Growth Variations of Tree Saplings in Relation to Species Diversity and Functional Traits in a Tree Diversity Pot Experiment


1
Faculty of Life Science and Technology, Central South University of Forestry and Technology, Changsha 410004, China
2
National Engineering Laboratory of Applied Technology for Forestry and Ecology in Southern China, Changsha 410004, China
3
Huitong National Field Station for Scientific Observation and Research of Chinese Fir Plantation Ecosystem in Hunan Province, Huitong 438107, China
*
Author to whom correspondence should be addressed.
Forests 2018, 9(7), 380; https://doi.org/10.3390/f9070380
Submission received: 23 April 2018
/
Revised: 11 June 2018
/
Accepted: 22 June 2018
/
Published: 26 June 2018
(This article belongs to the Special Issue Effects of Forest Management Practices on Forest Biodiversity)
Abstract
:Despite the accumulating evidence of the beneficial effects of diverse mixed species forests on ecosystem functioning and services, foresters in subtropical forest cultivation in China still prefer easily managed monocultures, which is also due to the complexity of mixed forests and the unknown underlying mechanisms related to relationships between biodiversity and forest growth. In a designed pot experiment, we selected two early-successional tree species (Pinus massoniana Lamb., Liquidambar formosana Hance.) and two late-successional tree species (Schima superba Champ., Elaeocarpus decipiens Hemsl.) and planted four saplings in one pot with regard to tree species diversity (monoculture, two species and four species mixtures), each combination replicated four times. In this three-year duration experiment, the effect of tree species diversity, tree identity, and functional traits on sapling growth (tree height, ground diameter, crown projection area), were analyzed. The results showed that the increments of ground diameter and crown projection area increased with tree species richness, whereas the mean tree height increment showed the opposite effect. This growth variation was species specific and related to functional traits (early or late succession), as the increments of the early successional tree species (P. massoniana Lamb. and L. formosana Hance.) had a positive correlation with tree species richness, while the late successional tree species (E. decipiens Hemsl. and S. superba Champ.) showed negative effects. In addition, our study provided evidence for the allometric differences between mixtures and monocultures, which have an important reference value on mixed-species forests.
Keywords:
species richness; complementarity; growth; successional stage; species identity; allometry1. Introduction
Biodiversity is an ancient issue, even derived earlier than ecology. Darwin [1] pointed out that grass in a mixture will gain more hay than monoculture. Over the next one hundred years, ecologists devoted themselves to the research of the relationships between diversity and productivity. Among them, many studies were conducted on grass ecosystems [2,3,4,5], and most of the results revealed over-yielding (i.e., a higher biomass or production in a mixture than in the corresponding monocultures), although some of those showed a negative or no correlation relationship. Over the past two decades, people have started to realize the importance of the forest ecosystem and services, for example the most important carbon sequestration and carbon sink in the terrestrial ecosystems [6], and more and more ecologists have begun to study the effects of biodiversity on ecosystem functioning (BEF) in the forest ecosystem. Many manipulated biodiversity experimental sites have been established and evidence of positive BEF is accumulating. However, people still adopt monoculture in plantation cultivation [7].
Compared to the boreal, temperate, and tropical forests where positive relationship between species richness and biomass (productivity) has been documented in many studies [8,9,10,11], the study of the biodiversity effect in the forest in the subtropical area is relatively scarce. Subtropical regions possess about a quarter of China’s territory [12], and species diversity is most abundant, while the terrain is a most complicated area with hilly mountains. It is so hard to control field conditions in this region with a range of tree species diversity and to disentangle the effects of species diversity on ecosystem services. It is not surprising that no species diversity effects on saplings growth were found in several studies carried out recently in hilly mountains or open fields in this region [7,13,14], however this sort of experiment undertaken by us in controlled conditions (e.g., pot experiment), to our knowledge, has never been reported.
Tree growth performance is often determined by tree height, stem diameter, and crown width [7]. The crown projection areas are quite sensitive to neighborhood interactions, and are strongly related to species identity (e.g., shade tolerant or not) [15,16,17]. Furthermore, the different successional trees may have different investment strategies in the growth process. For example, the early successional or mid-successional species grows faster than the late successional species at the early development stages. Thereby the early successional or mid-successional tree may invest more carbon on height growth to get more light while the late successional tree puts emphasis on diameter to accumulate nutrition. The different growth strategies help canopy stratification and increase light use efficiency in mixed communities.
Although the survival and growth of saplings is critical for the success of afforestation, few researches have emphasized plant growth in the manipulated biodiversity experiments [18]. In this study, four subtropical tree species, comprising two early successional species and two late successional species, were planted in monocultures and mixtures (two and four species) in the greenhouse and the growth status was monitored after two years. The objectives were to test two hypotheses that: (1) neighborhood species diversity enhances the overall growth of saplings; (2) the species identity and functional traits (early successional species or late successional species) respond differently to the presence of the other neighbor species in mixtures.
2. Materials and Methods
2.1. Experimental Design
The pot planting experiment was conducted in a cultivated greenhouse of the National Engineering Laboratory for Applied Technology and Forestry & Ecology in Southern China, Changsha, Hunan (112°98′ E, 28°13′ N). Round pots with inner size 30 cm in diameter and 30 cm height were filled with clay loam red soil derived from Lithocarpus glaber Thunb.-Cyclobalanopsis glauca Thunb. evergreen broadleaved forest [19]. The soils were mixed with organic manure at ratio 6:1 (soil:manure). Four species, consisting of two early successional species (Pinus massoniana Lamb., Liquidambar formosana Hance.) and two late successional species (Schima superba Champ., Elaeocarpus decipiens Hemsl.) were selected as they are commonly spotted in forests in this subtropical region. One year old saplings were bought from the same nursery company with similar height and diameter for each species. Four tree saplings were planted in a grid of 15 cm × 15 cm at the center of the pot, consisting of three species levels: one-species monocultures, two-species combinations, and four-species combinations. Therefore, there were a total of 11 species planting combinations, including 4 monocultures of each species, 6 possible two-species combinations, and 1 four-species combination (Figure 1). Each planting pattern was replicated four times, and in total there were 44 pots and 176 saplings. All the saplings were planted in March 2014. During the experiment, no further fertilization was applied and all the saplings were placed under natural light condition and watered when needed.
