Early Detection and Identification of the Main Fungal Pathogens for Resistance Evaluation of New Genotypes of Forest Trees

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Phytopathological Analysis
2.3. Species-Specific Molecular Genetic Identification of Phytopathogens
2.4. DNA Isolation and PCR Amplification
2.5. Gel Electrophoresis
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Schlarbaum, S.E.; Hebard, F.; Spaine, P.C.; Kamalay, J.C. Three American tragedies: Chestnut blight, butternut canker, and Dutch elm disease. In Proceedings of the Exotic Pests of Eastern Forests Conference, Nashville, TN, USA, 8–10 April 1997; pp. 45–54. [Google Scholar]
- Soularue, J.-P.; Robin, C.; Desprez-Loustau, M.-L.; Dutech, C. Short rotations in forest plantations accelerate virulence evolution in root-rot pathogenic fungi. Forests 2017, 8, 205. [Google Scholar] [CrossRef]
- Stephanson, C.A.; Ribarik Coe, N. Impacts of beech bark disease and climate change on American beech. Forests 2017, 8, 155. [Google Scholar] [CrossRef]
- Liu, Q.; Luo, L.; Zheng, L. Lignins: Biosynthesis and biological functions in plants. Int. J. Mol. Sci. 2018, 19, 335. [Google Scholar] [CrossRef] [PubMed]
- Rischer, H.; Oksman-Caldentey, K.-M. Unintended effects in genetically modified crops: Revealed by metabolomics? Trends Biotechnol. 2006, 24, 102–104. [Google Scholar] [CrossRef] [PubMed]
- Ladics, G.S.; Bartholomaeus, A.; Bregitzer, P.; Doerrer, N.G.; Gray, A.; Holzhauser, T.; Jordan, M.; Keese, P.; Kok, E.; Macdonald, P.; et al. Genetic basis and detection of unintended effects in genetically modified crop plants. Transgenic Res. 2015, 24, 587–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Den Herder, M.; Kouki, J.; Ruusila, V. The effects of timber harvest, forest fire, and herbivores on regeneration of deciduous trees in boreal pine-dominated forests. Can. J. For. Res. 2009, 39, 712–722. [Google Scholar] [CrossRef]
- Hynynen, J.; Niemiströ, P.; Viherä-Aarnio, A.; Brunner, A.; Hein, S.; Velling, P. Silviculture of birch (Betulapendula Roth and Betulapubescens Ehrh.) in northern Europe. Forestry 2010, 83, 103–119. [Google Scholar] [CrossRef]
- West, P.W. Choosing the species and site. In Growing Forest Plantations; West, P.W., Ed.; Springer International Publishing: Basel, Switzerland, 2014; pp. 45–54. ISBN 978-3-319-01826-3. [Google Scholar]
- Shcherbakova, L.A. Advanced methods of plant pathogen diagnostics. In Comprehensive and Molecular Phytopathology, 1st ed.; Dyakov, Y.T., Dzhavakhiya, V., Korpela, T., Eds.; Elsevier: Amsterdam, The Netherlands, 2007; pp. 75–116. ISBN 978-0-444-52132-3. [Google Scholar]
