Ungulate Browsing Limits Bird Diversity of the Central European Hardwood Floodplain Forests


1
Department of Development and Environmental Studies, Faculty of Science, Palacky University Olomouc, 77147 Olomouc, Czech Republic
2
Department of Forest Protection and Wildlife Management, Faculty of Forestry and Wood Technology, Mendel University in Brno, Zemedelska 3, 61300 Brno, Czech Republic
3
Department of Geoinformatics, Faculty of Science, Palacky University Olomouc, 77147 Olomouc, Czech Republic
*
Author to whom correspondence should be addressed.
Forests 2018, 9(7), 373; https://doi.org/10.3390/f9070373
Submission received: 20 April 2018
/
Revised: 8 June 2018
/
Accepted: 13 June 2018
/
Published: 21 June 2018
(This article belongs to the Special Issue Hardwood Reforestation and Restoration)
Abstract
:Temperate hardwood floodplain forests along lowland rivers are considered important forest biodiversity refugia in the European cultural landscape. The absence of apex predators combined with an artificial feeding of herbivore populations in winter seasons has caused an increase in browsing pressure on hardwood trees, nearly preventing their regeneration in some localities. There are still important knowledge gaps in understanding the relationships between deer abundance (and browsing pressure) and the abundance (and diversity) of forest bird species in unmanaged hardwood forests. We have studied the red deer and fallow deer browsing pressure in Central European unmanaged hardwood floodplain forests using a novel method based on monitoring browsing pressure along transects combined with bird census data in the Litovelské Pomoraví Protected Landscape Area (Czech Republic). The monitoring data suggested a very high browsing pressure on hardwood trees, causing a strong reduction of the shrub layer and young tree layer (30–210 cm above ground surface). The bird census data from the study area were collected using the territory mapping method. Our results revealed a bird diversity decline in all study plots and the bush nesters guild was found to be completely absent. As bird species from the bush nesters guild are generally common (usually dominant) in hardwood floodplain forest ecosystems with a rich shrub and young tree layer and low browsing pressure, we conclude that intense browsing by large herbivores represents a limiting factor to the bird diversity (especially bush nesters) of hardwood floodplain forests.
1. Introduction
Forest vegetation structure influences the diversity of forest avian communities [1,2,3,4,5]. The habitat characteristics of floodplain forests, modified by human activities, have influenced the bird diversity [6]. This is especially important at the local scale of riparian forest stands, because bird density has a direct relationship with site-scale resources, as pointed out by Zenzal et al. [7]. Bird species richness in hardwood floodplain forests differs among the habitat types, with mature forests supporting the largest number of species because of high stand heterogeneity based on diverse understorey tall shrubs and young trees [8] Natural forest regeneration, the presence or absence of understorey bush and herb cover, and even the general structure of forest ecosystems in the European hardwood temperate forests are affected by browsing pressure of large herbivores, typically deer [9]. The absence of large predators in hardwood forests of the European cultural landscape aggravates the effects of browsing pressure on forest biodiversity [10]. Ungulate browsing can interact with the local flooding regime of rivers to delay the recruitment of some tree species, resulting in shifts in successional trajectories, and leaving young forests vulnerable to invasion by exotic herbaceous species [11].
As shown in the study of Holt et al. [12], deer exclusion benefits birds which forage in the understorey layer. Several guilds or migrant species responded positively to deer exclusion and none responded negatively. The shrub-layer foraging guild was recorded less frequently in older and browsed vegetation, in both winter and spring. Exclusion of deer also increased the occurrence of ground-foraging species in both seasons, although these species showed no strong response to vegetation age. Newson et al. [13] have shown a strong association between deer densities and declines in understorey bird species. Their results indicate that deer-related habitat modification may be affecting some bird species on far larger scales than previously appreciated. Mainly through their effects on understorey vegetation, high deer populations are now likely to be affecting woodland biodiversity over large parts of lowland England and deer management plans, involving the integrated exclusion and culling of deer, need to be coordinated on large scales. It is suggested that such management plans could most usefully target areas that still support relatively high populations of species that are sensitive to deer. The density of understorey foliage is recognized as an important predictor of the distribution of forest birds [14]. Charchuk and Banes [15] have suggested that following understorey protection harvest, the retained forest regenerates quickly, rapidly providing a habitat to more mature forest species than natural disturbance harvest.
The understorey foliage can be seriously reduced by a high deer abundance, as Eichhorn et al. [16] revealed by a LiDAR survey. Their findings suggest that the reduction of deer populations is likely to have a strong impact on woodland structures and aid in restoring the complex understorey habitats required by many birds, whereas management interventions as currently practiced have limited and inconsistent effects. Thus, LiDAR seems to be a potentially important tool for forest bird conservation as it can help identify the full range of structural conditions associated with threshold responses [17].
Forest management decisions in hardwood forests should be made at a site level, and encompass factors such as browsing pressure and the dependence of species of conservation concern on particular habitats [18]. There are still important knowledge gaps in understanding relationships between deer abundance (and browsing pressure) and the abundance (and diversity) of forest bird species in unmanaged forests [19,20]. In order to address these knowledge gaps, we studied the impact of browsing pressure by red deer and fallow deer on avian communities in unmanaged hardwood floodplain forests with a high deer density in the Litovelské Pomoraví Protected Landscape Area, Czech Republic [21]. The main objective of this paper is to provide evidence of the relationship between deer browsing and bird diversity in hardwood floodplain forest ecosystems. We hypothesized that intensive ungulate browsing can be a significant limiting factor to bird diversity in hardwood floodplain forest in protected areas.
2. Materials and Methods
2.1. Study Area
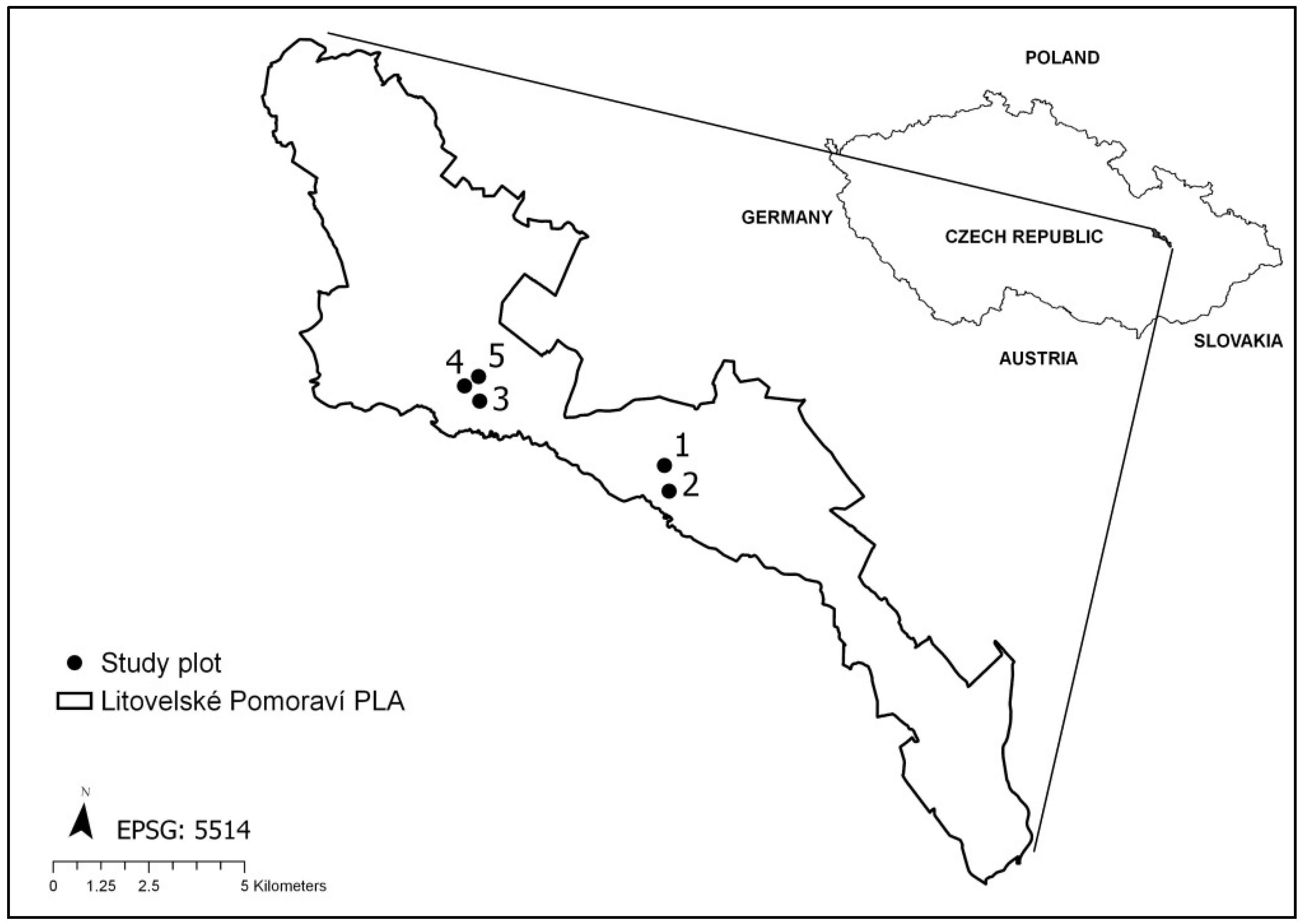
The study area—Litovelske Pomoravi Protected Landscape Area (LPPLA)—is formed by a large segment of hardwood floodplain forests in the eastern part of the Czech Republic along the lowland (240–249 m a.s.l.) meandering Morava River [22]. According to the Czech national classification of forest habitats, the hardwood floodplain forests in the study area [23] are classified as Ulmi-fraxineta carpini superior [24]. According to the European classification of forest natural habitats under the Natura 2000 network [25], they are classified as riparian mixed forests along the great rivers (habitat code 91F0). The dominant species are Quercus robur L. and Fraxinus excelsior L., with admixture of Tilia cordata Mill., Acer campestre L., Acer pseudoplatanus L., Acer platanoides L., Carpinus betulus L., Ulmus laevis Pallas, and Prunus padus L.