2.2. Measurements
The data sampling for all the saplings was conducted repeatedly three times (in November 2014, November 2015, and November 2016). Total sapling height was measured from ground to apical meristem by using linear tape and the stem diameter on the base was measured 5 cm above ground in two vertical directions with a vernier caliper while the mean values were calculated and used for analysis. The crown diameter was measured in the longest direction (a) and perpendicular direction to it (b), and the crown projection area was calculated as an ellipse with formula: π × a × b/4. The annual increment of diameter, total height, and crown expansion area were calculated for two growing seasons as: first season, value (November 2015)—value (November 2014) and second season, value (November 2016)—value (November 2015), respectively. The cumulative growth was calculated as the sums of both growth seasons. The final growth status and numbers of survival of saplings are listed in Table 1.
2.3. Data Analysis
The annual increment of tree height, ground diameter, and crown projection area at different species richness levels were analyzed for two specific growing seasons separately and the results showed a similar pattern (Figure 2). Therefore, we analyzed the data mainly based on the cumulative growth data after two growing seasons to detect the enhanced effects of species richness and functional traits on sapling growth. The cumulative increment of tree height, ground diameter, and crown projection area in different species richness were analyzed by means of one-way analysis of variance (ANOVA) with Tukey HSD to test whether there were significant differences among different tree species richness, and between different functional trait groups (α = 0.05). In order to understand the differences of growth dynamics and strategies between fast- and relatively slow-growing species, the relationships of height to the diameter were examined using general linear regressions for early successional (P. massoniana and L. formosana) and late successional tree species (Schima superba, Elaeocarpus decipiens) separately in monoculture and mixtures. Since there were only four replicates for the 4 species combinations, we grouped the 2 species combinations and the 4 species combinations as mixtures versus monocultures. Here the error data due to human factors during measurement or the dead saplings were excluded, i.e., one P. massoniana and L. formosana saplings in monoculture two Schima superba and Elaeocarpus decipiens saplings in 2-species combinations were dead during the experiment (Table 1). All the statistical analyses were performed using software R project (R 3.3.1) [20].
3. Results
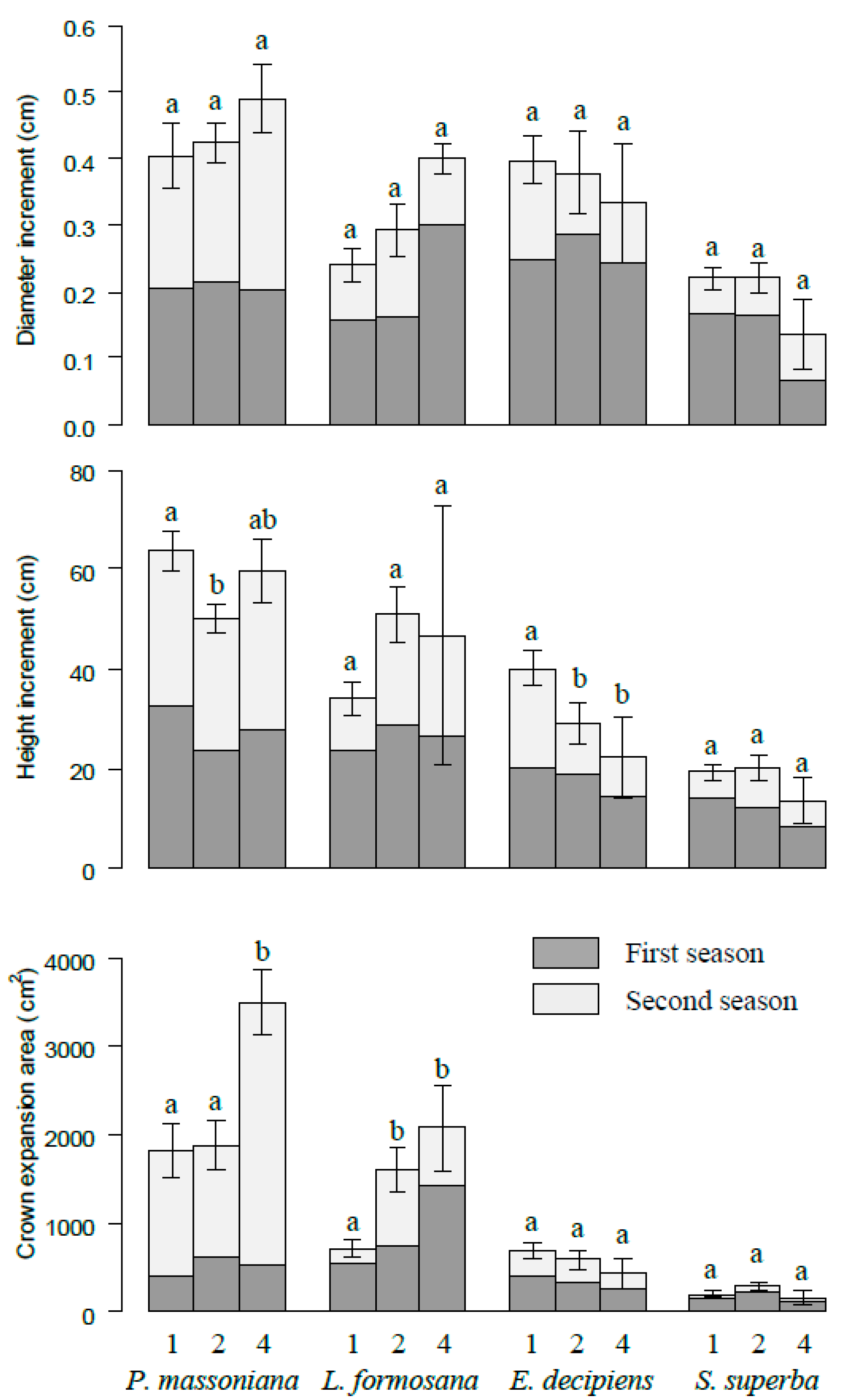
Cumulative increment of diameter increased slightly with increasing species richness at the pot level, although no significant differences were detected among different species diversity levels (Figure 2). The cumulative crown expansion area was significantly higher in four species combinations (6166.39 cm2) than that in monocultures (3438.58 cm2). In contrast, the cumulative height increment decreased with species diversity. These patterns were consistent in two growth periods here. On the species level, P. massoniana grew fastest, and the increments of diameter, height, and crown projection area on average were 0.42 cm, 56.01 cm, and 2007.53 cm2, respectively. L. formosana was the second fastest growth species, and S. superba grew slowest. The fastest growth species (P. massonian and L. formosana) increased with species diversity, while the slowest growth species (S. superba and E. decipiens) decreased with species diversity in terms of diameter increment and crown expansion area (Figure 3). The height increment showed a similar pattern except for slight fluctuation of P. massonian and L. formosana.
