
Edge Influence on Diversity of Orchids in Andean Cloud Forests


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sampling and Data Collection
2.3. Data Analysis
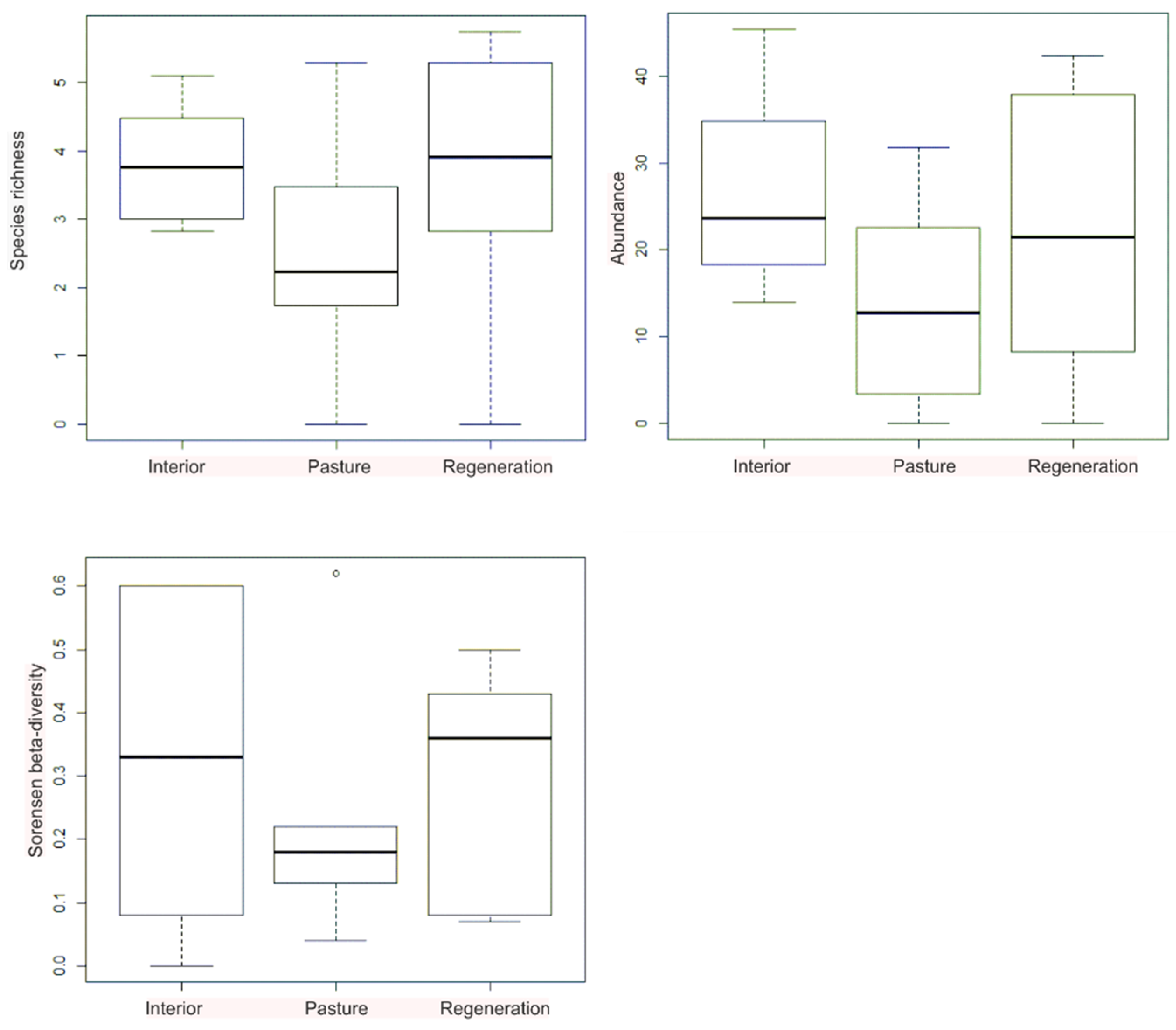
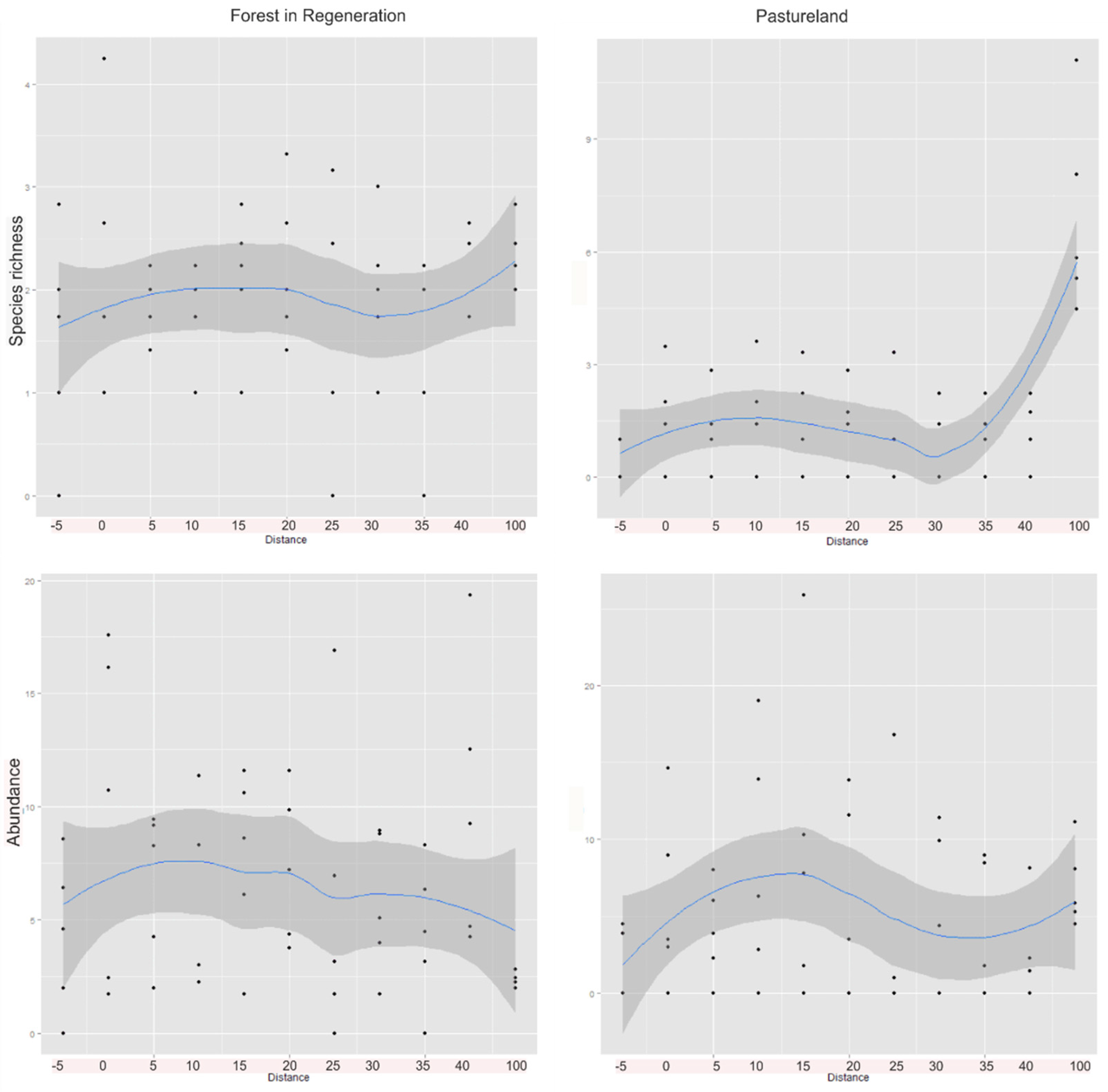
3. Results
4. Discussion
5. Conclusions
Supplementary Files
Supplementary File 1Supplementary File 2Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| DBH | Diameter at Breast Height |
| SU | Sampling Unit |
| CA | Area (ha) |
| TE | Total Edge |
| ED | Edge Density |
| MSI | Mean Shape Index |
| MPAR | Mean Patch Area Perimeter Ratio |
| MPFD | Mean Patch Fractal Dimension |
| DN | Tree density (trees per ha) |
| BA | Basal surface area (squared meters per ha) |
| Co | Cover (percentage %) |
| R | Richness (number of species) |
| A | Abundance (number individuals) |
| H’ | Shannon-Wiener index |
| GLM | Generalized linear model |
| ANOVA | Analysis of variance |
References
- Stadmüller, T. Cloud Forest in the Humid Tropics; United Nations University: Tokyo, Japan, 1987; pp. 17–27. [Google Scholar]
- Young, B.; Young, K.R.; Josse, C. Vulnerability of Tropical Andean Ecosystems to Climate Change. In Climate Change and Biodiversity in the Tropical Andes; Herzog, S.K., Martinez, R., Jorgensen, P.M., Tiessen, H., Eds.; Inter-American Institute for Global Change Research (IAI) and Scientific Committee on Problems of the Environment (SCOPE): San Jose dos Campos, Brazil, 2011; pp. 170–181. [Google Scholar]