2.2. Bird Census Data
In each study plot, birds were counted during nesting seasons (from the end of March to the end of June) in 2001, 2003, 2005, and 2010 using the ‘territory mapping’ method [28]. The field mapping involved seven to 10 repeated visits to each study plot. Only those birds were counted that were spotted no further than 50 m from the actual position of the surveying researcher to avoid mistakes based on the different detectability of birds in hardwood floodplain forests [29]. In order to obtain precise bird census results, we also searched all tree cavities and holes for bird nests in each of the study plots. To allow comparisons with other published studies, we used the field data to calculate the mean density (nesting pairs/10 ha). We excluded from our analyses bird species with obviously no relationship with the forest habitat, and which were observed only occasionally in study plots. Classification of bird species into four nesting guilds (ground nesters, bush nesters, canopy nesters, and tree hole nesters) was carried out a priori [30] based on field experience [31] and supported by relevant Czech ornithological literature [32]. Dominance values were calculated according to Aulak [33].
2.3. Ungulate Browsing Research
We established transects in all five study plots to assess herbivore browsing. All transects were 3 m wide and 30–70 m long (based on site conditions). The transect method is routinely used for unrepeated assessments of shrub layer density and browsing intensity [34,35].
The abundance of individual ungulates (IND) according to game management records (GMR 2003) was 63 IND/1000 ha for Capreolus capreolus and 11 IND/1000 ha for Dama dama in the study plots 1 and 2. In the study plots 3, 4, and 5, the abundance was higher: 98 IND/1000 ha for Capreolus capreolus and 63 IND/1000 ha for Dama dama [36].
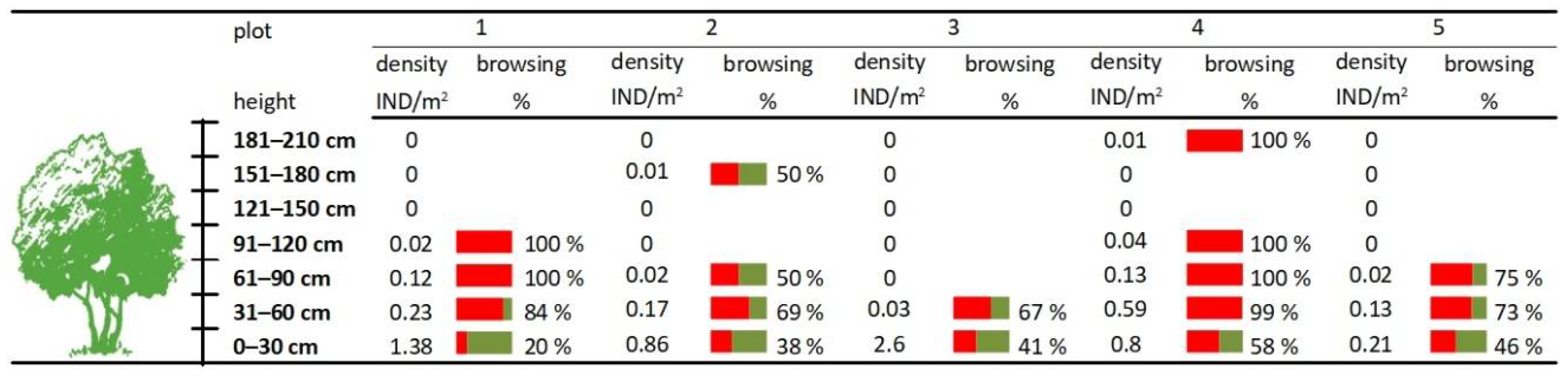
We assessed all trees smaller than 2.1 m. Woody plants higher than 2.1 m were completely absent in the shrub layer in all study plots. The trees were divided into seven height classes (<30 cm, 31–60 cm, 61–90 cm, 91–120 cm, 121–150 cm, 151–180 cm, 181–210 cm). Browsing % is the percentage of individual woody plants that have been browsed. “Individual” was classified as the browsed individual if the terminal shoot was damaged or more than half of the lateral shoots were damaged. The field data collection was conducted in 2003 and it only focused on present browsing damage, i.e., browsing from the past winter (2002/2003) and from the present growing season (2003). In 2005 and 2010, the ungulate browsing in transects was visually verified and found to be without any visible changes.
2.4. Statistical Analysis and Control Plots
To calculate the Pearson’s correlation between shrub layer parameters and browsing intensity and between deer density and bird density, we used the Statistica software (StatSoft s.r.o., Prague, Czech Republic) [37]. Statistically significant differences were compared using t-tests.
To analyse the similarity and diversity of nesting bird communities, we calculated the commonly used Jaccard similarity index [38]. We used rarefaction to standardize species richness of the bird communities in all five study plots and in all of the nesting seasons [39]. Based on rarefaction curves, this method makes it possible to compare the bird species diversity among different study plots and with different survey efforts. The rarefaction curve is constructed based on the expected number of species E(Sn) using the following equation:
where S is the total number of (nesting bird) species found in the study plot, and Ni is the number of nesting pairs of a particular bird species i. We used the EstimateS 8.0.0. software (StatSoft s.r.o., Prague, Czech Republic) to perform the calculations [40].
As control plots for our original results from study plots, we used localities in hardwood floodplain forests in the Czech Republic, where the bird communities had recently been studied by the territory mapping method and from which data were available. Data related to deer densities in these control plots were collected from legal hunting statistics in the archives of Regional Offices in Olomouc, Hradec Kralove, Ostrava, and Plzen.
3. Results
3.1. Ungulate Browsing and Woody Plants in Herb and Shrub Layers
The density of woody plants was relatively high in the 0–30 cm height class. Woody plants higher than 120 cm were absent, except for very low densities in the height classes 150–180 cm (plot 2) and 180–210 cm (plot 4). Beyond height class 0–30 cm, the browsing percentage was greater than 50% (Figure 2). The percentage of damaged woody plants was negatively correlated with the density of woody plants (Table 1).
Young trees strongly outweighed shrubs in herb and shrub layers. The dominant species were Fraxinus excelsior (55.5%), Acer campestre (21.3%), and Acer pseudoplatanus (15.1%). The remaining tree species accounted for only 6.6% (Carpinus betulus, Quercus petraea, Acer platanoides, and Tillia cordata) and shrub species for 1.4% (Swida sanguinea, Euonymus europaeus, and Crataegus laevigata) of the total records.
3.2. Bird Community
A total of 21 bird species have been found nesting in the study plots (Table 2). The species composition of nesting birds in all study plots was very similar (Jaccard coefficient 78.9–95.0%).
