
Predicting the Spatial Distribution of the Mangshan Pit Viper (Protobothrops mangshanensis) under Climate Change Scenarios Using MaxEnt Modeling
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
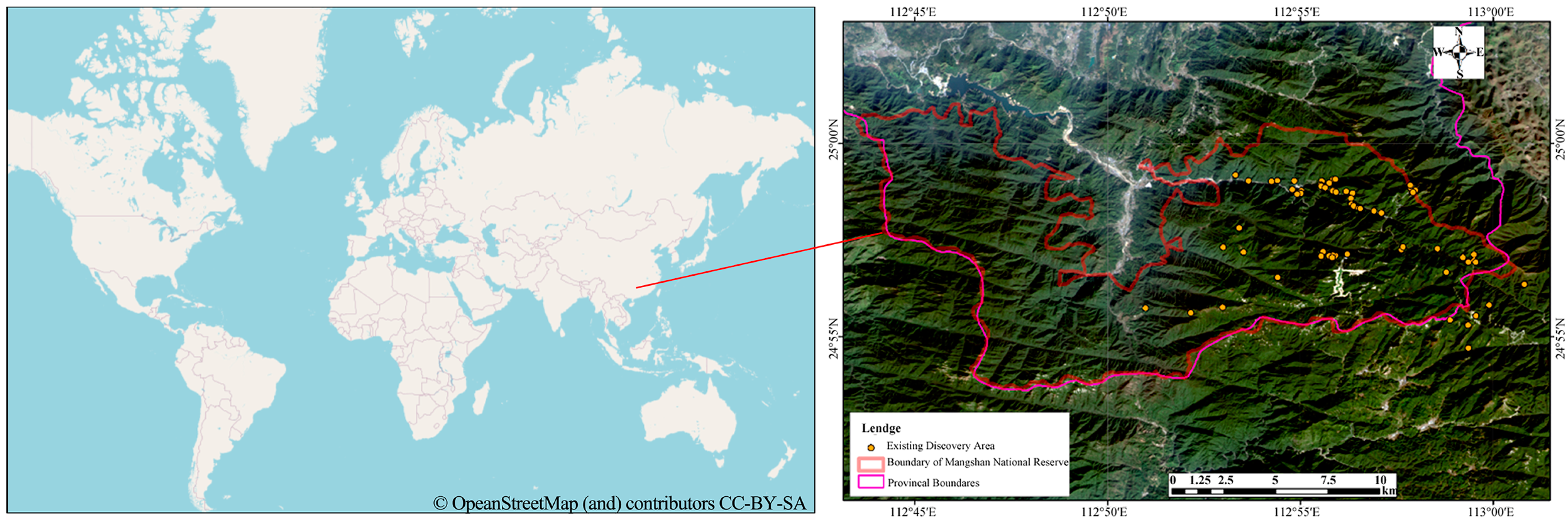
2.1. Habitat Coordinates of the Mangshan Pit Viper
2.2. Environmental Variables
2.3. Environmental Variable Processing
Map Vector Data and Software
2.4. MaxEnt Parameter Optimization
2.5. Calculating the Shifts in the Distribution Area
2.6. Analysis of Habitat Landscape Pattern Changes
3. Results
3.1. Accuracy Evaluation of the MaxEnt Niche Model
3.2. Critical Environmental Factors
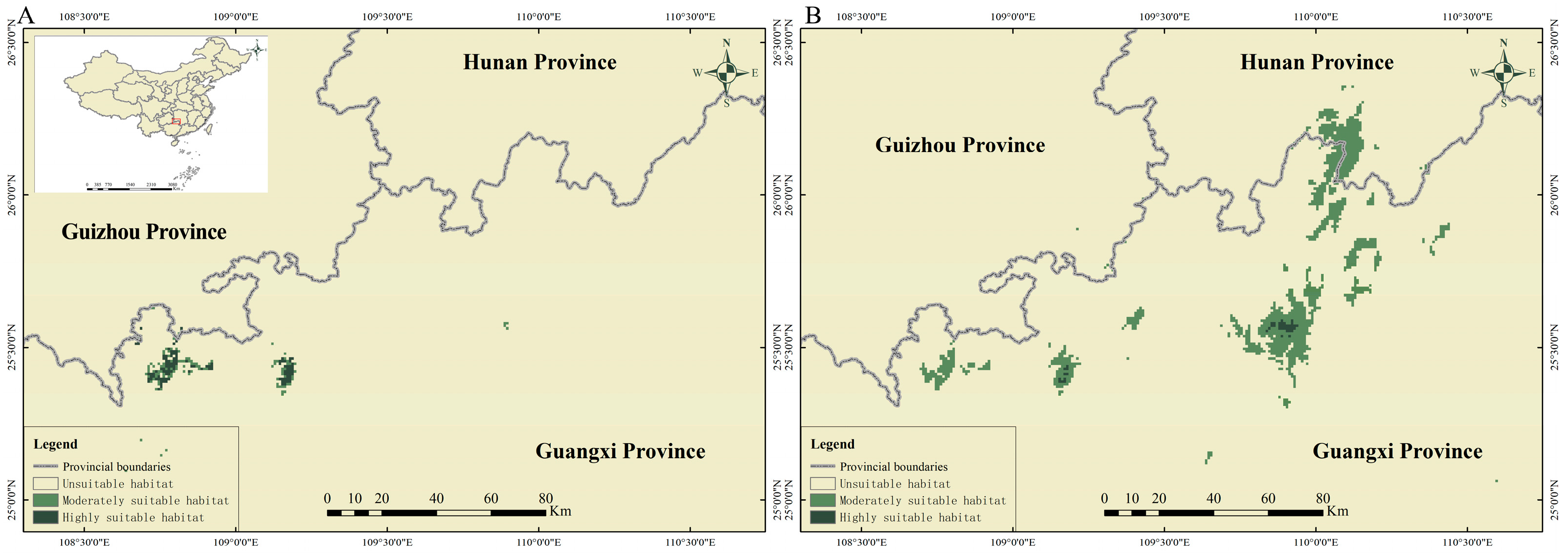
3.3. Current Potential Distribution of P. mangshanensis (2020)
3.4. Future Geographic Range (2040–2060 and 2060–2080)
3.5. Alteration in Habitat Landscape Patterns
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cong, M.; Xu, Y.; Tang, L.; Yang, W.; Jian, M. Predicting the dynamic distribution of Sphagnum bogs in China under climate change since the last interglacial period. PLoS ONE 2020, 15, e0230969. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Wang, B.; Chen, C.; Zhou, G. MaxEnt Modeling to Predict the Current and Future Distribution of Pomatosace filicula under Climate Change Scenarios on the Qinghai–Tibet Plateau. Plants 2022, 11, 670. [Google Scholar] [CrossRef] [PubMed]
- Wiens, J.J. Climate-Related Local Extinctions Are Already Widespread among Plant and Animal Species. PLoS Biol. 2016, 14, e2001104. [Google Scholar] [CrossRef] [PubMed]
- Gong, S.; Chen, Y.; Lau, M.; Wang, F. Population status, distribution and conservation needs of the Endangered Mangshan pit viper Protobothrops mangshanensis of China. Oryx 2013, 4, 122–127. [Google Scholar] [CrossRef]
- Duarte, M.; Guerrero, P.C.; Arroyo, M.T.; Bustamante, R.O. Niches and climate-change refugia in hundreds of species from one of the most arid places on Earth. PeerJ 2019, 7, e7409. [Google Scholar] [CrossRef] [PubMed]
- Orlov, N.; Ryabov, S.; Nguyen, T. Two new species of Genera Protobothrops Hoge et Romano-Hoge, 1983 and Viridovipera Malhotraet Thorpe. 2004 (Ophidia, Viperidae, Crotalinae) from Karst Region in Northeastern Vietnam. Part I. Description of a new species of Protobothrops Genus. Russ. J. Herpetol. 2009, 16, 69–82. [Google Scholar] [CrossRef]
- Guo, P.; Liu, Q.; Wen, T.; Xiao, R.; Fang, M.; Zhong, G.; Truong, N.Q.; Zhu, F.; Jadin, R.C.; Li, C. Multilocus phylogeny of the Asian Lance-headed pitvipers (Squamata, Viperidae, Protobothrops). Zootaxa 2016, 4093, 382–390. [Google Scholar] [CrossRef] [PubMed]
- IUCN (International Union for Conservation of Nature) Red List Committee. The IUCN Red List of Threatened Species™ Strategic Plan 2017–2020; IUCN: Gland, Switzerland, 2016. [Google Scholar] [CrossRef]
