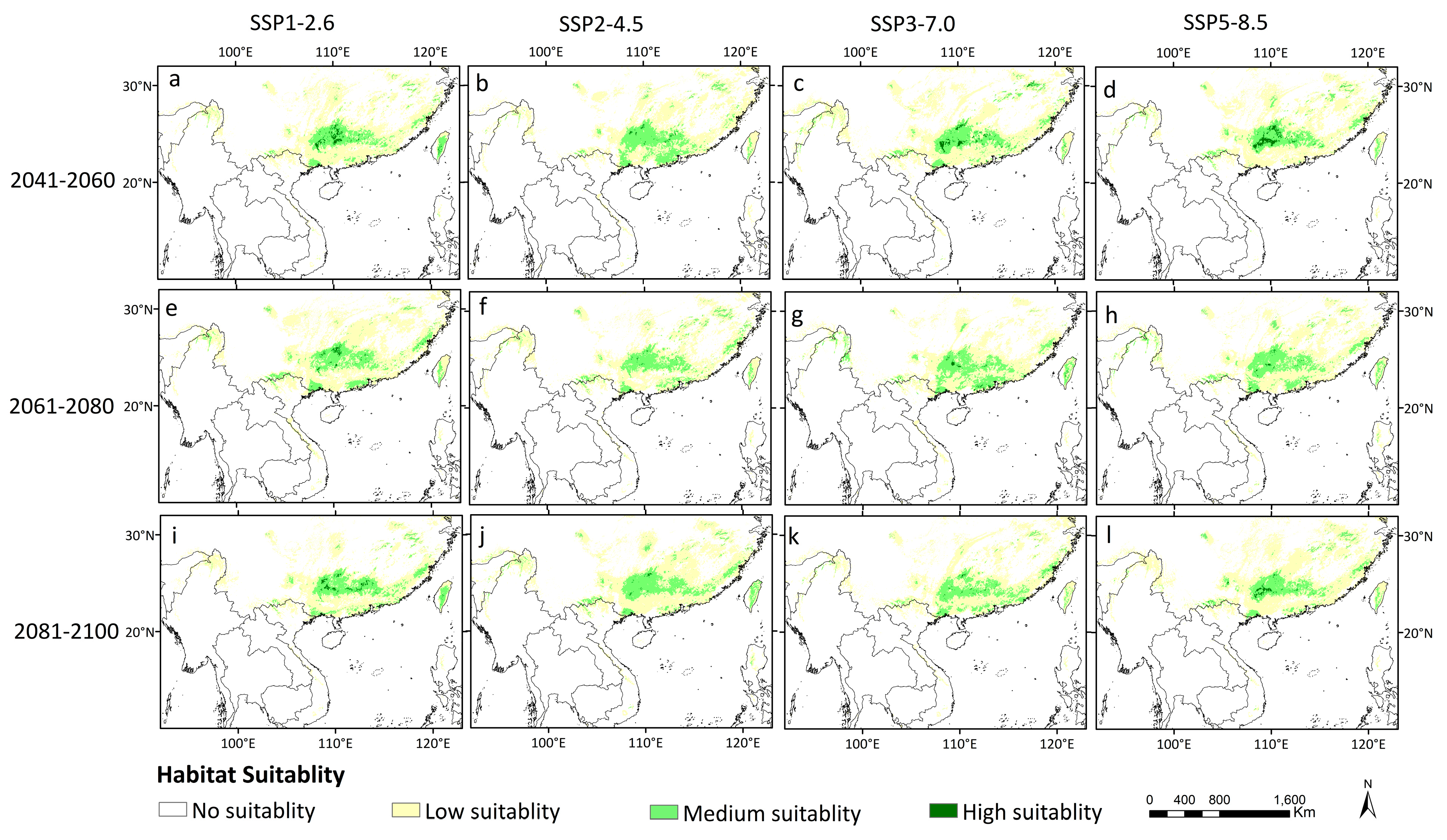
3.3. Potential Distribution of D. stachyanthus under Retrospective, Current and Prospective Climate Conditions
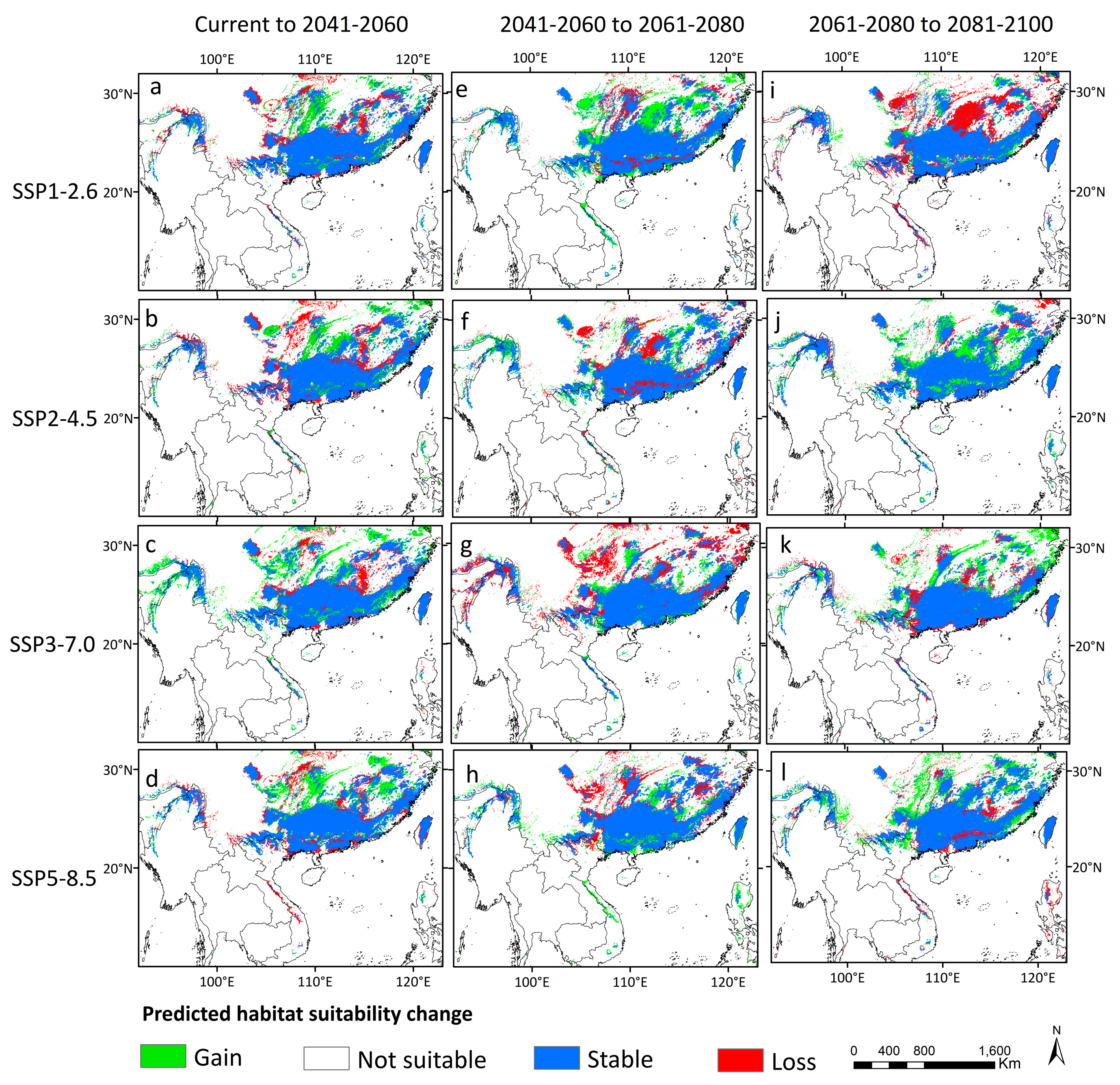
3.4. Distribution Dynamics
3.5. Centroid Shift under the Different Climatic Scenarios
4. Discussion
4.1. The Primary Climatic Factors Influencing the Distribution of D. stachyanthus
4.2. Distribution Dynamics of D. stachyanthus
4.3. Conservation and Forestry Management
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rotllan, P.X.; Traveset, A. Declining relict plants: Climate effect or seed dispersal disruption? A landscape-scale approach. Basic. Appl. Ecol. 2016, 17, 81–91. [Google Scholar] [CrossRef]
- Guo, Y.L.; Zhao, Z.F.; Li, X. Moderate warming will expand the suitable habitat of Ophiocordyceps sinensis and expand the area of O. sinensis with high adenosine content. Sci. Total Environ. 2021, 787, 147605. [Google Scholar] [CrossRef]
- Doxford, S.W.; Freckleton, R.P. Changes in the large-scale distribution of plants: Extinction, colonisation and the effects of climate. J. Ecol. 2012, 100, 519–529. [Google Scholar] [CrossRef]
- Jochum, G.M.; Mudge, K.W.; Thomas, R.B. Elevated temperatures increase leaf senescence and root secondary metabolite concentrations in the understory herb Panax quinquefolius (Araliaceae). Am. J. Bot. 2007, 94, 819–826. [Google Scholar] [CrossRef] [PubMed]
- Wei, B.; Wang, R.L.; Hou, K.; Wang, X.Y.; Wu, W. Predicting the current and future cultivation regions of Carthamus tinctorius L. using MaxEnt model under climate change in China. Glob. Ecol. Conserv. 2018, 16, e00477. [Google Scholar] [CrossRef]
- Warren, R.; Price, J.; Graham, E.; Forstenhaeusler, N.; VanDerWal, J. The projected effect on insects, vertebrates, and plants of limiting global warming to 1.5 degrees C rather than 2 degrees C. Science 2018, 360, 791–795. [Google Scholar] [CrossRef]
- Ali, H.; Din, J.U.; Bosso, L.; Hameed, S.; Kabir, M.; Younas, M.; Nawaz, M.A. Expanding or shrinking? Range shifts in wild ungulates under climate change in Pamir-Karakoram mountains, Pakistan. PLoS ONE 2021, 16, e0260031. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.B.; Xie, L.N.; Wang, H.W.; Zhong, J.B.; Li, Y.C.; Liu, J.L.; Ou, Z.; Liang, X.X.; Li, Y.S.; Huang, H.Y.; et al. Geographic distribution and impacts of climate change on the suitable habitats of Zingiber species in China. Ind. Crops Prod. 2019, 138, 111429. [Google Scholar] [CrossRef]
- Shen, Y.F.; Tu, Z.H.; Zhang, Y.L.; Zhong, W.P.; Xia, H.; Hao, Z.Y.; Zhang, C.G.; Li, H.G. Predicting the impact of climate change on the distribution of two relict Liriodendron species by coupling the MaxEnt model and actual physiological indicators in relation to stress tolerance. J. Environ. Manag. 2022, 322, 116024. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.M.; Ren, B.P.; Li, D.Y.; Garber, P.A.; Zhu, P.F.; Xiang, Z.F.; Grueter, C.C.; Liu, Z.J.; Li, M. Climate change, grazing, and collecting accelerate habitat contraction in an endangered primate. Biol. Conserv. 2019, 231, 88–97. [Google Scholar] [CrossRef]