- Josse, C.; Cuesta, F.; Navarro, G.; Barrena, V.; Cabrera, E.; Chacón-Moreno, E.; Ferreira, W.; Peralvo, M.; Saito, J.; Tovar, A. Ecosistemas de los Andes del norte y centro: Bolivia, Colombia, Ecuador, Peru y Venezuela; Secretaría General de la Comunidad Andina, Programa Regional ECOBONA-Intercooperation, CONDESAN, Proyecto Páramo Andino, Programa BioAndes, EcoCiencia, NatureServe, IAvH, LTAUNALM, ICAE-ULA, CDC-UNALM, and RUMBOL SRL. Secretaría General de la Comunidad Andina: Lima, Peru, 2009; pp. 44–48. [Google Scholar]
- Gentry, A.H.; Dodson, C.H. Diversity and biogeography of Neotropical vascular epiphytes. Ann. Mo. Bot. Gard. 1987, 74, 205–233. [Google Scholar] [CrossRef]
- Bubb, P.; May, I.; Miles, L.; Sayer, J. Cloud Forest Agenda; United Nations Environment Programme—World Conservation Monitoring Centre: Cambridge, UK, 2004; pp. 7–16. [Google Scholar]
- Bruijnzeel, L.A.; Mulligan, M.; Scatena, F. Hydrometeorology of tropical montane cloud forests: Emerging patterns. Hydrol. Process. 2011, 25, 465–498. [Google Scholar] [CrossRef]