- Li, S.; Lei, S.; Ren, X.; Liu, L.; Li, Q. Study on the incubation and rearing of eggs of the critically endangered pit viper Protobothrops mangshanensis. J. For. Environ. Sci. Chin. 2018, 34, 120–123. [Google Scholar] [CrossRef]
- Ding, X. Behavioral Rhythm of a Critically Endangered and Endemic Pitviper, Protobothrops mangshanensis. Master’s Thesis, Central South University of Forestry and Technology, Changsha, China, 2020; pp. 37–38. [Google Scholar] [CrossRef]
- Zhang, B. Multilevel Multiscale Habitat Selection Mechanisms of the Critically Endangered Mangshan Pit Viper (Protobothrops mangshanensis). Ph.D. Thesis, Central South University of Forestry and Technology, Changsha, China, 2021; pp. 38–39. [Google Scholar] [CrossRef]
- Ahmadi, M.; Hemami, M.R.; Kaboli, M.; Shabani, F. MaxEnt brings comparable results when the input data are being completed; Model parameterization of four species distribution models. Ecol. Evolution 2023, 13, e9827. [Google Scholar] [CrossRef]
- Zhang, J.; Jiang, F.; Li, G.; Qin, W.; Li, S.; Gao, H.; Cai, Z.; Lin, G.; Zhang, T. Maxent modeling for predicting the spatial distribution of three raptors in the Sanjiangyuan National Park, China. Ecol. Evol. 2019, 9, 6643–6654. [Google Scholar] [CrossRef]
- Qin, A.; Jin, K.; Batsaikhan, M.-E.; Nyamjav, J.; Li, G.; Li, J.; Xue, Y.; Sun, G.; Wu, L.; Indree, T.; et al. Predicting the current and future suitable habitats of the main dietary plants of the Gobi Bear using MaxEnt modeling. Glob. Ecol. Conserv. 2020, 22, e01032. [Google Scholar] [CrossRef]
- Li, M.; He, J.; Zhao, Z.; Lyu, R.; Yao, M.; Cheng, J.; Xie, L. Predictive modelling of the distribution of Clematis sect. Fruticella s. str. under climate change reveals a range expansion during the Last Glacial Maximum. PeerJ 2020, 8, e8729. [Google Scholar] [CrossRef] [PubMed]
- Prakash, S. Impact of Climate change on Aquatic Ecosystem and its Biodiversity: An overview. Int. J. Biol. Innov. 2021, 3. [Google Scholar] [CrossRef]
- Resano-Mayor, J.; Korner-Nievergelt, F.; Vignali, S.; Horrenberger, N.; Barras, A.G.; Braunisch, V.; Pernollet, C.A.; Arlettaz, R. Snow cover phenology is the main driver of foraging habitat selection for a high-alpine passerine during breeding: Implications for species persistence in the face of climate change. Biodivers. Conserv. 2019, 28, 2669–2685. [Google Scholar] [CrossRef]
- Zhang, M. Potential Distribution Prediction and Habitat Suitability Evaluation of Mangshan Pit Vipers. Master’s Thesis, Central South University of Forestry and Technology, Hunan, China, 2018. [Google Scholar] [CrossRef]