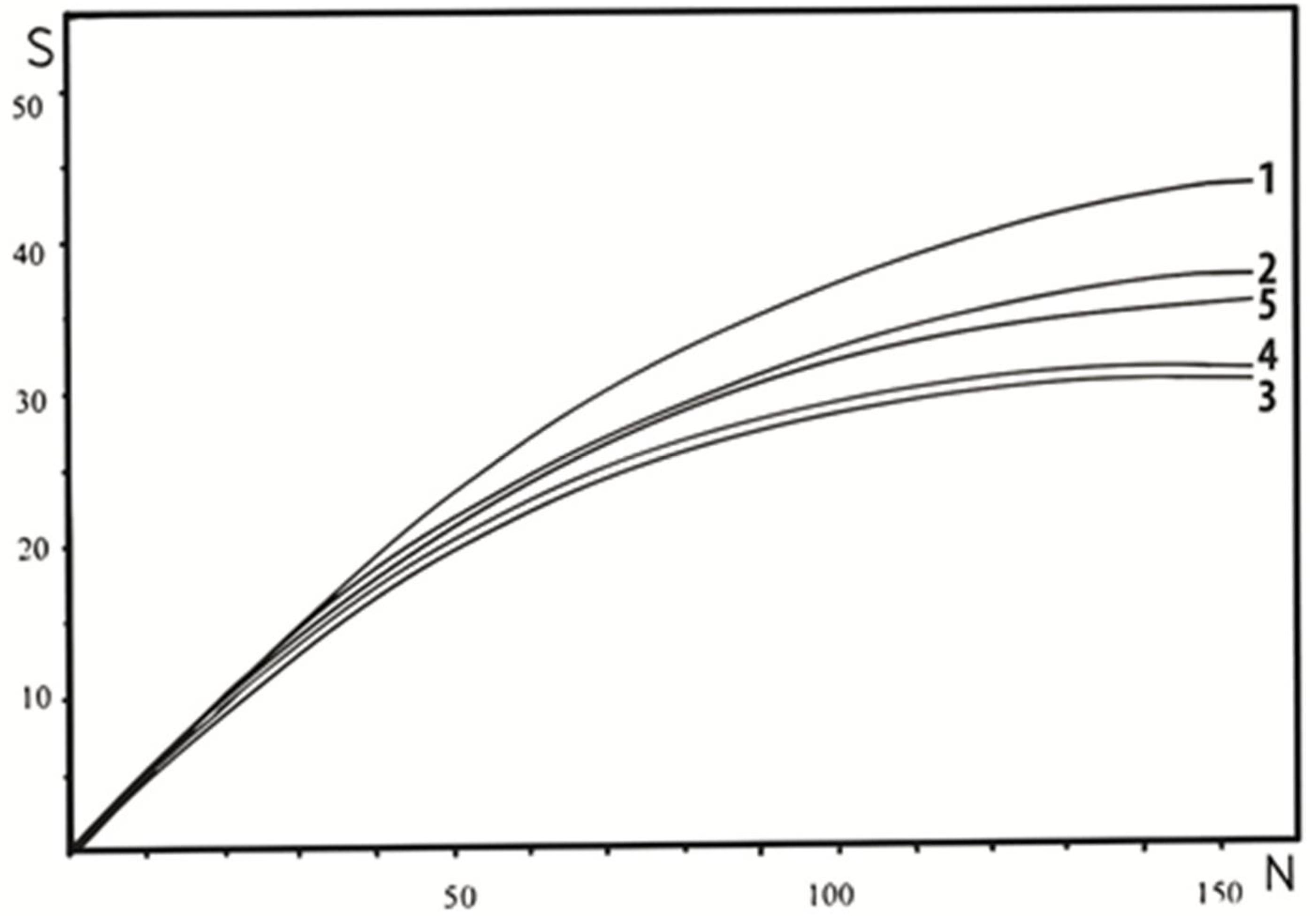
Using the rarefaction method (Figure 3), we found out that for the minimum abundance of nesting pairs, the highest species richness was reached in study plot 1. However, the differences in bird species richness between individual study plots were not visually distinct.
The rarefaction revealed that the efforts in count surveys in all plots were sufficient. The number of species recorded in all study plots was therefore sufficiently large and representative with respect to the method used.
The highest density of nesting birds was recorded in study plot 2 (69.1 pairs/10 ha), and the lowest in study plot 4 (56.7 pairs/10 ha). The densities calculated for all study plots (Table 2) were thus atypically low for the floodplain forest ecosystem. The representation of dominant species in all study plots reflected the general model of species dominance in bird communities of floodplain forests, in which only a few species (Sturnus vulgaris, Parus major) dominate the community, and the remaining species are considered attendant or accessory species. However, the count survey surprisingly revealed low dominance values for a few species (Phylloscopus collybyta, Erithacus rubecula) that usually form the dominant component of the community.
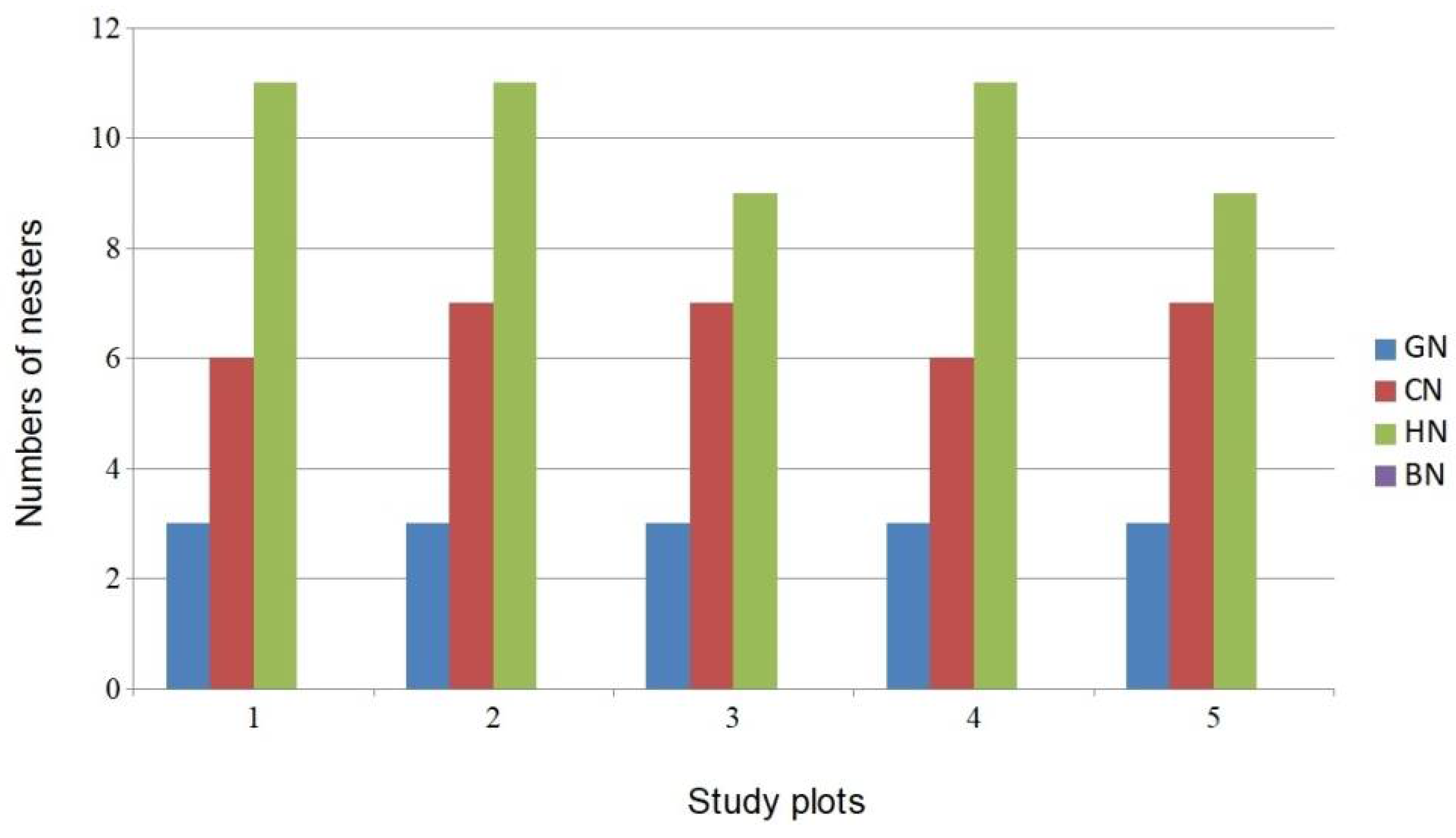
A close examination of the avian guild structure revealed that in all study plots, the bush nesters guild was completely absent (Figure 4). None of the species of this guild typical for the floodplain forests (e.g., Aegithalos caudatus, Coccothraustes, Hippolais icterina, Luscinia megarhynchos, Prunella modularis, Sylvia atricapilla, Troglodytes troglodytes, Turdus merula) were found in any of the study plots.
The bird density values in all our study plots within the LPPLA were significantly lower than the values obtained by other authors using the same mapping method in hardwood floodplain forests in the Czech Republic (Table 3). High negative correlation rates were found between Capreolus capreolus density and bird density (correlation coefficient = −0.6515, p-value = 0.0008) and between both of the deer species together (Capreolus capreolus and Dama dama) and bird density (correlation coefficient = −0.5641, p-value = 0.0051). A moderately high negative correlation rate was detected between Dama dama density and bird density (correlation coefficient = −0.5201, p-value = 0.0269). It is possible that the absence of the bush nesters guild may be the reason for a lower diversity of birds by approximately 30% in our study plots compared to other floodplain forest sites in the Czech Republic (Table 3).
4. Discussion
4.1. Impact of Ungulate Browsing on the Forest Understorey of Floodplain Forests
Browsing greater than 40% repeatedly led to important changes in woody plant composition in similar areas of the Czech Republic. More concretely, browsing led to a significant reduction of the abundance of the most affected species, especially if they had a low abundance [55]. Repeated browsing can significantly affect the forest dynamics [56]. Some authors [57] have observed that intensive ungulate browsing led to a significant prolongation of the regeneration period in mountain forests—new silver fir and Norway spruce were held to the height limit of 1–1.5 m in the long term (more than thirty years). Floodplain forests can generally better compensate for the lost woody-plant biomass than mountain forests, so the browsing has a lower impact on the growth of woody plants in the herb and shrub layers [58]. However, intensive browsing can promote an expansion of invasive herbaceous species. A strong herb competition can then complicate forest restoration in floodplain forests [59].