- Mulligan, M.; Burke, S.M. Global Cloud Forest and Environmental Change in a Hydrological Context. Final Report; United Kingdom Department for International Development: London, UK, 2005; pp. 7–20.
- Doumenge, C.; Gilmour, D.; Perez, M.R.; Blockhus, J. Tropical montane cloud forests: Conservation status and management issues. In Tropical Montane Cloud Forests Ecological Studies; Hamilton, L.S., Juvik, J.O., Scatena, F.N., Eds.; Springer-Verlag: Berlin, Germany, 1995; pp. 100–110. [Google Scholar]
- Pope, I.; Bowen, D.; Harbor, J.; Shao, G.; Zanotti, L.; Burniske, G. Deforestation of montane cloud forest in the Central Highlands of Guatemala: Contributing factors and implications for sustainability in Q’eqchi’ communities. Int. J. Sustain. Dev. World Ecol. 2015, 22, 201–212. [Google Scholar] [CrossRef]
- Forman, R.T. Land Mosaics. The Ecology of Landscapes and Regions; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- Driscoll, D.A.; Banks, S.C.; Barton, P.S.; Lindenmayer, D.B.; Smith, A.L. Conceptual domain of the matrix in fragmented landscapes. Trends Ecol. Evol. 2013, 28, 605–613. [Google Scholar] [CrossRef] [PubMed]
- Didham, R.K. Ecological consequences of habitat fragmentation. In Encyclopaedia of Life Sciences; Johansson, R., Ed.; Wiley: Hoboken, NJ, USA, 2010; Available Online: http://www.els.net (accessed on 25 June 2013).