- Tiffney, B.H. Perspectives on the origin of the floristic similarity between eastern Asia and eastern North America. J. Arnold Arbor. 1985, 66, 73–94. [Google Scholar] [CrossRef]
- Huang, Y.J.; Jacques, F.M.B.; Su, T.; Ferguson, D.K.; Tang, H.; Chen, W.Y.; Zhou, Z.K. Distribution of Cenozoic plant relicts in China explained by drought in dry season. Sci. Rep. 2015, 5, 14212. [Google Scholar] [CrossRef]
- Qiu, Y.X.; Lu, Q.X.; Zhang, Y.H.; Cao, Y.N. Phylogeography of East Asia’s Tertiary relict plants: Current progress and future prospects. Biodivers. Sci. 2017, 25, 136–146. [Google Scholar] [CrossRef]
- Milne, R.; Abbott, R. The origin and evolution of Tertiary relict flora. Adv. Bot. Res. 2002, 38, 281–314. [Google Scholar] [CrossRef]
- Zhekun, Z.; Momohara, A. Fossil history of some endemic seed plants of East Asia and its phytogeographical significance. Acta Bot. Yunnanica 2005, 27, 449–470. [Google Scholar]
- Manchester, S.R.; Chen, Z.D.; Lu, A.M.; Uemura, K. Eastern Asian endemic seed plant genera and their paleogeographic history throughout the Northern Hemisphere. J. Syst. Evol. 2009, 47, 1–42. [Google Scholar] [CrossRef]
- Eyde, R.H. Fossil record and ecology of Nyssa (Cornaceae). Bot. Rev. 1997, 63, 97–123. [Google Scholar] [CrossRef]
- Liu, Y.S.; Basinger, J.F. Fossil Cathaya (Pinaceae) pollen from the Canadian high arctic. Int. J. Plant Sci. 2000, 161, 829–847. [Google Scholar] [CrossRef]
- Heinemann, M. Warm and Sensitive Paleocene-Eocene Climate. Ph.D. Thesis, University of Hamburg, Hamburg, Spain, 2009. [Google Scholar]
- Chen, Z.L.; Ding, Z.L. A Review on the Paleocene-Eocene Thermal Maximum. Quat. Sci. 2011, 31, 937–950. [Google Scholar] [CrossRef]
- Jovane, L.G.; Coccioni, R.; Marsili, A.; Acton, G. The late Eocene greenhouse-icehouse transition: Observations from the Massignano global stratotype section and point (GSSP). Geol. Soc. Am. Spec. Pap. 2009, 452, 149–168. [Google Scholar]
- Collinson, M.E.; Hooker, J.J. Paleogene vegetation of Eurasia: Framework for mammalian faunas. Deinsea 2003, 10, 41–84. [Google Scholar]
- Ao, H.; Dupont-Nivet, G.; Rohling, E.J.; Zhang, P.; Ladant, J.B.; Roberts, A.P.; Licht, A.; Liu, Q.S.; Liu, Z.G.; Dekkers, M. Orbital climate variability on the northeastern Tibetan Plateau across the Eocene–Oligocene transition. Nat. Commun. 2020, 11, 5249. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.M.; Wang, M.Z.; Zhang, X.Q. Palynology Assemblages and Paleoclimatic Character of the Late Eocene to Early Oligocene in China. Earth Sci. 2005, 30, 309–316. [Google Scholar]
- Chen, C.; Qi, Z.C.; Xu, X.H.; Comes, H.P.; Koch, M.A.; Jin, X.J.; Fu, C.X.; Qiu, Y.X. Understanding the formation of Mediterranean–African–Asian disjunctions: Evidence for Miocene climate-driven vicariance and recent long-distance dispersal in the Tertiary relict Smilax aspera (Smilacaceae). New Phytol. 2014, 204, 243–255. [Google Scholar] [CrossRef] [PubMed]