The shrub layer is usually well developed in the studied type of floodplain forest. In our study plots, however, the shrub layer (woody plants higher than 1 m) was poorly developed (plots 1 to 3 with a maximum of one woody plant individual per 20 m2) or completely absent (plots 4 and 5) (Figure 2). Our opinion is that browsing is the main factor for the low abundance of shrubs. The situation of our study area may be compared with the floodplain forest near the Dyje river (locality Lednice hardwood floodplain forest) with the same type of hardwood forest, but with lower browsing pressure [60]. In Lednice, the percentage of browsing damaged woody plants (up to 150 cm of height) was 32% (more than 50% in our plots) and the relative abundance of shrub species was 9% (1.4% in our plots). The ungulates usually prefer some shrub woody plants (e.g., Swida sanguinea, especially during winter, or Eonymus europaeus) over others. Crataegus laevigata is less attractive, but it is strongly browsed, especially in locations where the woody-plant biomass of herb and shrub layers is limited. Most of the tree species present are also attractive [61].
The negative effect of browsing on shrub species can be intensified by their occurrence in herb layers, as browsers often prefer plant species occurring close to the forest floor [62,63]. The observed extreme browsing pressure was most likely the main factor leading to the absence or reduction of the shrub layer.
4.2. Bird Communities in Floodplain Forests
The avifauna of the LPPLA study area mainly consists of common bird species typical for the European hardwood floodplaorests [64]. Surprisingly, a few species otherwise commonly nesting in the understorey of this type of forest (Sylvia atricapilla, Turdus merula) [65] were found to be absent in all of the five study plots.
The rarefaction curves for all study plots approached their asymptotes and further field surveys would likely not reveal any new species [66]. The species composition of nesting birds was very similar in all study plots (measured by the Jaccard coefficient). These results are in contrast with the findings of Kolecek et al. [67], who have found relatively large differences in species composition between two floodplain-forest sites along the Morava River—the ‘Království’ site had about 30% more species than the ‘Žebračka’ site. As the authors themselves point out, these differences could have been caused by the different sizes of the studied sites, as larger sites can host more area-sensitive species [68]. Alternatively, the differences in species composition could have been caused by differences in environmental heterogeneity between the sites [69]. As a result of an intensive forest management, the ‘Království’ site (unlike the ‘Žebračka’ site) represents a diverse mosaic of various types of floodplain forest (clear-cut areas, young-growth and old-growth stands, forest margins). These consequences of forest management could explain the higher diversity of birds, because the edge effect on the diversity of nesting birds in the floodplain forests is very pronounced [70]. As all our study plots are located within the LPPLA conservation zone, no forest management has been applied in these sites since the establishment of the LPPLA in 1991. Implementation of multiple-use zoning [71] is generally considered as an important support tool for the conservation of forest diversity.
It is commonly known that past forest management of the European floodplain forests is very clearly reflected in the current structure of forest stands [72]. In our study plots, however, the historical development of forest stands was practically identical and therefore does not play an important role [73].
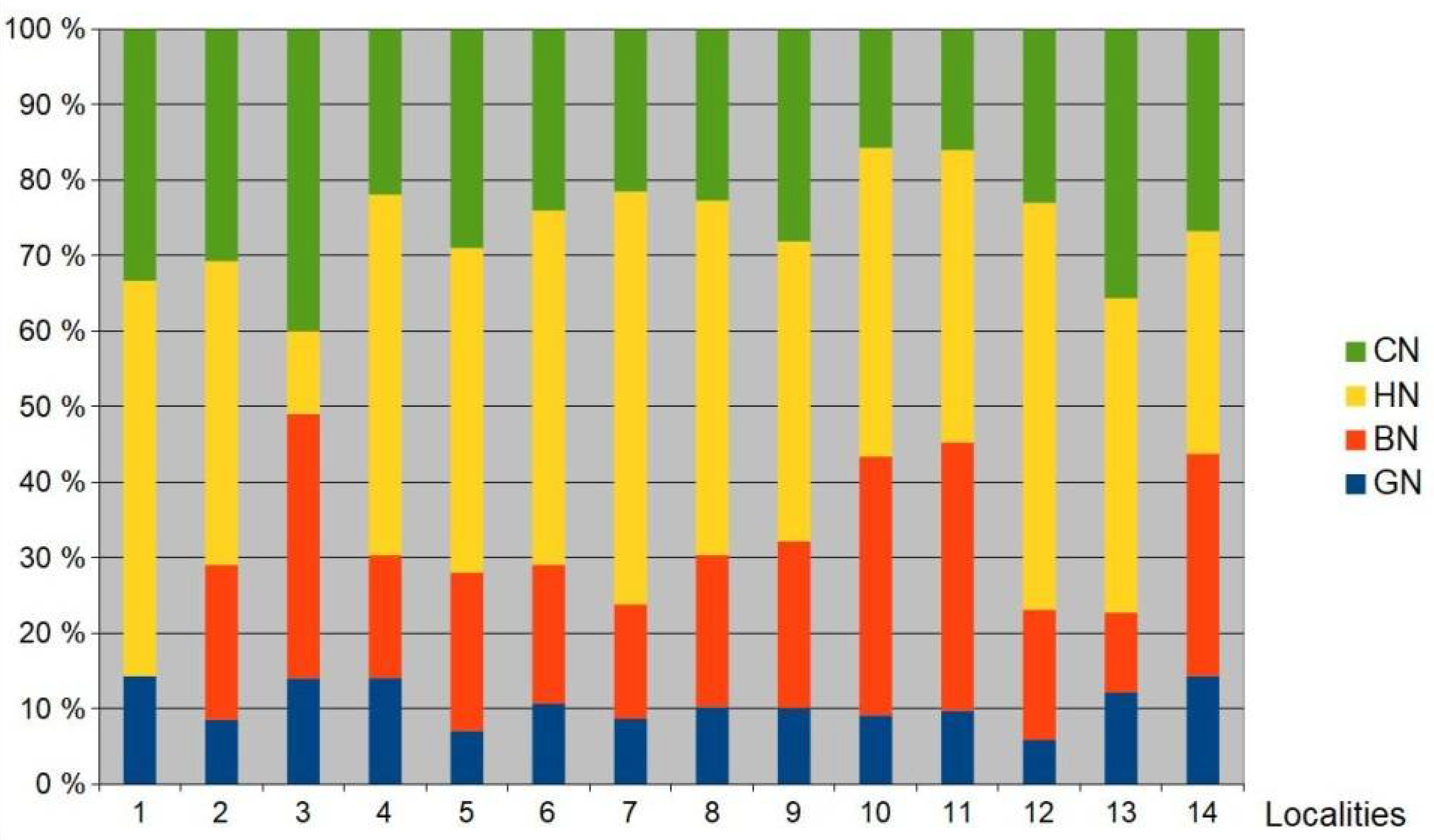
The structure of Central European hardwood forests dominated by oak (Quercus sp.) strictly influences the structure of breeding bird communities, especially in young forest stands [74]. Bird species typical for these stands are bush nesters (Phylloscopus trochilus, Sylvia atricapilla, Sylvia curruca, Prunella modularis, Aegithalos caudatus). In the LPPLA study plots, these bird species were absent. The comparison of guild structures in the LPPLA and in the hardwood floodplain forests in the Czech Republic (Figure 5) reveals a clear absence of common bird species from the bush nesters guild in the LPPLA study plots. Similarly, many studies have shown that the shrub layer is one of the key characteristics determining the alpha diversity and density of bird communities in the European oak-dominated hardwood forests [75,76,77,78,79].
Reductions in understorey foliage strongly indicate a browsing effect in forest areas of a high deer density. Terrestrial laser scanning applied to oak-dominated woods in the UK by Eichhorn et al. [16] revealed a reduction in understorey foliage of 68% at high deer density sites. In the context of the above-mentioned studies, the results from our study plots suggest that intensive ungulate browsing pressure can be considered a limiting factor to forest bird diversity. In our plots, the diversity of nesting birds was approximately 30% lower than the diversity found in a comparable forest type without an intense browsing pressure (Table 3).
Due to the absence of shrubs, our study plots had lower numbers of bird species otherwise commonly inhabiting the European temperate hardwood forests [80]. Although these species are considered neither rare/endangered in the Czech Republic [81] or habitat specialists, the results are significant [82]. As population trends for common forest bird species in the Czech Republic are otherwise remarkably stable in the long term (in contrast to the common bird species of the agricultural landscapes) [83], the intense ungulate browsing pressure in the LPPLA hardwood floodplain forests, reducing the diversity and density of common bird species, represents an extraordinary situation in terms of nature conservation. Here, we would like to point out that all five study plots are located within a protected area specifically aimed at the conservation of hardwood floodplain forest biodiversity [84].
To pursue the conservation targets for protected areas in the ecological conditions of Central European forests with extraordinary high deer densities, in which large predators are missing [85,86], it is necessary to take a step towards an intense targeted reduction of large herbivores by hunting. Understanding the types of relationships between deer abundance and habitat quality for birds, and other biodiversity, is an important knowledge gap that needs to be addressed if sound, collaborative deer management plans are to be developed.