- Bender, D.J.; Fahrig, L. Matrix structure obscures the relationship between interpatch movement and patch size and isolation. Ecology 2005, 86, 1023–1033. [Google Scholar] [CrossRef]
- Baker, S.C.; Barmuta, A.; Mcquillanc, P.B.; Richardson, M.M. Estimating edge effects on ground-dwelling beetles at clearfelled non-riparian stand edges in Tasmanian wet eucalypt forest. Forest Ecol. Manag. 2007, 239, 92–101. [Google Scholar] [CrossRef]
- Fischer, J.; Lindenmayer, D.B. Landscape modification and habitat fragmentation: A synthesis. Global Ecol. Biogeogr. 2007, 16, 265–280. [Google Scholar] [CrossRef]
- Lövei, G.L.; Magura, T.; Tothmeresz, B.; Ködöböcz, V. The influence of matrix and edges on species richness patterns of ground beetles (Coleoptera: Carabidae) in habitat islands. Global Ecol. Biogeogr. 2006, 15, 283–289. [Google Scholar] [CrossRef]
- Harper, K.A.; Macdonald, S.E.; Burton, P.J.; Chen, J.Q.; Brosofske, K.D.; Saunders, S.C.; Euskirchen, E.; Roberts, S.D.; Jaiteh, M.; Sandesseen, P.A. Edge influence on forest structure and composition in fragmented landscapes. Conserv. Biol. 2005, 19, 768–782. [Google Scholar] [CrossRef]
- Ewers, R.M.; Didham, R.K. The effect of fragment shape and species’ sensitivity to habitat edges on animal population size. Conserv. Biol. 2007, 21, 926–936. [Google Scholar] [CrossRef] [PubMed]
- Meiners, S.J.; Pickett, S.T.A. Changes in community and population responses across a forest-field gradient. Ecography 1999, 22, 261–267. [Google Scholar] [CrossRef]
- Gradstein, R. Epiphytes of tropical montane forests—impact of deforestation and climate change. In The Tropical Mountain Forest—Patterns and Processes in a Biodiversity Hotspot; Gradstein, S., Homeier, J., Gansert, D., Eds.; Göttingen Centre for Biodiversity and Ecology, Georg-August-Universität Göttingen: Göttingen, Germany, 2009; pp. 51–58. [Google Scholar]
- Nadkarni, N.M. Colonization of stripped branch surfaces by epiphytes in a lower montane cloud forest. Biotropica 2000, 32, 358–363. [Google Scholar] [CrossRef]
- Nöske, N.; Hilt, N.; Werner, F.; Brehm, G.; Fiedler, K.; Sipman, H.; Gradstein, S. Disturbance effects on diversity in montane forest of Ecuador: Sessile epiphytes versus mobile moths. Basic Appl. Ecol. 2009, 9, 4–12. [Google Scholar] [CrossRef]
- Barthlott, W.; Schmit-Neuerburg, V.; Nieder, J.; Engwald, S. Diversity and abundance of vascular epiphytes: A comparison of secondary vegetation and primary montane rain forest in the Venezuelan Andes. Plant Ecol. 2001, 152, 145–156. [Google Scholar] [CrossRef]
- Holz, I.; Gradstein, S.R. Cryptogamic epiphytes in primary and recovering upper montane oak forests of Costa Rica-species richness, community composition and ecology. Plant Ecol. 2005, 178, 89–109. [Google Scholar] [CrossRef]