- Nie, Z.L.; Sun, H.; Manchester, S.R.; Meng, Y.; Luke, Q.; Wen, J. Evolution of the intercontinental disjunctions in six continents in the Ampelopsis clade of the grape family (Vitaceae). BMC Evol. Biol. 2012, 12, 17. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Wagner, W.L.; Ree, R.H.; Berry, P.E.; Wen, J. Molecular phylogeny, divergence time estimates, and historical biogeography of Circaea (Onagraceae) in the Northern Hemisphere. Mol. Phylogenetics Evol. 2009, 53, 995–1009. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.Y.; Ree, R.H.; Salamin, N.; Xing, Y.W.; Silvestro, D. Fossil-informed models reveal a boreotropical origin and divergent evolutionary trajectories in the walnut family (Juglandaceae). Syst. Biol. 2022, 71, 242–258. [Google Scholar] [CrossRef] [PubMed]
- Tiffney, B.H.; Haggard, K.K. Fruits of Mastixioideae (Cornaceae) from the Paleogene of western North America. Rev. Palaeobot. Palyno 1996, 92, 29–54. [Google Scholar] [CrossRef]
- Eyde, R.H.; Xiang, Q.Y. Fossil Mastixioid (Cornaceae) alive in Eastern Asia. Am. J. Bot. 1990, 77, 689–692. [Google Scholar] [CrossRef]
- Matthew, K.M. A revision of the genus Mastixia (Cornaceae). Blumea Biodivers. Evol. Biogeogr. Plants 1976, 23, 51–93. [Google Scholar]
- He, J.; Zeng, C.J. Flora of China; Science Press: Beijing, China, 1978; Volume 54. [Google Scholar]
- Sodhi, N.S.; Posa, M.R.C.; Lee, T.M.; Bickford, D.; Koh, L.P.; Brook, B.W. The state and conservation of Southeast Asian biodiversity. Biodivers. Conserv. 2010, 19, 317–328. [Google Scholar] [CrossRef]
- Stibig, H.J.; Achard, F.; Carboni, S.; Raši, R.; Miettinen, J. Change in tropical forest cover of Southeast Asia from 1990 to 2010. Biogeosciences 2014, 11, 247–258. [Google Scholar] [CrossRef]
- Deb, J.C.; Phinn, S.; Butt, N.; McAlpine, C.A. Climate change impacts on tropical forests: Identifying risks for tropical Asia. J. Trop. For. Sci. 2018, 30, 182–194. [Google Scholar]
- Estoque, R.C.; Ooba, M.; Avitabile, V.; Hijioka, Y.; DasGupta, R.; Togawa, T.; Murayama, Y.J. The future of Southeast Asia’s forests. Nat. Commun. 2019, 10, 1829. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.Y.; Ci, X.Q.; Hu, J.L.; Bai, Y.; Thornhill, A.H.; Conran, J.G.; Li, J. Riparian areas as a conservation priority under climate change. Sci. Total Environ. 2023, 858, 159879. [Google Scholar] [CrossRef]
- Stockey, R.A.; LePage, B.A.; Pigg, K.B. Permineralized fruits of Diplopanax (Cornaceae, Mastixioideae) from the middle Eocene Princeton chert of British Columbia. Rev. Palaeobot. Palynol. 1998, 103, 223–234. [Google Scholar] [CrossRef]
- Martinetto, E. The first mastixioid fossil from Italy and its palaeobiogeographic implications. Rev. Palaeobot. Palyno 2011, 167, 222–229. [Google Scholar] [CrossRef]
- Mai, D.H. Entwicklung und klimatische Differenzierung der Laubwaldflora Mitteleuropas im Tertiär. Flora 1981, 171, 525–582. [Google Scholar] [CrossRef]
- Mai, D.H. On the extinct Mastixiaceae (Cornales) in Europe. Geophytology 1993, 23, 53–63. [Google Scholar]
- Seward, A.C. British Museum (Natural History) The London Clay Flora. Nature 1934, 134, 6–7. [Google Scholar] [CrossRef]
- Graham, A. Late Cretaceous and Cenozoic History of North American Vegetation: North of Mexico; Oxford University Press: Oxford, UK, 1999. [Google Scholar]