- Bilodeau, G.J.; Pelletier, G.; Pelletier, F.; Levesque, C.A.; Hamelin, R.C. Multiplex real-time polymerase chain reaction (PCR) for detection of Phytophthoraramorum, the causal agent of sudden oak death. Can. J. Plant Pathol. 2009, 31, 195–210. [Google Scholar] [CrossRef]
- Pilotti, M.; Lumia, V.; Di Lernia, G.; Brunetti, A. Development of real-time PCR for in wood-detection of Ceratocystisplatani, the agent of canker stain of Platanus spp. Eur. J. Plant Pathol. 2012, 134, 61–79. [Google Scholar] [CrossRef]
- Luchi, N.; Oliveira Longa, C.M.; Danti, R.; Capretti, P.; Maresi, G. Diplodiasapinea: The main fungal species involved in the colonization of pine shoots in Italy. For. Pathol. 2014, 44, 372–381. [Google Scholar] [CrossRef]
- Liu, B.; Zhang, Y.; He, D.; Li, Y. Identification of apple leaf diseases based on deep convolutional neural networks. Symmetry 2018, 10, 11. [Google Scholar] [CrossRef]
- Bakonyi, J.; Nagy, Z.Á.; Rsek, T.É. PCR-based DNA markers for identifying hybrids within Phytophthoraalni. J. Phytopathol. 2006, 154, 168–177. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.D.; Lee, S.B.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. ISBN 978-0123721815. [Google Scholar]
- Elder, J.F., Jr.; Turner, B.J. Concerted evolution of repetitive DNA sequences in eukaryotes. Q. Rev. Biol. 1995, 70, 297–320. [Google Scholar] [CrossRef] [PubMed]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc. Natl. Acad. Sci. USA. 2012, 109, 6241–6246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arteau, M.; Labrie, S.; Roy, D. Terminal-restriction fragment length polymorphism and automated ribosomal intergenic spacer analysis profiling of fungal communities in Camembert cheese. Int. Dairy J. 2010, 20, 545–554. [Google Scholar] [CrossRef]
- Callon, C.; Delbes, C.; Duthoit, F.; Montel, M.C. Application of SSCP-PCR fingerprinting to profile the yeast community in raw milk Salers cheeses. Syst. Appl. Microbiol. 2006, 29, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Gori, K.; Ryssel, M.; Arneborg, N.; Jespersen, L. Isolation and identification of the microbiota of Danish farmhouse and industrially produced surface-ripened cheeses. Microb. Ecol. 2013, 65, 602–615. [Google Scholar] [CrossRef] [PubMed]
- Padutov, V.E.; Baranov, O.Y.; Voropaev, E.V. Molecular Genetic Analysis Methods; Unipol: Minsk, Belarus, 2007; p. 176. ISBN 978-985-6768-12-8. [Google Scholar]
- Rogers, S.O.; Bendich, A.J. Extraction of total cellular DNA from plants, algae and fungi. In Plant Molecular Biology Manual; Gelvin, S.B., Schilperoort, R.A., Eds.; Springer: Heidelberg, Netherlands, 1994; pp. 1–8. ISBN 978-94-011-7654-5. [Google Scholar]
