Potential Roles of n-3 PUFAs during Skeletal Muscle Growth and Regeneration


Mary MacKillop Institute for Health Research, Exercise and Nutrition Research Program, Australian Catholic University, Melbourne, VIC 3000, Australia
*
Author to whom correspondence should be addressed.
Nutrients 2018, 10(3), 309; https://doi.org/10.3390/nu10030309
Submission received: 21 January 2018
/
Revised: 1 March 2018
/
Accepted: 2 March 2018
/
Published: 5 March 2018
(This article belongs to the Special Issue Nutrition and Ageing)
Abstract
:Omega-3 polyunsaturated fatty acids (n-3 PUFAs), which are commonly found in fish oil supplements, are known to possess anti-inflammatory properties and more recently alter skeletal muscle function. In this review, we discuss novel findings related to how n-3 PUFAs modulate molecular signaling responsible for growth and hypertrophy as well as the activity of muscle stem cells. Muscle stem cells commonly known as satellite cells, are primarily responsible for driving the skeletal muscle repair process to potentially damaging stimuli, such as mechanical stress elicited by exercise contraction. To date, there is a paucity of human investigations related to the effects of n-3 PUFAs on satellite cell content and activity. Based on current in vitro investigations, this review focuses on novel mechanisms linking n-3 PUFA’s to satellite cell activity and how they may improve muscle repair. Understanding the role of n-3 PUFAs during muscle growth and regeneration in association with exercise could lead to the development of novel supplementation strategies that increase muscle mass and strength, therefore possibly reducing the burden of muscle wasting with age.
1. Introduction
Skeletal muscle is a highly malleable tissue with the capacity to alter its phenotype in response to exercise and nutrient availability [1]. With increasing age, skeletal muscle becomes less responsive to anabolic stimuli, such as resistance exercise and protein feeding. It is thought that this reduced sensitivity to anabolic stimuli, termed ”anabolic resistance”, is implicated in the etiology of sarcopenia, which is the gradual loss of muscle mass with age [2,3,4,5,6,7]. Other factors that are known to contribute to sarcopenia include reductions in circulating sex hormones [8], physical inactivity [9], low grade inflammation [10,11], impairments to neuromuscular junctions [12], and reduced muscle stem cell (i.e., satellite cell) number and function [13]. Sarcopenia is phenotypically associated with decreased muscle fiber size and shifts in fiber type change from fast to slow, resulting in a decrease in maximal muscle force production [14,15]. These changes are also accompanied by a diminished regenerative capacity of the muscle due to a loss in the number and activity of satellite cells in type II “fast” fibers [14,16,17].
Satellite cells are muscle specific stem cells primarily responsible for the repair of muscle in response to injury [18,19,20]. Upon their activation, satellite cells enter the cell cycle, proliferate, differentiate to myoblasts and myocytes in a process termed myogenesis, and fuse to damaged muscle fibers. Myogenesis is regulated by changes in the expression of myogenic transcriptional regulatory factors (MRF) that dictate whether satellite cells are in a quiescent, activated, committed, or differentiated state [21,22] (Figure 1). Satellite cells play a role in skeletal muscle repair; however their role in muscle hypertrophy is still equivocal (for reviews see [23,24]). Some animal studies observe skeletal muscle hypertrophy occurring even in the absence of satellite cells [20,25], whereas other animal studies provide evidence to support their role during hypertrophy [26,27]. Although the extent to which satellite cells facilitate muscle hypertrophic response is still a topic of debate, predominating evidence indicates that the presence and more likely the cells’ activation and myogenic capacity are indispensable for supporting training adaptations and may be implicated in the events leading to sarcopenia. However, throughout the lifespan of satellite cell depleted mice, sarcopenia is neither exacerbated nor accelerated [28,29]. Notably, these mice are highly sedentary, and although this may be reflective of an elderly population it is still difficult to draw definitive conclusions in the context of exercise. While satellite cell depleted mice do not show signs of increased muscle loss, satellite cell depletion appears to cause a dysregulation in the surrounding muscle environment leading to increased fibrosis with ageing and a reduction in muscle quality and function. Indeed, satellite cells have been implicated in regulating extracellular matrix production during hypertrophy and regenerative processes, and therefore do play an important role in the maintenance of muscle mass with age [26,30,31].
The precise molecular mechanisms responsible for sarcopenia are yet to be elucidated, however, accumulating evidence indicates that nutritional supplements, in particular the omega 3 polyunsaturated fatty acids (n-3 PUFAs), have the potential to reduce muscle wasting and increase the functional capacity of muscle in older individuals by augmenting intracellular anabolic signalling [32,33,34]. Eicosapentaenoic acid (EPA), Docosahexaenoic acid (DHA), and α-linolenic acid (ALA) are the most commonly found n-3 PUFAs in the human diet [35]. ALA is most commonly abundant in plant foods, such as flax seed and nuts, while EPA and DHA are abundant in cold-water fish, such as salmon and tuna [36]. In addition, Docosapentaenoic acid (DPA) is an emerging, but lesser known, n-3 PUFA that possesses a similar structure to EPA and is also biochemically active [37]. Only ALA is considered truly essential to the human diet as both EPA and DHA can be synthesised in the endoplasmic reticulum in liver cells through conversion from ALA, though this conversion is limited due to enzyme availability [38,39,40].
The n-3 PUFAs EPA and DHA are well known for their anti-inflammatory properties [41,42]. They have also been reported to elicit positive effects on a wide range of other physiological processes and systems, such as visual signalling [43,44], insulin sensitivity, and glucose tolerance [45]. Accumulating evidence now indicates that n-3 PUFAs may additionally stimulate rates of muscle protein synthesis (MPS) by increasing both the intracellular activities of signalling molecules that are involved in the maintenance of skeletal muscle mass [46,47] and possibly satellite cell activity [48]. Chronic low-grade inflammation associated with ageing and other comorbidities known to increase inflammation, such as obesity, often exacerbate the effects of sarcopenia [49]. As such, the anti-inflammatory effects of n-3 PUFAs on skeletal muscle may be most beneficial to older individuals suffering from sarcopenia [32,50,51] and may complement other strategies, such as exercise and protein supplementation, to combat sarcopenia. In this review, we focus on the dose and treatment periods required for n-3 PUFAs, EPA, and DHA, to support the differentiation of satellite cells into myogenic cells (myogenesis) and elicit protective ant-inflammatory effects in vitro and in vivo. The combined effects of n-3 PUFA supplementation, exercise, and protein ingestion, and their associated mechanisms of action on muscle metabolism in both young and older individuals, will also be discussed.
2. The Beneficial Effects of n-3 PUFAs in Skeletal Muscle
2.1. Reducing Inflammation
Muscle wasting is a hallmark of various disease states that are often associated with increased levels of inflammation [52,53]. Acute transient increases in inflammation plays an important part in the early regenerative processes of muscle adaptation, particularly in response to exercise [54,55]. However, failure to effectively resolve inflammation at its onset which results in a chronic state of inflammation can be detrimental and is associated with insulin resistance, obesity, and muscle wasting [56,57,58]. Furthermore, elevated levels of inflammation during ageing may also exacerbate the effects of sarcopenia by impairing the regenerative capacity of muscle [59].
Upon their incorporation into membrane phospholipids, EPA and DHA act as substrates for the synthesis of lipid derived mediators of inflammation [60]. These mediators, termed eicosanoids, vary in their ability to mediate inflammation depending on the substrate used for their synthesis. For example, eicosanoids derived from arachidonic acid, an omega-6 fatty acid, are considered to be pro-inflammatory [61]. Conversely, eicosanoids derived from n-3 PUFAs are considered to be less potent, reducing the intensity and duration of the inflammatory responses by immune cells [62]. This divergent behaviour is due to the fact that eicosanoids derived from n-3 PUFAs are less biologically active than n-6 PUFAs, and possess a weaker affinity for eicosanoid receptors [63,64,65]. EPA and DHA can also inhibit the endogenous production of pro-inflammatory arachidonic acids by competing as substrates for the enzymes during eicosanoid synthesis [66,67].