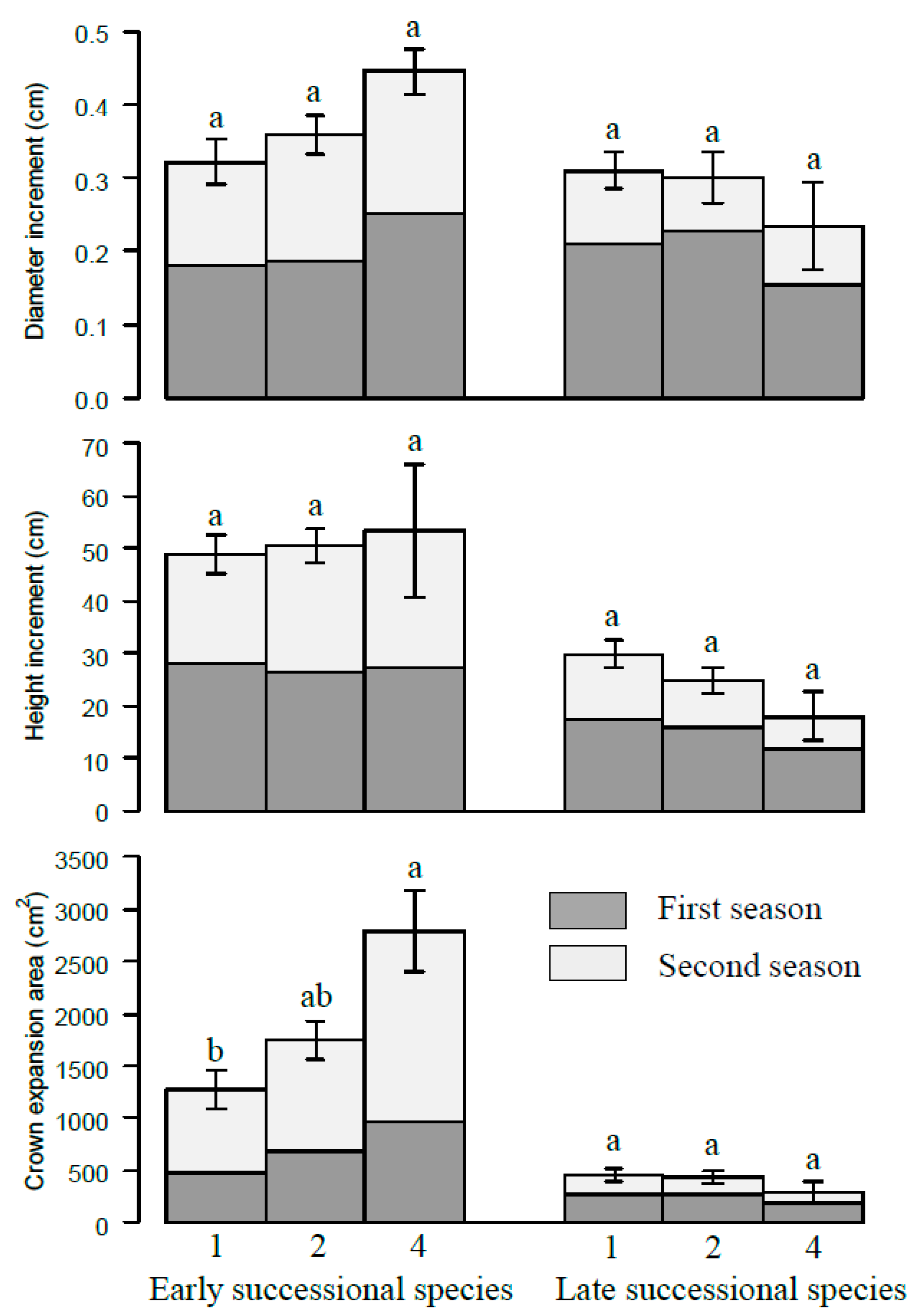
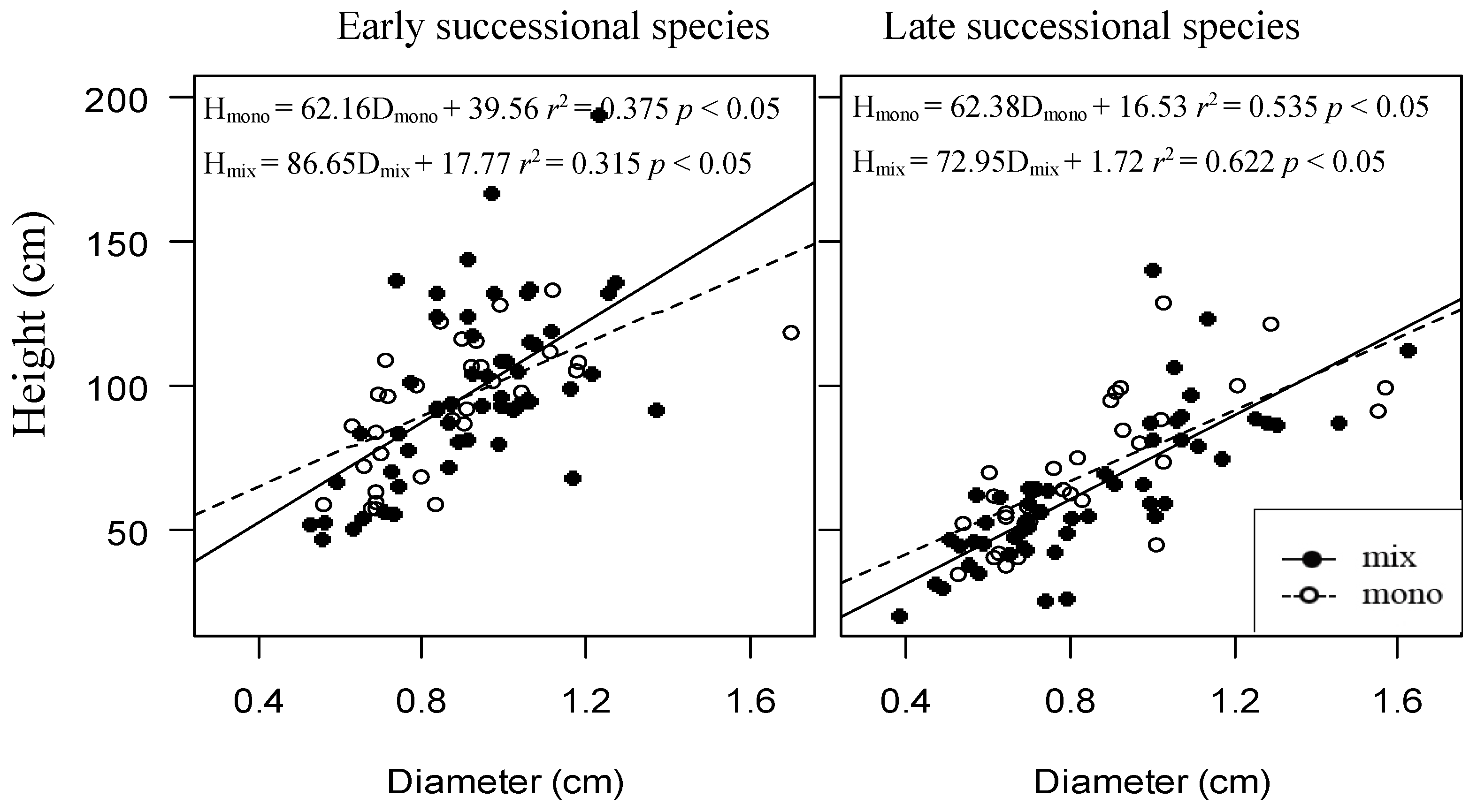
When these four species were divided into two functional groups according to the successional traits, early and late successional groups, the early successional groups (P. massoniana and L. formosana) exhibited higher growth status in more diverse combinations compared to those in less diverse combinations or monocultures in terms of increment of diameter, height, and crown projection area. Conversely, the late successional grouped species (S. superba and E. decipiens) decreased with species diversity (Figure 4). It seems that the early successional species benefited in mixtures at the expense of the late successional species’ suppressed growth in mixtures. Furthermore, the height–diameter relationships varied for the different species and neighboring species presented. The height to diameter ratio is higher in the mixture than in monocultures at the end of this pot experiment and the gap is larger for the early successional grouped species groups (Figure 5).
4. Discussion
4.1. The Relationship between Tree Growth and Species Richness
In this study, we did not find a consistent pattern between tree growth and species richness (p > 0.05). The increments of diameter and crown projection area showed a positive correlation with the species richness but the height increment showed the opposite trend. Our findings here partly support the previous demonstration that a greater diversity of neighborhood species moderately enhanced growth, and the neighborhood tree species identity also matters [21], but no significant positive correlation with the species richness. In the mixtures, the co-existing species interact with each other and facilitation is always together with competition. When complementary resource use and facilitation is bigger than competition, tree growth will be positively related to diversity and vice versa. Due to the tree species identity in mixtures, the four tree species showed higher positive effects on crown dimensions (coniferous, deciduous, and evergreen broad-leaved tree), and radial growth. In addition, what is worth mentioning is that the sensitivity of the crown projection area is bigger than height growth and diameter increment [22]. Quite a few studies have clearly found the prominent effects of species richness on sapling growth. The study of von Oheimb et al. [14] showed that a very weak or not significant effect of species richness on radial growth in subtropical forest stands. Pretzsch and Schütze [23] found a positive effect of local neighborhood mixing on biomass production in temperate forest stands of beech and Norway spruce. This disparity is likely due to the heterogeneous environmental and edaphic conditions in the open field. Here our study was conducted in relatively homogeneous conditions in the greenhouse, where it was possible to control some biotic or abiotic variables, on the other hand, planting saplings in the pot may hinder root proliferation and volume in this limiting pot size [24]. Meanwhile it could strengthen intraspecific or interspecific competition and enhance the mixing effects on the saplings growth under different species richness levels [23,25]. Initially, we designed the pot experiment only for two growing seasons, but the plant size was still relatively small. In order to detect the diversity effect and enhanced interaction between individuals, we prolonged the experiment for another growing season. The growth increment of diameter, height, and crown projection area at the second investigated season were equal or even higher than the first season. Thus we think the limiting pot size is acceptable. However, saplings may be very sensitive to species richness or not. We did not know to what extend the effects of species diversity for saplings were positive [26], negative [27] or had no relationship [28]. Maybe the different tree combinations in the diverse regions cause the variation in results in the assorted studies [29].
