Native and Invasive Woody Species Differentially Respond to Forest Edges and Forest Successional Age


1
Agronomy Department, University of Florida, 1676 McCarty Drive, Gainesville, FL 32611, USA
2
Department of Biology, Indiana University, 1001 East Third Street, Bloomington, IN 47405, USA
*
Author to whom correspondence should be addressed.
Forests 2018, 9(7), 381; https://doi.org/10.3390/f9070381
Submission received: 15 May 2018
/
Revised: 20 June 2018
/
Accepted: 21 June 2018
/
Published: 26 June 2018
(This article belongs to the Special Issue Ecology and Management of Invasive Species in Forest Ecosystems)
Abstract
:Forest fragmentation can promote non-native plant invasions by increasing invasive plant seed dispersal and resource availability along edges. These effects can vary based on forest age and may be influenced by differences in traits of native and invasive plant species. To determine how native versus invasive plant species respond to forest edges and forest successional age, we experimentally evaluated germination, survival, and growth of three native and three invasive woody plant species in eastern USA forests. Across all species, increasing distance from the edge resulted in more germination and less seedling growth, but had no effect on seedling survival. Generally, seedling growth was greater in younger forests and invasive species outperformed native species; however, there were significant species-specific differences in performance. For example, among native species, spicebush had poor growth performance but high survival, while redbud had low germination but high growth performance and survival. By contrast, the invasive privet and autumn olive produced more biomass with high relative growth rates, and autumn olive had exceptionally high germination but the lowest survival. Overall, our results suggest that while there are some general characteristics of invasive species, species-specific traits may better inform management strategies and improve predictions about biological invasions along forest edges.
Keywords:
edge effects; seedling growth; survival; invasion; Elaeagnus; Lonicera; Ligustrum; Cornus; Cercis; Lindera1. Introduction
The susceptibility of forests to plant invasions is determined by both characteristics of invaders and environmental conditions of forest stands [1]. Natural and anthropogenic disturbances have increased forest fragmentation such that, globally, approximately 70% of forests are within 1 km of a forest edge [2]. Forest fragmentation and proximity to forest edges have frequently been associated with non-native plant invasions (e.g., [3,4,5]), predominantly due to a combination of greater invasive plant propagule dispersal and higher resource availability (e.g., light) along edges compared to forest interiors [6,7,8]. Anthropogenic disturbances that fragment forests and create edges may increase propagule pressure through direct introductions on contaminated forestry or road building equipment. In addition, fragmenting forests can create travel corridors where animals and human activity disperse propagules along edges [9,10,11].
Forest edges can have unique biotic and abiotic conditions compared to forest interiors [12,13], including differences in productivity, nutrient cycling, and evapotranspiration due to modification of tree density and canopy cover along edges [14]. These effects in turn influence invasive and native plant species recruitment, growth, reproduction, and exposure to herbivory, often resulting in distinct plant communities at forest edges compared to forest interiors [14,15,16]. The magnitude of edge effects can vary with distance from edge, age, and site-specific characteristics such as topography and soil type [13,14,17]. Younger forests and newer, more recent, edges may be more prone to invasion due in part to a greater light availability and greater exposure to invader propagules compared to mature forests [18,19]. Mature forests also may exhibit more biotic resistance due to greater abundance of resident native species and generalist herbivores that suppress invader establishment [20].
In addition to edge effects, plant community composition may be influenced by plant traits (e.g., phenology and fecundity) that are related to species origin (native vs. non-native invasive). Traits of invasive plant species, such as extended leaf phenology, rapid growth, high fecundity, and persistence in marginal habitats, can enable these species to capitalize on higher resource availability such as canopy gaps and soil disturbance [21,22,23,24]. Additionally, invasive species may benefit from enemy-release in their new habitat [25,26]. In contrast, many native species have later spring and earlier fall phenology, are slower growing, and experience more damage from natural enemies, and therefore are slower to respond to greater resource availability across forest successional ages and proximity to forest edges.
Traits that might explain dominance of invasive over native species, particularly at forest edges, often are inferred from observations where the invader already is dominant (e.g., [5,27,28]). Yet, the high abundance of invasive species observed near forest edges could be simply due to high propagule pressure at edges compared to forest interiors, not because of particular invader traits [27]. Moreover, it is unclear whether the invader characteristics identified in invader-dominated communities provide advantages over native species during the early stage of invasion (i.e., seed germination and seedling growth) or if such differences are only realized later in the succession process. Experimental tests of seed germination and seedling survival and growth are needed to disentangle propagule pressure from traits that may drive differences in the establishment and performance of invasive versus native species at forest edges compared to forest interiors.
Here, we experimentally introduced native and non-native invasive (hereafter “invasive”) woody plant species to deciduous forests of the eastern USA to compare germination, survival, and growth among the species across distance from forest edge and forest successional age. Three native woody species (Cornus florida, Cercis canadensis, Lindera benzoin) and three invasive woody species (Elaeagnus umbellata, Ligustrum obtusifolium, and Lonicera maackii) common to these forests were used in this experiment. These invasive species have been identified as some of the most problematic invaders in the region [19,29]. Previously, Flory and Clay [30] showed that as a group these invasive species generally outperformed the native species group in germination, survival, and growth, but they did not evaluate how the individual species contributed to differences between groups. Here, we examined seed germination and seedling survival and performance at the species level to identify particular species that contributed to the broader differences between these invasive and native species groups. Then, we focused on particular traits, such as shade tolerance, phenology, and higher growth rates, that may enable species to take advantage of greater resource availability associated with forest edge effects and forest successional age. Examining how edge effects influence both native and invasive seedling establishment and performance can help elucidate how understory plant community structure is realized.
2. Materials and Methods
2.1. Species Description
We selected three invasive woody species currently impacting eastern deciduous forests and three native woody species that occupy the forest understory. Elaeagnus umbellata (autumn olive), Ligustrum obtusifolium (border privet), and Lonicera maackii (Amur honeysuckle) share many traits of other invasive species such as high fecundity, germination rates, and relative growth rates, moderate to high shade tolerance, and the ability to grow in low nutrition and variable soil moisture conditions [31,32,33]. They have also benefited from an apparent release from natural enemies and are potentially allelopathic [34,35,36,37]. All three invasive shrubs occupy a temporal resource niche in deciduous forests by leafing out in early spring before the canopy trees and retaining leaves late in the growing season [22]. For example, privet is described as “semi-evergreen” [38] and honeysuckle often retains green, frost tolerant leaves through November or even December [39].
The native species, Cercis canadensis (redbud), Cornus florida (flowering dogwood), and Lindera benzoin (spicebush), also share common characteristics including high shade tolerance, lower relative growth rates, and susceptibility to native and introduced natural enemies [40,41,42]. For example, dogwoods infected with the introduced fungus Discula destructiva suffer from anthracnose, which has up to a 90% mortality rate [40,43]. Numerous damaging enemies (wood borers, various Lepidoptera, spittlebugs, and pathogens) have been observed on redbud [44]. Unlike the invasive species in this study, these native species flower before leaf-out, often in phase with the budburst of the overstory trees [45]. The native species allocate to early reproduction at a time when a more open canopy promotes pollinator visitation [46].
2.2. Site Selection and Habitat Characterization
In June of 2004, we located 12 research sites bordering two-lane public roads that were mixtures of Quercus-Carya (oak-hickory) and Fagus-Acer (beech-maple) dominated forests in southern Indiana, USA. Site selection was stratified to have six mature forest sites (trees >60 years old) and six young forest sites (trees <40 years old). At each of the 12 sites we selected a random point along the road and ran a 60 m transect into the forest perpendicular to the forest edge. Along each transect, light availability, soil moisture, and litter were measured at 0, 5, 10, 20, 40, and 60 m from the edge. These distances were selected because previous observations showed high abundances of these invasive species within 10 m of forest edges and extending at least 30 m into the forest [19]. Light availability was calculated from measurements of photosynthetically active radiation (PAR) made at each location relative to PAR measurements taken in open sun (AccuPAR Linear PAR/LAI ceptometer, Decagon Devices, Inc., Pullman, WA, USA). A 125 cm3 mineral soil sample was collected at each location to calculate the gravimetric water content (soil moisture). Litter was collected from within a 25 cm × 25 cm area at each location, dried to a constant mass, and weighed. We characterized forest structure at each point by quantifying tree basal area using a wedge prism. See Flory and Clay [30] for additional details on the study area and site selection.
2.3. Experimental Design
To assess species-level differences in survival, growth, and incidence of herbivory related to edge effects, we transplanted four seedlings of each species at each of the distances from the edge at each site, i.e., 24 plants of each species for each combination of distance and forest age. Seedlings were reared under common conditions in an Indiana University shaded greenhouse and were between 10 and 15 cm height at the time of planting in June 2004. Seedlings that died within two weeks of planting were replaced. In September 2004, 2005, and 2006 we scored each seedling for survivorship and evaluated incidence of herbivory. The height, basal diameter, and above-ground biomass of all seedlings surviving to September 2006 were measured. Above-ground biomass was dried to a constant mass at 60 °C and weighed (±0.01 g). All seedlings were removed before they reached a reproductive age. To assess germination related to edge effects, ten seeds of each species were enclosed in 10 cm × 10 cm seed bags (0.09 mm2 mesh size) and placed under leaf litter (0.5–1.9 kg m2) at each distance from the forest edge. Seeds were set out in October 2004 and collected in late June 2005 to score germination.
We calculated three growth metrics, survival, and germination to assess species-level responses to edge effects in the two forest successional ages. To characterize seedling growth, we calculated averages of biomass, basal diameter, and relative growth rate of height for each species at each distance from edge at each site. Relative growth rate was calculated as:
where height 1 and height 2 are plant heights at the time of transplanting in 2004 (time 1) and at the end of the study in 2006 (time 2), respectively.
Relative growth rate = [ln(height 2) − ln(height 1)]/(time 2 − time 1),
Survival was calculated as the proportion of seedlings alive in 2006 out of the initial number of seedlings planted. Germination was calculated as the number of seeds that germinated divided by the total number of seeds recovered because some seeds may have been lost to rodents. We did not estimate seed viability, so it is possible that the proportion of germinated seeds was underestimated because the rodents may have preferred viable seeds. We calculated the average herbivory incidence of invertebrate foliar herbivory from three yes/no observations (2004, 2005, 2006) for each species at each distance and site to use as an explanatory variable in growth response models.