Inflammatory cytokines produced by immune cells post-exercise can have a profound effect on skeletal muscle protein turnover and myogenesis. In animal and cell models, cytokines, such as tumour necrosis factor-α (TNF-α) and interleukin-6 (IL-6), bind to receptors on the muscle, activating the transcription factor nuclear factor kappa-light-chain-enhancer of activated B cells (NFκB) [68,69]. Activation of NFκB subsequently increases the expression of the atrogenes muscle RING-finger protein-1 (MuRF1) and atrogin-1, promoting muscle wasting [68]. n-3 PUFAs, in addition to modulating eicosanoid synthesis, can also inhibit the activation of NFκB in skeletal muscle cells and other cell types in vitro (see Section 2.3) [69,70,71,72]. Whether n-3 PUFAs can inhibit the activation of NFκB in human skeletal muscle in response to inflammation remains to be determined. Although being considered pro-inflammatory, TNF-α and IL-6 also appear to play important role in the early stages of muscle regeneration following injury by enhancing myoblast proliferation and inhibiting differentiation [73,74,75]. This inhibition involves the down regulation of the MRFs, MyoD and Myogenin [73,74,75], with TNF-α receptor double knock out mice [74] and with IL-6 non-specific knock out mice [73] shown to present impaired myogenic differentiation and hypertrophy when compared to wild type mice during functional overloading. This highlights the importance of inflammatory processes during muscle regeneration following injury (i.e., exercise).
On the other hand, chronic inflammation observed during ageing is associated with an increase in pro-inflammatory cytokines (compared to younger individuals) such as TNF-α and IL-6 due to possible dysregulation in immune cell function [59,76]. Elevations in circulating pro-inflammatory cytokines reported in older individuals negatively impacts muscle regeneration by impairing satellite cell differentiation and fusion and by increasing NF-κB activation. These factors contribute to deterioration of muscle mass with age [59,77]. Despite their purported anti-inflammatory effects, which are thought to underlie the beneficial effects of n-3 PUFAs in muscle, Da Boit, et al. [78] and others [33,47], were unable to detect any differences in plasma TNF-α or IL-6 between the placebo and n-3 PUFA treatment groups post-intervention in both older and younger men and women. A possible explanation for this discrepancy is that the participants were healthy and had low levels of inflammation to begin with, making it difficult to detect any changes.
Mechanisms Underlying the Anti-Inflammatory Properties of n-3 PUFAs during Myogenesis
The potential positive effects of n-3 PUFA’s on myogenesis may relate to their capacity to alter the cell membrane lipid composition, which in turn, changes the profile of membrane bound proteins of lipid rafts [79,80]. These changes impact membrane fluidity and assist in myoblast fusion during myotube formation [81], allowing for n-3 PUFAs to simultaneously modulate several signalling pathways. One such pathway involves the Peroxisome proliferator-activated receptors (PPARs), which are a group of nuclear receptors that are well characterised in preventing metabolic disorders, promoting adaptations to skeletal muscle following fasting and physical exercise, and have a novel role in regulating satellite cell activity [82,83,84]. PPARs are also critical regulators of genes that are involved in development, metabolism of lipids and carbohydrates, as well as inflammation [85]. Recent evidence suggests that lipids, such as n-3 PUFAs, are capable of binding to PPARs inducing their activation and altering the expression of pro-inflammatory genes [86]. Results from several studies support the potential for n-3 PUFAs to regulate inflammatory events in muscle cells in association with the PPAR signalling pathway. TNF-α has been shown to elicit negative effects on differentiating myoblasts, resulting in decreases in myotube size and number [87]. However, pre-treating or co-treating myoblasts with EPA prevents the cytotoxic effects of TNF-α [87]. These protective effects of EPA are associated with increases in PPARγ expression and decreases in NF-κB [88]. Moreover, treating C2C12 myotubes with high concentrations (400–600 µM) of EPA or DHA has also been shown to increase the gene expression of PPARγ and reduce muscle breakdown (Huang et al., 2011; Wang et al., 2013). These findings collectively indicate that n-3 PUFAs are able to increase the expression of PPAR and reduce the expression of NF-κB in skeletal muscle resulting in reduced muscle wasting [69,72].
PPARδ is another isoform of PPAR abundantly expressed in skeletal muscle that has a novel role in regulating satellite cell activity [89]. In response to injury PPARδ-KO mice when compared to wild type mice, display reductions in proliferating satellite cell number and increases in differentiating cell number [89]. Whether n-3 PUFA supplementation is able to augment satellite cell regeneration in a PPARδ regulated manner following injury or exercise remains to be determined but is likely considering that n-3 PUFAs are known PPARs agonists in skeletal muscle [69].
Like PPARs, the peroxisome proliferator-activated receptor gamma co-activator 1-alpha (PGC-1α) is also capable of repressing the transcriptional activity of NF-kB lowering inflammation in muscle cells [90]. In differentiated C2C12 myotubes, n-3 PUFAs have been found to increase the expression of PGC-1α in a dose and time dependent manner with 50 μM for 24 h having the greatest effect [91]. PGC-1α is also indirectly involved in regulating the expression of mitochondrial DNA (mtDNA) by increasing the transcription of mitochondrial transcription factor A (Tfam) and nuclear respiratory factor 1 (NRF1) [92]. In fully formed myotubes, the expression of both Tfam and NRF1 have been shown to increase following both EPA and DHA treatment [91]. Moreover, overexpressing PGC-1α in C2C12 cells has been shown to promote differentiation and increase the expression of MyoD and Myogenin, however, it remains to be seen whether n-3 PUFAs can elicit similar responses in both PGC-1α and MRF expression during differentiation [93].
As fatty acid oxidation occurs within mitochondria, the increases in mitochondrial biogenesis seen following n-3 PUFA treatment could be compensatory as a means of disposing of excess fatty acids more efficiently, which requires more mitochondria [94]. Indeed, EPA and DHA ameliorate the lipotoxic effects of other fatty acids such as palmitate in fully formed myotubes and increase the expression that is associated with mitochondrial-β oxidation, such as carnitine palmitol transferase 1α and β (CPT1α; CPT1β) [95]. Likewise, others have shown that DHA treatment of C2C12 myoblasts attenuates the inhibitory effects of palmitate on PGC-1α activity and preserves oxidative capacity by maintaining citrate synthase activity [96]. Thus, PGC-1α appears to play an important role overcoming the deleterious effects of palmitate lipotoxcity, though it is remains to be determined whether DHA or EPA can modulate the expression of PGC-1α during myoblast differentiation. In Vitro, both PPAR and PGC-1α are capable of attenuating inflammation via downregulating NF-kB following exposure to n-3 PUFAs during myogenic differentiation in C2C12 cells. More work is required to elucidate whether n-3 PUFAs can modulate the same pathways in human satellite cells.
In summary, the n-3 PUFAs EPA and DHA can reduce inflammation via their incorporation into membrane phospholipids, where they inhibit the production of pro-inflammatory eicosanoids, reducing of activation of immune cells and the associated release of pro-inflammatory cytokines [71,97,98]. Therefore, n-3 PUFAs may assist in muscle regeneration in older individuals by reducing excessive inflammation and promoting a systemic environment more conducive to growth [99] (see Section 2.3). The anti-inflammatory properties of n-3 PUFAs and their incorporation into membrane phospholipids may also underpin some of their other beneficial effects in muscle, such as altering protein metabolism, increasing muscle strength and modulating myogenesis.
2.2. Protein Metabolism Regulation
The combination of resistance exercise and protein ingestion is known to maximally stimulate rates of muscle protein synthesis (MPS) [100,101,102] and also lead to increases in satellite cell activity [103] and content in younger men [104]. Moreover, n-3 PUFA supplementation has been shown to exert a positive effect on rates of MPS in both young and old individuals. For instance, when combined with amino acids, n-3 PUFAs enhance rates of MPS in both young and old people to a greater extent than amino acid ingestion alone [33,47]. However, it should be noted that the findings were under non-physiological conditions of a hyperaminoacidemic-hyperinsulinemic clamp [33,47]. Nonetheless, these findings provide evidence that n-3 PUFAs may be an effective strategy to rescue anabolic sensitivity in healthy older individuals purported to be ‘anabolic resistance’, as observed by attenuated MPS responses when compared to healthy young individuals in response to protein ingestion [3,7] and exercise [105].