- Zhang, B.; Wu, B.; Yang, D.; Tao, X.; Zhang, M.; Hu, S.; Chen, J.; Zheng, M. Habitat association in the critically endangered Mangshan pit viper (Protobothrops mangshanensis), a species endemic to China. PeerJ 2020, 8, e9439. [Google Scholar] [CrossRef]
- Carne, C. Predicting habitat suitability for the wart-biter bush cricket (Decticus verrucivorus) in Europe. J. Insect Conserv. 2017, 21, 287–295. [Google Scholar] [CrossRef]
- Mafuwe, K.; Broadley, S.; Moyo, S. Use of maximum entropy (Maxent) niche modelling to predict the occurrence of threatened freshwater species in a biodiversity hotspot of Zimbabwe. Afr. J. Ecol. 2021, 60, 557–565. [Google Scholar] [CrossRef]
- Zhang, L.; Huang, S.; Yuan, Y.; Wu, X.; Tan, Z.; Yao, L.; Hong, Z.; Cai, Q.; Wang, Y.; Xiang, H. Geographical distribution and predict potential distribution of Cerasus serrulata. Environ. Sci. Pollut. Res. 2023, 30, 43369–43376. [Google Scholar] [CrossRef]
- Peterson, A.T. perspective: Phylogeography is not enough: The need for multiple lines of evidence. Front. Biogeogr. 2012, 1, 19–25. [Google Scholar] [CrossRef]
- Li, Q.; Ren, H.; Zheng, L.; Cao, W.; Zhang, A.; Zhuang, D.; Lu, L.; Jiang, H. Ecological Niche Modeling Identifies Fine-Scale Areas at High Risk of Dengue Fever in the Pearl River Delta, China. Int. J. Environ. Res. Public Health 2017, 14, 619. [Google Scholar] [CrossRef]
- Wang, R.; Li, Q.; He, S.; Liu, Y.; Wang, M.; Jiang, G. Modeling and mapping the current and future distribution of Pseudomonas syringae pv. actinidiae under climate change in China. PLoS ONE 2018, 13, e0192153. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Park, D.S.; Liang, Y.; Pandey, R.; Papeş, M. Collinearity in ecological niche modeling: Confusions and challenges. Ecol. Evol. 2019, 9, 10365–10376. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; White, M.; Newell, G. Selecting thresholds for the prediction of species occurrence with presence-only data. J. Biogeogr. 2013, 40, 778–789. [Google Scholar] [CrossRef]
- Freeman, E.A.; Moisen, G.G. A comparison of the performance of threshold criteria for binary classification in terms of predicted prevalence and kappa. Ecol. Model. 2008, 217, 48–58. [Google Scholar] [CrossRef]
- Wang, M.; Chen, H.; Lei, M. Identifying potentially contaminated areas with MaxEnt model for petrochemical industry in China. Environ. Sci. Pollut. Res. 2022, 29, 54421–54431. [Google Scholar] [CrossRef] [PubMed]
- Yousefi, M.; Kafash, A.; Khani, A.; Nabati, N. Applying species distribution models in public health research by predicting snakebite risk using venomous snakes’ habitat suitability as an indicating factor. Sci. Rep. 2020, 10, 18073. [Google Scholar] [CrossRef] [PubMed]