- Manchester, S.R. Extinct ulmaceous fruits from the Tertiary of Europe and western North America. Rev. Palaeobot. Palynol. 1987, 52, 119–129. [Google Scholar] [CrossRef]
- Manchester, S.R. Fruits and seeds of the Middle Eocene nut beds flora, Clarno Formation, Oregon. Paleoutographica Am. 1994, 58, 1–205. [Google Scholar]
- Brikiatis, L. Late Mesozoic North Atlantic land bridges. Earth-Sci. Rev. 2016, 159, 47–57. [Google Scholar] [CrossRef]
- Tiffney, B.H.; Manchester, S.R. The use of geological and paleontological evidence in evaluating plant phylogeographic hypotheses in the Northern Hemisphere tertiary. Int. J. Plant Sci. 2001, 162, S3–S17. [Google Scholar] [CrossRef]
- Czeczott, H.; Skirgiello, A. Dicotyledoneae. Foss. Flora Turów Near Bogatynia Second. Part Syst. Descr. Plant Remain. No. 4 1975, 24, 25–56. [Google Scholar]
- Khan, M.A.; Bera, M.; Spicer, R.A.; Spicer, T.E.V.; Bera, S. First occurrence of mastixioid (Cornaceae) fossil in India and its biogeographic implications. Rev. Palaeobot. Palyno 2017, 247, 83–96. [Google Scholar] [CrossRef]
- Handel-Mazzetti, H.F.V. Plantae novae Chingianae. Sinensia 1933, 3, 185–198. [Google Scholar]
- Li, Y.L.; Zhu, H.; Yang, J.B. Systematic position of the genus Mastixia: Evidence from rbc L gene sequences. Acta Bot. Yunnanica 2002, 24, 352–358. [Google Scholar]
- Zhu, W.H.; Xiang, Q.B. The Origin and Distribution of Genus Diplopanax Hand.-Mazz. J. Nanjing For. Univ. 2001, 25, 35. [Google Scholar]
- Averyanov, L.V.; Hiep, N.T. Diplopanax vietnamensis, a New Species of Nyssaceae from Vietnam: One More Living Representative of the Tertiary Flora of Eurasia. Novon 2002, 12, 433–436. [Google Scholar] [CrossRef]
- Ševčík, J.; Kvaček, Z.; Mai, D.H. A new mastixioid florula from tektite-bearing deposits in South Bohemia, Czech Republic (Middle Miocene, Vrábče Member). Bull. Geosci. 2007, 82, 429–436. [Google Scholar] [CrossRef]
- Manchester, S.R.; Collinson, M.E. Mastixioid fruits (Cornales) from the early Eocene London Clay Flora:morphology, anatomy and nomenclatural revision. Foss. Impr. 2022, 78, 310–328. [Google Scholar] [CrossRef]
- Manchester, S.R.; McINTOSH, W.C. Late Eocene silicified fruits and seeds from the John Day Formation near Post, Oregon. PaleoBios 2007, 27, 7–17. [Google Scholar]
- Feng, L.; Xu, Z.Y.; Wang, L. Genetic diversity and demographic analysis of an endangered tree species Diplopanax stachyanthus in subtropical China: Implications for conservation and management. Conserv. Genet. 2019, 20, 315–327. [Google Scholar] [CrossRef]
- Khan, M.A.; Spicer, R.A.; Bera, S.; Ghosh, R.; Yang, J.; Spicer, T.E.V.; Guo, S.X.; Su, T.; Jacques, F.M.B.; Grote, P.J. Miocene to Pleistocene floras and climate of the Eastern Himalayan Siwaliks, and new palaeoelevation estimates for the Namling–Oiyug Basin, Tibet. Glob. Planet. Chang. 2014, 113, 1–10. [Google Scholar] [CrossRef]
- IUCN. Diplopanax stachyanthus. The IUCN Red List of Threatened Species 1998: E.T32339A9699334. Available online: https://www.iucnredlist.org/species/32339/9699334 (accessed on 24 March 2024).