When acute resistance exercise, protein ingestion and n-3 PUFA supplementation (5 g/day for eight weeks) are combined under physiological conditions; rates of MPS are not further enhanced compared to a placebo group in younger adults [46]. Interestingly, the activity of Akt and p70S6K1, signalling kinases involved in translation initiation in muscle, is attenuated following resistance exercise and n-3 PUFA supplementation. This could indicate that anabolic kinase efficiency is increased, suggesting less protein is required to elicit post translational modifications (e.g., phosphorylation) and that these posttranslational modifications occur at a faster rate with n-3 PUFA supplementation. This may be due to the increased incorporation of unsaturated fatty acids into membrane phospholipids thereby changing their composition and altering the activities of proteins that are associated with stimulating MPS tethered to cell membrane [79,80].
While it is possible that changes to protein signalling efficiency occur, a major limitation of the study conducted by McGlory et al. [46] is that they did not measure any changes in the total protein content of Akt and p70S6K1 nor did they measure any specific post-translational modifications of these proteins (i.e., altered phosphorylation at numerous sites). Measuring these parameters would confirm whether n-3 PUFAs are indeed capable of altering kinase efficiency and which post-translational modifications are altered following n-3 PUFA supplementation. Lastly, the supplementation protocol (dose and duration) used by McGlory et al. [46] should have been sufficient to induce changes in the muscle. Previously, the same group has shown that after four weeks of supplementation, levels of EPA and DHA continue to rise in muscle (~3.0% of total fatty acids in muscle at week 2 and ~5.0% of total fatty acids in muscle at week 4), whereas levels of EPA and DHA in blood plateau at two weeks with no further increase at four weeks (~8.0% at week 2 and ~8.0% at week 4) [106]. Future research in older individuals is required to investigate whether the combined effects of protein ingestion, resistance exercise, and n-3 PUFA supplementation under physiological conditions is more beneficial than resistance exercise and/or protein ingestion in isolation. It also remains to be seen whether changes in inflammatory markers underlie changes to MPS following n-3 PUFA supplementation.
2.3. Increasing Muscle Strength
The central nervous system (CNS) contains a high concentration of the unsaturated lipids, EPA and DHA, which define the structure and function of its cellular and subcellular components. These fatty acids are known to increase nerve conduction velocity in both young and older individuals through the modulation of sarcolemma ion channels which, in turn, improves contractile activity of the muscle [106,107,108,109,110]. In older women, dietary n-3 PUFA supplementation of 2 g/day for 90 days in conjunction with strength training has been shown to exert additional increases to strength and functional capacity (i.e., increased peak torque and rate of torque development) than strength training alone [34]. Rodacki and co-workers [34] also found these increases to occur through changes in neuromuscular junction conductivity, as evidenced by increases in electromyographic activation of the muscle and decreases in electromechanical delay. Even in the absence of regular resistance training, muscular strength and thigh volume can increase with n-3 PUFA supplementation in older men and women. Indeed, supplementation of 4 g/day for six months results in increased whole thigh muscle area, 1-RM strength and isokinetic power [32]. However, in the study that was conducted by Smith et al. diet was not controlled, which may magnify and thus misrepresent the effects of n-3 PUFA supplementation on muscle area [32]. Furthermore, without the collection of muscle biopsies, it is difficult to determine the precise molecular mechanism underlying these responses as well as the potential for similar shifts in muscle fiber types which a known to occur in rat muscle as a result of n-3 PUFA supplementation [111].
There is also evidence to suggest that older women may respond better than men to combined n-3 PUFA supplementation and resistance exercise training. For example, when compared to both male and female placebo groups, Da Boit and co-workers observed greater improvement in muscle quality (strength/unit of muscle area) in the quadriceps of older women following combined resistance exercise and fish-oil supplementation [78]. A possible explanation for this sexual dimorphism is that older women do not respond as well to resistance exercise as men and also have less muscle mass, therefore allowing a greater effect in women than men with n-3 PUFA supplementation due to their greater capacity for improvement. This theory however remains highly speculative as others have shown that older men and women respond equally as well to resistance exercise-based training and that no such sex differences exist in older individuals [112].
Furthermore, n-3 PUFAs have been shown to exert similar effects as the female sex hormone oestrogen, preventing bone loss in ovariectomised rats [113]. As such, another possible explanation regarding the sexual dimorphisms in fish oil efficacy is that n-3 PUFAs may function in a similar fashion to oestrogen, promoting anabolism or at least preventing catabolism. However, it should be emphasised here that this study was conducted using an animal model and further human research is required as sexual dimorphisms in protein metabolism are not yet fully understood [114]. Whether n-3 PUFAs induce effects similar to male or female sex hormones in human skeletal muscle related to anabolism remain to be determined.
2.4. Potential to Alter Satellite Cell Activation
Satellite cells are primarily involved in muscle regeneration in response to injury, such as those sustained following resistance exercise [18]. Upon their activation, satellite cells enter the cell cycle, proliferate, and differentiate to myoblasts and myocytes in a process termed myogenesis, before fusing to existing muscle fibers. Circulating systemic factors, such as hormones and inflammatory marker, can positively and negatively influence satellite cell activation. For instance, circulating growth factors, such as GDF11 and myostatin, as well as inflammatory markers, such as TNF-α and IL-6, increase with age, impairing the regenerative capacity of satellite cells in human skeletal muscle [10,115]. Though it is important to note that while GDF11 and myostatin have been correlated with increased ageing in humans, no study to date has demonstrated a causal link between these circulating factors and their ability to affect the regenerative capacity of muscle. Further evidence from parabiosis experiments in mice indicate that ageing reduces satellite cell activity; however exposing cells to a more youthful systemic environment and improves the regenerative capacity of satellite cells indicating that the age-related reduction in satellite cell activity can be modulated by systemic factors that change with age [116,117]. Thus, n-3 PUFAs with their anti-inflammatory properties could promote a systemic environment that improves satellite cell responsiveness to injury during aging by attenuating systemic inflammation in a similar manner to nonsteroidal anti-inflammatory drugs (NSAIDs) [118,119,120]. Currently, there is a paucity of human data to support the latter argument, nevertheless, studies investigating the influence of n-3 PUFAs myogenesis in vitro have been conducted and these findings are summarised in Table 1.
The Mechanisms Underlying the Modulatory Effects of n-3 PUFAs on Protein Metabolism Signalling and Myogenesis
Activation of the mechanistic target of rapamycin (mTOR) and its downstream signalling targets the 70 kDa ribosomal protein S6 kinase-1 (p70S6K1) and eIF4E-binding protein 1 (4E-BP1) are involved in the regulation of translation initiation responses that are required for skeletal muscle growth, hypertrophy, and also have a role in myogenesis [121]. In isolation, n-3 PUFAs do not appear to have any effect on mTOR signalling pathway in fully formed C2C12 cells [80]. However, EPA exclusively augments leucine induced MPS in fully differentiated C2C12 myotubes and increases the phosphorylation of p70S6K1, with no changes in the phosphorylation of any of the other anabolic mTOR pathway members [122]. Others have also shown EPA and DHA to induce similar increases in p70S6K1 phosphorylation without leucine stimulation during differentiation [79]. The activation of p70S6K1 following n-3 PUFA treatment in this case is not associated with an increase in Akt activity, which is an upstream effector of mTOR that is implicated in skeletal muscle hypertrophy [79,123]. The fact that changes to p70S6K1 activity [46] and phosphorylation [79] can occur without prior changes to Akt, suggests that n-3 PUFAs are capable of modulating differentiation and MPS independently of canonical mTOR signalling. This alternate activation may possibly come through signalling proteins, such as the mitogen-activated protein kinase (MAPKs), which have been shown to phosphorylate members of the mTOR signalling pathway [124].
The MAPK cascade(s), particularly the extracellular signal-regulated kinases (ERK) 1 and 2 (ERK1/2), play a duel role in myogenesis [125,126]. During proliferation, ERK1/2 expression is increased, promoting proliferation and inhibiting differentiation [127,128]. In contrast to this inhibitory effect during the early stages of differentiation ERK1/2 becomes phosphorylated during the late stages of differentiation promoting the fusion and formation of myotubes [129,130].