The authors of this paper suggest concentrating all forest and wildlife management practice on deer exclusion in regions with a high browsing pressure. This is especially important in protected areas focused on conserving the biodiversity of European hardwood forests, in which the absence of large carnivores limits the spontaneous succession of the forest ecosystem. A survey [87] on the near-natural hardwood floodplain forest of Cahnov-Soutok in the Czech Republic, which has been left to spontaneous development since the beginning of the 1930s, showed that (1) the most significant trend is a decreased representation of Quercus robur in all monitored indicators and conversely an expanding representation of Acer campestre, Carpinus betulus, and Tilia cordata; and (2) that the floodplain forest ecosystem demonstrates a high-level stability in the total volume of tree biomass with an essential change in the tree species composition, spatial structure, and average stem volume of individual trees. However, if the spontaneous gap dynamics and natural regeneration in the forest are limited by ungulate browsing, we cannot protect the biodiversity of hardwood floodplain forest, as pointed out in this paper, which seems to be a general problem in forest biodiversity conservation in the Czech Republic [88]. This problem is increased by the predictions of climate change impacts to European hardwood forests [89], because of an increase in the effect of factors limiting natural forest development and forest natural regeneration. The authors of this paper suggest that deer exclusion in European hardwood forests—especially in protected areas focusing on forest biodiversity conservation [90]—is necessary for putting the basic principles of sustainable forest management [91] into practice.
5. Conclusions
The European hardwood floodplain forests are characterized by a high avian diversity and density. In the unmanaged hardwood floodplain forest of the Litovelské Pomoraví PLA, however, an atypically low bird density was identified in five study plots with a very high degree of red deer and fallow deer browsing pressure on hardwood trees. The heavy browsing pressure has caused a strong reduction of the shrub and young tree layer in the understorey (30–210 cm above ground surface). In all five study plots, bird species from the bush nesters guild—usually dominant in hardwood floodplain forest ecosystems with a rich shrub and young tree layer and low browsing pressure—were found to be absent. As the decline in bird diversity is clearly a consequence of the browsing pressure, the intense ungulate browsing can be considered an important limiting factor to bird diversity (especially to bush nesters) in hardwood floodplain forests. Based on these results, we suggest that deer exclusion become an important component of forest management practice, in order to conserve hardwood floodplain forest biodiversity.
Author Contributions
I.M. and P.C. conceived and designed the sampling protocol; V.P. analyzed the data; I.M. wrote the paper.
Funding
This study was funded by the research grant “Significant Trees—Living Symbols of National and Cultural Identity” (no. DG18P02OVV027), within the NAKI II Program founded by the Czech Ministry of Culture.
Acknowledgments
Authors are grateful to Otto Cizek for providing data related to deer density in control plots.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Doyon, F.; Giroux, J.F.; Gagnon, D. Effects of different types of management on the diversity of birds and small mammals in a hardwood forest. In Landscape Ecology in Land use Planning Methods and Practice, Proceedings of the 4th Workshop of the Canadian-Society-for-Landscape-Ecology-and-Management, Laval, QC, Canada, 5–7 June 1995; Université Laval: Sanite Foy, QC, Canada; pp. 211–219.
- Laiolo, P. Effects of habitat structure, floral composition and diverzity on a forest bird community in north-western Italy. Folia Zool. 2002, 51, 121–128. [Google Scholar]
- Lindbladh, M.; Lindstrom, A.; Hedwall, P.O.; Felton, A. Avian diversity in Norway spruce production forests—How variation in structure and composition reveals pathways for improving habitat quality. For. Ecol. Manag. 2017, 397, 48–56. [Google Scholar] [CrossRef]
- Perry, R.W.; Jenkins, J.M.A.; Thill, R.E.; Thompson, F.R. Long-term effects of different forest regeneration methods on mature forest birds. For. Ecol. Manag. 2018, 408, 183–194. [Google Scholar] [CrossRef]
- Sallabanks, R.; Haufler, J.B.; Mehl, C.A. Influence of forest vegetation structure on avian community composition in west-central Idaho. Wildl. Soc. B 2006, 34, 1079–1093. [Google Scholar] [CrossRef]
- Pierce, A.R.; King, S.L. A comparison of avian communities and habitat characteristics in floodplain forests associated with valley plugs and unchannelized streams. River Res. Appl. 2011, 27, 1315–1324. [Google Scholar] [CrossRef]
- Zenzal, T.J.; Smith, R.J.; Ewert, D.N.; Diehl, R.H.; Buler, J.J. Fine-scale heterogeneity drives forest use by spring migrant landbirds across a broad, contiguous forest matrix. Condor 2018, 120, 166–184. [Google Scholar] [CrossRef]
- Knutson, M.G.; McColl, L.E.; Suarez, S.A. Breeding bird assemblages associated with stages of forest succession in large river floodplains. Nat. Areas J. 2005, 25, 55–70. [Google Scholar]
- Vacek, Z.; Vacek, S.; Bilek, L.; Kral, J.; Remes, J.; Bulusek, D.; Kralicek, I. Ungulate impact on natural regeneration in spruce-beech-fir stands in Cerny Dul nature reserve in the Orlicke Hory mountains, Case study from Central Sudetes. Forests 2014, 5, 2929–2946. [Google Scholar] [CrossRef]
- Chapron, G.; Kaczensky, P.; Linnell, J.D.C.; von Arx, M.; Huber, D.; Andren, H.; Lopez-Bao, J.V.; Adamec, M.; Alvares, F.; Anders, O.; et al. Recovery of large carnivores in Europe’s modern human-dominated landscapes. Science 2014, 346, 1517–1519. [Google Scholar] [CrossRef] [PubMed]
- De Jager, N.R.; Cogger, B.J.; Thomsen, M.A. Interactive effects of flooding and deer (Odocoileus virginianus) browsing on floodplain forest recruitment. For. Ecol. Manag. 2013, 303, 11–19. [Google Scholar] [CrossRef]
- Holt, C.A.; Fuller, R.J.; Dolman, P.M. Exclusion of deer affects responses of birds to woodland regeneration in winter and summer. IBIS 2014, 156, 116–131. [Google Scholar] [CrossRef]
- Newson, S.E.; Johnston, A.; Renwick, A.R.; Baillie, S.R.; Fuller, R.J. Modelling large-scale relationship between changes in woodland deer and bird populations. J. Anim. Ecol. 2012, 49, 278–286. [Google Scholar] [CrossRef]
- Hinsley, S.A.; Hill, R.A.; Fuller, R.J.; Bellamy, P.E.; Rothery, P. Bird species distributions across woodland canopy structure gradients. Community Ecol. 2009, 10, 99–110. [Google Scholar] [CrossRef]
- Charchuk, C.; Bayne, E.M. Avian community response to understory protection harvesting in the boreal forest of Alberta, Canada. For. Ecol. Manag. 2018, 407, 9–15. [Google Scholar] [CrossRef]
- Eichhorn, M.P.; Ryding, J.; Smith, M.J.; Gill, R.M.A.; Siriwardena, G.M.; Fuller, R.J. Effects of deer on woodland structure revealed through terrestrial laser scanning. J. Appl. Ecol. 2017. [Google Scholar] [CrossRef]
- Garabedian, J.E.; Moorman, C.E.; Peterson, M.N.; Kilgo, J.C. Use of LiDAR to define habitat thresholds for forest bird conservation. For. Ecol. Manag. 2017, 399, 24–36. [Google Scholar] [CrossRef]
- Fuller, R.J. Searching for biodiversity gains through woodfuel and forest management. J. Appl. Ecol. 2013, 50, 1295–1300. [Google Scholar] [CrossRef]
- Petty, S.J.; Avery, M.I. Forest Bird Communities. A Review of the Ecology and Management of Forest Bird Communities in Relation to Silvicultural Practices in the British Uplands; Forestry Commission Paper; Forestry Commission: Edinburgh, UK, 1990; Volume 26, pp. 1–41. [Google Scholar]
- Fuller, R.J.; Smith, K.W.; Hinsley, S.A. Temperate western European woodland as a dynamic environments for birds: A resource-based review. In Birds and Habitat: Relationship in Changing Landscapes; Fuller, R.J., Ed.; Cambridge University Press: Cambridge, UK, 2012; pp. 352–380. [Google Scholar]