2.4. Data Analysis
We fit linear mixed-effects models to the averages of each growth response variable: biomass, diameter, and relative growth rate of height. Models were of the following form:
where y is the n-length vector of observed values for the response variable, which is multivariate normal conditional on B = b; X is the n × p fixed-effects matrix of predictor variables, with p equal to the number of explanatory variables; β is the p-length vector of fixed-effects coefficients to be estimated and apply to all species and sites; Z is the n × q random-effects model matrix for the q-dimensional unobserved random-effects variable B whose value is fixed at b, where q is the total number of levels in the grouping factors (i.e., 12 sites plus six species). B has an unconditional multivariate normal distribution with a mean of zero and covariance matrix D; 2 is the residual error.
(y|B = b) ~ N(X × β + Z × b, σ2)
B ~ N(0,D)
Our main goal was to compare species responses to the influences of edge effects and forest successional age. We assessed all potential explanatory variables for collinearity using scatterplots and Pearson’s correlation coefficients (threshold value of ±0.5) between continuous variables, and box and whisker plots between continuous and categorical variables. We selected forest successional age, distance from edge, average herbivory incidence, and the averaged percentage of available light as fixed effects. We included random effects for site and species, to account for unmeasured species-level and site-level variation (random intercepts). We also allowed the relationship between herbivory incidence and the responses to vary by species (random slopes), reflecting potential interaction between species and herbivory due to species differences in defense or compensatory responses. Soil moisture and forest basal area were excluded from the models due to correlations with distance from edge and forest successional age, respectively. Biomass and diameter response variables were natural logarithm-transformed to improve homogeneity of residual variance and normality.
We fit binomial mixed-effects logistic regressions to model the probability of survival and germination for each species:
where y is binomially distributed with an occurrence probability p, which is modeled as a function of linear predictors using the logit link function. The details of the linear predictors are the same as those in linear model described above. In all models we allowed the intercepts to vary for species and site, enabling comparison of species performance relative to each other and accounting for unmeasured site characteristics. We checked the residual plots of the final models to ensure no major violations of homogeneity in the residual variance.
(y|B = b) ~ Binomial(n, p|b)
logit(p|b) = X × β + Z × b
All models were fit using the “lme4” [47] and “lmerTest” [48] packages in the R statistical computing environment [49]. The coefficient estimates from rescaled continuous variables are reported in the figures. Percent light, litter mass, and distance from edge were centered on a mean of zero and divided by two standard deviations, and distance was rescaled to be between zero and one by dividing by the maximum distance value. Dividing continuous input variables by two standard deviations instead of one standard deviation effectively puts them on the same scale as binary predictors, facilitating a more direct comparison of relative importance [50].
3. Results
3.1. Habitat Conditions
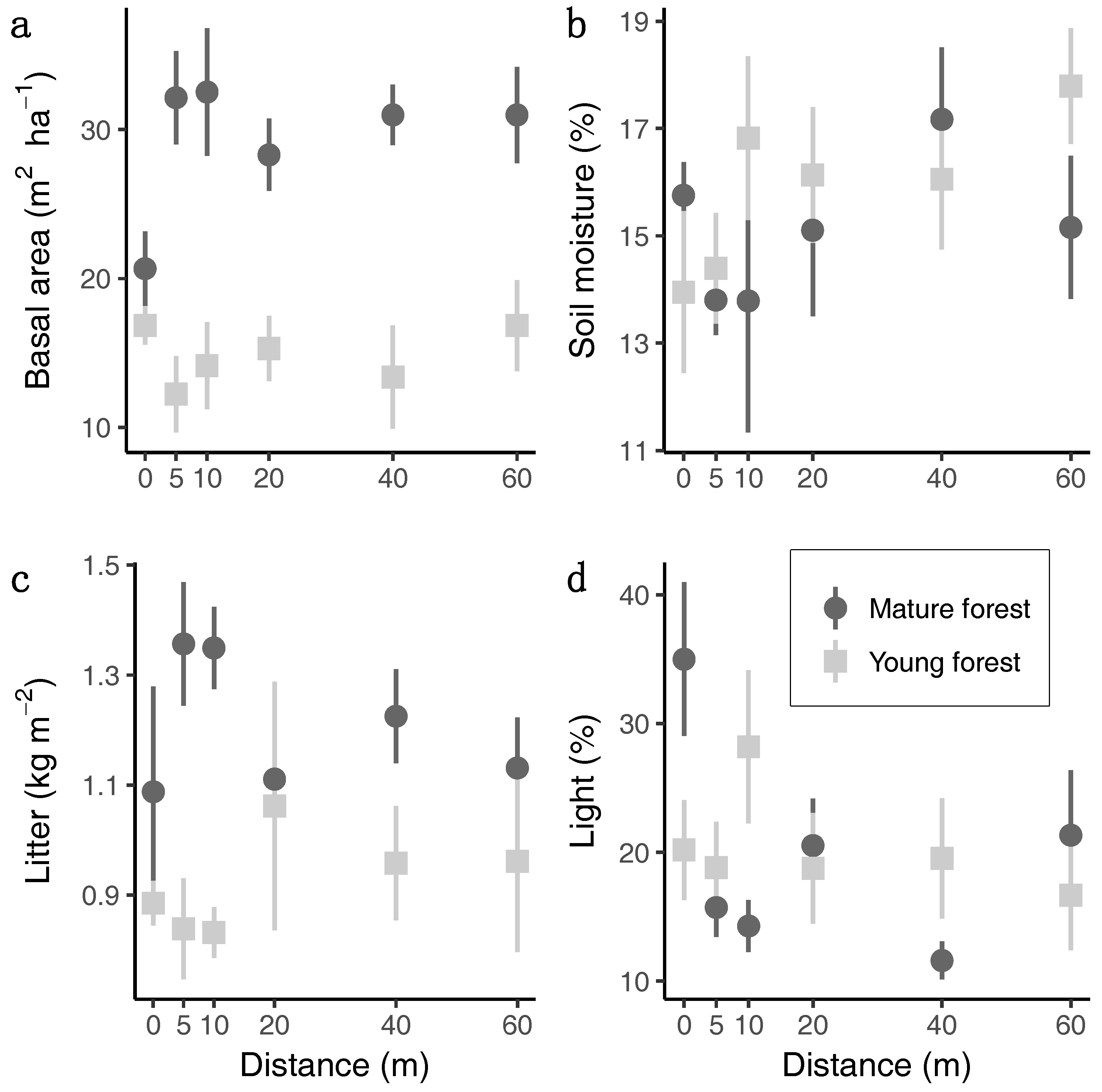
Basal area and litter mass were both greater in mature than in young forests and neither showed a relationship with distance (Figure 1a,c). Soil moisture and light availability showed no distinctive pattern between forest successional ages (Figure 1b,d). Soil moisture tended to increase with distance in both forest successional ages (Figure 1b). Light availability tended to decrease with distance in mature but not young forests (Figure 1c).
3.2. Germination and Survival
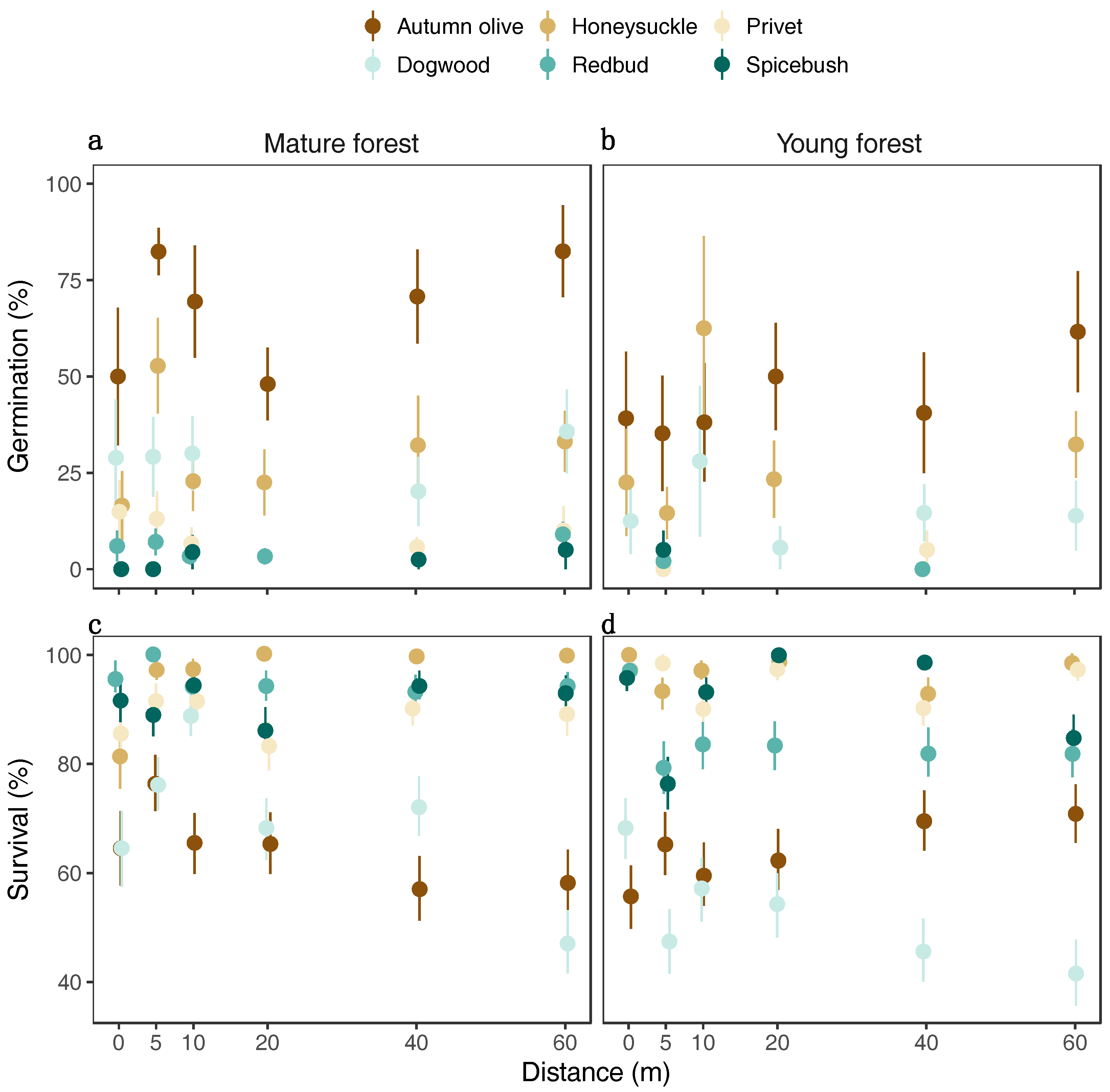
Seed germination was higher in mature compared to young forests across all species except for spicebush and was overall higher for invasive than native species (Figure 2a). The average proportion of seeds germinated ranged from 1% for redbud in young forests to 67% for autumn olive in mature forests (Table A1). The greater germination success of invasive species was driven by autumn olive, which had the highest germination in both mature and young forest types.