- Panteleev, S.V. Molecular genetic diagnosis and identification of fungal pathogens of planting material of tree species in forest nurseries of Belarus. Ph.D. Thesis, Forest Institute, Gomel, Republic of Belarus, 2013. (In Russian). [Google Scholar]
- Poteri, M. Screening of Birch, Betula spp., for Rust Resistance to Melampsoridiumbetulinum; Research Papers; Finnish Forest Research Institute: Helsinki, Finland, 1998; Volume 689, p. 44. [Google Scholar]
- Jalkanen, R. Synthesis and new observations on needle pathogens of larch in Northern Finland. Forests 2016, 7, 25. [Google Scholar] [CrossRef]
- Organisation for Economic Co-operation and Development (OECD). Consensus document on the biology of European White Birch (Betulapendula Roth). In Series on Harmonisation of Regulatory Oversight in Biotechnology; No. 28; OECD Environment Directorate: Paris, France, 2003; p. 46. [Google Scholar]
- Phillips, D.H.; Burdekin, D.A. Diseases of ash (Fraxinus spp.), birch (Betula spp.) and alder (Alnus spp.). In Diseases of Forest and Ornamental Trees; Phillips, D.H., Burdekin, D.A., Eds.; Palgrave Macmillan: London, UK, 1992; pp. 284–298. ISBN 978-1-349-10955-5. [Google Scholar]
- Baranov, O.Y.; Panteleev, S.V. Molecular genetic assay of rDNA loci of main forest tree species pathogens in Belarus. Challenge For. 2012, 72, 220–223. (In Russian) [Google Scholar]
- Broders, K.; Munck, I.; Wyka, S.; Iriarte, G.; Beaudoin, E. Characterization of fungal pathogens associated with white pine needle damage (WPND) in Northeastern North America. Forests 2015, 6, 4088–4104. [Google Scholar] [CrossRef]
- Hauptman, T.; Ogris, N.; de Groot, M.; Piškur, B.; Jurc, D. Individual resistance of Fraxinus angustifolia clones to ash dieback. For. Pathol. 2016, 46, 269–280. [Google Scholar] [CrossRef]
- Cleary, M.R.; Blomquist, M.; Vetukuri, R.R.; Bohlenius, H.; Witzell, J. Susceptibility of common tree species in Sweden to Phytophthora cactorum, P. cambivora and P. plurivora. For. Pathol. 2017, 47, e12329. [Google Scholar] [CrossRef]
- Ding, T.; Palmer, M.W.; Melcher, U. Community terminal restriction fragment length polymorphisms reveal insights into the diversity and dynamics of leaf endophytic bacteria. BMC Microbiol. 2013, 13, 1. [Google Scholar] [CrossRef] [PubMed]
- Kovalitskaya, Y.A.; Dayanova, L.K.; Azarova, A.B.; Shestibratov, K.A. RNA interference-mediated down-regulation of 4-coumarate: coenzyme A ligase in Populus tremula alters lignification and plant growth. Int. J. Envir. Sci. Educat. 2016, 11, 12259–12271. [Google Scholar]



{kind=link}
{kind=link}
| Locus | Primer | Primer Sequence (5′–3′) |
|---|---|---|
| ITS1 | ITS1 ITS2 | FAM-TCCGTAGGTGAACCTGCGG GCTGCGTTCTTCATCGATGC |