- Portillo, J.T.d.M.; Barbo, F.E.; Sawaya, R.J. Climatic niche breadths of the Atlantic Forest snakes do not increase with increasing latitude. Curr. Zool. 2021, 68, 535–540. [Google Scholar] [CrossRef] [PubMed]
- Ceron, K.; Mângia, S.; Guedes, T.B.; Alvares, D.J.; Neves, M.O.; Moroti, M.D.T.; Torello, N.; Borges-Martins, M.; Ferreira, V.L.; Santana, D.J. Ecological Niche Explains the Sympatric Occurrence of Lined Ground Snakes of the Genus Lygophis (Serpentes, Dipsadidae) in the South American Dry Diagonal. Herpetologica 2021, 77, 239–248. [Google Scholar] [CrossRef]
- Sunny, A.; Manjarrez, J.; Caballero-Viñas, C.; Bolom-Huet, R.; Gómez-Ortiz, Y.; Domínguez-Vega, H.; Heredia-Bobadilla, R.L.; Torres-Romero, E.J.; González-Fernández, A. Modelling the effects of climate and land-cover changes on the potential distribution and landscape connectivity of three earth snakes (Genus Conopsis, Günther 1858) in central Mexico. Sci. Nat. 2023, 110, 52. [Google Scholar] [CrossRef]
- Andrade, S.; Sarquis, A.; Loiselle, A.; Giraudo, A.R.; Díaz-Gómez, J.M. Expansion of the agricultural frontier in the largest South American Dry Forest: Iden-tifying priority conservation areas for snakes before everything is lost. PLoS ONE 2019, 14, e0221901. [Google Scholar] [CrossRef]
- Pyron, R.A.; Burbrink, F.T. Early origin of viviparity and multiple reversions to oviparity in squamate reptiles. Ecol. Lett. 2013, 17, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Walther, G.-R.; Post, E.; Convey, P.; Menzel, A.; Parmesan, C.; Beebee, T.J.C.; Fromentin, J.-M.; Hoegh-Guldberg, O.; Bairlein, F. Ecological responses to recent climate change. Nature 2002, 416, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Pearson, R.G. Climate change and the migration capacity of species. Trends Ecol. Evol. 2006, 21, 111–113. [Google Scholar] [CrossRef] [PubMed]
- Marshall, B.M.; Strine, C.T. Exploring snake occurrence records: Spatial biases and marginal gains from accessible social media. PeerJ 2019, 7, e8059. [Google Scholar] [CrossRef] [PubMed]
- Román-Palacios, C.; Wiens, J.J. Recent responses to climate change reveal the drivers of species extinction and survival. Proc. Natl. Acad. Sci. USA 2020, 117, 4211–4217. [Google Scholar] [CrossRef]
- Li, Y.; Xie, Z.; Qin, Y.; Zheng, Z. Estimating Relations of Vegetation, Climate Change, and Human Activity: A Case Study in the 400 mm Annual Precipitation Fluctuation Zone, China. Remote Sens. 2019, 11, 1159. [Google Scholar] [CrossRef]
- Pei, F.; Zhou, Y.; Xia, Y. Application of Normalized Difference Vegetation Index (NDVI) for the Detection of Extreme Precipitation Change. Forests 2021, 12, 594. [Google Scholar] [CrossRef]
- Chen, Z.; Wang, W.; Fu, J. Vegetation response to precipitation anomalies under different climatic and biogeographical conditions in China. Sci. Rep. 2020, 10, 830. [Google Scholar] [CrossRef]