Average seedling survival ranged from 42% for native dogwood in young forests to 96% for invasive honeysuckle in mature forests (Table A2). The native dogwood and invasive autumn olive seedlings had the lowest overall survival: 51% and 50%, respectively. Autumn olive survival decreased with increasing distance from forest edge in mature but not young forests (Figure 2c,d), whereas other species’ survival was minimally affected by distance from edge in both forest types.
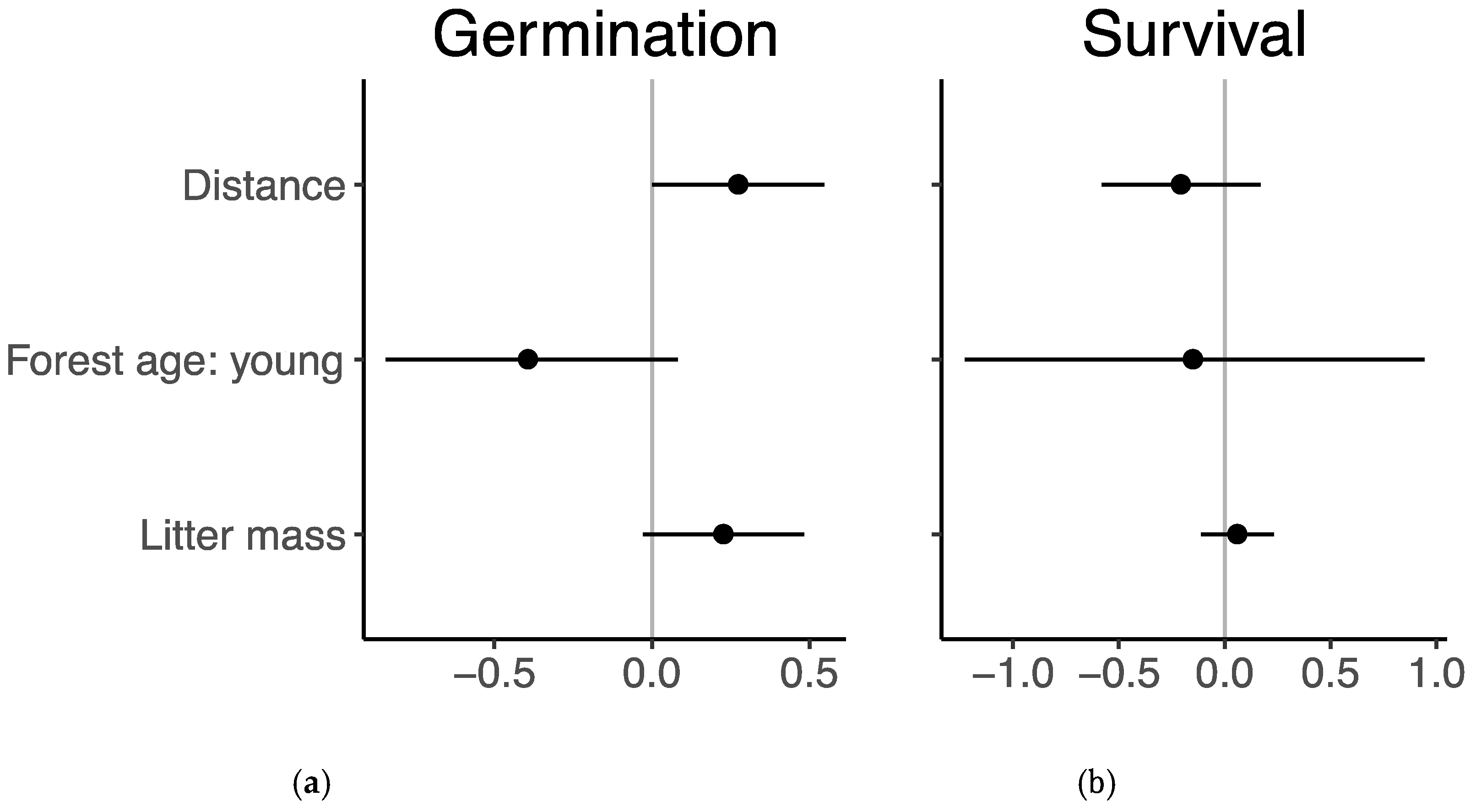
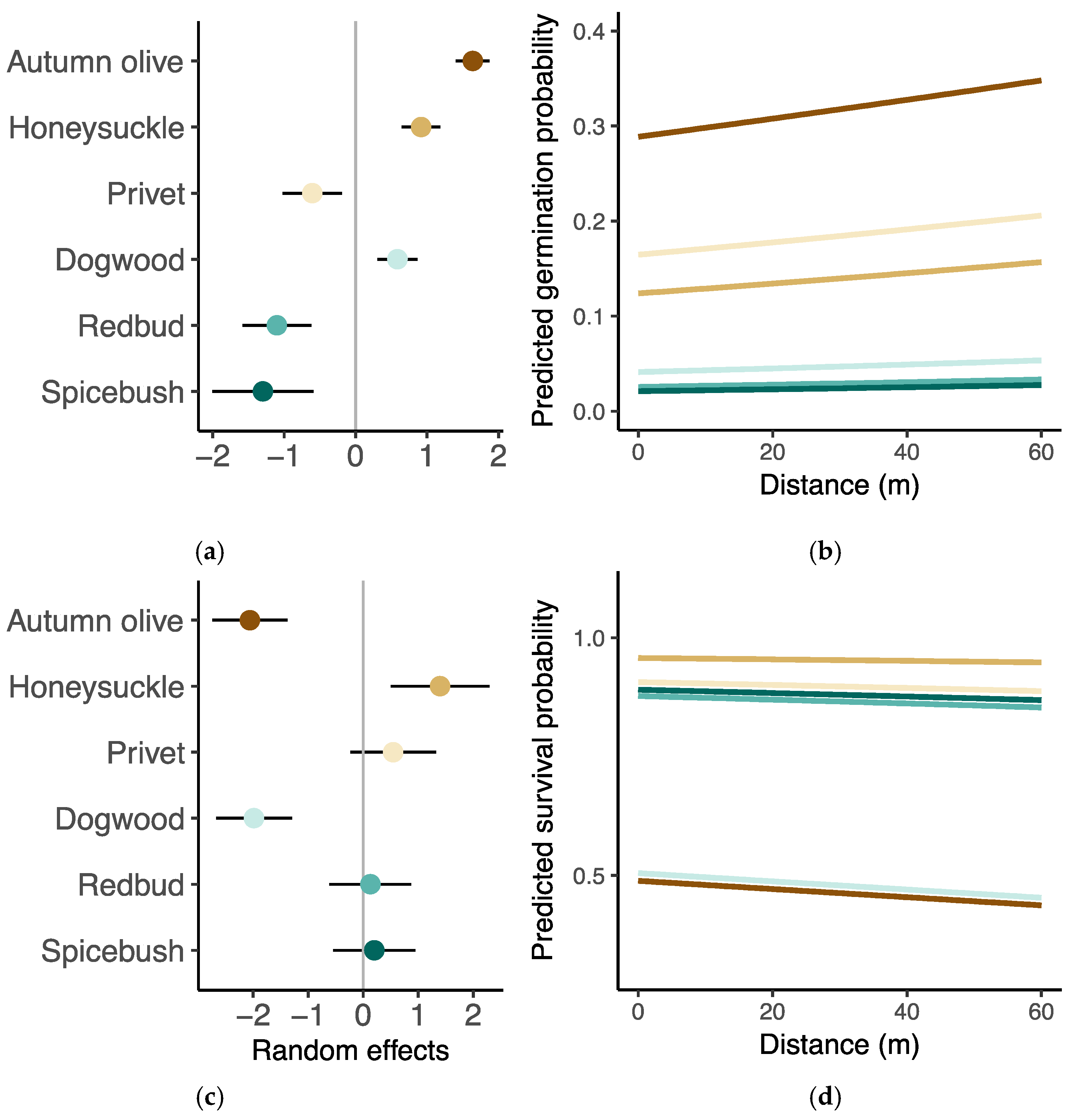
Model results indicate that the probability of germination was significantly greater with greater distance from edge and tended to be lower in young forests and greater with higher litter mass, although these two relationships were not statistically significant (Figure 3a, Table A3). No explanatory variables had a statistically significant effect on seedling survival (Figure 3b, Table A3). Species differences accounted for much of the variation in both germination and survival, with some having a high proportion of germination in the seed experiment but low survival in the seedling transplant experiment (Figure 4). Autumn olive had the highest proportion of germinated seeds in the germination experiment but the lowest survival in the seedling transplant experiment. Dogwood showed a similar pattern of relatively greater germination but lower survival. Privet had the opposite pattern with greater survival, though not significantly different from the average, but lower germination. Redbud and spicebush both had low germination and seedling survival was near the overall average. Honeysuckle stood out as having above average germination (trailing only autumn olive) and the greatest survival with over 95% of seedlings surviving to the end of the experiment (Table A2).