The actions of MAPK signalling in the regulation of myogenesis have been observed in cells treated with n-3 PUFAs. Exposing proliferating C2C12 cells to DHA and EPA for 24 h decreases MAPK/ERK1/2 phosphorylation, preventing the progression of myoblasts from the G1 to S phase [131]. When the same cells are proliferated in the absence of n-3 PUFAs for 24 h, they continue through the cell cycle, which is noteworthy as it demonstrates that n-3 PUFAs can regulate a return to quiescence, keeping the cells in the G1 phase. This is noteworthy as there are particular muscle wasting conditions where satellite cells are constitutively active as inflammation is elevated, thus impairing their ability to self-renew [132,133]. Constitutively active satellite cells also cause precocious differentiation (i.e., premature and excessive differentiation), further reducing the satellite cell pool and regenerative capacity of muscle [132,133]. Thus, n-3 PUFAs may have the potential to preserve muscle mass by keeping cells in a quiescent state preserving the satellite pool or at least delay their inevitable activation following the induction of differentiation in vitro. Whether n-3 PUFAs are able to prevent precocious differentiation under pathophysiological conditions in vivo where inflammation is elevated remains to be determined.
Aside from regulating proliferation and differentiation, MAPKs can also initiate cell death responses in the face of noxious stimuli, such as palmitate and TNF-α [134,135]. EPA combined with palmitate and/or TNF-α treatment during C2 differentiation partially rescues cell death and differentiation via the suppression of MAPK [135]. Saini and co-workers [135] also show that EPA promotes differentiation by increasing the expression of MyoD and Myogenin. Whether the other n-3 PUFAs, such as DHA and DPA, elicit the same inhibitory effects on MAPK induced cell death and increase MRFs during differentiation remains unclear. The γ isoform of p38 can also modulate myogenesis by repressing the transcriptional activity of MyoD promoting proliferation [136] although it is unknown whether EPA and DHA alter the activation of p38 MAPK in the context of myogenesis. Future research incorporating in vitro models is required to fully understand the effect that n-3 PUFAs exert on MAPKs during myogenesis and how this affects other pathways that are involved in growth, such as mTOR signalling during this process.
3. Limitations and Differences between In Vitro Cell Culture and In Vivo Human Studies
It is evident that EPA and DHA can elicit both similar and differential effects during various stages of myogenesis (Table 1), but also in full differentiated myotubes (Table 2). The differences identified could be due to structural characteristics of the two fatty acids as well as their ability to produce different metabolite species [137,138]. It is important to highlight that the effects of n-3 PUFA in vitro are dose and time dependent and an accurate balance between those parameters is required to significantly augment in vitro proliferation, differentiation, and post-differentiation events. Inconsistency in these parameters may also account for significant differences between published reports. For example, very high concentrations (200–600 μM) of EPA added to mice C2C12 myoblasts during proliferation for up to 10 days induce transdifferentiation to adipocytes [139]. In contrast, EPA has no effect on proliferation rate of C2C12 cells at concentration of 0.1–10 µM even though treated for 48 h, while DHA proliferative stimulation starts at 10 µM [140]. Exposure to EPA and DHA at higher concentrations (20–100 µM) for 24 h slows down the cell growth rate by preventing the transition of C2C12 cells from G1 to S phase of the cell cycle with DHA having a more pronounced effect [131]. Additionally, myotubes can be more resilient than myoblasts as concentrations of EPA and DHA, ranging from 400 to 600 µM in myotubes, do not cause protein degradation [69,72] (Table 2). While 50 μM of both EPA and DHA promote myogenic responses during differentiation in C2C12 cells (Table 1), other cell lines such as L6 rat cells are sensitive to 20 μM of EPA and DHA. Furthermore, while studying in vitro myogenesis provides mechanistic insight into the processes underlying muscle development and regeneration, it more closely resembles embryonic muscle development and does not necessarily recapitulate what occurs during myogenesis in adult muscle following injury or exercise. Nevertheless, in vivo human studies investigating the effects on n-3 PUFAs on satellite cell activation are warranted to compliment the in vitro data.
One of the major discrepancies in human studies investigating the effects of n-3 PUFAs alone or combined with exercise and/or protein ingestion is the n-3 PUFA dosage and supplementation duration (Table 3). This discrepancy makes it difficult to elucidate the optimal supplementation duration and the dosage that is required to have an effect on intracellular signalling and muscle anabolism responses. These dose discrepancies may arise due to regional differences in the availability of certain n-3 PUFA supplements, whereas discrepancies in the duration of the supplementation may due to the research question being asked (i.e., acute MPS responses vs chronic changes in lean mass).
4. Future Directions
With respect to muscle fiber hypertrophy, studies concluding that satellite cells are necessary for muscle fiber hypertrophy utilize young or developing mice [146,147] (<4 months of age). However, in models of fully grown/ developed mice (>4 months of age), short-term hypertrophy occurs in the absence of satellite cells [20,24,25]. While satellite cells play an integral role in regulating the extracellular matrix during remodeling [26,30,31], it is yet to be determined whether excessive extracellular matrix (ECM) accumulation or a “myonuclear domain ceiling” limits prolonged hypertrophy in the absence of satellite cells. As such, it is likely that satellite cells are necessary for muscle fiber hypertrophy beyond a certain threshold. With regards to n-3 PUFAs, six months of supplementation in older individuals has been shown to increase gene expression of various pathways that are involved in growth and structural support (ECM organization) [148]. These small n-3 PUFA-induced transcriptional changes in pathways related to growth and ECM organization could modulate satellite cell function given the reciprocal relationship between fibroblasts and satellite cells [31]. Thus, n-3 PUFAs may possess the capacity to alter satellite cell activity directly by altering proliferation and differentiation or indirectly via changes to the ECM.
Furthermore, the in vitro effects of DPA on myogenesis remain largely understudied. This represents an area for future investigations as DPA is a biologically active n-3 PUFA with similar properties to DHA and EPA and can also be readily converted to EPA [37,149,150]. Thus, further research is required to elucidate whether DPA can also induce myogenesis in a dose dependant manner by regulating the same molecular targets as EPA and DHA.
Exploring the effects of n-3 PUFAs on human satellite cell myogenesis in vitro as well as ex vivo would provide new insight into the effects timing and dose elicit during the muscle repair process. Understanding precisely how n-3 PUFAs are able to alter anabolic signaling following hypertrophic stimuli, such as protein ingestion and resistance exercise could see novel n-3 PUFA supplementation strategies developed to maximize these responses increasing or at the very least maintaining skeletal muscle mass.
5. Summary and Conclusions
The mechanisms by which n-3 PUFAs exert their multiple beneficial effects center largely on their increased incorporation into cellular membranes which induce changes to various phospholipid species used as substrates for various signalling cascades. n-3 PUFAs are also capable of reducing systemic inflammation by inhibiting the release of pro-inflammatory cytokines from immune cells [151,152,153] and improving the signalling efficiency of proteins that are involved in growth and hypertrophy [46,72,79]. However, several questions remain unanswered regarding n-3 PUFA supplementation and human skeletal muscle metabolism. Firstly, the optimal dosage and duration required to elicit beneficial responses in skeletal muscle remains unclear. As it stands, supplementation dosages ranging from 2 to 5 g/day for a minimum of four weeks results in improvements in anabolic signalling efficiency and muscle strength outcomes (Table 3). In older individuals sex differences in muscle quality have been observed following n-3 PUFA supplementation [78]. The mechanistic basis for these differences remains to be determined. Other sexual dimorphisms may also exist following n-3 PUFA supplementation, such as changes to anabolic signalling activity and rates of MPS, though no study to date has been able to detect these changes.
Secondly, it is also possible that doses within ranges of 2–5 g/day may affect satellite cell activity particularly after exercise, although experimental evidence is required to confirm this hypothesis. The anti-lipotoxic and anti-cytotoxic properties of EPA and DHA during in vitro myogenesis have been established with 50 µM being an effective concentration to induce protective effects. Whether EPA, DHA, and DPA similarly or differentially modulate the activity and expression of myogenic regulatory factors at 50 µM during myogenesis remains unclear.
Lastly, the capacity for n-3 PUFAs to activate satellite cells in human skeletal muscle also remains to be determined. This knowledge possesses important health and clinical implications by providing further support for n-3 PUFA supplementation to augment the growth, development and regenerative capabilities of skeletal muscle. Establishing how EPA, DHA, and DPA modulate myogenesis and alters intracellular anabolic signaling would provide a greater understanding as to how n-3 PUFA supplementation can be used as an effective nutritional strategy in conjunction with exercise to increase and maintain muscle mass.
Acknowledgments
O.L.-K. is supported by an Australian Catholic University Research Fund (ACURF) 2016000363.