- Liu, H.; Jia, J.; Lin, Z.; Wang, Z.; Gong, H. Relationship between net primary production and climate change in different vegetation zones based on EEMD detrending—A case study of Northwest China. Ecol. Indic. 2020, 122, 107276. [Google Scholar] [CrossRef]
- Kong, D.; Miao, C.; Wu, J.; Zheng, H.; Wu, S. Time lag of vegetation growth on the Loess Plateau in response to climate factors: Estimation, distribution, and influence. Sci. Total. Environ. 2020, 744, 140726. [Google Scholar] [CrossRef]
- Bonnet, X.; Naulleau, G.; Shine, R. The dangers of leaving home: Dispersal and mortality in snakes. Biol. Conserv. 1999, 89, 39–50. [Google Scholar] [CrossRef]
- Shiferaw, H.; Bewket, W.; Alamirew, T.; Zeleke, G.; Teketay, D.; Bekele, K.; Schaffner, U.; Eckert, S. Implications of land use/land cover dynamics and Prosopis invasion on ecosystem service values in Afar Region, Ethiopia. Sci. Total. Environ. 2019, 675, 354–366. [Google Scholar] [CrossRef] [PubMed]
- Kelly, L.T.; Giljohann, K.M.; Duane, A.; Aquilué, N.; Archibald, S.; Batllori, E.; Bennett, A.F.; Buckland, S.T.; Canelles, Q.; Clarke, M.F.; et al. Fire and biodiversity in the Anthropocene. Science 2020, 370, 929. [Google Scholar] [CrossRef] [PubMed]
- Lian, X.; Jiao, L.; Liu, Z.; Jia, Q.; Zhong, J.; Fang, M.; Wang, W. Multi-spatiotemporal heterogeneous legacy effects of climate on terrestrial vegetation dynamics in China. GIScience Remote Sens. 2022, 59, 164–183. [Google Scholar] [CrossRef]
- Sui, Y.; Huang, W.; Yang, X.; Li, M. Characteristics adaptation of seasonal drought in southern China under the background of climate change: I. Evolution of precipitation resources. Chin. J. Appl. Ecol. 2012, 23, 1875–1882. [Google Scholar] [CrossRef]
- Valencia-Aguilar, A.; Cortés-Gómez, A.M.; Ruiz-Agudelo, C.A. Ecosystem services provided by amphibians and reptiles in Neotropical ecosystems. Int. J. Biodivers. Sci. Ecosyst. Serv. Manag. 2013, 9, 257–272. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Definitions of the 19 Bioclimatic Variables |
|---|
| BIO1 = Annual Mean Temperature |
| BIO2 = Mean Diurnal Range [Mean of monthly (max temp–min temp)] |
| BIO3 = Isothermality [(BIO2/BIO7) × 100] |
| BIO4 = Temperature Seasonality (standard deviation × 100) |
| BIO5 = Max Temperature in the Warmest Month |
| BIO6 = Min Temperature in the Coldest Month |
| BIO7 = Temperature Annual Range (BIO5–BIO6) |
| BIO8 = Mean Temperature in the Wettest Quarter |
| BIO9 = Mean Temperature in the Driest Quarter |
| BIO10 = Mean Temperature in the Warmest Quarter |
| BIO11 = Mean Temperature in the Coldest Quarter |
| BIO12 = Annual Precipitation |
| BIO13 = Precipitation in the Wettest Month |