4.2. Species-Specific Growth Variation
The early successional or pioneer tree species usually grow fast while the late successional or shade-tolerant trees are slow growing, especially at the sapling stage. Our result showed the increment of early successional species (P. massoniana and L. formosana) had a positive correlation with the species richness but it was otherwise for late successional trees (S. superba and E. decipiens). In other studies, S. superba [30] and L. formosana showed high growth rates [7] but Q. serrata showed very slow-growth [31]. In the natural ecosystem succession, the late successional tree will replace the early successional tree, so there are different living strategies for them. In order to get more light in the limited time the early successional saplings have height growth priority, but the late successional saplings focus on diameter growth like spur with long accumulation.
The ratio of height to diameter (H/D) directly reflects the way of allometry. Our results showed that the H/D were different not only in mixture and monoculture but also in different successional trees (Figure 5). In monoculture, the regression equations of early and late successional trees were similar at this sapling stage. However, in mixtures, the height to diameter ratios of early successional species were bigger than for late successional species, suggesting the different functional trees responded differently to the presence of the foreign neighbor tree species and then changed the allometry [32]. It could lead to bias when simply extrapolating the allometric relationship from monocultures to mixed forests [33]. H/D can also indicate the competition for light or for below-ground resources [34], which indicates that the diverse communities may change the limiting factor for saplings growth. This can partially explain why the increment of diameter and crown projection area had a positive correlation with the species richness onwards (Figure 2). Therefore, we concluded that there were tradeoffs between vertical space occupation and radial growth, indicating the growth strategies used by different tree species were determined by the species identity or the growth traits in different tree combinations. Coincidentally, the different tree combinations change the limiting factors of tree growth. Although pot size might have constrained rooting growth in this experiment, our results revealed a consistent pattern based on the annual growth and cumulative growth data and showed agreement with previous findings reported in natural forests in terms of allometry in European forests [32]. However, it still requires extended research to further identify this.
Author Contributions
P.L. and W.X. conceived and designed the experiments; J.Y., X.H. and S.O. performed the experiments; J.Y. and P.L. analyzed the data; W.X. and S.O. contributed technical advice. J.Y and P.L. wrote the paper.
Funding
This study was supported by the National Natural Science Foundation of China (31670448) and Young Talent Introduction Program of CSUFT (Central South University of Forestry and Technology).