- Acebey, A.; Gradstein, S.R.; Krömer, T. Species richness and habitat diversification of bryophytes in submontane rain forest and fallows of Bolivia. J. Trop. Ecol. 2003, 19, 9–18. [Google Scholar] [CrossRef]
- Krömer, T.; Gradstein, S.R. Species richness of vascular epiphytes in two primary forest and fallows in the Bolivian Andes. Selbyana 2003, 24, 190–195. [Google Scholar]
- Jacquemyn, H.; Brys, R.; Vandepitte, K.; Honnay, O.; Roldan-Ruiz, I.; Wiegand, T. A spatially explicit analysis of seedlings recruitment in a terrestrial orchid Orchis purpurea. New Phytol. 2007, 176, 448–459. [Google Scholar] [CrossRef] [PubMed]
- Olmsted, I.; Juárez, M.G. Distribution and conservation of epiphytes on the Yucatan Peninsula. Selbyana 1996, 17, 58–70. [Google Scholar]
- Dodson, C.H.; Gentry, A.H. Biological extinction in western Ecuador. Ann. Mo. Bot. Gard. 1991, 78, 273–295. [Google Scholar] [CrossRef]
- Sydes, M. Orchids: indicators of management success? Vic. Nat. 1994, 111, 213–217. [Google Scholar]
- Bergman, E.; Ackerman, J.; Thomson, J.; Zimmerman, J. Land-use history affects the distribution of the saprophytic orchid Wullschlaegelia calacrata in Puerto Rico’s Tabonuco Forest. Biotropica 2006, 38, 492–499. [Google Scholar] [CrossRef]
- Kull, T.; Hutchings, M. A comparative analysis of decline in the distribution ranges of orchid species in Estonia and the United Kingdom. Biol. Conserv. 2006, 129, 31–39. [Google Scholar] [CrossRef]
- Wotavova, K.; Balaunova, Z.; Kindlmann, P. Factors affecting persistence of terrestrial orchids in wet meadows and implications for their conservation in a changing rural landscape. Biol. Conserv. 2004, 118, 271–279. [Google Scholar] [CrossRef]
- Rumpff, L.; Coates, F.; Messina, A.; Morgan, J. Potential Biological Indicators of Climate Change: Evidence from Phenology Records of Plants along the Victorian Coast; Victorian Government Department of Sustainability and Environment: Melbourne, Australia, 2008; pp. 17–47.
- Armenteras, D.; Cadena, V.C.; Moreno, R.P. Evaluación del Estado de los Bosques de Niebla y de la Meta 2010 en Colombia; Instituto de Investigación de Recursos Biológicos Alexander von Humboldt: Bogotá, Colombia, 2007; pp. 10–22. [Google Scholar]
- Betancur, J.; Sarmiento-L., H.; Toro-González, L.; Valencia, J. (Eds.) Ministerio De Ambiente Y Desarrollo Sostenible Y Universidad Nacional De Colombia. In Plan Para el Estudio y la Conservación de las Orquídeas en Colombia; Ministerio de Ambiente y Desarrollo Sostenible, Universidad Nacional de Colombia: Bogotá, Colombia, 2015; p. 336.