| BIO14 = Precipitation in the Driest Month |
| BIO15 = Precipitation Seasonality (Coefficient of Variation) |
| BIO16 = Precipitation in the Wettest Quarter |
| BIO17 = Precipitation in the Driest Quarter |
| BIO18 = Precipitation in the Warmest Quarter |
| BIO19 = Precipitation in the Coldest Quarter |
| Variables | Percentage Contribution | Permutation Importance |
|---|---|---|
| BIO14 | 56.5 | 57.0 |
| BIO5 | 22.9 | 3.7 |
| BIO15 | 15.5 | 6.4 |
| BIO13 | 2.9 | 1.2 |
| BIO6 | 1.2 | 31.2 |
| BIO2 | 1.1 | 0.5 |
| Period | Climate Model | Suitable Range | Total (km2) | |
|---|---|---|---|---|
| Moderately (km2) | Highly (km2) | |||
| 1970–2023 | 531.14 | 491.99 | 1023.12 | |
| 2050 | SSPs126 | 529.36 | 347.86 | 877.22 |
| 2050 | SSPs370 | 459.44 | 34.26 | 459.44 |
| 2050 | SSPs585 | 107.05 | 99.02 | 107.05 |
| 2070 | SSPs126 | 241.45 | 33.10 | 1174.55 |
| 2070 | SSPs370 | 506.67 | 2.93 | 509.60 |
| 2070 | SSPs585 | 235.33 | 0 | 235.33 |
| Model | SSPs126 | SSPs370 | SSPs585 | ||||
|---|---|---|---|---|---|---|---|
| Year | 1970–2023 | 2050 | 2070 | 2050 | 2070 | 2050 | 2070 |
| NP | 48 | 40 | 33 | 23 | 131 | 67 | 37 |
| MPS | 21.315 | 21.931 | 8.320 | 21.465 | 3.890 | 3.076 | 6.36 |
| LPI | 0.631 | 0.100 | 0.461 | 0.060 | 0.232 | 0.135 | 0.094 |
| SHAPE_MN | 1.249 | 1.243 | 1.242 | 1.160 | 1.149 | 1.148 | 1.059 |
| PROX | 12.167 | 12.144 | 11.203 | 4.716 | 6.887 | 2.463 | 1.034 |
| Model | SSPs126 | SSPs370 | SSPs585 | ||||
|---|---|---|---|---|---|---|---|
| Year | 1970–2023 | 2050 | 2070 | 2050 | 2070 | 2050 | 2070 |
| PC | 0.098 | 0.072 | 0.036 | 0.020 | 0.175 | 0.057 | 0.014 |
| Number of VIPs | 163 | 117 | 46 | 30 | 314 | 58 | 0 |
| Number of IPs | 102 | 46 | 26 | 23 | 131 | 31 | 0 |
| Number of UPs | 198 | 165 | 98 | 41 | 340 | 157 | 17 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deng, Z.; Xia, X.; Zhang, M.; Chen, X.; Ding, X.; Zhang, B.; Deng, G.; Yang, D. Predicting the Spatial Distribution of the Mangshan Pit Viper (Protobothrops mangshanensis) under Climate Change Scenarios Using MaxEnt Modeling. Forests 2024, 15, 723. https://doi.org/10.3390/f15040723
Deng Z, Xia X, Zhang M, Chen X, Ding X, Zhang B, Deng G, Yang D. Predicting the Spatial Distribution of the Mangshan Pit Viper (Protobothrops mangshanensis) under Climate Change Scenarios Using MaxEnt Modeling. Forests. 2024; 15(4):723. https://doi.org/10.3390/f15040723
Chicago/Turabian StyleDeng, Zeshuai, Xin Xia, Mu Zhang, Xiangying Chen, Xiangyun Ding, Bing Zhang, Guoxing Deng, and Daode Yang. 2024. "Predicting the Spatial Distribution of the Mangshan Pit Viper (Protobothrops mangshanensis) under Climate Change Scenarios Using MaxEnt Modeling" Forests 15, no. 4: 723. https://doi.org/10.3390/f15040723