- Machar, I. Conservation and Management of Floodplain Forests in the Protected Landscape Area Litovelske Pomoravi (Czech Republic) Introduction. In Conservation and Management of Floodplain Forests in the Protected Landscape Area Litovelske Pomoravi (Czech Republic); Accession Number: WOS:000331015800001; Machar, I., Ed.; Palacky University: Olomouc, Czech Republic, 2009; pp. 7–108. ISBN 978-80-244-2355-5. [Google Scholar]
- Kilianova, H.; Pechanec, V.; Svobodova, J.; Machar, I. Analysis of the evolution of the floodplain forests in the aluvium of the Morava river. In Proceedings of the 12th International Multidisciplinary Scientific Geoconference (SGEM 2012), Albena, Bulgaria, 17–23 June 2012; SGEM: Albena, Bulgaria48535300001, 2012. Accession Number: WOS:000348535300001. Volume IV, pp. 1–8. [Google Scholar]
- Simon, J.; Machar, I.; Bucek, A. Linking the historical research with the growth simulation model of hardwood floodplain forests. Pol. J. Ecol. 2014, 62, 273–288. [Google Scholar] [CrossRef]
- Kusbach, A.; Friedl, M.; Zouhar, V.; Mikita, T.; Šebesta, J. Assessing Forest Classification in a Landscape-Level Framework: An Example from Central European Forests. Forests 2017, 8, 461. [Google Scholar] [CrossRef]
- Miko, L. Nature and landscape protection in the European context. In Ochrana prirody a krajiny v Ceske Republice, Vols I and II; Accession Number: WOS:000334387900004; Machar, I., Drobilova, L., Eds.; Palacky University: Olomouc, Czech Republic, 2012; pp. 43–49. ISBN 978-80-244-3041-6. [Google Scholar]
- Hanzelka, J.; Reif, J. Responses to the black locust (Robinia pseudoacacia) invasion differ between habitat specialist and generalist in central European forest birds. J. Ornithol. 2015, 156, 1015–1024. [Google Scholar] [CrossRef]
- Kroftova, M.; Reif, J. Management implications of bird responses to variation in non-native/native tree ratios within central European forest stands. For. Ecol. Manag. 2017, 391, 330–337. [Google Scholar] [CrossRef]
- Bibby, C.J.; Burges, N.D.; Hill, D.A.; Mustoe, S. Bird Census Techniques; Academic Press: London, UK, 2007; pp. 42–64. ISBN 978-0-12-095831-3. [Google Scholar]
- Poprach, K.; Vrbkova, J.; Machar, I. Detectability as an important factor influencing the knowledge of bird diversity in a floodplain forest ecosystem. J. For. Sci. 2015, 61, 89–97. [Google Scholar] [CrossRef]
- Wiens, J.A. The Ecology of Bird Communities, Vol. 1, Foundation and Patterns; Cambridge University Press: Cambridge, UK, 1989; pp. 1–539. ISBN 0-521-26030. [Google Scholar]
- Machar, I. Changes in ecological stability and biodiversity in a floodplain landscape. In Applying Landscape Ecology in Conservation and Management of the Floodplain Forest (Czech Republic); Accession Number: WOS:000325436900004; Machar, I., Ed.; Palacky University: Olomouc, Czech Republic, 2012; pp. 73–87. ISBN 978-80-244-2997-7. [Google Scholar]
- Stastny, K.; Hudec, K. Fauna of the Czech Republic. Birds 2 and 3, 2nd ed.; Academia: Prague, Czech Republic, 2011; pp. 1–1178. ISBN 80-200-1113-7. [Google Scholar]
- Aulak, W. Small mammal communities of the Białowieża National Park. Acta Theriol. 1970, 15, 465–515. [Google Scholar] [CrossRef] [Green Version]
- Cermak, P.; Horsak, P.; Spirik, M.; Mrkva, R. Relationships between browsing damage and woody species dominance. J. For. Sci. (Prague) 2009, 55, 23–31. [Google Scholar] [Green Version]
- Cermak, P.; Beranova, P.; Oralkova, J.; Horsak, P.; Plsek, J. Relationships between browsing damage and the species dominance by the highly food-attractive and less food-attractive trees. Acta Univ. Agric. Mendel. Brun. 2011, 59, 29–36. [Google Scholar] [CrossRef] [Green Version]
- Cermak, P.; Mrkva, R. Browsing damage to broadleaves in some national nature reserves (Czech Republic) in 2000–2001. Ekológia (Bratislava) 2003, 22, 132–141. [Google Scholar]
- StatSoft, s.r.o. Statistica [software, CD-ROM]. Ver. 12. Praha, 2013. [Google Scholar]
- Jaccard, P. Étude comparative de la distribution florale dans une portion des Alpes et des Jura. Bull. Soc. Vaud. Sci. Nat. 1901, 37, 547–579. [Google Scholar]
- James, F.C.; Rathbun, S. Rarefaction, relative abundance and diversity of avian communities. Auk 1981, 98, 785–800. [Google Scholar]
- Author Colwell, R.K. EstimateS: Statistical Estimation of Species Richness and Shared Species from Samples. Version 8.0.0. 2005. Available online: http://viceroy.eeb.uconn.edu/estimates (accessed on 12 January 2018).
- Bures, S.; Maton, K. Ptaci slozka segmentu skupiny typu geobiocenu Ulmi-fraxineta populi v navrhovane CHKO Pomoravi [Birds of hardwood floodplain forest in proclaimed PLA Pomoravi—In Czech]. Sylvia 1984, 23–24, 37–46. [Google Scholar]
- Bures, S. Analyza ptaci slozky navrhovane SPR Sargoun [Birds of harwood floodplain forest in Sargoun locality—In Czech]. Unpublished Work. 1996. [Google Scholar]
- Horak, Z. Ptactvo okolí Starého Labe u Cihelny u Pardubic [Birds of Stare Labe u Cihelny locality—In Czech]. Panurus 1988, 9, 53–61. [Google Scholar]
- Chytil, J. Srovnání Produkce Savců a ptáků v Lužním lese [Comparison of the Production of Birds and Mammals in Floodplain Forest—In Czech]. Ph.D. Thesis, University JE Purkyne, Brno, Czech Republic, 1991. [Google Scholar]
- Kubecka, D. Avifauna lužního lesa na lokalitě Horní Záseky v CHKO Litovelské Pomoraví [Birds of floodplain forest in Horni Zaseky locality—In Czech]. Ph.D. Thesis, Palacky University, Olomouc, Czech Republic, 2003. [Google Scholar]
- Lemberk, V. Srovnání ornitocenóz čtyř lužních lesů ve východních Čechách [Comparison of avian communities in floodplain forests—In Czech]. Panurus 2001, 11, 69–79. [Google Scholar]
- Machar, I. The effect of floodplain forest fragmentation on bird community. J. For. Sci. 2012, 58, 213–224. [Google Scholar] [CrossRef]
- Machar, I. The impact of floodplain forest habitat conservation on the structure of bird breeding communities. Ekológia (Bratislava) 2011, 30, 36–50. [Google Scholar] [CrossRef]
- Pavelka, J. Hnízdní ornitocenóza v lužním lese u řeky Odry [Birds in floodplain forest along Odra river—In Czech]. Zprávy MOS (Přerov) 1987, 46, 115–118. [Google Scholar]
- Polasek, V. Výzkum Ornitocenózy lužního lesa v Litovelském Pomoraví [Birds of Floodplain Forest in Litovelske Pomoravi—In Czech]. Ph.D. Thesis, Palacky University, Olomouc, Czech Republic, 2001. [Google Scholar]
- Pykal, J. Ornitocenosy různých typů přirozených lesních společenstev v pahorkatině jihozápadních Čech [Avian community in hardwood forests in Czechia—In Czech]. Panurus 1991, 3, 67–76. [Google Scholar]
- Ruzicka, I. Ornitologický výzkum lokality Chrbovský les u Záříčí [Birds of Floodplain Forest Near Zarici—In Czech]. Ph.D. Thesis, Palacky University, Olomouc, Czech Republic, 2005. [Google Scholar]
- Storch, D. Densities and territory of birds in two different lowland communities in eastern Bohemia. Folia Zool. 1998, 47, 181–188. [Google Scholar]
- Toman, A. Avifauna SPR Zastudanci [Avian Community in Floodplain Forest Zastudanci Locality]. Ph.D. Thesis, Palacky University, Olomouc, Czech Republic, 2004. [Google Scholar]
- Nascher, F.A. Zur Waldbaulichen Bedeutung des Rothirschverbisses in der Waldgesellschaft des subalpinen Fichtenwalds in der Umgebung des Schweizerischen Nationalparks. Ph.D. Thesis, ETH Zürich, Rämistr, Zürich Switzerland, 1979. Diss. Nr. 6373. [Google Scholar]
- Heuze, P.; Schnitzler, A.; Klein, F. Is browsing the major factor of silver fir decline in the Vosges Mountains of France? For. Ecol. Manag. 2005, 217, 219–228. [Google Scholar] [CrossRef]
- Barancekova, M.; Krojerova-Prokesova, J.; Homolka, M. Impact of deer browsing on natural and artificial regeneration in floodplain forest. Folia Zool. 2007, 56, 354–364. [Google Scholar]
- Kubicek, F.; Simonovic, V.; Kollar, J.; Kanka, R. Herb layer biomass of the Morava river floodplain forests. Ekológia (Bratislava) 2008, 27, 23–30. [Google Scholar]