- Fang, X.M.; Yan, M.D.; Zhang, W.L.; Nie, J.S.; Han, W.X.; Wu, F.L.; Song, C.H.; Zhang, T.; Zan, J.B.; Yang, Y.P. Paleogeography control of Indian monsoon intensification and expansion at 41 Ma. Sci. Bull. 2021, 66, 2320–2328. [Google Scholar] [CrossRef] [PubMed]
- Boria, R.A.; Olson, L.E.; Goodman, S.M.; Anderson, R.P. Spatial filtering to reduce sampling bias can improve the performance of ecological niche models. Ecol. Model. 2014, 275, 73–77. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Phillips, S.; Leathwick, J.; Elith, J.; Hijmans, M.R. Package ‘dismo’. Circles 2017, 9, 1–68. [Google Scholar]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Parding, K.M.; Dobler, A.; McSweeney, C.F.; Landgren, O.A.; Benestad, R.; Erlandsen, H.B.; Mezghani, A.; Gregow, H.; Raty, O.; Viktor, E.; et al. GCMeval-An interactive tool for evaluation and selection of climate model ensembles. Clim. Serv. 2020, 18, 100167. [Google Scholar] [CrossRef]
- Wu, T.W.; Lu, Y.X.; Fang, Y.J.; Xin, X.G.; Li, L.; Li, W.P.; Jie, W.H.; Zhang, J.; Liu, Y.M.; Zhang, L.; et al. The Beijing Climate Center Climate System Model (BCC-CSM): The main progress from CMIP5 to CMIP6. Geosci. Model. Dev. 2019, 12, 1573–1600. [Google Scholar] [CrossRef]
- Chen, G.W.; Ling, J.; Zhang, R.W.; Xiao, Z.N.; Li, C.Y. The MJO From CMIP5 to CMIP6: Perspectives from Tracking MJO Precipitation. Geophys. Res. Lett. 2022, 49, e2021GL095241. [Google Scholar] [CrossRef]
- Lin, L.; Jiang, X.L.; Guo, K.Q.; Byrne, A.; Deng, M. Climate change impacts the distribution of Quercus section Cyclobalanopsis (Fagaceae), a keystone lineage in East Asian evergreen broadleaved forests. Plant Divers. 2023, 45, 552–568. [Google Scholar] [CrossRef] [PubMed]
- Pearson, R.G.; Raxworthy, C.J.; Nakamura, M.; Peterson, A.T. Predicting species distributions from small numbers of occurrence records: A test case using cryptic geckos in Madagascar. J. Biogeogr. 2007, 34, 102–117. [Google Scholar] [CrossRef]
- Yang, Y.J.W.; Lin, L.; Tan, Y.H.; Deng, M. How Climate Change Impacts the Distribution of Lithocarpus hancei (Fagaceae), a Dominant Tree in East Asian Montane Cloud Forests. Forests 2023, 14, 1049. [Google Scholar] [CrossRef]
- Cao, Y.T.; Lu, Z.P.; Gao, X.Y.; Liu, M.L.; Sa, W.; Liang, J.; Wang, L.; Yin, W.; Shang, Q.H.; Li, Z.H. Maximum entropy modeling the distribution area of Morchella dill. Ex Pers. species in China under changing climate. Biology 2022, 11, 1027. [Google Scholar] [CrossRef]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Naimi, B.; Araújo, M.B. sdm: A reproducible and extensible R platform for species distribution modelling. Ecography 2016, 39, 368–375. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Wang, Y.S.; Wang, Z.H.; Xing, H.F.; Li, J.W.; Sun, S. Prediction of potential suitable distribution of Davidia involucrata Baill in China based on MaxEnt. Chin. J. Ecol. 2019, 38, 1230–1237. [Google Scholar] [CrossRef]
- Yang, J.T.; Jiang, P.; Huang, Y.; Yang, Y.L.; Wang, R.L.; Yang, Y.X. Potential geographic distribution of relict plant Pteroceltis tatarinowii in China under climate change scenarios. PLoS ONE 2022, 17, e0266133. [Google Scholar] [CrossRef] [PubMed]
- O‘Donnell, J.; Gallagher, R.V.; Wilson, P.D.; Downey, P.O.; Hughes, L.; Leishman, M.R. Invasion hotspots for non-native plants in Australia under current and future climates. Glob. Chang. Biol. 2012, 18, 617–629. [Google Scholar] [CrossRef]
- Zeng, C.Y.; Zhong, Q.J.; Wang, C.Y.; Hu, Y.P.; Wu, M.H.; Meng, W.; Peng, M.C. Ecologically suitable habitats and population characteristics of Cercidiphyllum japonicum in China. Chin. J. Ecol. 2020, 39, 2704–2712. [Google Scholar] [CrossRef]
- Liu, Y.; Huang, P.; Lin, F.R.; Yang, W.Y.; Gaisberger, H.; Christopher, K.; Zheng, Y.Q. MaxEnt modelling for predicting the potential distribution of a near threatened rosewood species (Dalbergia cultrata Graham ex Benth). Ecol. Eng. 2019, 141, 105612. [Google Scholar] [CrossRef]