3.3. Seedling Performance
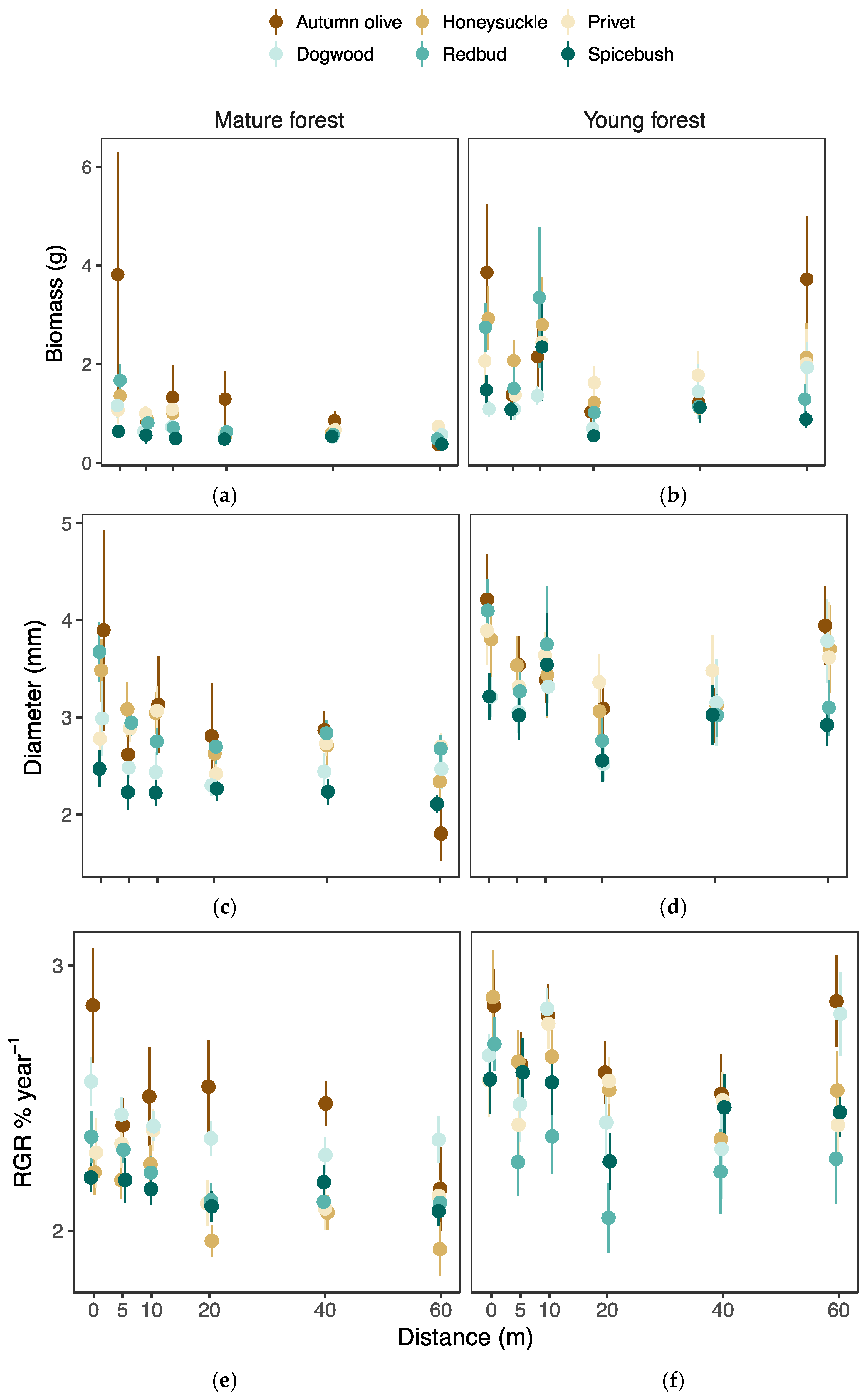
Invasive species generally performed better than the native species across forest successional age; however, the performance of just two or three species drove this pattern. For example, the invasive autumn olive and privet consistently produced larger amounts of biomass, while the native spicebush produced little biomass (Figure 5a,b). Similar patterns were observed for diameter (Figure 5c,d) and relative growth rate (Figure 5e,f).
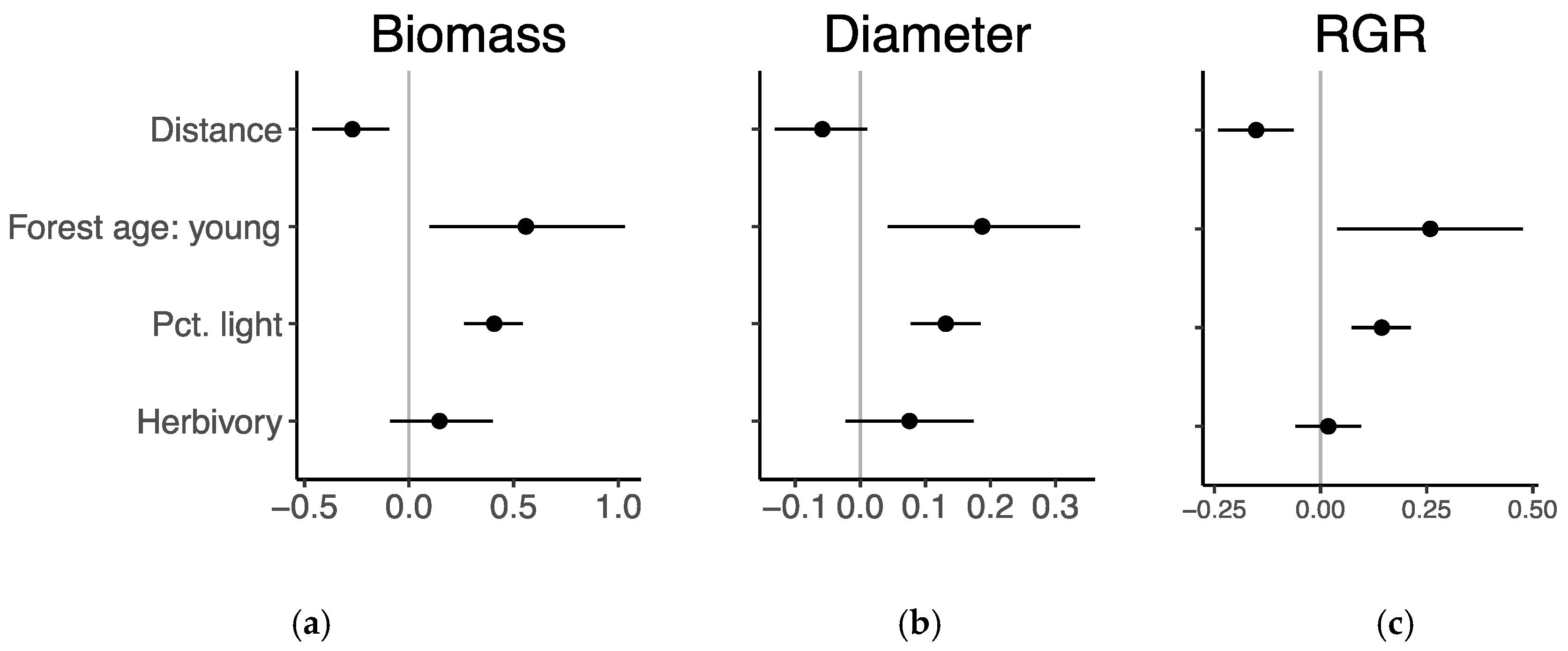
Model results indicated that proximity to forest edge, forest successional age, light availability, and herbivory all similarly influenced the biomass, diameter, and relative growth rate of height of transplanted seedlings (Figure 6, Table A4). Across all species, performance was greater closer to forest edge, in young forests, and under higher light availability.
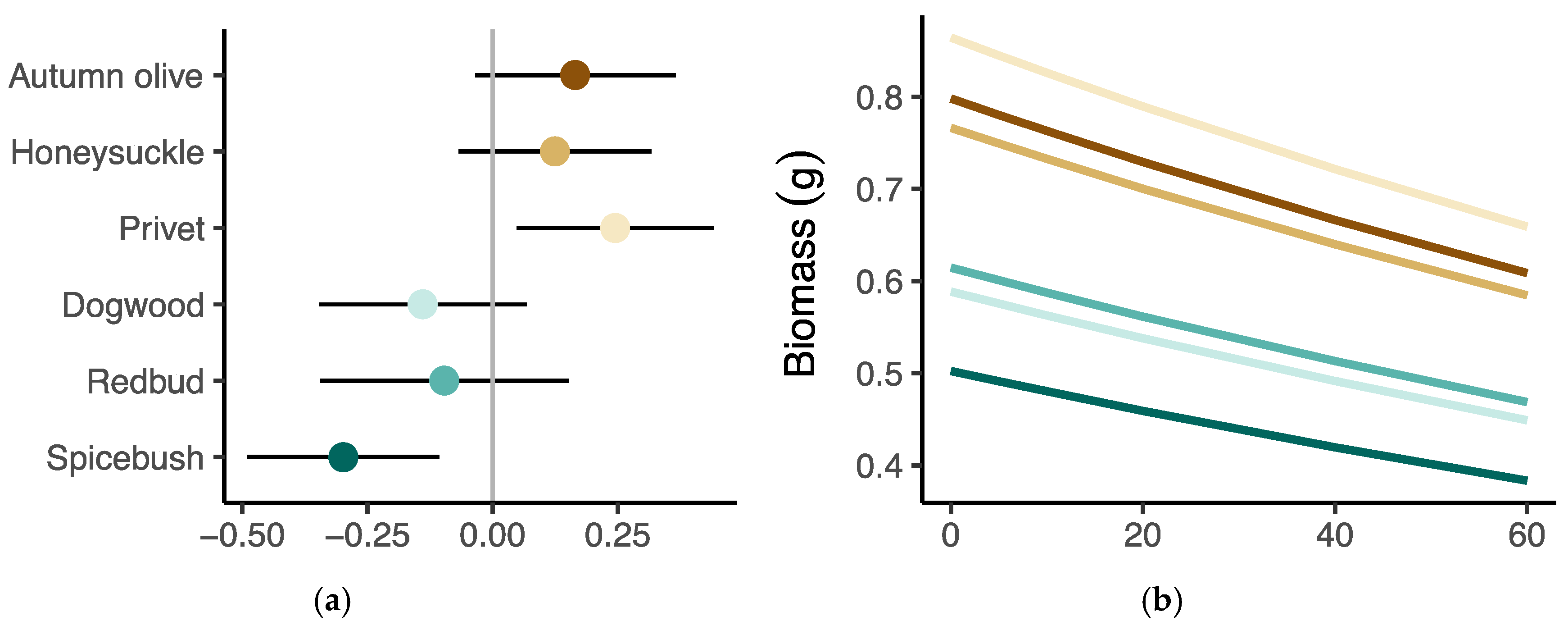
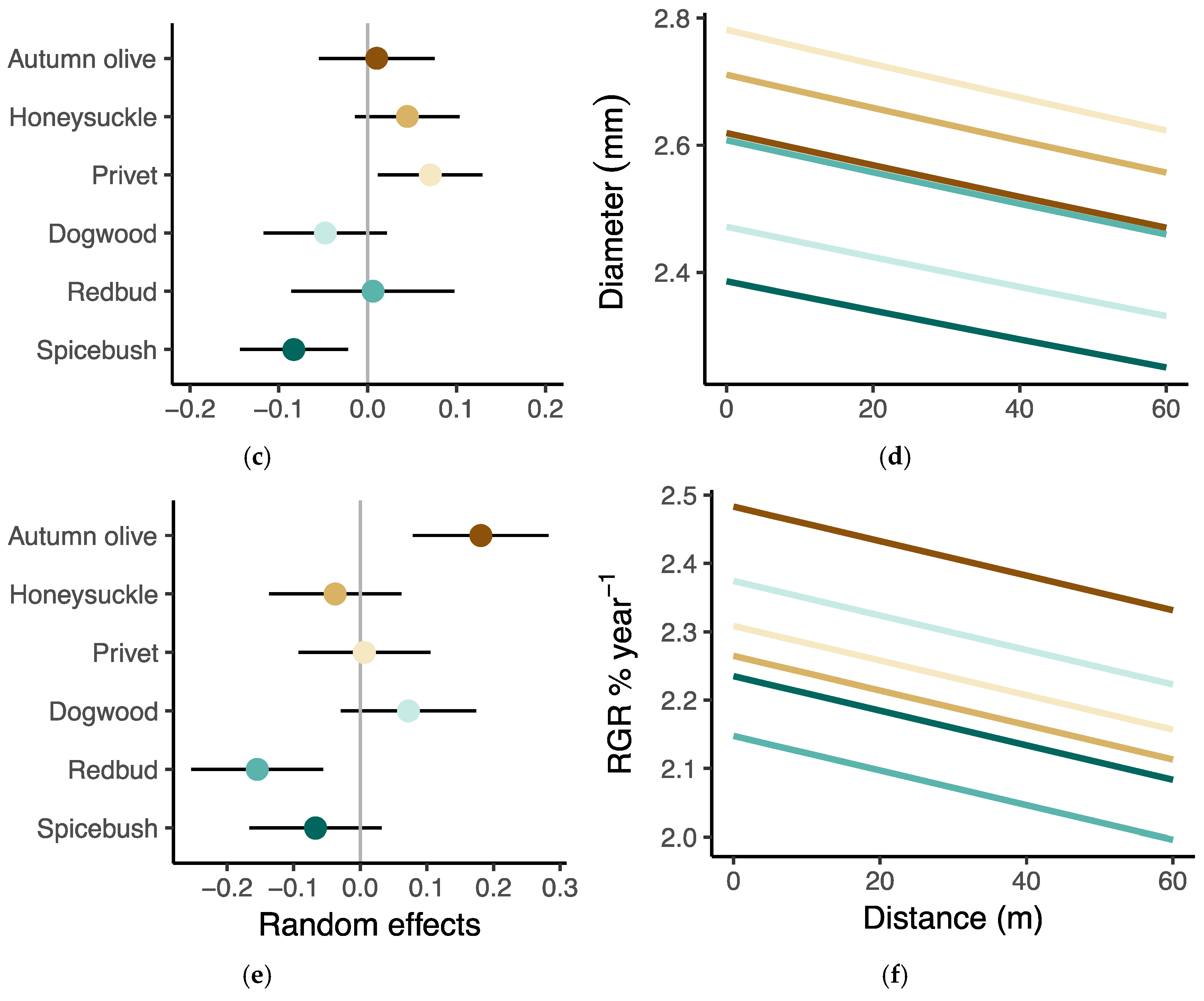
Model estimated average biomass of privet, autumn olive, and honeysuckle was greater than the mean across all species, though only privet had 95% confidence intervals that did not cross zero (Figure 7a). The average biomass estimated from the model was less than the overall mean for all the native species, though only spicebush had confidence intervals that did not overlap zero (Figure 7a). On average, invasive privet had the greatest relative diameter while native spicebush had the smallest relative diameter, while the model estimates for all other species had confidence intervals crossing zero (Figure 7c). Model results indicated that autumn olive had the highest average relative growth rate in height while spicebush had the lowest (Figure 7e,f).
3.4. Growth Response to Herbivory
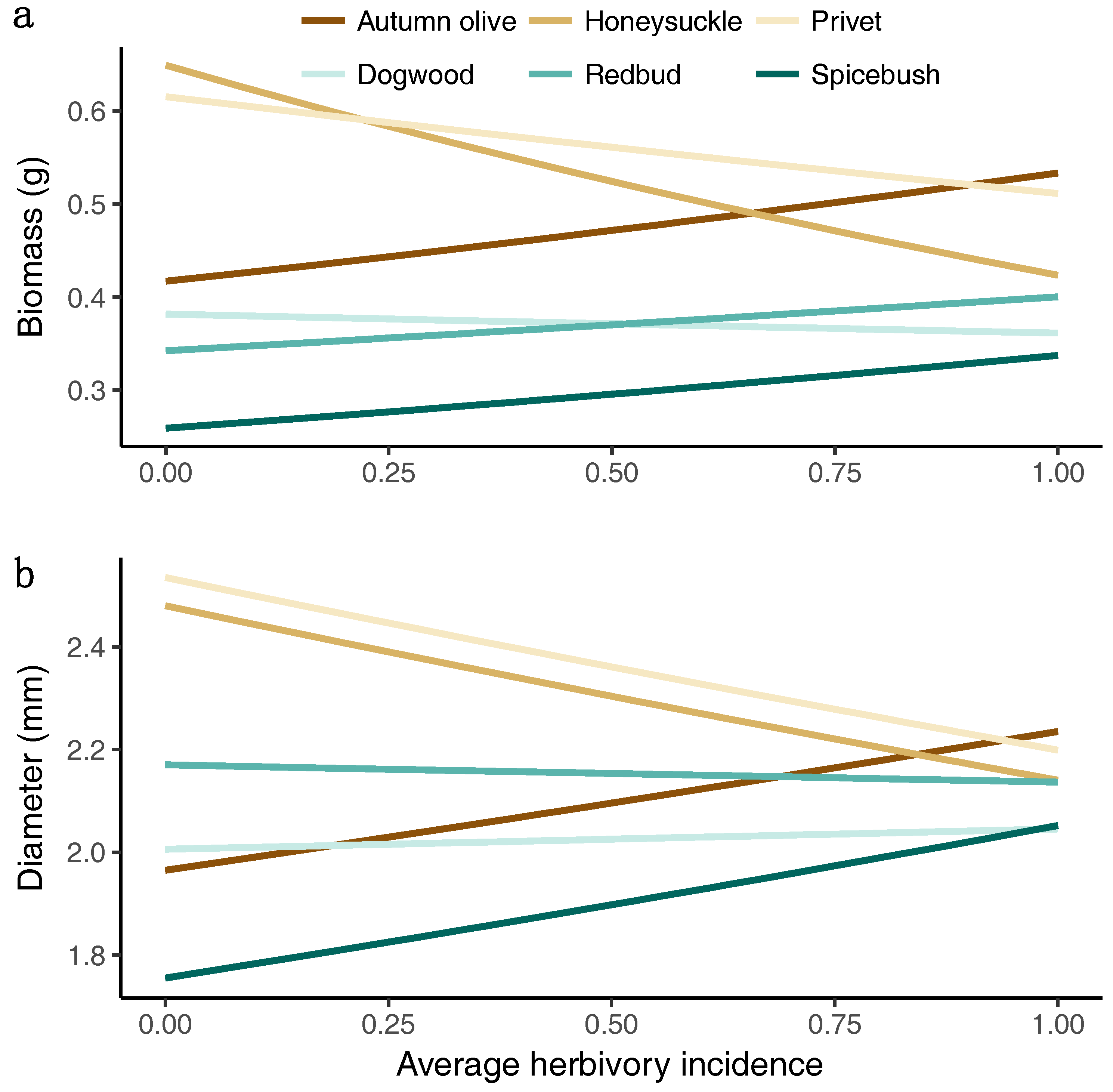
Model results showed that species also responded differently to observed average herbivory incidence (Figure 8). Native redbud and dogwood tended to maintain biomass regardless of herbivory, while the invasive privet and honeysuckle produced less biomass with greater herbivory. The invasive autumn olive and native spicebush both tended to have greater biomass and diameter with more herbivory. The relative growth rate was maintained for all species regardless of average herbivory incidence (not shown).
4. Discussion