- Cogger, B.J.; De Jager, N.R.; Thomsen, M.; Adams, C.R. Winter Browse Selection by White-Tailed Deer and Implications for Bottomland Forest Restoration in the Upper Mississippi River Valley, USA. Nat. Areas J. 2014, 34, 144–153. [Google Scholar] [CrossRef]
- Cermak, P.; Mrkva, R.; Horsak, P.; Spirik, M.; Beranova, P.; Oralkova, J.; Plsek, J.; Kadlec, M.; Zarybnicky, O.; Svatos, M. Impact of Ungulate Browsing on Forest Dynamics, 1st ed.; Lesnická Práce, Folia Forestalia Bohemica: Kostelec nad Černými lesy, Czech Republic, 2011; pp. 61–66. ISBN 978-80-87154-94-6. [Google Scholar]
- Frerker, K.; Sonnier, G.; Waller, D.M. Browsing rates and ratios provide reliable indices of ungulate impacts on forest plant communities. For. Ecol. Manag. 2013, 291, 55–64. [Google Scholar] [CrossRef]
- Ammer, C. Impact of ungulates on structure and dynamics of natural regeneration of mixed mountain forests in the Bavarian Alps. For. Ecol. Manag. 1996, 88, 45–53. [Google Scholar] [CrossRef]
- Stergar, M. Objedenost Mladja Drevesnih Vrstv Odvisnosti od Zgradbe Sestoja. Ph.D. Thesis, Biotehniška fakulteta, Univerza v Ljubljani, Ljubljana, Slovenia, 2005; p. 70. [Google Scholar]
- Hubalek, Z. Seasonal variation of forest habitat preferences by birds in a lowland riverine ecosystem. Folia Zool. 2001, 50, 281–289. [Google Scholar]
- Schlaghamersky, J.; Hudec, K. The fauna of temperate European floodplain forests. In Floodplain Forests of the Temperate Zone of Europe; Klimo, E., Hager, H., Matic, S., Anic, I., Kulhavy, J., Eds.; Lesnicka Prace: Kostelec, Czech Republic, 2008; pp. 160–230. ISBN 978-80-87154-16-8. [Google Scholar]
- Walther, B.A.; Martin, J.L. Species richness estimation of bird communities: How to control for sampling effort? IBIS 2001, 143, 413–419. [Google Scholar] [CrossRef]
- Kolecek, J.; Paclik, M.; Weidinger, K.; Reif, J. Abundance and species richness of birds in two lowland riverine forests in Central Moravia—Possibilities for analyses of point-count data. Sylvia 2010, 46, 71–85. [Google Scholar]
- Freemark, K.E.; Collins, B. Landscape ecology of birds breeding in temperate forest fragments. In Ecology ans Conservation of Neotropical Migrant Landbird; Hagan, J.M., Johnston, D.W., Eds.; Smithsomian Institution Press: Washington, DC, USA, 1992; pp. 443–454. [Google Scholar]
- Pechanec, V.; Brus, J.; Kilianova, H.; Machar, I. Decision support tool for the evaluation of landscapes. Ecol. Inform. 2015, 30, 305–308. [Google Scholar] [CrossRef]
- Kornan, M. Comparison of bird assemblage structure between forest ecotone and interior of an alder swamp. Sylvia 2009, 45, 151–176. [Google Scholar]
- Truax, B.; Gagnon, D.; Lambert, F.; Fortier, J. Multiple-Use Zoning Model for Private Forest Owners in Agricultural Landscapes: A Case Study. Forests 2015, 6, 3614–3664. [Google Scholar] [CrossRef] [Green Version]
- Anic, I.; Mestrovic, S.; Matic, S. Important events in the history of forestry in Croatia. Sumar. List 2012, 136, 169–177. [Google Scholar]
- Kilianova, H.; Pechanec, V.; Brus, J.; Kirchner, K.; Machar, I. Analysis of the development of land use in the Morava River floodplain, with special emphasis on the landscape matrix. Morav. Geogr. Rep. 2017, 25. [Google Scholar] [CrossRef]
- Leso, P. Breeding bird communities of two succession stages of young oak forests. Sylvia 2003, 39, 67–78. [Google Scholar]
- Fulller, R.J. Bird Life of Woodland and Forest, 1st ed.; Cambridge University Press: Cambridge, UK, 2003; pp. 126–127. [Google Scholar]
- Glowacinski, Z. Succession of bird communities in the Niepolomice Forest (Southern Poland). Ekol. Polska 1975, 23, 231–263. [Google Scholar]
- Kropil, R. Struktura a Produkcia Ornitocenoz Vybranych Prirodnych Lesov Slovenska [Structure and Production of Bird Communities in Natural Forests in Slovakia]. Ph.D. Thesis, Technical University, Zvolen, Zvolen, Slovakia, 1993. [Google Scholar]
- Waliczky, Z. Bird community changes in different-aged oak forest stands in the Buda-hills. Ornis Hung. 1991, 1, 1–9. [Google Scholar]
- Wesolovski, T.; Rowinski, P.; Mitrus, C.; Czeszczewik, D. Breeding bird community of a primeval temperate forest (Bialowieza National Park, Poland) at the beginning of the 21st century. Acta Ornithol. 2006, 41, 55–70. [Google Scholar] [CrossRef]
- Tucker, G.M.; Evans, M.I. Habitats for Birds in Europe; Birdlife International: Cambridge, UK, 1997; pp. 1–464. ISBN 978-0-94688-8320. [Google Scholar]
- Stastny, K.; Bejcek, V.; Hudec, K. Atlas of Breeding Bird Distribution in the Czech Republic; Academia: Prague, Czech Republic, 2006; pp. 1–463. ISBN 80-86858-19-7. [Google Scholar]
- Reif, J.; Jiguet, F.; Šťastný, K. Habitat specialization of birds in the Czech Republic: Comparison of objective measures with expert opinion. Bird Study 2010, 57, 197–212. [Google Scholar] [CrossRef]
- Reif, J.; Storch, D.; Šímová, I. The effect of scale-dependent habitat gradients on the structure of bird assemblages in the Czech Republic. Acta Ornithol. 2008, 43, 197–206. [Google Scholar] [CrossRef]
- Machar, I. Attempt to summarize the problems: Is a sustainable management of floodplain forest geobiocenoses possible? In Biodiversity and Target Management of Floodplain Forests in the Morava River Basin (Czech Republic); Accession Number: WOS:000328003200016; Machar, I., Ed.; Palacky University: Olomouc, Czech Republic, 2010; pp. 189–226. ISBN 978-80-244-2530-6. [Google Scholar]
- Kovarik, P.; Kutal, M.; Machar, I. Sheep and wolves: Is the occurrence of large predators a limiting factor for sheep grazing in the Czech Carpathians? J. Nat. Conserv. 2014, 22, 5. [Google Scholar] [CrossRef]
- Machar, I.; Harmacek, J.; Vrublova, K.; Filippovova, J.; Brus, J. Biocontrol of common vole populations by avian predators versus rodenticide application. Pol. J. Ecol. 2017, 65, 434–444. [Google Scholar] [CrossRef]
- Janik, D.; Adam, D.; Vrska, T.; Hort, L.; Unar, P.; Kral, K.; Samonil, P.; Horal, D. Tree layer dynamics of the Cahnov-Soutok near-natural floodplain forest after 33 years (1973–2006). Eur. J. For. Res. 2008, 127, 337–345. [Google Scholar] [CrossRef]
- Machar, I. Protection of nature and landscapes in the Czech Republic Selected current issues and possibilities of their solution. In Ochrana Prirody a Krajiny v Ceske Republice, Vols I and II; Accession Number: WOS:000334387900001; Machar, I., Drobilova, L., Eds.; Palacky University: Olomouc, Czech Republic, 2012; p. 9-+. ISBN 978-80-244-3041-6. [Google Scholar]
- Machar, I.; Vlckova, V.; Bucek, A.; Vozenilek, V.; Salek, L.; Jerabkova, L. Modelling of Climate Conditions in Forest Vegetation Zones as a Support Tool for Forest Management Strategy in European Beech Dominated Forests. Forests 2017, 8, 82. [Google Scholar] [CrossRef]
- Machar, I.; Simon, J.; Rejsek, K.; Pechanec, V.; Brus, J.; Kilianova, H. Assessment of Forest Management in Protected Areas Based on Multidisciplinary Research. Forests 2016, 7, 285. [Google Scholar] [CrossRef]
- Spathelf, P. Sustainable Forest Management as a Model for Sustainbale Development: Conclusions Toward a Concrete Vision. Sustain. For. Manag. Chang. World Manag. For. Ecosyst. 2009, 19, 237–240. [Google Scholar] [CrossRef]
Figure 1.
Study area and location of study plots 1–5.

Figure 2.
Ungulate density and browsing in study plots 1–5.

Figure 3.
Standardized rarefaction curves for species richness in study plots (S = number of bird species, N = number of nesting pairs, 1–5 = rarefaction curves for study plots no. 1–5).
Figure 3.
Standardized rarefaction curves for species richness in study plots (S = number of bird species, N = number of nesting pairs, 1–5 = rarefaction curves for study plots no. 1–5).

Figure 4.
Structure of bird nesting guilds in study plots. (GN—ground nesters, CN—canopy nesters, HN—hole nesters, BN—bush nesters).
Figure 4.
Structure of bird nesting guilds in study plots. (GN—ground nesters, CN—canopy nesters, HN—hole nesters, BN—bush nesters).

Figure 5.