Author Contributions
B.T., D.C. and O.L.-K. conceived and wrote the manuscript. All authors approved the final manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hawley, J.A.; Hargreaves, M.; Joyner, M.J.; Zierath, J.R. Integrative biology of exercise. Cell 2014, 159, 738–749. [Google Scholar] [CrossRef] [PubMed]
- Cuthbertson, D.; Smith, K.; Babraj, J.; Leese, G.; Waddell, T.; Atherton, P.; Wackerhage, H.; Taylor, P.M.; Rennie, M.J. Anabolic signaling deficits underlie amino acid resistance of wasting, aging muscle. FASEB J. 2005, 19, 422–424. [Google Scholar] [CrossRef] [PubMed]
- Wall, B.T.; Gorissen, S.H.; Pennings, B.; Koopman, R.; Groen, B.B.; Verdijk, L.B.; van Loon, L.J. Aging is accompanied by a blunted muscle protein synthetic response to protein ingestion. PLoS ONE 2015, 10, e0140903. [Google Scholar] [CrossRef] [PubMed]
- Brook, M.S.; Wilkinson, D.J.; Mitchell, W.K.; Lund, J.N.; Phillips, B.E.; Szewczyk, N.J.; Greenhaff, P.L.; Smith, K.; Atherton, P.J. Synchronous deficits in cumulative muscle protein synthesis and ribosomal biogenesis underlie age-related anabolic resistance to exercise in humans. J. Physiol. 2016, 594, 7399–7417. [Google Scholar] [CrossRef] [PubMed]
- Fry, C.S.; Drummond, M.J.; Glynn, E.L.; Dickinson, J.M.; Gundermann, D.M.; Timmerman, K.L.; Walker, D.K.; Dhanani, S.; Volpi, E.; Rasmussen, B.B. Aging impairs contraction-induced human skeletal muscle mtorc1 signaling and protein synthesis. Skelet Muscle 2011, 1, 11. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, I.H. Sarcopenia: Origins and clinical relevance. J. Nutr. 1997, 127, 990S–991S. [Google Scholar] [CrossRef] [PubMed]
- Volpi, E.; Mittendorfer, B.; Rasmussen, B.B.; Wolfe, R.R. The response of muscle protein anabolism to combined hyperaminoacidemia and glucose-induced hyperinsulinemia is impaired in the elderly. J. Clin. Endocrinol. Metab. 2000, 85, 4481–4490. [Google Scholar] [CrossRef] [PubMed]
- Janssen, H.C.; Samson, M.M.; Verhaar, H.J. Vitamin d deficiency, muscle function, and falls in elderly people. Am. J. Clin. Nutr. 2002, 75, 611–615. [Google Scholar] [CrossRef] [PubMed]
- Szulc, P.; Duboeuf, F.; Marchand, F.; Delmas, P.D. Hormonal and lifestyle determinants of appendicular skeletal muscle mass in men: The minos study. Am. J. Clin. Nutr. 2004, 80, 496–503. [Google Scholar] [CrossRef] [PubMed]
- Visser, M.; Pahor, M.; Taaffe, D.R.; Goodpaster, B.H.; Simonsick, E.M.; Newman, A.B.; Nevitt, M.; Harris, T.B. Relationship of interleukin-6 and tumor necrosis factor-alpha with muscle mass and muscle strength in elderly men and women: The health abc study. J. Gerontol. A Biol. Sci. Med. Sci. 2002, 57, M326–M332. [Google Scholar] [CrossRef] [PubMed]
- McKay, B.R.; Ogborn, D.I.; Baker, J.M.; Toth, K.G.; Tarnopolsky, M.A.; Parise, G. Elevated SOCS3 and altered IL-6 signaling is associated with age-related human muscle stem cell dysfunction. Am. J. Physiol. Cell Physiol. 2013, 304, C717–C728. [Google Scholar] [CrossRef] [PubMed]
- Vandervoort, A.A. Aging of the human neuromuscular system. Muscle Nerve 2002, 25, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Verdijk, L.B.; Snijders, T.; Drost, M.; Delhaas, T.; Kadi, F.; van Loon, L.J. Satellite cells in human skeletal muscle; from birth to old age. Age 2014, 36, 545–547. [Google Scholar] [CrossRef] [PubMed]
- Verdijk, L.B.; Koopman, R.; Schaart, G.; Meijer, K.; Savelberg, H.H.; van Loon, L.J. Satellite cell content is specifically reduced in type II skeletal muscle fibers in the elderly. Am. J. Physiol. Endocrinol. Metab. 2007, 292, E151–E157. [Google Scholar] [CrossRef] [PubMed]
- Larsson, L.; Sjodin, B.; Karlsson, J. Histochemical and biochemical changes in human skeletal muscle with age in sedentary males, age 22–65 years. Acta Physiol. Scand. 1978, 103, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Frontera, W.R.; Zayas, A.R.; Rodriguez, N. Aging of human muscle: Understanding sarcopenia at the single muscle cell level. Phys. Med. Rehabil. Clin. N. Am. 2012, 23, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Miljkovic, N.; Lim, J.Y.; Miljkovic, I.; Frontera, W.R. Aging of skeletal muscle fibers. Ann. Rehabil. Med. 2015, 39, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Relaix, F.; Marcelle, C. Muscle stem cells. Curr. Opin. Cell Biol. 2009, 21, 748–753. [Google Scholar] [CrossRef] [PubMed]
- Sambasivan, R.; Yao, R.; Kissenpfennig, A.; Van Wittenberghe, L.; Paldi, A.; Gayraud-Morel, B.; Guenou, H.; Malissen, B.; Tajbakhsh, S.; Galy, A. Pax7-expressing satellite cells are indispensable for adult skeletal muscle regeneration. Development 2011, 138, 3647–3656. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, J.J.; Mula, J.; Miyazaki, M.; Erfani, R.; Garrison, K.; Farooqui, A.B.; Srikuea, R.; Lawson, B.A.; Grimes, B.; Keller, C.; et al. Effective fiber hypertrophy in satellite cell-depleted skeletal muscle. Development 2011, 138, 3657–3666. [Google Scholar] [CrossRef] [PubMed]
- Olguin, H.C.; Olwin, B.B. Pax-7 up-regulation inhibits myogenesis and cell cycle progression in satellite cells: A potential mechanism for self-renewal. Dev. Biol. 2004, 275, 375–388. [Google Scholar] [CrossRef] [PubMed]
- Venuti, J.M.; Morris, J.H.; Vivian, J.L.; Olson, E.N.; Klein, W.H. Myogenin is required for late but not early aspects of myogenesis during mouse development. J. Cell Biol. 1995, 128, 563–576. [Google Scholar] [CrossRef] [PubMed]
- Fukada, S.I. The roles of muscle stem cells in muscle injury, atrophy, and hypertrophy. J. Biochem. 2018. [Google Scholar] [CrossRef] [PubMed]
- Murach, K.A.; Fry, C.S.; Kirby, T.J.; Jackson, J.R.; Lee, J.D.; White, S.H.; Dupont-Versteegden, E.E.; McCarthy, J.J.; Peterson, C.A. Starring or supporting role? Satellite cells and skeletal muscle fiber size regulation. Physiology 2018, 33, 26–38. [Google Scholar] [CrossRef] [PubMed]
- Murach, K.A.; White, S.H.; Wen, Y.; Ho, A.; Dupont-Versteegden, E.E.; McCarthy, J.J.; Peterson, C.A. Differential requirement for satellite cells during overload-induced muscle hypertrophy in growing versus mature mice. Skelet Muscle 2017, 7, 14. [Google Scholar] [CrossRef] [PubMed]
- Fry, C.S.; Lee, J.D.; Jackson, J.R.; Kirby, T.J.; Stasko, S.A.; Liu, H.; Dupont-Versteegden, E.E.; McCarthy, J.J.; Peterson, C.A. Regulation of the muscle fiber microenvironment by activated satellite cells during hypertrophy. FASEB J. 2014, 28, 1654–1665. [Google Scholar] [CrossRef] [PubMed]
- Egner, I.M.; Bruusgaard, J.C.; Gundersen, K. Satellite cell depletion prevents fiber hypertrophy in skeletal muscle. Development 2016, 143, 2898–2906. [Google Scholar] [CrossRef] [PubMed]
- Keefe, A.C.; Lawson, J.A.; Flygare, S.D.; Fox, Z.D.; Colasanto, M.P.; Mathew, S.J.; Yandell, M.; Kardon, G. Muscle stem cells contribute to myofibers in sedentary adult mice. Nat. Commun. 2015, 6, 7087. [Google Scholar] [CrossRef] [PubMed]