The functional traits of invasive species and local environmental conditions are key determinants of habitat susceptibility to invasion. Areas of greater resource availability, such as forest edges, can be more susceptible to invasions if the functional traits of invaders enable them to outperform native species under such conditions [1,3,51,52]. Here, we compared the performance of commonly occurring native and invasive species at multiple distances from forest edges in both young and mature forests. We found that species performance was variable across the range of conditions regardless of nativity. The invasive species generally outperformed native species, but the differences between the groups were largely driven by a few species. The variability in species responses is likely related to different resource allocation strategies among species. Factors contributing to better performance by invaders may include advantageous use of temporally variable light availability, enemy release, allelopathy, and association with nitrogen-fixing bacteria [53]. Using experimental introductions to evaluate if invasive and native species respond differently to variable forest conditions can help identify non-native species likely to be invasive and the most vigorous native species.
Many invasive plants exhibit common traits that can contribute to their success, such as extended leaf phenology, rapid growth, tolerance to low resource conditions, and relatively little damage from natural enemies [21,22,23,24,26], but individual invaders likely only benefit from a few of these traits. For example, one invasive species may experience enemy release and high fecundity while another tolerates low resource conditions and has extended leaf phenology. These trait differences may explain some of the species-specific variation in invader performance we observed in this study. All three invasive species in this study have been characterized as having longer leaf duration during the growing season than native species, so the relatively greater growth of autumn olive, privet, and honeysuckle is perhaps due to advantages conferred by early photosynthesis before native species leaf out [38,39,54,55]. In contrast to shade-tolerance exhibited by privet and honeysuckle, autumn-olive showed characteristics of a sun-demanding species, with the greatest growth at forest edges and less growth in the forest interior. Despite the shade-intolerance of autumn olive, our results indicate that this species performs as well or better than the native species in most situations, perhaps due to other common invader traits such as drought tolerance or a nitrogen-fixing symbiont [3,31]. Dechoum et al. [56] found that Hovenia dulcis, an invasive tree in Brazil, exhibited a growth response similar to autumn olive, growing at about the same rate in closed and semi-open successional habitats, while having somewhat greater growth in more open areas. Greater production of fine roots by invasive species has also been linked to their greater production of aboveground biomass [57].