- Brown, J.L.; Bennett, J.R.; French, C.M. SDMtoolbox 2.0: The next generation Python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. PeerJ 2017, 5, e4095. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.K.; Wei, H.Y.; Lu, C.Y.; Gao, B.; Gu, W. Predictions of potential geographical distribution and quality of Schisandra sphenanthera under climate change. PeerJ 2016, 4, e2554. [Google Scholar] [CrossRef] [PubMed]
- Callaway, R.M.; Cipollini, D.; Barto, K.; Thelen, G.C.; Hallett, S.G.; Prati, D.; Stinson, K.; Klironomos, J. Novel weapons: Invasive plant suppresses fungal mutualists in America but not in its native Europe. Ecology 2008, 89, 1043–1055. [Google Scholar] [CrossRef] [PubMed]
- Callaway, R.M.; Mahall, B.E.; Wicks, C.; Pankey, J.; Zabinski, C. Soil fungi and the effects of an invasive forb on grasses: Neighbor identity matters. Ecology 2003, 84, 129–135. [Google Scholar] [CrossRef]
- Wang, L.; You, Z.P.; Xu, Y.; Zhang, D.H. Research Status and Endangered Causes of Diplopanax stachyanthus. Bull. Bot. Res. 2010, 30, 344–348. [Google Scholar]
- Liu, X.D.; Yin, Z.Y. Sensitivity of East Asian monsoon climate to the uplift of the Tibetan Plateau. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2002, 183, 223–245. [Google Scholar] [CrossRef]
- Zhou, Z.K.; Huang, J.; Ding, W.N. The impact of major geological events on Chinese flora. Biodivers. Sci. 2017, 25, 123–135. [Google Scholar] [CrossRef]
- Yang, Q.; Yuan, M.Q.; Feng, B.X. Study on Community Structure and Resources of Rare Species Diplopanax stachyanthus on the Moon Mountain, Rongjiang. Seed 2013, 32, 55–59. [Google Scholar] [CrossRef]
- Zhu, W.H. Systematic Position and Evolution of Genus Diplopanax stachyanthus Hand.-Mazz. Doctoral Dissertation, Nanjing Forestry University, Nanjing, China, 1998. [Google Scholar]
- Lei, N.F.; Su, Z.X.; Chen, J.S.; Guo, J.H. Germination inhibitors in fruit of rare and endangered Davidia involucrata. Chin. J. Appl. Environ. Biol. 2003, 9, 607–610. [Google Scholar]
- Qian, C.M.; Jiang, Z.; Zhou, J.H.; Dai, S.; Su, Y.Y.; Li, S.X. Changes of inhibitory activity during stratification in Davidia involucrata seeds. J. Nanjing For. Univ. Nat. Sci. Ed. 2016, 40, 188–192. [Google Scholar]
- Mosbrugger, V.; Utescher, T.; Dilcher, D.L. Cenozoic continental climatic evolution of Central Europe. Proc. Natl. Acad. Sci. USA 2005, 102, 14964–14969. [Google Scholar] [CrossRef] [PubMed]
- Eronen, J.T.; Fortelius, M.; Micheels, A.; Portmann, F.T.; Puolamaki, K.; Janis, C.M. Neogene aridification of the Northern Hemisphere. Geology 2012, 40, 823–826. [Google Scholar] [CrossRef]
- Sunderlin, D.; Loope, G.; Parker, N.E.; Williams, C.J. Paleoclimatic and Paleoecological implications of a Paleocene-Eocene fossil leaf assemblage, Chickaloon formation, Alaska. Palaios 2011, 26, 335–345. [Google Scholar] [CrossRef]
- Taylor, D.W. Paleobiogeographic relationships of angiosperms from the Cretaceous and early Tertiary of the North American area. Bot. Rev. 1990, 56, 279–417. [Google Scholar] [CrossRef]
- Vieira, M.; Poças, E.; Pais, J.; Pereira, D. Pliocene flora from S. Pedro da Torre deposits (Minho, NW Portugal). Geodiversitas 2011, 33, 71–85. [Google Scholar] [CrossRef]
- Zhou, P.; Li, X.Z.; Shi, Z.G.; Sha, Y.Y.; Lei, J.; An, Z.S. Strengthened East Asian Winter Monsoon Regulated by Insolation and Arctic Sea Ice Since the Middle Holocene. Geophys. Res. Lett. 2023, 50, e2023GL105440. [Google Scholar] [CrossRef]
- LePage, B.A.; Yang, H.; Matsumoto, M. The evolution and biogeographic history of Metasequoia in the geobiology and ecology of Metasequoia. Geobiol. Ecol. Metasequoia 2005, 22, 3–114. [Google Scholar] [CrossRef]
- Lai, W.F.; Shi, C.Y.; Wen, G.W.; Ln, Z.W.; Ye, L.Q.; Huang, Q.L.; Zhang, G.F. Potential impacts of climate change on the distribution of the relict plant Shaniodendron subaequale. Heliyon 2023, 9, e14402. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.Q.; Dong, Y.F.; Herrando-Moraira, S.; Matsui, T.; Ohashi, H.; He, L.Y.; Nakao, K.; Tanaka, N.; Tomita, M.; Li, X.S.; et al. Potential effects of climate change on geographic distribution of the Tertiary relict tree species Davidia involucrata in China. Sci. Rep. 2017, 7, 43822. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.G.; Lu, J.J. Potential Effects of Climate Change on the Distribution of Dove Trees (Davidia Involucrata Baill) in China. Res. Environ. Sci. 2009, 22, 1371–1381. [Google Scholar]