Acknowledgments
We thank the Yan Liu and Xiaoxiao Shen for assistance in data collection in the greenhouse work. We are also grateful for two anonymous reviewers’ helpful comments.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Darwin, C. The Origin of Species by Means of Natural Selection; John Murray: London, UK, 1859. [Google Scholar]
- Naeem, S.; Thompson, L.J.; Lawler, S.P.; Woodfin, R.M. Declining biodiversity can alter the performance of ecosystems. Nature 1994, 368, 734–737. [Google Scholar] [CrossRef]
- Tilman, D.; Wedin, D.; Knops, J. Productivity and sustainability influenced by biodiversity in grassland ecosystems. Nature 1996, 379, 718–720. [Google Scholar] [CrossRef]
- Hooper, D.U.; Vitousek, P.M. The effects of plant composition and diversity on ecosystem processes. Science 1997, 277, 1302–1305. [Google Scholar] [CrossRef]
- Hector, A.; Schmid, B.; Beierkuhnlein, C.; Caldeira, M.; Diemer, M.; Dimitrakopoulos, P.G.; Finn, J.A.; Freitas, H.; Giller, P.S.; Good, J.; et al. Plant diversity and productivity experiments in European grasslands. Science 1999, 286, 1123–1127. [Google Scholar] [CrossRef] [PubMed]
- Kyoto Protocol to the United Nations Framework Convention on Climate Change. Available online: https://unfccc.int/resource/docs/convkp/kpeng.pdf (accessed on 20 April 2018).
- Li, Y.; Härdtle, W.; Bruelheide, H.; Nadrowski, K.; Scholten, T.; Wehrden, H.V.; Oheimb, G.V. Site and neighborhood effects on growth of tree saplings in subtropical plantations (China). For. Ecol. Manag. 2014, 327, 118–127. [Google Scholar] [CrossRef]
- Vilà, M.; Carrillo-Gavilán, A.; Vayreda, J.; Bugmann, H.; Fridman, J.; Grodzki, W.; Haase, J.; Kunstler, G.; Schelhaas, M.J.; Trasobares, A. Disentangling biodiversity and climatic determinants of wood production. PLoS ONE 2007, 8, e53530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paquette, A.; Messier, C. The effect of biodiversity on tree productivity: From temperate to boreal forests. Glob. Ecol. Biogeogr. 2011, 20, 170–180. [Google Scholar] [CrossRef] [Green Version]
- Gamfeldt, L.; Snäll, T.; Bagchi, R.; Jonsson, M.; Gustafsson, L.; Kjellander, P.; Ruiz-Jaen, M.C.; Fröberg, M.; Stendahl, J.; Philipson, C.D.; et al. Higher levels of multiple ecosystem services are found in forests with more tree species. Nat. Commun. 2013, 4, 1340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vilà, M.; Vayreda, J.; Comas, L.; Ibáñez, J.J.; Mata, T.; Obón, B. Species richness and wood production: A positive association in Mediterranean forests. Ecol. Lett. 2007, 10, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.H.; Kent, M.; Fang, X.F. Evergreen broad-leaved forest in Eastern China: Its ecology and conservation and the importance of resprouting in forest restoration. For. Ecol. Manag. 2007, 245, 76–87. [Google Scholar] [CrossRef]
- Lang, A.C.; Härdtle, W.; Baruffol, M.; Böhnke, M.; Bruelheide, H.; Schmid, B.; Von-Wehrden, H.; von Oheimb, G. Mechanisms promoting tree species co-existence: Experimental evidence with saplings of subtropical forest ecosystems of China. J. Veg. Sci. 2012, 23, 837–846. [Google Scholar] [CrossRef] [Green Version]
- Von Oheimb, G.; Lang, A.C.; Bruelheide, H.; Forrester, D.I.; Wäsche, I.; Yu, M.; Härdtle, W. Individual-tree radial growth in a subtropical broad-leaved forest: The role of local neighborhood competition. For. Ecol. Manag. 2011, 261, 499–507. [Google Scholar] [CrossRef]
- Lang, A.C.; Härdtle, W.; Bruelheide, H.; Geißler, C.; Nadrowski, K.; Schuldt, A.; Yu, M.J.; Von-Oheimb, G. Tree morphology responds to neighborhood competition and slope in species-rich forests of subtropical China. For. Ecol. Manag. 2010, 260, 1708–1715. [Google Scholar] [CrossRef]
- Longuetaud, F.; Seifert, T.; Leban, J.M.; Pretzsch, H. Analysis of long-term dynamics of crowns of sessile oaks at the stand level by means of spatial statistics. For. Ecol. Manag. 2008, 255, 2007–2019. [Google Scholar] [CrossRef]
- Longuetaud, F.; Piboule, A.; Wernsdorfer, H.; Collet, C. Crown plasticity reduces inter-tree competition in a mixed broadleaved forest. Eur. J. For. Res. 2013, 132, 621–634. [Google Scholar] [CrossRef]
- He, Y.J.; Liang, X.Y.; Qin, L.; Li, Z.Y.; Shao, M.X.; Tan, L. Community characteristics and soil properties of coniferous plantation forest monocultures in the early stages after close-to-nature transformation management in southern subtropical China. Acta Ecol. Sin. 2013, 33, 2484–2495. [Google Scholar]