The invasive species in this study all tend to have greater abundance at forest edges than interiors. Previous studies on honeysuckle have shown that it produces abundant fruit and seed in high light environments but still produces fruit and seed in forest interiors [32], and that density is positively related to amount of edge in the landscape [58]. Similar relationships of greater abundance along forest edges have been observed for autumn olive [59] and privet [60,61], and all of these species can dominate the understory and reduce diversity and abundance of native species. In each case, the abundance of the invasive species declined but still persisted farther from the forest edge. Our results showing less growth farther from forest edges support these previous findings.
While proximity to edge often correlates positively with the occurrence and abundance of invasive species [19,62], the relatively poor performance of native species under the same conditions may be due to adaptations to their interior understory niche. The relatively low germination but high survivorship of native species in combination with traits such as shade tolerance and slow growth enable these species to persist in the resource-limited understory of interior eastern USA forests. A similar response of a native tree was observed in a tropical forest in Amazonia where seedling survival and leaf number increased with distance from edge for native Chrysophyllum pomiferum (Sapotaceae) seedlings over a 9.5-year period [63]. Yet, the traits enabling the success of native species in resource-limited interior forest habitats may put them at a disadvantage to phenotypically plastic invaders that are more effective at acquiring and allocating resources when availability increases due to large-scale forest fragmentation [64].
Although some invasive species are persistent, the density and distribution of invasive plants may decline over time under certain conditions. For example, Flory et al. [65] found that experimental Microstegium vimineum invasions declined over eight years after dominating the herbaceous layer for the first four years. Multiple factors may contribute to the decline of Microstegium and other invasive species, such as succession of competitive native species or accumulation of pathogens [66,67]. Of the invasive species in our study, we are aware of the potential decline of only one species, honeysuckle, which exhibited leaf-blight in the midwestern USA [68], suggesting that this species could be suffering from pathogen accumulation. Additional research is needed to determine how patterns in success of early life-history stages relate to longer-term invasive species success.
By conducting a multi-year field experiment we were able to examine the influence of natural herbivory on plant performance over multiple growing seasons. The maintenance of biomass by native species regardless of herbivory incidence suggests greater defense against herbivory, e.g., high phenolic content in dogwood [69]. Meanwhile, the invasive privet and honeysuckle produced less biomass with greater herbivory, indicating some susceptibility to herbivory in the seedling stage. However, Lieurance and Cipollini [70] demonstrated that while honeysuckle experienced lower biomass in response to generalist herbivory, it also had a greater root to shoot ratio, and more foliar protein with greater herbivory, indicating both a tolerance and resistance to herbivory. The greater biomass with more herbivory shown by the invasive autumn olive and native spicebush indicates a possible compensatory growth response to damage. These findings add to the increasing body of work documenting the changes in herbivory across the landscape, particularly along forest edges [13,71].
5. Conclusions
Experimentally introducing multiple native and co-occurring invasive species in a natural setting enabled the assessment of differences in species performance under similar environmental conditions. These results indicate that accounting for species-specific traits may improve the efficacy of invasive species management. Differences in species characteristics and traits enable some plants to more rapidly utilize resources that become available after disturbance. The native redbud performed on par with the invasive species under some conditions, indicating it may be a good species for restoration management in these forests. Still, forest edges were most beneficial to invasive species due to the relatively rapid growth response of invasive compared to native species. Thus, new edges that are created through road building or development may increase the susceptibility of forests to invasive plant species. Efficient invasive species management requires vigilance to detect invasive species at the earliest stages, which can be aided by identification of species likely to become problematic due to their specific traits. While species-specific differences confound the ability to generalize mechanisms of invasion, they also highlight the importance of understanding species-specific processes contextually in the research and management of invasive plants.
Author Contributions
Conceptualization, D.L., K.C. and S.L.F.; Data curation, S.L.F.; Formal analysis, W.W.D.; Investigation, K.C. and S.L.F.; Methodology, K.C. and S.L.F.; Resources, K.C.; Visualization, W.W.D. and D.T.H.; Writing—original draft, W.W.D., D.L., D.T.H. and S.L.F.; Writing—review & editing, W.W.D., D.L., D.T.H., K.C. and S.L.F.
Funding
This project was funded by the Indiana Graduate and Professional Student Organization, the Indiana Academy of Sciences, and the Indiana University Research and Teaching Preserve.
Acknowledgments
We thank the owners and managers of the properties where the field experiment was implemented who provided access and logistical support.
Conflicts of Interest
The authors declare no conflict of interest. The funding sponsors had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, and in the decision to publish the results.
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table A1.
Average germination (%) of seeds for each species in each forest age and overall.
| Species | Status | Mature Forest | Young Forest | Average |
|---|---|---|---|---|
| Autumn olive | Invasive | 67.19 | 44.07 | 55.63 |
| Honeysuckle | Invasive | 29.99 | 25.34 | 27.66 |
| Privet | Invasive | 8.21 | 0.97 | 4.59 |
| Dogwood | Native | 23.85 | 12.34 | 18.09 |
| Redbud | Native | 4.77 | 0.96 | 2.86 |
| Spicebush | Native | 2.18 | 10.00 | 6.09 |
Table A2.
Average percentage of seedlings of each species surviving after three years (to 2006) in mature and young forests.
Table A2.
Average percentage of seedlings of each species surviving after three years (to 2006) in mature and young forests.
| Species | Status | Mature Forest | Young Forest | Average |
|---|---|---|---|---|
| Autumn olive | Invasive | 48.53 | 50.69 | 49.61 |
| Honeysuckle | Invasive | 95.56 | 95.14 | 95.35 |
| Privet | Invasive | 85.29 | 93.06 | 89.17 |
| Dogwood | Native | 61.03 | 41.67 | 51.35 |
| Redbud | Native | 91.91 | 80.56 | 86.23 |
| Spicebush | Native | 86.76 | 88.19 | 87.48 |
Table A3.
Results from generalized linear mixed effects models of germination and survival. Odds ratios and 95% confidence intervals (CI) are reported using the scaled predictor variables.
Table A3.
Results from generalized linear mixed effects models of germination and survival. Odds ratios and 95% confidence intervals (CI) are reported using the scaled predictor variables.
| Fixed Parts | Germination | Survival | ||
|---|---|---|---|---|
| Odds Ratio | CI | Odds Ratio | CI | |
| (Intercept) | 0.11 | 0.04–0.30 | 5.92 | 1.74–20.16 |
| Distance | 1.31 | 1.00–1.73 | 0.82 | 0.57–1.20 |
| Forest age (Young) | 0.67 | 0.44–1.03 | 0.87 | 0.32–2.37 |
| Litter mass | 1.25 | 0.97–1.62 | 1.08 | 0.91–1.29 |
| Light availability | 1.08 | 0.92–1.28 | ||
| Random Parts | ||||
| Variance, site_code | 0.087 | 0.710 | ||
| Variance, common_name | 1.269 | 1.514 | ||
Table A4.
Results from linear mixed effects models of biomass, diameter, and relative growth rate. Estimates (B) and 95% confidence intervals (CI) are reported using the scaled predictor variables.
Table A4.
Results from linear mixed effects models of biomass, diameter, and relative growth rate. Estimates (B) and 95% confidence intervals (CI) are reported using the scaled predictor variables.