- Fry, C.S.; Lee, J.D.; Mula, J.; Kirby, T.J.; Jackson, J.R.; Liu, F.; Yang, L.; Mendias, C.L.; Dupont-Versteegden, E.E.; McCarthy, J.J.; et al. Inducible depletion of satellite cells in adult, sedentary mice impairs muscle regenerative capacity but does not contribute to sarcopenia. Nat. Med. 2015, 21, 76–80. [Google Scholar] [CrossRef] [PubMed]
- Fry, C.S.; Kirby, T.J.; Kosmac, K.; McCarthy, J.J.; Peterson, C.A. Myogenic progenitor cells control extracellular matrix production by fibroblasts during skeletal muscle hypertrophy. Cell Stem Cell 2017, 20, 56–69. [Google Scholar] [CrossRef] [PubMed]
- Murphy, M.M.; Lawson, J.A.; Mathew, S.J.; Hutcheson, D.A.; Kardon, G. Satellite cells, connective tissue fibroblasts and their interactions are crucial for muscle regeneration. Development 2011, 138, 3625–3637. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.I.; Julliand, S.; Reeds, D.N.; Sinacore, D.R.; Klein, S.; Mittendorfer, B. Fish oil-derived n-3 pufa therapy increases muscle mass and function in healthy older adults. Am. J. Clin. Nutr. 2015, 102, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.I.; Atherton, P.; Reeds, D.N.; Mohammed, B.S.; Rankin, D.; Rennie, M.J.; Mittendorfer, B. Omega-3 polyunsaturated fatty acids augment the muscle protein anabolic response to hyperinsulinaemia-hyperaminoacidaemia in healthy young and middle-aged men and women. Clin. Sci. 2011, 121, 267–278. [Google Scholar] [CrossRef] [PubMed]
- Rodacki, C.L.; Rodacki, A.L.; Pereira, G.; Naliwaiko, K.; Coelho, I.; Pequito, D.; Fernandes, L.C. Fish-oil supplementation enhances the effects of strength training in elderly women. Am. J. Clin. Nutr. 2012, 95, 428–436. [Google Scholar] [CrossRef] [PubMed]
- Simopoulos, A.P. Human requirement for n-3 polyunsaturated fatty acids. Poult. Sci. 2000, 79, 961–970. [Google Scholar] [CrossRef] [PubMed]
- Baker, E.J.; Miles, E.A.; Burdge, G.C.; Yaqoob, P.; Calder, P.C. Metabolism and functional effects of plant-derived omega-3 fatty acids in humans. Prog. Lipid Res. 2016, 64, 30–56. [Google Scholar] [CrossRef] [PubMed]
- Kaur, G.; Sinclair, A.J.; Cameron-Smith, D.; Barr, D.P.; Molero-Navajas, J.C.; Konstantopoulos, N. Docosapentaenoic acid (22:5n-3) down-regulates the expression of genes involved in fat synthesis in liver cells. Prostaglandins Leukot Essent Fatty Acids 2011, 85, 155–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burdge, G.C.; Jones, A.E.; Wootton, S.A. Eicosapentaenoic and docosapentaenoic acids are the principal products of alpha-linolenic acid metabolism in young men. Br. J. Nutr. 2002, 88, 355–363. [Google Scholar] [CrossRef] [PubMed]
- Burdge, G.C.; Wootton, S.A. Conversion of alpha-linolenic acid to eicosapentaenoic, docosapentaenoic and docosahexaenoic acids in young women. Br. J. Nutr. 2002, 88, 411–420. [Google Scholar] [CrossRef] [PubMed]
- Arterburn, L.M.; Hall, E.B.; Oken, H. Distribution, interconversion, and dose response of n-3 fatty acids in humans. Am. J. Clin. Nutr. 2006, 83, 1467S. [Google Scholar] [CrossRef] [PubMed]
- Morin, C.; Charbonneau, L.; Ouellet, N.; Ouellet, H.; Blier, P.U.; Dufresne, F.; Fortin, S. Eicosapentaenoic acid monoglyceride resolves inflammation in an ex vivo model of human peripheral blood mononuclear cell. Eur. J. Pharmacol. 2017, 807, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Vedin, I.; Cederholm, T.; Freund Levi, Y.; Basun, H.; Garlind, A.; Faxen Irving, G.; Jonhagen, M.E.; Vessby, B.; Wahlund, L.O.; Palmblad, J. Effects of docosahexaenoic acid-rich n-3 fatty acid supplementation on cytokine release from blood mononuclear leukocytes: The omegad study. Am. J. Clin. Nutr. 2008, 87, 1616–1622. [Google Scholar] [CrossRef] [PubMed]
- Shindou, H.; Koso, H.; Sasaki, J.; Nakanishi, H.; Sagara, H.; Nakagawa, K.M.; Takahashi, Y.; Hishikawa, D.; Iizuka-Hishikawa, Y.; Tokumasu, F.; et al. Docosahexaenoic acid preserves visual function by maintaining correct disc morphology in retinal photoreceptor cells. J. Biol. Chem. 2017, 292, 12054–12064. [Google Scholar] [CrossRef] [PubMed]
- Litman, B.J.; Niu, S.L.; Polozova, A.; Mitchell, D.C. The role of docosahexaenoic acid containing phospholipids in modulating G protein-coupled signaling pathways: Visual transduction. J. Mol. Neurosci. 2001, 16. [Google Scholar] [CrossRef]
- Albert, B.B.; Derraik, J.G.; Brennan, C.M.; Biggs, J.B.; Smith, G.C.; Garg, M.L.; Cameron-Smith, D.; Hofman, P.L.; Cutfield, W.S. Higher omega-3 index is associated with increased insulin sensitivity and more favourable metabolic profile in middle-aged overweight men. Sci. Rep. 2014, 4, 6697. [Google Scholar] [CrossRef] [PubMed]
- McGlory, C.; Wardle, S.L.; Macnaughton, L.S.; Witard, O.C.; Scott, F.; Dick, J.; Bell, J.G.; Phillips, S.M.; Galloway, S.D.; Hamilton, D.L.; et al. Fish oil supplementation suppresses resistance exercise and feeding-induced increases in anabolic signaling without affecting myofibrillar protein synthesis in young men. Physiol. Rep. 2016, 4, e12715. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.I.; Atherton, P.; Reeds, D.N.; Mohammed, B.S.; Rankin, D.; Rennie, M.J.; Mittendorfer, B. Dietary omega-3 fatty acid supplementation increases the rate of muscle protein synthesis in older adults: A randomized controlled trial. Am. J. Clin. Nutr. 2011, 93, 402–412. [Google Scholar] [CrossRef] [PubMed]
- Bhullar, A.S.; Putman, C.T.; Mazurak, V.C. Potential role of omega-3 fatty acids on the myogenic program of satellite cells. Nutr. Metab. Insights 2016, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kalinkovich, A.; Livshits, G. Sarcopenic obesity or obese sarcopenia: A cross talk between age-associated adipose tissue and skeletal muscle inflammation as a main mechanism of the pathogenesis. Ageing Res. Rev. 2017, 35, 200–221. [Google Scholar] [CrossRef] [PubMed]
- Millward, D.J. Nutrition and sarcopenia: Evidence for an interaction. Proc. Nutr. Soc. 2012, 71, 566–575. [Google Scholar] [CrossRef] [PubMed]
- Phillips, S.M. Nutritional supplements in support of resistance exercise to counter age-related sarcopenia. Adv. Nutr. 2015, 6, 452–460. [Google Scholar] [CrossRef] [PubMed]
- Soysal, P.; Stubbs, B.; Lucato, P.; Luchini, C.; Solmi, M.; Peluso, R.; Sergi, G.; Isik, A.T.; Manzato, E.; Maggi, S.; et al. Inflammation and frailty in the elderly: A systematic review and meta-analysis. Ageing Res. Rev. 2016, 31, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Bano, G.; Trevisan, C.; Carraro, S.; Solmi, M.; Luchini, C.; Stubbs, B.; Manzato, E.; Sergi, G.; Veronese, N. Inflammation and sarcopenia: A systematic review and meta-analysis. Maturitas 2017, 96, 10–15. [Google Scholar] [CrossRef] [PubMed]