- Etter, A.; McAlpine, C.; Wilson, K.; Phinn, S.; Possingham, H. Regional patterns of agricultural land use and deforestation in Colombia. Agric. Ecosyst. Environ. 2006, 114, 369–386. [Google Scholar] [CrossRef]
- Armenteras, D.; Rodríguez-Eraso, N.; Alumbreros, J.R.; Morales, M. Understanding deforestation in montane and lowland forests of the Colombian Andes. Reg. Environ. Change. 2011, 11, 693–705. [Google Scholar] [CrossRef]
- Meisel, J.E.; Woodward, C.L. Andean Orchid Conservation and the Role of Private Lands: A Case Study from Ecuador. Selbyana 2010, 26, 49–57. [Google Scholar]
- Magurran, A.E. Ecological Diversity and Its Measurement; Princeton University Press: Princeton, NJ, USA, 1988; pp. 7–45. [Google Scholar]
- Pielou, E.C. 2k contingency tables in ecology. J. Theor. Biol. 1972, 34, 337–330. [Google Scholar] [CrossRef]
- Shannon, C.E.; Weaver, W. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef]
- Baselga, A. Partitioning the turnover and nestedness components of beta-diversity. Glob. Ecol. Biogeogr. 2010, 19, 134–143. [Google Scholar] [CrossRef]
- Baselga, A.; Leprieur, F. Comparing methods to separate components of beta diversity. Methods Ecol. Evol. 2015, 6, 1069–1079. [Google Scholar] [CrossRef]
- Cottam, G.; Curtis, J.T. The use of distance measures in phytosociological sampling. Ecology 1956, 37, 451–460. [Google Scholar] [CrossRef]
- Mitchell, K. Quantitative Analysis by the Point-Centered Quarter Method; Department of Mathematics and Computer Science, Hobart and William smith Colleges: Geneva, NY, USA, 2007. [Google Scholar]
- Krömer, T.; Kessler, M.; Gradstein, S.R. Vertical stratification of vascular epiphytes in submontane and montane forest of the Bolivian Andes: The importance of the understory. Plant Ecol. 2007, 189, 261–278. [Google Scholar]
- Sáyago, R.; Lopezaraiza-Mikel, M.; Quesada, M.; Álvarez-Añorve, M.Y.; Cascante-Marín, A.; Bastida, J.M. Evaluating factors that predict the structure of a commensalistic epiphyte-phorophyte network. Proc. Biol. Sci. 2013, 280. [Google Scholar] [CrossRef] [PubMed]
- Wagner, K.; Bogusch, W.; Zotz, G. The role of the regeneration niche for the vertical stratification of vascular epiphytes. J. Trop. Ecol. 2013, 29, 277–290. [Google Scholar] [CrossRef]
- Souza, F.M.; Gandolfi, S.; Rodrigues, R.R. Deciduousness Influences the Understory Community in a Semideciduous Tropical Forest. Biotropica 2014, 46, 512–515. [Google Scholar] [CrossRef]
- Environment Systems Research Institute. ArcGis—ArcView 9; Environment Systems Research Institute: Redlands, CA, USA, 2008. [Google Scholar]
- Mcgarigal, K. Landscape pattern metrics. In Encyclopaedia of Environmentrics; El-Shaarawi, A.H., Piegorsch, W.W., Eds.; John Wiley & Sons: Sussex, UK, 2002. [Google Scholar]
- Rempel, R.S.; Kaukinen, D.; Carr, A.P. Patch Analyst and Patch Grid. Ontario Ministry of Natural Resources; Centre for Northern Forest Ecosystem Research: Thunder Bay, Ontario, ON, Canada, 2012. [Google Scholar]
- Bianchi, J.S.; Kersten, R.D.A.; Araucaria, T.; Forest, A. Edge effect on vascular epiphytes in a subtropical Atlantic Forest. Acta Bot. Bras. 2014, 28, 120–126. [Google Scholar] [CrossRef]
- Laurance, W.F.; Yensen, E. Predicting the impacts of edge effects in fragmented habitats. Biol. Conserv. 1991, 55, 77–92. [Google Scholar] [CrossRef]
- Cadenasso, M.L.; Traynor, M.M.; Pickett, S.T.A. Functional location of forest edges: Gradients of multiple physical factors. Can. J. Forest Res. 1997, 27, 774–782. [Google Scholar] [CrossRef]
- Harper, K.A.; Macdonald, S.E. Quantifying distance of edge influence: A comparison of methods and a new randomization method. Ecosphere 2011, 2, 1–17. [Google Scholar] [CrossRef]
- RStudio: Integrated Development Environment for R; Version 0. 98.1091; RStudio: Boston, MA, USA, 2015.