| Fixed Parts | Biomass | Diameter | RGR | |||
|---|---|---|---|---|---|---|
| B | CI | B | CI | B | CI | |
| (Intercept) | −0.39 | −0.77–−0.01 | 0.95 | 0.83–1.07 | 2.30 | 2.12–2.49 |
| Distance | −0.27 | −0.45–−0.09 | −0.06 | −0.13–0.01 | −0.15 | −0.24–−0.06 |
| Light availability | 0.41 | 0.27–0.55 | 0.13 | 0.08–0.19 | 0.14 | 0.08–0.21 |
| Forest age (Young) | 0.56 | 0.10–1.02 | 0.19 | 0.04–0.33 | 0.26 | 0.04–0.47 |
| Herbivory incidence | 0.14 | −0.08–0.37 | 0.08 | −0.02–0.17 | 0.02 | −0.06–0.10 |
| Random Parts | ||||||
| σ2 | 0.391 | 0.059 | 0.098 | |||
| Variance, site | 0.155 | 0.015 | 0.034 | |||
| Variance, species | 0.051 | 0.004 | 0.015 | |||
References
- Davis, M.A.; Grime, J.P.; Thompson, K. Fluctuating resources in plant communities: A general theory of invasibility. J. Ecol. 2000, 88, 528–534. [Google Scholar] [CrossRef]
- Haddad, N.M.; Brudvig, L.A.; Clobert, J.; Davies, K.F.; Gonzalez, A.; Holt, R.D.; Lovejoy, T.E.; Sexton, J.O.; Austin, M.P.; Collins, C.D.; et al. Habitat fragmentation and its lasting impact on Earth’s ecosystems. Sci. Adv. 2015, 1, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Yates, E.D.; Levia, D.F.; Williams, C.L. Recruitment of three non-native invasive plants into a fragmented forest in southern Illinois. For. Ecol. Manag. 2004, 190, 119–130. [Google Scholar] [CrossRef]
- Alston, K.P.; Richardson, D.M. The roles of habitat features, disturbance, and distance from putative source populations in structuring alien plant invasions at the urban/wildland interface on the Cape Peninsula, South Africa. Biol. Conserv. 2006, 132, 183–198. [Google Scholar] [CrossRef]
- Moser, W.K.; Fan, Z.; Hansen, M.H.; Crosby, M.K.; Fan, S.X. Invasibility of three major non-native invasive shrubs and associated factors in Upper Midwest U.S. forest lands. For. Ecol. Manag. 2016, 379, 195–205. [Google Scholar] [CrossRef] [Green Version]
- Brothers, T.S.; Spingarn, A. Forest Fragmentation and Alien Plant Invasion of Central Indiana Old-Growth Forests. Conserv. Biol. 1992, 6, 91–100. [Google Scholar] [CrossRef]
- Hansen, M.J.; Clevenger, A.P. The influence of disturbance and habitat on the presence of non-native plant species along transport corridors. Biol. Conserv. 2005, 125, 249–259. [Google Scholar] [CrossRef]
- Von Der Lippe, M.; Kowarik, I. Long-Distance Dispersal of Plants by Vehicles as a Driver of Plant Invasions. Conserv. Biol. 2007, 21, 986–996. [Google Scholar] [CrossRef] [PubMed]
- Rew, L.J.; Brummer, T.J.; Pollnac, F.W.; Larson, C.D.; Taylor, K.T.; Taper, M.L.; Fleming, J.D.; Balbach, H.E. Hitching a ride: Seed accrual rates on different types of vehicles. J. Environ. Manag. 2018, 206, 547–555. [Google Scholar] [CrossRef] [PubMed]
- Simberloff, D. The Role of Propagule Pressure in Biological Invasions. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 81–102. [Google Scholar] [CrossRef]
- Damschen, E.I.; Baker, D.V.; Bohrer, G.; Nathan, R.; Orrock, J.L.; Turner, J.R.; Brudvig, L.A.; Haddad, N.M.; Levey, D.J.; Tewksbury, J.J. How fragmentation and corridors affect wind dynamics and seed dispersal in open habitats. Proc. Natl. Acad. Sci. USA 2014. [Google Scholar] [CrossRef] [PubMed]
- Murcia, C. Edge effects in fragmented forests: Implications for conservation. Trends Ecol. Evol. 1995, 10, 58–62. [Google Scholar] [CrossRef]
- Cadenasso, M.L.; Pickett, S.T.A. Effect of Edge Structure on the Flux of Species into Forest Interiors. Conserv. Biol. 2001, 15, 91–97. [Google Scholar] [CrossRef]
- Harper, K.A.; Macdonald, S.E.; Burton, P.J.; Chen, J.; Brosofske, K.D.; Saunders, S.C.; Euskirchen, E.S.; Roberts, D.; Jaiteh, M.S.; Esseen, P.A. Edge Influence on Forest Structure and Composition in Fragmented Landscapes. Conserv. Biol. 2005, 19, 768–782. [Google Scholar] [CrossRef]
- Wirth, R.; Meyer, S.T.; Leal, I.R.; Tabarelli, M. Plant Herbivore Interactions at the Forest Edge. In Progress in Botany; Lüttge, U., Beyschlag, W., Murata, J., Eds.; Springer: Berlin, Germany, 2008; Volume 69, pp. 423–448. ISBN 978-3-540-72953-2. [Google Scholar]
- De Carvalho Guimarães, C.D.; Viana, J.P.R.; Cornelissen, T. A Meta-Analysis of the Effects of Fragmentation on Herbivorous Insects. Environ. Entomol. 2014, 43, 537–545. [Google Scholar] [CrossRef] [PubMed]
- Matlack, G.R. Microenvironment variation within and among forest edge sites in the eastern United States. Biol. Conserv. 1993, 66, 185–194. [Google Scholar] [CrossRef]
- Funk, J.L.; McDaniel, S. Altering Light Availability to Restore Invaded Forest: The Predictive Role of Plant Traits. Restor. Ecol. 2010, 18, 865–872. [Google Scholar] [CrossRef]
- Flory, S.L.; Clay, K. Invasive shrub distribution varies with distance to roads and stand age in eastern deciduous forests in Indiana, USA. Plant Ecol. 2006, 184, 131–141. [Google Scholar] [CrossRef]
- Levine, J.M.; Adler, P.B.; Yelenik, S.G. A meta-analysis of biotic resistance to exotic plant invasions. Ecol. Lett. 2004, 7, 975–989. [Google Scholar] [CrossRef]
- Rejmanek, M.; Richardson, D.M. What Attributes Make Some Plant Species More Invasive? Ecology 1996, 77, 1655–1661. [Google Scholar] [CrossRef] [Green Version]
- Fridley, J.D. Extended leaf phenology and the autumn niche in deciduous forest invasions. Nature 2012, 485, 359–362. [Google Scholar] [CrossRef] [PubMed]
- Funk, J.L. The physiology of invasive plants in low-resource environments. Conserv. Physiol. 2013, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theoharides, K.A.; Dukes, J.S. Plant invasion across space and time: Factors affecting nonindigenous species success during four stages of invasion. New Phytol. 2007, 176, 256–273. [Google Scholar] [CrossRef] [PubMed]
- Keane, R.M.; Crawley, M.J. Exotic plant invasions and the enemy release hypothesis. Trends Ecol. Evol. 2002, 17, 164–170. [Google Scholar] [CrossRef]
- Mitchell, C.E.; Power, A.G. Release of invasive plants from fungal and viral pathogens. Nature 2003, 421, 625–627. [Google Scholar] [CrossRef] [PubMed]
- Rouget, M.; Richardson, D.M. Inferring Process from Pattern in Plant Invasions: A Semimechanistic Model Incorporating Propagule Pressure and Environmental Factors. Am. Nat. 2003, 162, 713–724. [Google Scholar] [CrossRef] [PubMed]
- Dreiss, L.M.; Volin, J.C. Influence of leaf phenology and site nitrogen on invasive species establishment in temperate deciduous forest understories. For. Ecol. Manag. 2013, 296, 1–8. [Google Scholar] [CrossRef]
- Kaufman, S.R.; Kaufman, W. Invasive Plants: A Guide to Identification and the Impacts and Control of Common North American Species; Stackpole Books: Mechanicsburg, PA, USA, 2013. [Google Scholar]
- Flory, S.L.; Clay, K. Effects of roads and forest successional age on experimental plant invasions. Biol. Conserv. 2009, 142, 2531–2537. [Google Scholar] [CrossRef]
- Naumann, J.C.; Bissett, S.N.; Young, D.R.; Edwards, J.; Anderson, J.E. Diurnal patterns of photosynthesis, chlorophyll fluorescence, and PRI to evaluate water stress in the invasive species, Elaeagnus umbellata Thunb. Trees 2010, 24, 237–245. [Google Scholar] [CrossRef]
- Lieurance, D.; Landsbergen, K. The influence of light habitat on the physiology, biomass allocation, and fecundity of the invasive shrub Amur honeysuckle (Lonicera maackii, Caprifoliaceae). J. Torrey Bot. Soc. 2016, 143, 415–426. [Google Scholar] [CrossRef]
- Wang, H.H.; Grant, W.E. Determinants of Chinese and European Privet (Ligustrum sinense and Ligustrum vulgare) Invasion and Likelihood of Further Invasion in Southern U.S. Forestlands. Invasive Plant Sci. Manag. 2012, 5, 454–463. [Google Scholar] [CrossRef]
- Pisula, N.L.; Meiners, S.J. Relative allelopathic potential of invasive plant species in a young disturbed woodland. J. Torrey Bot. Soc. 2010, 137, 81–87. [Google Scholar] [CrossRef]
- Montti, L.; Ayup, M.M.; Aragón, R.; Qi, W.; Ruan, H.; Fernández, R.; Casertano, S.A.; Zou, X. Herbivory and the success of Ligustrum lucidum: Evidence from a comparison between native and novel ranges. Aust. J. Bot. 2016, 64, 181–192. [Google Scholar] [CrossRef]
- Dorning, M.; Cipollini, D. Leaf and Root Extracts of the Invasive Shrub, Lonicera maackii, Inhibit Seed Germination of Three Herbs with No Autotoxic Effects. Plant Ecol. 2006, 184, 287–296. [Google Scholar] [CrossRef]
- Lieurance, D.; Cipollini, D. Exotic Lonicera species both escape and resist specialist and generalist herbivores in the introduced range in North America. Biol. Invasions 2013, 15, 1713–1724. [Google Scholar] [CrossRef]
- Nesom, G.L. Taxonomic Overview of Ligustrum (Oleaceae) Naturalizaed in North America North of Mexico. Phytologia 2009, 91, 467–482. [Google Scholar]
- McEwan, R.W.; Birchfield, M.K.; Schoergendorfer, A.; Arthur, M.A. Leaf Phenology and Freeze Tolerance of the Invasive Shrub Amur Honeysuckle and Potential Native Competitors. J. Torrey Bot. Soc. 2009, 136, 212–220. [Google Scholar] [CrossRef]
- Jenkins, M.A.; White, P.S. Cornus florida L. Mortality and Understory Composition Changes in Western Great Smoky Mountains National Park. J. Torrey Bot. Soc. 2002, 129, 194. [Google Scholar] [CrossRef]
- Niesenbaum, R.A. The Effects of Light Environment on Herbivory and Growth in the Dioecious Shrub Lindera benzoin (Lauraceae). Am. Midl. Nat. 1992, 128, 270. [Google Scholar] [CrossRef]
- Luken, J.O.; Kuddes, L.M.; Tholemeier, T.C.; Haller, D.M. Comparative Responses of Lonicera maackii (Amur honeysuckle) and Lindera benzoin (Spicebush) to Increased Light. Am. Midl. Nat. 1997, 138, 331. [Google Scholar] [CrossRef]
- Holzmueller, E.; Jose, S.; Jenkins, M.; Camp, A.; Long, A. Dogwood Anthracnose in Eastern Hardwood Forests: What Is Known and What Can Be Done? J. For. 2006, 104, 21–26. [Google Scholar]
- Burns, R.M.; Honkala, B.H. Silvics of North America; United States Department of Agriculture: Washington, DC, USA, 1990; Volume 1–2.
- Lieth, H.; Radford, J.S. Phenology, Resource Management, and Synagraphic Computer Mapping. BioScience 1971, 21, 62–70. [Google Scholar] [CrossRef]
- Herrera, C.M. Microclimate and Individual Variation in Pollinators: Flowering Plants are More than Their Flowers. Ecology 1995, 76, 1516–1524. [Google Scholar] [CrossRef] [Green Version]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–51. [Google Scholar] [CrossRef]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest Package: Tests in Linear Mixed Effects Models. J. Stat. Softw. 2017, 82, 1–26. [Google Scholar] [CrossRef]
- R: A Language and Environment for Statistical Computing. Available online: http://citeseerx.ist.psu.edu/viewdoc/download?doi=10.1.1.470.5851&rep=rep1&type=pdf (accessed on 15 September 2017).