- Wu, X.T.; Wang, M.Q.; Li, X.Y.; Yan, Y.D.; Dai, M.J.; Xie, W.Y.; Zhou, X.F.; Zhang, D.L.; Wen, Y.F. Response of distribution patterns of two closely related species in Taxus genus to climate change since last inter-glacial. Ecol. Evol. 2022, 12, e9302. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhao, H.X.; Xu, C.G. The potential geographical distribution of Alsophila spinulosain under climate change in China. Chin. J. Ecol. 2021, 40, 968–979. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Wei, H.Y.; Zhang, X.H.; Liu, J.; Zhang, Q.Z.; Gu, W. Non-pessimistic predictions of the distributions and suitability of Metasequoia glyptostroboides under climate change using a random forest model. Forests 2020, 11, 62. [Google Scholar] [CrossRef]
- Alexander, J.M.; Chalmandrier, L.; Lenoir, J.; Burgess, T.I.; Essl, F.; Haider, S.; Kueffer, C.; McDougall, K.; Milbau, A.; Nuñez, M.A.; et al. Lags in the response of mountain plant communities to climate change. Glob. Chang. Biol. 2018, 24, 563–579. [Google Scholar] [CrossRef] [PubMed]
- Seddon, A.W.R.; Macias, F.M.; Long, P.R.; Benz, D.; Willis, K.J. Sensitivity of global terrestrial ecosystems to climate variability. Nature 2016, 531, 229–232. [Google Scholar] [CrossRef] [PubMed]
- Ernakovich, J.G.; Hopping, K.A.; Berdanier, A.B.; Simpson, R.T.; Kachergis, E.J.; Steltzer, H.; Wallenstein, M.D. Predicted responses of arctic and alpine ecosystems to altered seasonality under climate change. Glob. Chang. Biol. 2014, 20, 3256–3269. [Google Scholar] [CrossRef] [PubMed]
- Rudgers, J.A.; Chung, Y.A.; Maurer, G.E.; Moore, D.I.; Muldavin, E.H.; Litvak, M.E.; Collins, S.L. Climate sensitivity functions and net primary production: A framework for incorporating climate mean and variability. Ecology 2018, 99, 576–582. [Google Scholar] [CrossRef] [PubMed]
- Ge, J.L.; Berg, B.; Xie, Z.Q. Climatic seasonality is linked to the occurrence of the mixed evergreen and deciduous broad—Leaved forests in China. Ecosphere 2019, 10, e02862. [Google Scholar] [CrossRef]
- Song, C.Y.; Liu, H.M.; Gao, J.X. Habitat preference and potential distribution of Magnolia officinalis subsp. officinalis and M. o. subsp. biloba in China. Nat. Conserv. Bulg. 2019, 36, 93–111. [Google Scholar] [CrossRef]
- Guan, X.Y.; Shi, W.; Cao, K.F. Effect of Climate Change in Future on Geographical Distribution of Widespread Quercus acutissima and Analysis of Dominant Climatic Factors. J. Trop. Subtrop. Bot. 2018, 26, 661–668. [Google Scholar] [CrossRef]
- Shi, X.D.; Yin, Q.; Sang, Z.Y.; Zhu, Z.L.; Jia, Z.K.; Ma, L.Y. Prediction of potentially suitable areas for the introduction of Magnolia wufengensis under climate change. Ecol. Indic. 2021, 127, 107762. [Google Scholar] [CrossRef]
- Xu, J.; Deng, M.; Jiang, X.L.; Westwood, M.; Song, Y.G.; Turkington, R. Phylogeography of Quercus glauca (Fagaceae), a dominant tree of East Asian subtropical evergreen forests, based on three chloroplast DNA interspace sequences. Tree Genet. Genomes 2015, 11, 805. [Google Scholar] [CrossRef]
- Song, C.Y.; Liu, H.M. Habitat differentiation and conservation gap of Magnolia biondii, M. denudata, and M. sprengeri in China. PeerJ 2019, 6, e6126. [Google Scholar] [CrossRef]
- Holbourn, A.E.; Kuhnt, W.; Clemens, S.C.; Kochhann, K.G.D.; Jöhnck, J.; Lübbers, J.; Andersen, N. Late Miocene climate cooling and intensification of southeast Asian winter monsoon. Nat. Commun. 2018, 9, 1584. [Google Scholar] [CrossRef] [PubMed]
- Willis, K.J.; Niklas, K.J. The role of Quaternary environmental change in plant macroevolution: The exception or the rule? Philos. Trans. R. Soc. London B Biol. Sci. 2004, 359, 159–172. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.W.; Zhang, Y.; Wang, X.J. Phylogeographic history of broad-leaved forest plants in subtropical China. Acta Ecol. Sin. 2017, 37, 5894–5904. [Google Scholar] [CrossRef]