- Fox, J.; Weisberg, S. An {R} Companion to Applied Regression, 2nd ed.; Sage: Thousand Oaks, CA, USA, 2011. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2009. [Google Scholar]
- Baselga, A.; Orme, C.D.L. betapart: An R package for the study of beta diversity. Methods Ecol. Evol. 2012, 3, 808–812. [Google Scholar] [CrossRef]
- Leibold, M.A.; Holyoak, M.; Mouquet, N.; Amarasekare, P.; Chase, J.M.; Hoopes, M.F.; Holt, R.D.; Shurin, J.B.; Law, R.; Tilman, D.; et al. The metacommunity concept: A framework for multi-scale community ecology. Ecol. Lett. 2004, 7, 601–613. [Google Scholar] [CrossRef]
- Flores-Palacios, A.; García-Franco, J.G. The relationship between tree size and epiphyte species richness: Testing four different hypotheses. J. Biogeogr. 2006, 33, 323–330. [Google Scholar] [CrossRef]
- Taylor, A.; Burns, K. Epiphyte community development throughout tree ontogeny: An island ontogeny framework. J. Veg. Sci. 2015, 26, 902–910. [Google Scholar] [CrossRef]
- Günter, S.; Weber, M.; Erreis, R.; Aguirre, N. Influence of forest edges on the regeneration of abandoned pastures in the tropical mountain rain forest of Southern Ecuador. Eur. J. Forest Res. 2007, 126, 67–75. [Google Scholar] [CrossRef]
- Ruiz-Cordova, J.P.; Toledo-Hernández, V.H.; Flores-Palacios, A. The effect of substrate abundance in the vertical stratification of Bromeliad epiphytes in a tropical dry forest (Mexico). Flora 2014, 209, 375–384. [Google Scholar] [CrossRef]
- Wagner, K.; Mendieta-Leiva, G.; Zotz, G. Host specificity in vascular epiphytes: A review of methodology, empirical evidence and potential mechanisms. AoB Plants 2015. [Google Scholar] [CrossRef] [PubMed]
- Qian, H.; Ricklefs, R.E.; White, P.S. Beta diversity of angiosperms in temperate floras of eastern Asia and eastern North America. Ecol. Lett. 2005, 8, 15–22. [Google Scholar] [CrossRef]
- Dodson, C.H. Why Are There So Many Orchid Species? Lankesteriana 2003, 7, 99–103. [Google Scholar] [CrossRef]
- Jost, L. Explosive Local Radiation of the Genus Teagueia (Orchidaceae) in the Upper Pastaza Watershed of Ecuador. Lyonia 2004, 7, 41–47. [Google Scholar]
- Perez-Escobar, O.; Parra Sanchez, E.; Valdivieso, O. Inventario orquideológico de la Reserva Bosque de Yotoco Valle del Cauca. Acta Agron. 2009, 58, 189–196. [Google Scholar]
- Larrea, L.; Werner, F. Response of vascular epiphyte diversity to different land-use intensities in a neotropical montane wet forest. Forest Ecol. Manag. 2010, 260, 1950–1955. [Google Scholar] [CrossRef]
- Flores-Palacios, A.; García-Franco, J.G. Habitat isolation changes the beta-diversity of the vascular epiphyte community in lower montane forest, Veracruz, Mexico. Biodivers. Conserv. 2008, 17, 191–207. [Google Scholar] [CrossRef]
- Ewers, R.; Banks-Leite, C. Fragmentation impairs the microclimate buffering effect of tropical forests. PLoS ONE 2013, 8, e58093. [Google Scholar] [CrossRef] [PubMed]
- Etter, A.; McAlpine, C.; Possingham, H. Historical patterns and drivers of landscape change in Colombia since 1500: A regionalized spatial approach. Ann. Assoc. Am. Geogr. 2008, 98, 2–23. [Google Scholar] [CrossRef]
- Rodríguez Eraso, N.; Armenteras, D.; Alumbreros, J.R. Land use and land cover change in the Colombian Andes: Dynamics and future scenarios. J. Land Use Sci. 2013, 8, 154–174. [Google Scholar] [CrossRef]