- Gelman, A. Scaling regression inputs by dividing by two standard deviations. Stat. Med. 2008, 27, 2865–2873. [Google Scholar] [CrossRef] [PubMed]
- Delgado, J.D.; Arroyo, N.L.; Arévalo, J.R.; Fernández-Palacios, J.M. Edge effects of roads on temperature, light, canopy cover, and canopy height in laurel and pine forests (Tenerife, Canary Islands). Landsc. Urban Plan. 2007, 81, 328–340. [Google Scholar] [CrossRef]
- Van Kleunen, M.; Weber, E.; Fischer, M. A meta-analysis of trait differences between invasive and non-invasive plant species. Ecol. Lett. 2010, 13, 235–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Funk, J.L.; Vitousek, P.M. Resource-use efficiency and plant invasion in low-resource systems. Nature 2007, 446, 1079–1081. [Google Scholar] [CrossRef] [PubMed]
- Miller-Rushing Abraham, J.; Primack Richard, B. Global warming and flowering times in thoreau’s concord: A community perspective. Ecology 2008, 89, 332–341. [Google Scholar] [CrossRef]
- Brym, Z.T.; Lake, J.K.; Allen, D.; Ostling, A. Plant functional traits suggest novel ecological strategy for an invasive shrub in an understorey woody plant community: Plant functional traits and invasion. J. Appl. Ecol. 2011, 48, 1098–1106. [Google Scholar] [CrossRef]
- Dechoum, M.S.; Zenni, R.D.; Castellani, T.T.; Zalba, S.M.; Rejmánek, M. Invasions across secondary forest successional stages: Effects of local plant community, soil, litter, and herbivory on Hovenia dulcis seed germination and seedling establishment. Plant Ecol. 2015, 216, 823–833. [Google Scholar] [CrossRef]
- Jo, I.; Fridley, J.D.; Frank, D.A. Linking above- and belowground resource use strategies for native and invasive species of temperate deciduous forests. Biol. Invasions 2015, 17, 1545–1554. [Google Scholar] [CrossRef]
- Bartuszevige, A.M.; Gorchov, D.L.; Raab, L. The relative importance of landscape and community features in the invasion of an exotic shrub in a fragmented landscape. Ecography 2006, 29, 213–222. [Google Scholar] [CrossRef]
- Moore, M.R.; Buckley, D.S.; Klingeman, W.E.; Saxton, A.M. Distribution and growth of autumn olive in a managed forest landscape. For. Ecol. Manag. 2013, 310, 589–599. [Google Scholar] [CrossRef]
- Ward, R.W. Extent and Dispersal Rates of Chinese Privet (Ligustrum sinense) Invasion on the Upper Oconee River Floodplain, North Georgia. Southeast. Geogr. 2002, 42, 29–48. [Google Scholar] [CrossRef]
- Merriam, R.W.; Feil, E. The Potential Impact of an Introduced Shrub on Native Plant Diversity and Forest Regeneration. 5. Biol. Invasions 2002, 4, 369–373. [Google Scholar] [CrossRef]
- Riitters, K.; Potter, K.; Iannone, B.V.; Oswalt, C.; Fei, S.; Guo, Q. Landscape correlates of forest plant invasions: A high-resolution analysis across the eastern United States. Divers. Distrib. 2018, 24, 274–284. [Google Scholar] [CrossRef]
- Benitez-Malvido, J.; Lázaro, A.; Ferraz, I.D.K. Effect of Distance to Edge and Edge Interaction on Seedling Regeneration and Biotic Damage in Tropical Rain Forest Fragments: A Long-Term Experiment. J. Ecol. 2018. [Google Scholar] [CrossRef]
- Davidson, A.M.; Jennions, M.; Nicotra, A.B. Do invasive species show higher phenotypic plasticity than native species and, if so, is it adaptive? A meta-analysis. Ecol. Lett. 2011, 14, 419–431. [Google Scholar] [CrossRef] [PubMed]
- Flory, S.L.; Bauer, J.; Phillips, R.P.; Clay, K. Effects of a non-native grass invasion decline over time. J. Ecol. 2017, 105, 1475–1484. [Google Scholar] [CrossRef]
- Luke Flory, S.; Clay, K. Pathogen accumulation and long-term dynamics of plant invasions. J. Ecol. 2013, 101, 607–613. [Google Scholar] [CrossRef] [Green Version]
- Stricker, K.B.; Harmon, P.F.; Goss, E.M.; Clay, K.; Luke Flory, S. Emergence and accumulation of novel pathogens suppress an invasive species. Ecol. Lett. 2016, 19, 469–477. [Google Scholar] [CrossRef] [PubMed]
- Boyce, R.L.; Brossart, S.N.; Bryant, L.A.; Fehrenbach, L.A.; Hetzer, R.; Holt, J.E.; Parr, B.; Poynter, Z.; Schumacher, C.; Stonebraker, S.N.; et al. The beginning of the end? Extensive dieback of an open-grown Amur honeysuckle stand in northern Kentucky, USA. Biol. Invasions 2014, 16, 2017–2023. [Google Scholar] [CrossRef]
- Dudt, J.F.; Shure, D.J. The Influence of Light and Nutrients on Foliar Phenolics and Insect Herbivory. Ecology 1994, 75, 86–98. [Google Scholar] [CrossRef]
- Lieurance, D.; Cipollini, D. Environmental influences on growth and defence responses of the invasive shrub, Lonicera maackii, to simulated and real herbivory in the juvenile stage. Ann. Bot. 2013, 112, 741–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meiners, S.J.; Handel, S.N.; Pickett, S.T.A. Tree seedling establishment under insect herbivory: Edge effects and inter- annual variation. Plant Ecol. 2000, 151, 161–170. [Google Scholar] [CrossRef]
Figure 1.
The mean and standard error of the forest structure and environmental variables measured at each distance at each site. (a) Basal area, (b) soil moisture, (c) litter mass, (d) light availability.
Figure 1.
The mean and standard error of the forest structure and environmental variables measured at each distance at each site. (a) Basal area, (b) soil moisture, (c) litter mass, (d) light availability.

Figure 2.
The mean and standard error of the observed seed germination and seedling survival of each species in (a,c) mature or (b,d) young forests at each distance from the forest edge. The brown shading indicates the group of invasive species while the blue shading indicates the group of native species. Points are slightly jittered to reduce overlap at each distance.
Figure 2.
The mean and standard error of the observed seed germination and seedling survival of each species in (a,c) mature or (b,d) young forests at each distance from the forest edge. The brown shading indicates the group of invasive species while the blue shading indicates the group of native species. Points are slightly jittered to reduce overlap at each distance.

Figure 3.
Coefficient estimates ±95% confidence intervals of scaled coefficients for distance, forest successional age, and litter mass on (a) germination of seeds and (b) survival of seedlings.
Figure 3.
Coefficient estimates ±95% confidence intervals of scaled coefficients for distance, forest successional age, and litter mass on (a) germination of seeds and (b) survival of seedlings.

Figure 4.
Graphical results from mixed effects models of survival and germination. (a,c) Differences between species intercepts ±95% confidence intervals and overall model intercepts and (b,d) the marginal effect of distance on germination and survival for each species. The brown shading indicates the group of invasive species while the blue shading indicates the group of native species.
Figure 4.
Graphical results from mixed effects models of survival and germination. (a,c) Differences between species intercepts ±95% confidence intervals and overall model intercepts and (b,d) the marginal effect of distance on germination and survival for each species. The brown shading indicates the group of invasive species while the blue shading indicates the group of native species.

Figure 5.
The mean and standard error of biomass, diameter, and relative growth rate (RGR) of each species in (a,c,e) mature and (b,d,f) young forests calculated from the observed data at each distance. The brown shading indicates the group of invasive species while the blue shading indicates the group of native species. Points are slightly jittered to reduce overlap at each distance.
Figure 5.
The mean and standard error of biomass, diameter, and relative growth rate (RGR) of each species in (a,c,e) mature and (b,d,f) young forests calculated from the observed data at each distance. The brown shading indicates the group of invasive species while the blue shading indicates the group of native species. Points are slightly jittered to reduce overlap at each distance.

Figure 6.
Coefficient estimates ±95% confidence intervals of scaled coefficients for distance, forest successional age, percent available light, and herbivory across all species from mixed-effects regression models fit to (a) biomass, (b) diameter, and (c) relative growth rate of height (RGR).
Figure 6.
Coefficient estimates ±95% confidence intervals of scaled coefficients for distance, forest successional age, percent available light, and herbivory across all species from mixed-effects regression models fit to (a) biomass, (b) diameter, and (c) relative growth rate of height (RGR).

Figure 7.
Results from linear mixed effects models of growth measurements. (a,c,e) Differences between the overall model intercept and the estimated intercept for each species ±95% confidence intervals, and (b,d,f) the marginal effect of distance on three of the performance responses, biomass, diameter, and relative growth rate. The brown shading indicates the group of invasive species while the blue shading indicates the group of native species.
Figure 7.
Results from linear mixed effects models of growth measurements. (a,c,e) Differences between the overall model intercept and the estimated intercept for each species ±95% confidence intervals, and (b,d,f) the marginal effect of distance on three of the performance responses, biomass, diameter, and relative growth rate. The brown shading indicates the group of invasive species while the blue shading indicates the group of native species.


Figure 8.
Random-slope-intercept plots for the model estimated marginal between average herbivory incidence and (a) biomass or (b) diameter of each species. The brown shading indicates the group of invasive species while the blue shading indicates the group of native species.
Figure 8.
Random-slope-intercept plots for the model estimated marginal between average herbivory incidence and (a) biomass or (b) diameter of each species. The brown shading indicates the group of invasive species while the blue shading indicates the group of native species.

© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Dillon, W.W.; Lieurance, D.; Hiatt, D.T.; Clay, K.; Flory, S.L. Native and Invasive Woody Species Differentially Respond to Forest Edges and Forest Successional Age. Forests 2018, 9, 381. https://doi.org/10.3390/f9070381
AMA Style
Dillon WW, Lieurance D, Hiatt DT, Clay K, Flory SL. Native and Invasive Woody Species Differentially Respond to Forest Edges and Forest Successional Age. Forests. 2018; 9(7):381. https://doi.org/10.3390/f9070381
Chicago/Turabian StyleDillon, Whalen W., Deah Lieurance, Drew T. Hiatt, Keith Clay, and S. Luke Flory. 2018. "Native and Invasive Woody Species Differentially Respond to Forest Edges and Forest Successional Age" Forests 9, no. 7: 381. https://doi.org/10.3390/f9070381
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.