- McKay, B.R.; De Lisio, M.; Johnston, A.P.; O’Reilly, C.E.; Phillips, S.M.; Tarnopolsky, M.A.; Parise, G. Association of interleukin-6 signalling with the muscle stem cell response following muscle-lengthening contractions in humans. PLoS ONE 2009, 4, e6027. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Li, Y.; Wu, Y.; Wang, L.; Wang, X.; Du, J. Interleukin-6/signal transducer and activator of transcription 3 (STAT3) pathway is essential for macrophage infiltration and myoblast proliferation during muscle regeneration. J. Biol. Chem. 2013, 288, 1489–1499. [Google Scholar] [CrossRef] [PubMed]
- Sindhu, S.; Thomas, R.; Shihab, P.; Sriraman, D.; Behbehani, K.; Ahmad, R. Obesity is a positive modulator of IL-6R and IL-6 expression in the subcutaneous adipose tissue: Significance for metabolic inflammation. PLoS ONE 2015, 10, e0133494. [Google Scholar] [CrossRef] [PubMed]
- Feinstein, R.; Kanety, H.; Papa, M.Z.; Lunenfeld, B.; Karasik, A. Tumor necrosis factor-alpha suppresses insulin-induced tyrosine phosphorylation of insulin receptor and its substrates. J. Biol. Chem. 1993, 268, 26055–26058. [Google Scholar] [PubMed]
- Hotamisligil, G.S.; Shargill, N.S.; Spiegelman, B.M. Adipose expression of tumor necrosis factor-alpha: Direct role in obesity-linked insulin resistance. Science 1993, 259, 87–91. [Google Scholar] [CrossRef] [PubMed]
- Merritt, E.K.; Stec, M.J.; Thalacker-Mercer, A.; Windham, S.T.; Cross, J.M.; Shelley, D.P.; Craig Tuggle, S.; Kosek, D.J.; Kim, J.S.; Bamman, M.M. Heightened muscle inflammation susceptibility may impair regenerative capacity in aging humans. J. Appl. Physiol. 2013, 115, 937–948. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Long-chain fatty acids and inflammation. Proc. Nutr. Soc. 2012, 71, 284–289. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. N-3 fatty acids, inflammation and immunity: New mechanisms to explain old actions. Proc. Nutr. Soc. 2013, 72, 326–336. [Google Scholar] [CrossRef] [PubMed]
- Wada, M.; DeLong, C.J.; Hong, Y.H.; Rieke, C.J.; Song, I.; Sidhu, R.S.; Yuan, C.; Warnock, M.; Schmaier, A.H.; Yokoyama, C.; et al. Enzymes and receptors of prostaglandin pathways with arachidonic acid-derived versus eicosapentaenoic acid-derived substrates and products. J. Biol. Chem. 2007, 282, 22254–22266. [Google Scholar] [CrossRef] [PubMed]
- Kelley, D.S.; Taylor, P.C.; Nelson, G.J.; Schmidt, P.C.; Mackey, B.E.; Kyle, D. Effects of dietary arachidonic acid on human immune response. Lipids 1997, 32, 449–456. [Google Scholar] [CrossRef] [PubMed]
- Kelley, D.S.; Taylor, P.C.; Nelson, G.J.; Mackey, B.E. Arachidonic acid supplementation enhances synthesis of eicosanoids without suppressing immune functions in young healthy men. Lipids 1998, 33, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Phang, M.; Garg, M.L.; Sinclair, A.J. Inhibition of platelet aggregation by omega-3 polyunsaturated fatty acids is gender specific-redefining platelet response to fish oils. Prostaglandins Leukot Essent Fatty Acids 2009, 81, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Bartram, H.P.; Gostner, A.; Scheppach, W.; Reddy, B.S.; Rao, C.V.; Dusel, G.; Richter, F.; Richter, A.; Kasper, H. Effects of fish oil on rectal cell proliferation, mucosal fatty acids, and prostaglandin e2 release in healthy subjects. Gastroenterology 1993, 105, 1317–1322. [Google Scholar] [CrossRef]
- Calder, P.C. Polyunsaturated fatty acids and inflammatory processes: New twists in an old tale. Biochimie 2009, 91, 791–795. [Google Scholar] [CrossRef] [PubMed]
- Cai, D.; Frantz, J.D.; Tawa, N.E., Jr.; Melendez, P.A.; Oh, B.C.; Lidov, H.G.; Hasselgren, P.O.; Frontera, W.R.; Lee, J.; Glass, D.J.; et al. Ikkbeta/nf-kappab activation causes severe muscle wasting in mice. Cell 2004, 119, 285–298. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Lin, Q.W.; Zheng, P.P.; Zhang, J.S.; Huang, F.R. Dha inhibits protein degradation more efficiently than epa by regulating the ppargamma/nfkappab pathway in c2c12 myotubes. Biomed. Res. Int. 2013, 2013, 318981. [Google Scholar] [PubMed]
- Camandola, S.; Leonarduzzi, G.; Musso, T.; Varesio, L.; Carini, R.; Scavazza, A.; Chiarpotto, E.; Baeuerle, P.A.; Poli, G. Nuclear factor kb is activated by arachidonic acid but not by eicosapentaenoic acid. Biochem. Biophys. Res. Commun. 1996, 229, 643–647. [Google Scholar] [CrossRef] [PubMed]
- Williams-Bey, Y.; Boularan, C.; Vural, A.; Huang, N.N.; Hwang, I.Y.; Shan-Shi, C.; Kehrl, J.H. Omega-3 free fatty acids suppress macrophage inflammasome activation by inhibiting nf-κb activation and enhancing autophagy. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Wei, H.; Luo, H.; Jiang, S.; Peng, J. EPA inhibits the inhibitor of kappabalpha (IκBα)/NF-κB/muscle ring finger 1 pathway in C2C12 myotubes in a PPARγ-dependent manner. Br. J. Nutr. 2011, 105, 348–356. [Google Scholar] [CrossRef] [PubMed]
- Serrano, A.L.; Baeza-Raja, B.; Perdiguero, E.; Jardi, M.; Munoz-Canoves, P. Interleukin-6 is an essential regulator of satellite cell-mediated skeletal muscle hypertrophy. Cell Metab. 2008, 7, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.E.; Gerken, E.; Zhang, Y.; Zhan, M.; Mohan, R.K.; Li, A.S.; Reid, M.B.; Li, Y.P. Role of tnf-{alpha} signaling in regeneration of cardiotoxin-injured muscle. Am. J. Physiol. Cell Physiol. 2005, 289, C1179–C1187. [Google Scholar] [CrossRef] [PubMed]
- Langen, R.C.; Van Der Velden, J.L.; Schols, A.M.; Kelders, M.C.; Wouters, E.F.; Janssen-Heininger, Y.M. Tumor necrosis factor-alpha inhibits myogenic differentiation through myod protein destabilization. FASEB J. 2004, 18, 227–237. [Google Scholar] [CrossRef] [PubMed]
- Domingues-Faria, C.; Vasson, M.P.; Goncalves-Mendes, N.; Boirie, Y.; Walrand, S. Skeletal muscle regeneration and impact of aging and nutrition. Ageing Res. Rev. 2016, 26, 22–36. [Google Scholar] [CrossRef] [PubMed]
- Wahlin-Larsson, B.; Carnac, G.; Kadi, F. The influence of systemic inflammation on skeletal muscle in physically active elderly women. Age 2014, 36, 9718. [Google Scholar] [CrossRef] [PubMed]
- Da Boit, M.; Sibson, R.; Sivasubramaniam, S.; Meakin, J.R.; Greig, C.A.; Aspden, R.M.; Thies, F.; Jeromson, S.; Hamilton, D.L.; Speakman, J.R.; et al. Sex differences in the effect of fish-oil supplementation on the adaptive response to resistance exercise training in older people: A randomized controlled trial. Am. J. Clin. Nutr. 2017, 105, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Briolay, A.; Jaafar, R.; Nemoz, G.; Bessueille, L. Myogenic differentiation and lipid-raft composition of l6 skeletal muscle cells are modulated by pufas. Biochim. Biophys. Acta 2013, 1828, 602–613. [Google Scholar] [CrossRef] [PubMed]
- Jeromson, S.; Mackenzie, I.; Doherty, M.K.; Whitfield, P.D.; Bell, G.; Dick, J.; Shaw, A.; Rao, F.; Ashcroft, S.; Philp, A.; et al. Lipid remodelling and an altered membrane proteome may drive the effects of epa and dha treatment on skeletal muscle glucose uptake and protein accretion. Am. J. Physiol. Endocrinol. Metab. 2017. [Google Scholar] [CrossRef] [PubMed]