- Liu, C.; Xiang, W.H.; Lei, P.F.; Deng, X.W.; Tian, D.L.; Fang, X.; Peng, C.H. Standing fine root mass and production in four Chinese subtropical forests along a succession and species diversity gradient. Plant Soil 2014, 376, 445–459. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Ratcliffe, S.; Holzwarth, F.; Nadrowski, K.; Levick, S.; Wirth, C. Tree neighbourhood matters—Tree species composition drives diversity–productivity patterns in a near-natural beech forest. For. Ecol. Manag. 2015, 335, 225–234. [Google Scholar] [CrossRef]
- Simard, S.W.; Zimonick, B.J. Neighborhood size effects on mortality, growth and crown morphology of paper birch. For. Ecol. Manag. 2005, 214, 251–265. [Google Scholar] [CrossRef]
- Pretzsch, H.; Schütze, G. Transgressive overyielding in mixed compared with pure stands of Norway spruce and European beech in Central Europe: Evidence on stand level and explanation on individual tree level. Eur. J. For. Res. 2009, 128, 183–204. [Google Scholar] [CrossRef]
- Poorter, H.; Bühler, J.; van Dusschoten, D.; Climent, J.; Postma, J.A. Pot size matters: A meta-analysis of the effects of rooting volume on plant growth. Funct. Plant Biol. 2012, 39, 839–850. [Google Scholar] [CrossRef]
- Potvin, C.; Gotelli, N.J. Biodiversity enhances individual performance but does not affect survivorship in tropical trees. Ecol. Lett. 2008, 11, 217–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-Benito, P.; Gómez-Aparicio, L.; Paquette, A.; Messier, C.; Kattge, J.; Zavala, M.A. Diversity increases carbon storage and tree productivity in Spanish forests. Glob. Ecol. Biogeogr. 2014, 23, 311–322. [Google Scholar] [CrossRef]
- Jacob, M.; Leuschner, C.; Thomas, F.M. Productivity of temperate broad-leaved forest stands differing in tree species diversity. Ann. For. Sci. 2010, 67, 503. [Google Scholar] [CrossRef]
- Nguyen, H.; Herbohn, J.; Firn, J.; Lamb, D. Biodiversity-productivity relationships in small-scale mixed-species plantations using native species in Leyte province, Philippines. For. Ecol. Manag. 2012, 274, 81–90. [Google Scholar] [CrossRef]
- Bruelheide, H.; Nadrowski, K.; Assmann, T.; Bauhus, J.; Both, S.; Buscot, F.; Chen, X.Y.; Ding, B.Y.; Durka, W.; Erfmeier, A.; et al. Designing forest biodiversity experiments: General considerations illustrated by a new large experiment in subtropical China. Methods Ecol. Evol. 2014, 5, 74–89. [Google Scholar] [CrossRef]
- Chen, Z.H.; Wang, B.S.; Zhang, H.D. Growth of the trees and saplings in the lower subtropical evergreen broad-leaved forest in Heishiding, Guangdon Province. Acta Phytoecol. Sin. 1999, 23, 441–450. [Google Scholar]
- Xing, P.; Zhang, Q.B.; Baker, P.J. Age and radial growth pattern of four tree species in a subtropical forest of China. Trees 2012, 26, 283–290. [Google Scholar] [CrossRef]
- Forrester, D.I.; Benneter, A.; Bouriaud, O.; Bauhus, J. Diversity and competition influence tree allometric relationships: Developing functions for mixed-species forests. J. Ecol. 2017, 105, 761–774. [Google Scholar] [CrossRef]
- Forrester, D.I.; Pretzsch, H. Tamm Review: On the strength of evidence when comparing ecosystem functions of mixtures with monocultures. For. Ecol. Manag. 2015, 356, 41–53. [Google Scholar] [CrossRef]
- Bauhus, J.; Khanna, P.K.; Menden, N. Aboveground and belowground interactions in mixed plantations of Eucalyptus globulus and Acacia mearnsii. Can. J. For. Res. 2000, 30, 1886–1894. [Google Scholar] [CrossRef]
Figure 1.
The planting pattern of the pot experiment. Different symbols indicate different species.
Figure 2.
Total increments of diameter, height and crown projection area at different species richness levels on the pot level. The number 1, 2, and 4 represent monoculture, two species and four species mixtures, respectively. Bars represent means ± SE. Different letters indicate significant differences among different species richness levels, p < 0.05.
Figure 2.
Total increments of diameter, height and crown projection area at different species richness levels on the pot level. The number 1, 2, and 4 represent monoculture, two species and four species mixtures, respectively. Bars represent means ± SE. Different letters indicate significant differences among different species richness levels, p < 0.05.
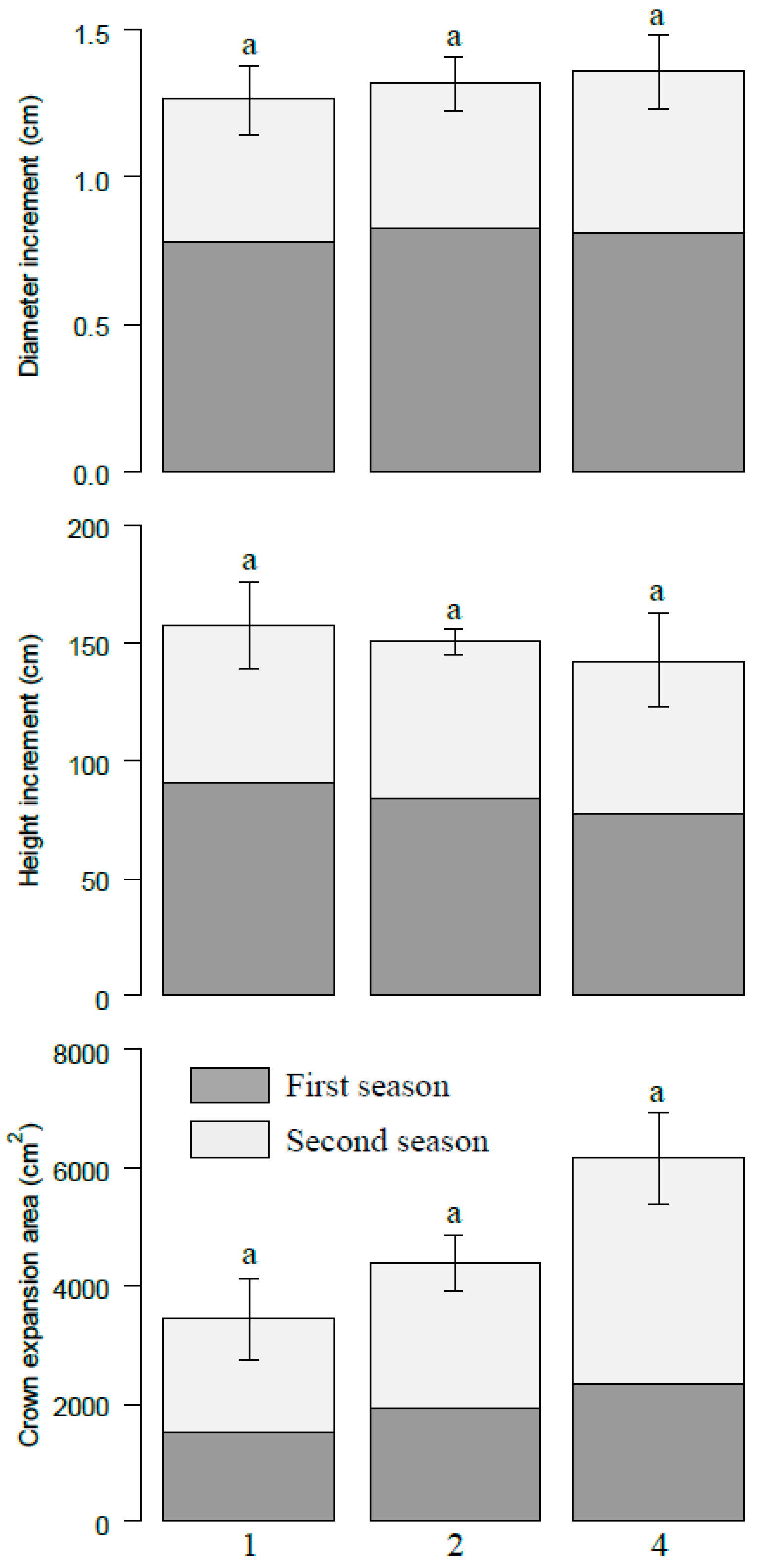
Figure 3.