- Hewitt, G.M. Genetic consequences of climatic oscillations in the Quaternary. Philos. T R. Soc. B 2004, 359, 183–195. [Google Scholar] [CrossRef] [PubMed]
- Qian, H.; Ricklefs, R.E. Diversity of temperate plants in east Asia. Nature 2001, 413, 130. [Google Scholar] [CrossRef]
- Shi, Y. Characteristics of late Quaternary monsoonal glaciation on the Tibetan Plateau and in East Asia. Quat. Int. 2002, 97–98, 79–91. [Google Scholar] [CrossRef]
- Shi, Y.F.; Ren, B.H.; Wang, J.T.; Edward, D. Quaternary glaciation in China. Quat. Sci. Rev. 1986, 5, 503–507. [Google Scholar] [CrossRef]
- Tian, Z.; Jiang, D.B. Revisiting last glacial maximum climate over China and East Asian monsoon using PMIP3 simulations. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2016, 453, 115–126. [Google Scholar] [CrossRef]
- Qian, H.; Ricklefs, R.E. Large-scale processes and the Asian bias in species diversity of temperate plants. Nature 2000, 407, 180–182. [Google Scholar] [CrossRef] [PubMed]
- Ricklefs, R.E. A comprehensive framework for global patterns in biodiversity. Ecol. Lett. 2004, 7, 1–15. [Google Scholar] [CrossRef]
- Tian, Z.P.; Jiang, D.B. Mid-Holocene and last glacial maximum changes in monsoon area and precipitation over China. Chin. Sci. Bull. 2015, 60, 400–410. [Google Scholar] [CrossRef]
- Zhang, Y.K.; Zhang, S.S. Effects of seed soaking with salicylic acid on seed germination and early seeding growth of Mastixia euonymoides under drought stress. Hunan For. Sci. Technol. 2022, 49, 24–29. [Google Scholar]
- Zhang, S.S.; Yuan, C.M.; Chen, J.; Zhang, Y.K. Population Status of Wild Mastixia euonymoides as an Extremely Small Population. For. Inventory Plan. 2020, 45, 82–87. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Relative Contribution Rate (%) | Description |
|---|---|---|
| Bio16 | 32.6 | Precipitation of Wettest Quarter (mm) |
| Bio17 | 21.2 | Precipitation of Driest Quarter (mm) |
| Bio19 | 17.3 | Precipitation of Coldest Quarter (mm) |
| Bio1 | 13.5 | Annual Mean Temperature(°C) |
| Bio2 | 13.4 | Mean Diurnal Range (Mean of monthly (max temp–min temp)) (°C) |
| Bio11 | 9.5 | Mean Temperature of Coldest Quarter (°C) |
| Climate Scenario | Time | High Suitable | Medium Suitable | Low Suitable | Total |
|---|---|---|---|---|---|
| (104 km2) | (104 km2) | (104 km2) | (104 km2) | ||
| LGM | 0.779 | 19.807 | 72.477 | 90.063 | |
| MH | 1.824 | 20.668 | 67.326 | 89.818 | |
| Current | 1.113 | 20.391 | 64.470 | 85.974 | |
| SSP1-2.6 | 2041–2060 | 1.161 | 16.292 | 66.477 | 83.931 |
| 2061–2080 | 0.564 | 19.427 | 79.066 | 99.057 | |
| 2081–2100 | 0.816 | 18.335 | 57.361 | 76.512 | |
| SSP2-4.5 | 2041–2060 | 0.269 | 20.146 | 64.641 | 85.056 |
| 2061–2080 | 0.092 | 18.035 | 63.076 | 81.203 | |
| 2081–2100 | 0.170 | 21.175 | 74.519 | 95.865 | |
| SSP3-7.0 | 2041–2060 | 1.248 | 18.589 | 76.691 | 96.528 |
| 2061–2080 | 0.389 | 20.503 | 57.205 | 78.097 | |
| 2081–2100 | 0.234 | 18.170 | 66.457 | 84.861 | |
| SSP5-8.5 | 2041–2060 | 1.625 | 16.359 | 69.005 | 86.990 |
| 2061–2080 | 0.229 | 20.821 | 64.898 | 85.948 | |
| 2081–2100 | 0.719 | 16.875 | 78.139 | 95.733 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, M.; Yang, Y.; Lin, L.; Tan, Y.; Deng, M.; Zuo, Y. Distribution Dynamics of Diplopanax stachyanthus Hand.-Mazz. (Mastixiaceae) and Its Implications in Relict Mastixioid Flora Conservation. Forests 2024, 15, 766. https://doi.org/10.3390/f15050766
Chen M, Yang Y, Lin L, Tan Y, Deng M, Zuo Y. Distribution Dynamics of Diplopanax stachyanthus Hand.-Mazz. (Mastixiaceae) and Its Implications in Relict Mastixioid Flora Conservation. Forests. 2024; 15(5):766. https://doi.org/10.3390/f15050766
Chicago/Turabian StyleChen, Menglin, Yongjingwen Yang, Lin Lin, Yunhong Tan, Min Deng, and Yunjuan Zuo. 2024. "Distribution Dynamics of Diplopanax stachyanthus Hand.-Mazz. (Mastixiaceae) and Its Implications in Relict Mastixioid Flora Conservation" Forests 15, no. 5: 766. https://doi.org/10.3390/f15050766