Comparison of guild structures in bird communities in the hardwood floodplain forests in the Czech Republic. (GN—ground nesters, CN—canopy nesters, HN—hole nesters, BN—bush nesters). Localities: 1 = average data from LPPLA, 2 = [41], 3 = [42], 4 = [44], 5 = [45], 6 = [46] Bosin, 7 = [46] Dubno, 8 = [46] Choltice, 9 = [46] Zbytka, 10 = [48] Sargoun, 11 = [48] Vrapac, 12 = [49], 13 = [49], 14 = [54]).
Figure 5.
Comparison of guild structures in bird communities in the hardwood floodplain forests in the Czech Republic. (GN—ground nesters, CN—canopy nesters, HN—hole nesters, BN—bush nesters). Localities: 1 = average data from LPPLA, 2 = [41], 3 = [42], 4 = [44], 5 = [45], 6 = [46] Bosin, 7 = [46] Dubno, 8 = [46] Choltice, 9 = [46] Zbytka, 10 = [48] Sargoun, 11 = [48] Vrapac, 12 = [49], 13 = [49], 14 = [54]).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Correlation matrix for woody plants height, density, and intensity of browsing damage (% of damaged woody plants).
Table 1.
Correlation matrix for woody plants height, density, and intensity of browsing damage (% of damaged woody plants).
| Height | Density | Browsing % | |
|---|---|---|---|
| Height | - | −0.5653 1 | 0.4271 |
| Density | - | −0.5318 1 | |
| Browsing % | - |
1 Significant values are shown in bold; significant level: p < 0.05.
Table 2.
Densities and dominance of bird nesting guilds in study plots.
| Bird Species | Nesting Guilds 1 | Study Plot | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | |||||||
| DE 2 | DO 3 | DE | DO | DE | DO | DE | DO | DE | DO | ||
| Anthus trivialis | GN | 0.5 | 0.8 | 0.4 | 0.6 | 0.7 | 1.2 | 0.3 | 0.5 | 0.5 | 0.8 |
| Certhia brachydactyla | HN | 2.1 | 3.4 | 2.7 | 3.9 | 0.4 | 0.6 | 2.9 | 5.1 | 0.0 | 0.0 |
| Columba palumbus | CN | 1.6 | 2.6 | 2.2 | 3.2 | 2.6 | 4.3 | 0.6 | 1.1 | 2.0 | 3.2 |
| Cyanistes caeruleus | HN | 3.2 | 4.9 | 0.9 | 1.3 | 2.7 | 4.5 | 3.9 | 6.9 | 3.0 | 4.7 |
| Dendrocopos major | HN | 2.8 | 4.3 | 3.9 | 5.6 | 1.5 | 2.5 | 2.8 | 4.9 | 3.1 | 4.9 |
| Dendrocopos medius | HN | 0.0 | 0.0 | 0.7 | 1.0 | 0.0 | 0.0 | 0.5 | 0.9 | 0.6 | 0.9 |
| Erithacus rubecula | GN | 2.4 | 3.8 | 2.7 | 3.9 | 2.1 | 3.5 | 1.3 | 2.3 | 2.9 | 4.6 |
| Ficedula albicollis | HN | 7.0 | 10.8 | 4.3 | 6.2 | 7.6 | 12.7 | 7.9 | 13.9 | 3.8 | 6.0 |
| Fringilla coelebs | CN | 4.3 | 6.6 | 3.2 | 4.6 | 3.9 | 6.5 | 1.9 | 3.4 | 2.7 | 4.2 |
| Garrulus glandarius | CN | 0.6 | 0.9 | 0.9 | 1.3 | 0.5 | 0.8 | 1.2 | 2.0 | 0.7 | 1.1 |
| Muscicapa striata | HN | 0.6 | 0.9 | 1.1 | 1.7 | 1.3 | 2.3 | 0.9 | 1.6 | 0.5 | 0.8 |
| Oriolus oriolus | CN | 1.8 | 2.8 | 1.1 | 1.7 | 0.7 | 1.1 | 1.1 | 1.9 | 1.6 | 2.5 |
| Parus major | HN | 10.3 | 16.1 | 7.1 | 10.2 | 8.2 | 13.7 | 8.9 | 15.7 | 9.6 | 15.1 |
| Phylloscopus collybita | GN | 2.2 | 3.4 | 2.9 | 4.2 | 2.2 | 3.7 | 3.7 | 6.5 | 1.8 | 2.8 |
| Picus viridis | HN | 0.3 | 0.4 | 0.9 | 1.3 | 0.0 | 0.0 | 1.0 | 1.8 | 0.0 | 0.0 |
| Poecile palustris | HN | 0.9 | 1.4 | 0.2 | 0.3 | 0.7 | 1.2 | 0.8 | 1.4 | 0.6 | 0.9 |
| Sitta europaea | HN | 5.3 | 8.2 | 6.7 | 9.7 | 5.1 | 8.5 | 6.9 | 12.2 | 5.0 | 7.9 |
| Steptopelia turtur | CN | 0.8 | 1.2 | 1.5 | 2.2 | 0.6 | 1.0 | 0.6 | 1.1 | 0.9 | 1.4 |
| Sturnus vulgaris | HN | 17.2 | 26.6 | 19.0 | 27.5 | 15.4 | 25.7 | 9.1 | 16.1 | 18.3 | 28.8 |
| Turdus philomelos | CN | 0.6 | 0.9 | 0.5 | 0.7 | 0.6 | 0.6 | 0.4 | 0.7 | 0.7 | 1.1 |
| Turdus pilaris | CN | 0.0 | 0.0 | 6.2 | 8.9 | 3.1 | 5.2 | 0.0 | 0.0 | 5.3 | 8.3 |
| Total of DE and DO | 64.5 | 100 | 69.1 | 100 | 59.9 | 100 | 56.7 | 100 | 63.6 | 100 | |
| Total of bird species | 19 | 21 | 19 | 20 | 19 | ||||||
1 Nesting guilds: GN—ground nesters, CN—canopy nesters, HN—hole nesters, BN—bush nesters; 2 DE = density [amount of nesting pairs/10 ha]; 3 DO = dominance [%].
Table 3.
Comparison of bird and deer density between LPPLA and other localities in hardwood floodplain forests in the Czech Republic.
Table 3.
Comparison of bird and deer density between LPPLA and other localities in hardwood floodplain forests in the Czech Republic.
| Source 1 | Size of Study Plot (ha) | Deer Density (Individuals/1000 ha) | Species Richness of Nesting Birds in Study Plot | Density of Birds (Nesting Pairs/10 ha) | |
|---|---|---|---|---|---|
| Capreolus capreolus | Dama dama | ||||
| Results of this study in LPPLA (study plot 1) | 1 | 63 | 11 | 19 | 64.5 |
| Results of this study in LPPLA (study plot 2) | 1 | 63 | 11 | 21 | 69.1 |
| Results of this study in LPPLA (study plot 3) | 1 | 98 | 63 | 19 | 59.9 |
| Results of this study in LPPLA (study plot 4) | 1 | 98 | 63 | 20 | 56.7 |
| Results of this study in LPPLA (study plot 5) | 1 | 98 | 63 | 19 | 63.6 |
| [41] | 15 | 21 | 5 | 39 | 161 |
| [42] | 5 | 21 | 5 | 48 | 79 |
| [43] | 9 | 39 | - | 44 | 39 |
| [44] | 10 | 18 | 15 | 37 | 177 |
| [45] | 12 | 44 | - | 28 | 93 |
| [46] locality Bosin | 32 | 32 | 3 | 44 | 103 |
| [46] locality Dubno | 51 | 29 | 3 | 40 | 101 |
| [46] locality Choltice | 52 | 37 | 3 | 42 | 109 |
| [46] locality Zbytka | 43 | 40 | 3 | 41 | 80.5 |
| [47] locality Panensky les I | 9 | 35 | - | 31 | 139 |
| [47] locality Panensky les II | 9 | 35 | - | 36 | 118 |
| [48] locality Sargoun | 12 | 21 | 5 | 33 | 135 |
| [48] locality Vrapac | 13 | 35 | 11 | 31 | 126 |
| [49] | 10 | 20 | 3 | 25 | 113 |
| [50] | 10 | 50 | 11 | 23 | 92 |
| [51] | 22 | 41 | 5 | 36 | 112 |
| [52] | 10 | 36 | - | 23 | 102 |
| [53] | 12 | ? | - | 30 | 270 |
| [54] | 5 | 33 | 3 | 30 | 174 |
1 Sources in brackets: see References. If there is more than one study site in cited literature, this is indicated.
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Machar, I.; Cermak, P.; Pechanec, V. Ungulate Browsing Limits Bird Diversity of the Central European Hardwood Floodplain Forests. Forests 2018, 9, 373. https://doi.org/10.3390/f9070373
AMA Style
Machar I, Cermak P, Pechanec V. Ungulate Browsing Limits Bird Diversity of the Central European Hardwood Floodplain Forests. Forests. 2018; 9(7):373. https://doi.org/10.3390/f9070373
Chicago/Turabian StyleMachar, Ivo, Petr Cermak, and Vilem Pechanec. 2018. "Ungulate Browsing Limits Bird Diversity of the Central European Hardwood Floodplain Forests" Forests 9, no. 7: 373. https://doi.org/10.3390/f9070373
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.