The increments of diameter, height, and crown projection area for P. massoniana, L. formosana, E. decipiens, and S. superba at different species richness levels. The number 1, 2, and 4 represent monoculture, two species and four species mixtures, respectively. Bars represent means ± SE. Different letters indicate significant differences among different species richness levels, p < 0.05.
Figure 3.
The increments of diameter, height, and crown projection area for P. massoniana, L. formosana, E. decipiens, and S. superba at different species richness levels. The number 1, 2, and 4 represent monoculture, two species and four species mixtures, respectively. Bars represent means ± SE. Different letters indicate significant differences among different species richness levels, p < 0.05.
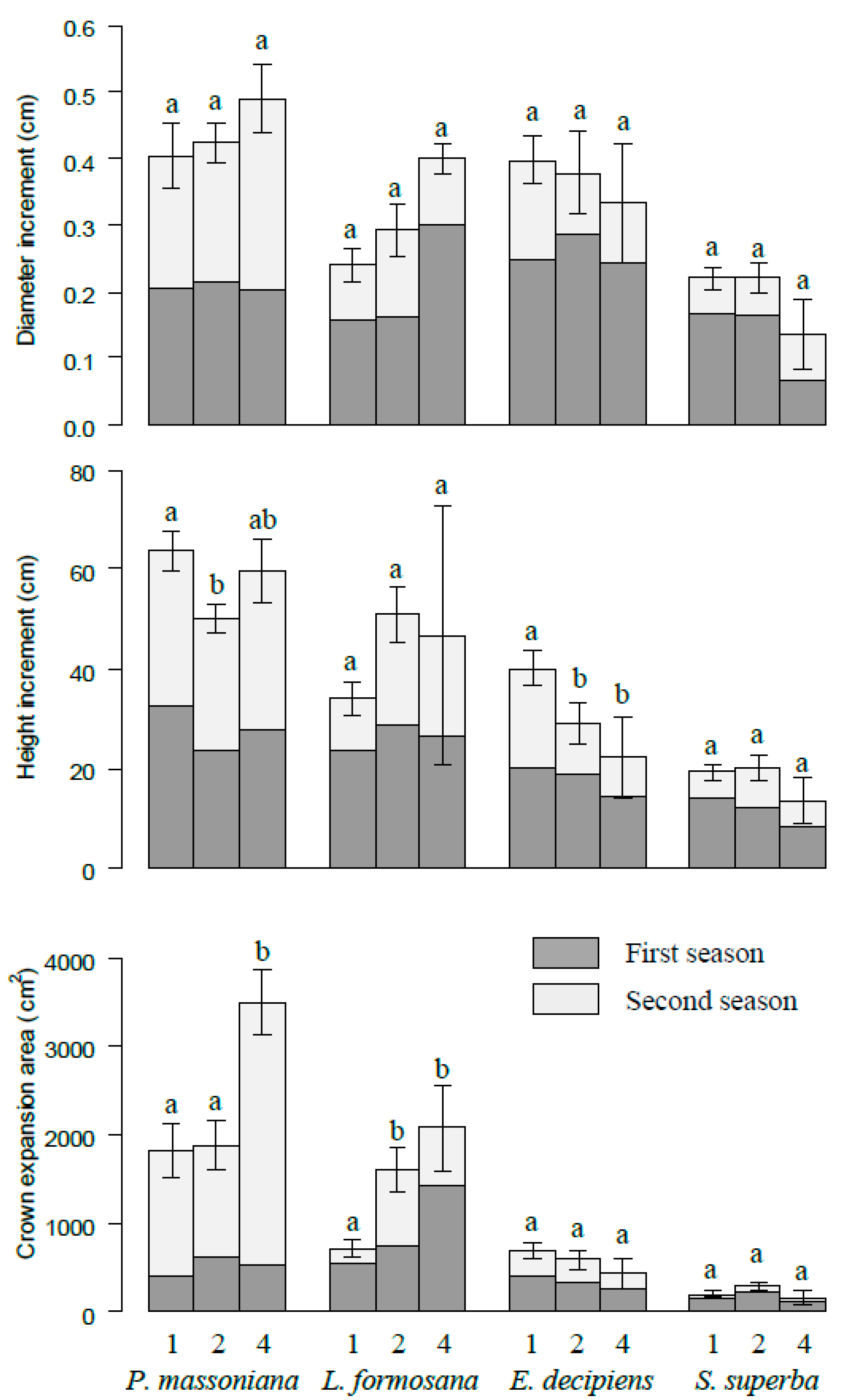
Figure 4.
The increments of diameter, height, and crown projection area of different successional species at different species richness levels. The numbers 1, 2, and 4 represent monoculture, two species and four species mixtures, respectively. Bars represent means ± SE. Different letters indicate significant differences among different species richness levels, p < 0.05.
Figure 4.
The increments of diameter, height, and crown projection area of different successional species at different species richness levels. The numbers 1, 2, and 4 represent monoculture, two species and four species mixtures, respectively. Bars represent means ± SE. Different letters indicate significant differences among different species richness levels, p < 0.05.