| ITS2 | ITS3 ITS4 | FAM-GCATCGATGAAGAACGCAGC TCCTCCGCTTATTGATATGC |
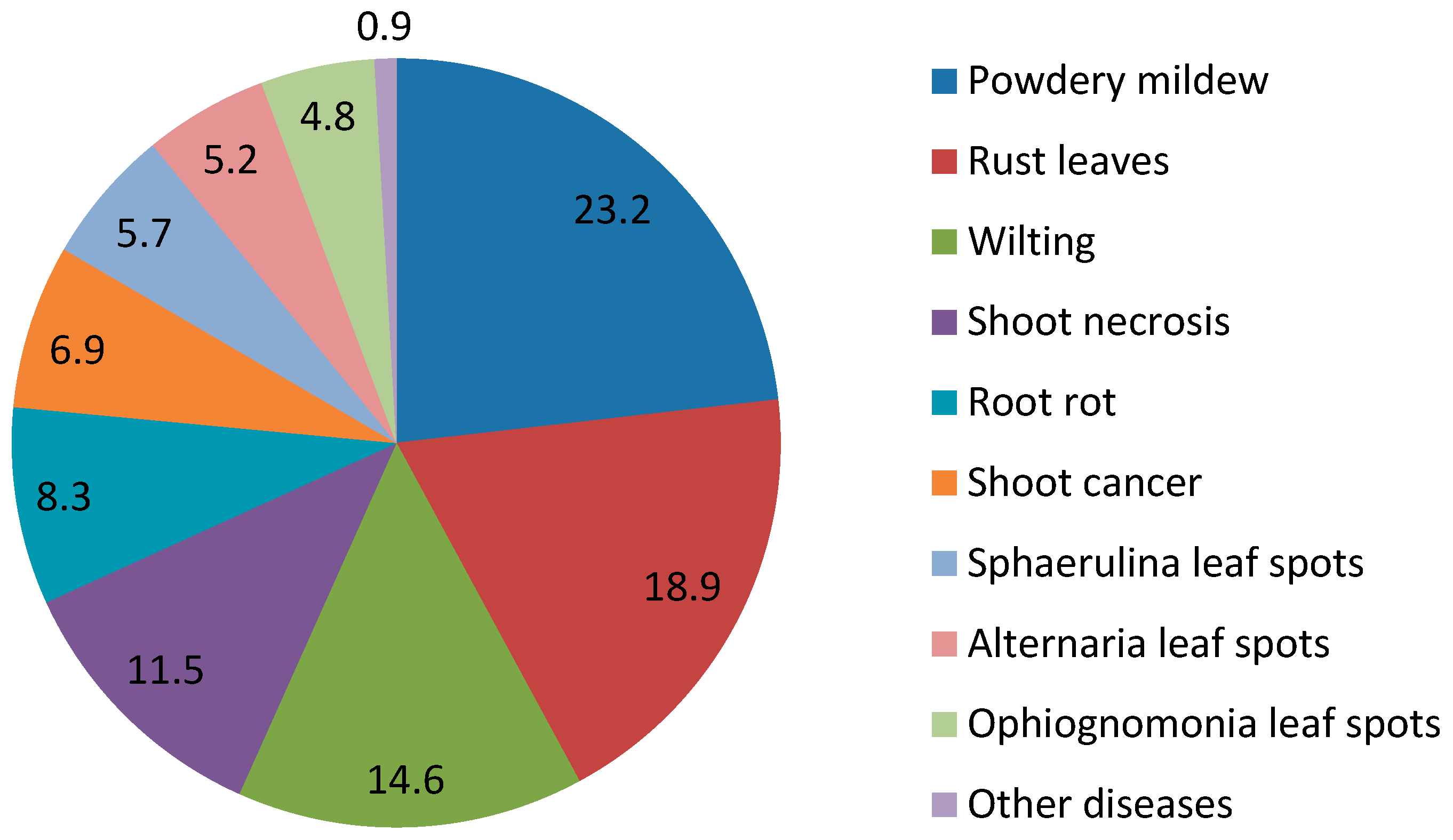
| Phytopathogen | Disease Type | Occurrence, % |
|---|---|---|
| Phyllactinia guttata (Wallr.) Lev. | Powdery mildew | 13.2 |
| Erysiphe ornate (U. Braun) U. Braun & S. Takam. | Powdery mildew | 10.0 |
| Melampsoridium betulinum (Pers.) Kleb. | Rust leaves | 18.9 |
| Fusarium avenaceum (Fr.) Sacc. | Wilting | 13.2 |
| Nectria sp. (Fr.) Fr. | Shoot necrosis | 4.1 |
| Melanconium bicolor Nees. | Shoot necrosis | 3.7 |
| Phytophthora cactorum (Leb. & Cohn) Schroeter | Shoot necrosis | 3.5 |
| Pythium sp. Pringsheim | Root rot | 7.6 |
| Botryosphaeria dothidea (Moug. & Fr.) Ces. & DeNot. | Shoot cancer | 6.2 |
| Ophiognomonia intermedia (Rehm) Sogonov | Ophiognomonia leaf spots | 4.8 |
| Sphaerulina betulae (Pass.) Quaedvlieg, Verkley & Crous | Sphaerulina leaf spots | 5.7 |
| Alternaria alternata (Fr.) Keissl. | Alternaria leaf spots | 4.9 |
| Other species | Other diseases | 0.9 |
| Phytopathogenic Species | ITS1–ITS2, bp | ITS3–ITS4, bp |
|---|---|---|
| Sphaerulina betulae (Pass.) Quaedvlieg, Verkley & Crous | 225 | 231 |
| Ophiognomonia intermedia (Rehm) Sogonov | 268 | 351 |
| Alternaria alternata (Fr.) Keissl. | 244 | 346 |
| Phyllactinia guttata (Wallr.) Lev. | 314 | 364 |
| Botryosphaeria dothidea (Moug. & Fr.) Ces. & DeNot. | 259 | 344 |
| Erysiphe ornate (U. Braun) U. Braun & S. Takam. | 298 | 362 |
| Melampsoridium betulinum (Pers.) Kleb. | 328 | 406 |
| Pythium sp. Pringsheim | 298 | 633 |
| Phytophthora cactorum (Leb. & Cohn) Schroeter | 295 | 602 |
| Fusarium avenaceum (Fr.) Sacc. | 233 | 355 |
| Melanconium bicolor Nees. | 270 | 349 |
| Nectria sp. (Fr.) Fr. | 217 | 348 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shestibratov, K.A.; Baranov, O.Y.; Subbotina, N.M.; Lebedev, V.G.; Panteleev, S.V.; Krutovsky, K.V.; Padutov, V.E. Early Detection and Identification of the Main Fungal Pathogens for Resistance Evaluation of New Genotypes of Forest Trees. Forests 2018, 9, 732. https://doi.org/10.3390/f9120732
Shestibratov KA, Baranov OY, Subbotina NM, Lebedev VG, Panteleev SV, Krutovsky KV, Padutov VE. Early Detection and Identification of the Main Fungal Pathogens for Resistance Evaluation of New Genotypes of Forest Trees. Forests. 2018; 9(12):732. https://doi.org/10.3390/f9120732
Chicago/Turabian StyleShestibratov, Konstantin A., Oleg Yu. Baranov, Natalya M. Subbotina, Vadim G. Lebedev, Stanislav V. Panteleev, Konstantin V. Krutovsky, and Vladimir E. Padutov. 2018. "Early Detection and Identification of the Main Fungal Pathogens for Resistance Evaluation of New Genotypes of Forest Trees" Forests 9, no. 12: 732. https://doi.org/10.3390/f9120732