- Ledo, A.; Montes, F.; Condes, S. Species dynamics in a montane cloud forest: Identifying factors involved in changes in tree diversity and functional characteristics. For. Ecol. Manag. 2009, 258, 75–84. [Google Scholar] [CrossRef]
- Didham, R.K.; Kapos, V.; Ewers, R. Rethinking the conceptual foundations of habitat fragmentation research. Oikos 2012, 121, 161–170. [Google Scholar] [CrossRef]
- Rasmussen, H.N.; Dixon, K.W.; Jersáková, J.; Tesitelova, T. Germination and seedling establishment in orchids: A complex of requirements. Ann. Bot. Lond. 2015, 116, 391–402. [Google Scholar] [CrossRef] [PubMed]
- Morales, M.; Armenteras, D. Estado De Conservación De Los Bosques De Niebla De Los Andes Colombianos, Un Análisis Multiescalar. Bol. Cient. Mus. Hist. Nat. 2013, 17, 64–72. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reserve | Coordinates | Geographical Situation | Altitude (m.a.s.l.) | Surface Area (m2) | Perimeter (m) |
|---|---|---|---|---|---|
| Arenillo | 3°29′31.88″ N 76°09′55.44″ O | Central Cordillera | 2015–2350 | 985.000 | 4344.6 |
| Sevilla | 4°12′23.37″ N 75°55’03.27″ O | Central Cordillera | 2011–2378 | 255.000 | 8093.7 |
| La Iberia | 4°04′06.73″ N 76°05´18.39″ O | Central Cordillera | 1950–2065 | 12.300 | 1568.4 |
| Dapa | 3°32′51.89″ N 76°35′12.29″ O | Western Cordillera | 1950–2210 | 92.000 | 1313.5 |
| Yotoco | 3°49′25.32″ N 76°25′59.03″ O | Western Cordillera | 1880–2160 | 149.000 | 1838.4 |
| Roldanillo | 4°25′56.46″ N 76°12′35.24″ O | Western Cordillera | 2050–2100 | 193.000 | 2604.8 |
| Patch Geometry | |||
|---|---|---|---|
| Variable | Description | Code | Type |
| Area | Class area (ha) | CA | CONT |
| Shape | Mean Shape Index | MSI | CONT |
| Mean Patch Fractal Dimension | MPFD | CONT | |
| Mean Patch Area Perimeter Ratio | MPAR | CONT | |
| Edge | Total edge (m) | TE | CONT |
| Edge density m (ha) | ED | CONT | |
| Forest structure | |||
| Density | Number of trees per hectare | DN | CONT |
| Canopy cover | Estimation of the percentage (%) of canopy cover of each tree | Cover | CONT |
| Height | Estimation of the height of each tree at each point in meters | H | CONT |
| Basal area | Quantification of the basal area at DBH for every tree in cm | BA | CONT |
| Neighbouring cover type | |||
| Pasture | Plots randomly located at the boundary within the fragments and the neighboring cover of pasture dominant of herbaceaus of the botanical families Cyperaceae, Poaceae, y Fabaceae, typically below 100 cm height. | P | CAT |
| Regeneration | Plots randomly located at the boundary within the fragment and the neighboring cover characterized by low tree density, high distance among trees, low basal area, and low height. | R | CAT |
| Interior | Located in the centroid of the patch, 100 m away from the edge, where is expected a low edge effect | I | CAT |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parra Sánchez, E.; Armenteras, D.; Retana, J. Edge Influence on Diversity of Orchids in Andean Cloud Forests. Forests 2016, 7, 63. https://doi.org/10.3390/f7030063
Parra Sánchez E, Armenteras D, Retana J. Edge Influence on Diversity of Orchids in Andean Cloud Forests. Forests. 2016; 7(3):63. https://doi.org/10.3390/f7030063
Chicago/Turabian StyleParra Sánchez, Edicson, Dolors Armenteras, and Javier Retana. 2016. "Edge Influence on Diversity of Orchids in Andean Cloud Forests" Forests 7, no. 3: 63. https://doi.org/10.3390/f7030063