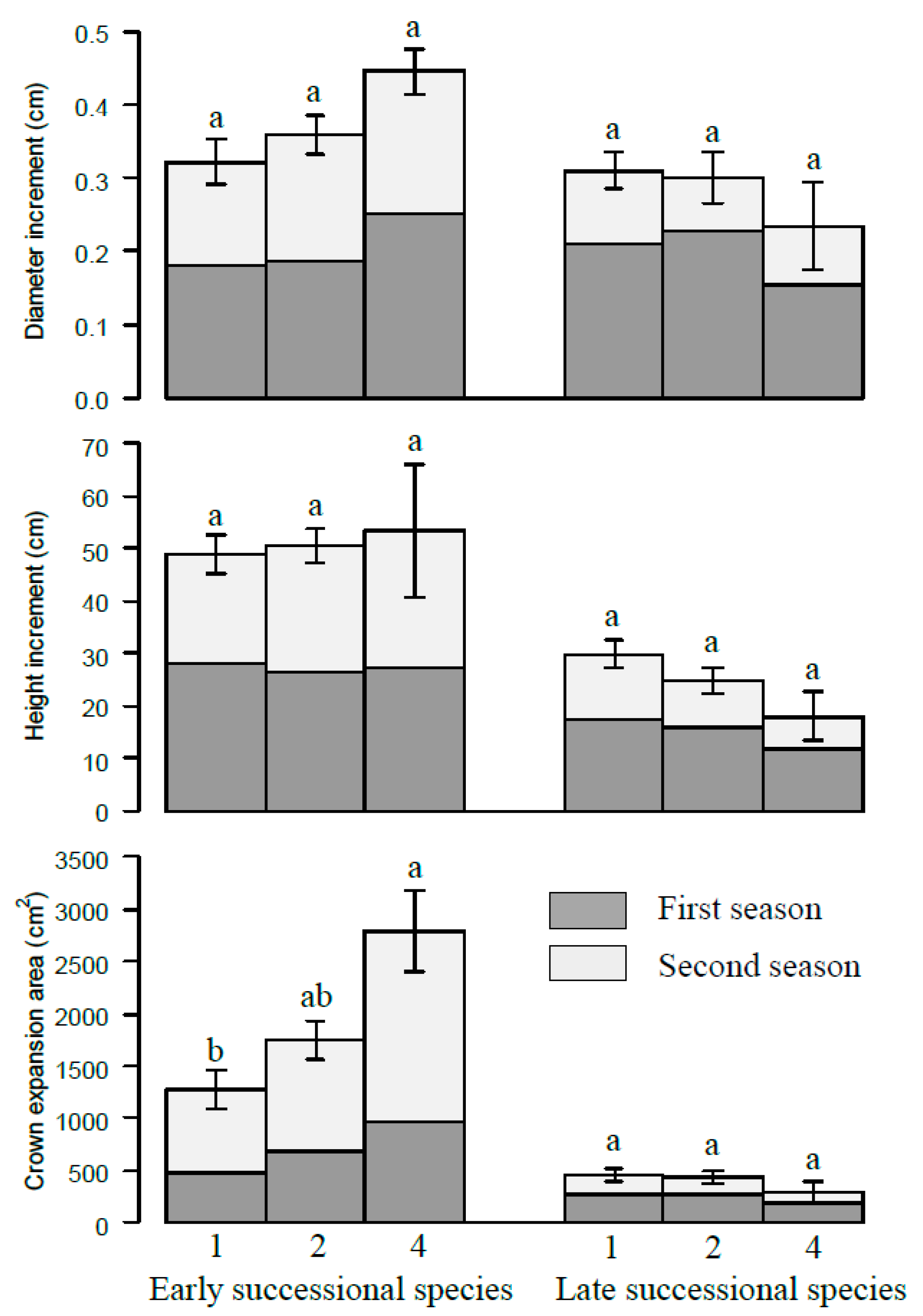
Figure 5.
The relationships between height and diameter of different successional species as monoculture and mixture. Dashed line and empty circles represent the monoculture, solid line and filled circles represent the mixture.
Figure 5.
The relationships between height and diameter of different successional species as monoculture and mixture. Dashed line and empty circles represent the monoculture, solid line and filled circles represent the mixture.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The mean diameter, height, crown projection area, and number of saplings in different species diversity levels at the end the pot experiment. Given are means ± SE (standard error).
Table 1.
The mean diameter, height, crown projection area, and number of saplings in different species diversity levels at the end the pot experiment. Given are means ± SE (standard error).
| Species | Richness | Diameter (cm) | Height (cm) | Crown Projection Area (cm2) | n |
|---|---|---|---|---|---|
| P. massoniana Lamb. | 1 | 0.88 ± 0.07 | 92.62 ± 5.28 | 2381.79 ± 359.88 | 15 |
| 2 | 0.87 ± 0.04 | 84.68 ± 3.98 | 2332.70 ± 281.20 | 24 | |
| 4 | 1.01 ± 0.09 | 100.35 ± 11.00 | 3906.15 ± 405.29 | 4 | |
| L. formosana Hance. | 1 | 0.83 ± 0.04 | 92.51 ± 6.76 | 859.08 ± 106.40 | 15 |
| 2 | 0.89 ± 0.04 | 105.84 ± 6.32 | 1879.22 ± 283.89 | 24 | |
| 4 | 1.21 ± 0.03 | 116.35 ± 30.22 | 2747.92 ± 451.11 | 4 | |
| S. superba Champ. | 1 | 0.67 ± 0.02 | 52.97 ± 2.72 | 602.94 ± 58.81 | 16 |
| 2 | 0.64 ± 0.02 | 50.54 ± 2.10 | 604.17 ± 57.49 | 22 | |
| 4 | 0.49 ± 0.06 | 27.10 ± 2.32 | 326.33 ± 26.35 | 4 | |
| E. decipiens Hemsl | 1 | 1.09 ± 0.07 | 90.37 ± 4.95 | 1101.05 ± 136.35 | 16 |
| 2 | 1.04 ± 0.05 | 75.58 ± 4.94 | 969.66 ± 183.31 | 22 | |
| 4 | 0.76 ± 0.09 | 62.88 ± 17.61 | 418.12 ± 254.01 | 4 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Yang, J.; Lei, P.; Xiang, W.; Ouyang, S.; Hui, X. Growth Variations of Tree Saplings in Relation to Species Diversity and Functional Traits in a Tree Diversity Pot Experiment. Forests 2018, 9, 380. https://doi.org/10.3390/f9070380
AMA Style
Yang J, Lei P, Xiang W, Ouyang S, Hui X. Growth Variations of Tree Saplings in Relation to Species Diversity and Functional Traits in a Tree Diversity Pot Experiment. Forests. 2018; 9(7):380. https://doi.org/10.3390/f9070380
Chicago/Turabian StyleYang, Jingjun, Pifeng Lei, Wenhua Xiang, Shuai Ouyang, and Xinrong Hui. 2018. "Growth Variations of Tree Saplings in Relation to Species Diversity and Functional Traits in a Tree Diversity Pot Experiment" Forests 9, no. 7: 380. https://doi.org/10.3390/f9070380
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.