- Mukai, A.; Kurisaki, T.; Sato, S.B.; Kobayashi, T.; Kondoh, G.; Hashimoto, N. Dynamic clustering and dispersion of lipid rafts contribute to fusion competence of myogenic cells. Exp. Cell Res. 2009, 315, 3052–3063. [Google Scholar] [CrossRef] [PubMed]
- Grimaldi, P.A. Regulatory role of peroxisome proliferator-activated receptor delta (ppar delta) in muscle metabolism. A new target for metabolic syndrome treatment? Biochimie 2005, 87, 5–8. [Google Scholar] [CrossRef] [PubMed]
- Chandrashekar, P.; Manickam, R.; Ge, X.; Bonala, S.; McFarlane, C.; Sharma, M.; Wahli, W.; Kambadur, R. Inactivation of pparbeta/delta adversely affects satellite cells and reduces postnatal myogenesis. Am. J. Physiol. Endocrinol. Metab. 2015, 309, E122–E131. [Google Scholar] [CrossRef] [PubMed]
- Deng, B.; Zhang, F.; Chen, K.; Wen, J.; Huang, H.; Liu, W.; Ye, S.; Wang, L.; Yang, Y.; Gong, P.; et al. Myod promotes porcine ppargamma gene expression through an e-box and a myod-binding site in the ppargamma promoter region. Cell Tissue Res. 2016, 365, 381–391. [Google Scholar] [CrossRef] [PubMed]
- Varga, T.; Czimmerer, Z.; Nagy, L. Ppars are a unique set of fatty acid regulated transcription factors controlling both lipid metabolism and inflammation. Biochim. Biophys. Acta 2011, 1812, 1007–1022. [Google Scholar] [CrossRef] [PubMed]
- Fritsche, K.L. The science of fatty acids and inflammation. Adv. Nutr. 2015, 6, 293S–301S. [Google Scholar] [CrossRef] [PubMed]
- Magee, P.; Pearson, S.; Allen, J. The omega-3 fatty acid, eicosapentaenoic acid (EPA), prevents the damaging effects of tumour necrosis factor (TNF)-alpha during murine skeletal muscle cell differentiation. Lipids Health Dis. 2008, 7, 24. [Google Scholar] [CrossRef] [PubMed]
- Magee, P.; Pearson, S.; Whittingham-Dowd, J.; Allen, J. Ppargamma as a molecular target of epa anti-inflammatory activity during tnf-alpha-impaired skeletal muscle cell differentiation. J. Nutr. Biochem. 2012, 23, 1440–1448. [Google Scholar] [CrossRef] [PubMed]
- Angione, A.R.; Jiang, C.; Pan, D.; Wang, Y.-X.; Kuang, S. Pparδ regulates satellite cell proliferation and skeletal muscle regeneration. Skeletal Muscle 2011, 1, 33. [Google Scholar] [CrossRef] [PubMed]
- Eisele, P.S.; Salatino, S.; Sobek, J.; Hottiger, M.O.; Handschin, C. The peroxisome proliferator-activated receptor gamma coactivator 1α/β (PGC-1) coactivators repress the transcriptional activity of NF-κB in skeletal muscle cells. J. Biol. Chem. 2013, 288, 2246–2260. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.S.; Shin, Y.; Moon, S.; Kim, S.; Kim, Y. Effects of eicosapentaenoic acid and docosahexaenoic acid on mitochondrial DNA replication and PGC-1α gene expression in C(2)C(12) muscle cells. Prev. Nutr. Food Sci. 2016, 21, 317–322. [Google Scholar] [CrossRef] [PubMed]
- Virbasius, J.V.; Scarpulla, R.C. Activation of the human mitochondrial transcription. Proc. Natl. Acad. Sci. USA 1994, 91, 1309–1313. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Zhao, Y.; Li, R.; Gong, J.; Zheng, Y.; Wang, Y. Pgc-1α is associated with c2c12 myoblast differentiation. Cent. Eur. J. Biol. 2014, 9, 1030–1036. [Google Scholar] [CrossRef]
- Hancock, C.R.; Han, D.H.; Chen, M.; Terada, S.; Yasuda, T.; Wright, D.C.; Holloszy, J.O. High-fat diets cause insulin resistance despite an increase in muscle mitochondria. Proc. Natl. Acad. Sci. USA 2008, 105, 7815–7820. [Google Scholar] [CrossRef] [PubMed]
- Pinel, A.; Rigaudiere, J.P.; Laillet, B.; Pouyet, C.; Malpuech-Brugere, C.; Prip-Buus, C.; Morio, B.; Capel, F. N-3pufa differentially modulate palmitate-induced lipotoxicity through alterations of its metabolism in C2C12 muscle cells. Biochim. Biophys. Acta 2016, 1861, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Bryner, R.W.; Woodworth-Hobbs, M.E.; Williamson, D.L.; Alway, S.E. Docosahexaenoic acid protects muscle cells from palmitate-induced atrophy. ISRN Obes. 2012, 2012, 647348. [Google Scholar] [CrossRef] [PubMed]
- Endres, S.; Ghorbani, R.; Kelley, V.E.; Georgilis, K.; Lonnemann, G.; van der Meer, J.W.; Cannon, J.G.; Rogers, T.S.; Klempner, M.S.; Weber, P.C.; et al. The effect of dietary supplementation with n-3 polyunsaturated fatty acids on the synthesis of interleukin-1 and tumor necrosis factor by mononuclear cells. N. Engl. J. Med. 1989, 320, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Oh, D.Y.; Talukdar, S.; Bae, E.J.; Imamura, T.; Morinaga, H.; Fan, W.; Li, P.; Lu, W.J.; Watkins, S.M.; Olefsky, J.M. GPR120 is an omega-3 fatty acid receptor mediating potent anti-inflammatory and insulin-sensitizing effects. Cell 2010, 142, 687–698. [Google Scholar] [CrossRef] [PubMed]
- Rieu, I.; Magne, H.; Savary-Auzeloux, I.; Averous, J.; Bos, C.; Peyron, M.A.; Combaret, L.; Dardevet, D. Reduction of low grade inflammation restores blunting of postprandial muscle anabolism and limits sarcopenia in old rats. J. Physiol. 2009, 587, 5483–5492. [Google Scholar] [CrossRef] [PubMed]
- Moore, D.R.; Robinson, M.J.; Fry, J.L.; Tang, J.E.; Glover, E.I.; Wilkinson, S.B.; Prior, T.; Tarnopolsky, M.A.; Phillips, S.M. Ingested protein dose response of muscle and albumin protein synthesis after resistance exercise in young men. Am. J. Clin. Nutr. 2009, 89, 161–168. [Google Scholar] [CrossRef] [PubMed]
- MacDougall, J.D.; Gibala, M.J.; Tarnopolsky, M.A.; MacDonald, J.R.; Interisano, S.A.; Yarasheski, K.E. The time course for elevated muscle protein synthesis following heavy resistance exercise. Can. J. Appl. Physiol. 1995, 20, 480–486. [Google Scholar] [CrossRef] [PubMed]
- Witard, O.C.; Jackman, S.R.; Breen, L.; Smith, K.; Selby, A.; Tipton, K.D. Myofibrillar muscle protein synthesis rates subsequent to a meal in response to increasing doses of whey protein at rest and after resistance exercise. Am. J. Clin. Nutr. 2014, 99, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Hulmi, J.J.; Kovanen, V.; Lisko, I.; Selanne, H.; Mero, A.A. The effects of whey protein on myostatin and cell cycle-related gene expression responses to a single heavy resistance exercise bout in trained older men. Eur J. Appl. Physiol. 2008, 102, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Farup, J.; Rahbek, S.K.; Knudsen, I.S.; de Paoli, F.; Mackey, A.L.; Vissing, K. Whey protein supplementation accelerates satellite cell proliferation during recovery from eccentric exercise. Amino Acids 2014, 46, 2503–2516. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Selby, A.; Rankin, D.; Patel, R.; Atherton, P.; Hildebrandt, W.; Williams, J.; Smith, K.; Seynnes, O.; Hiscock, N.; et al. Age-related differences in the dose-response relationship of muscle protein synthesis to resistance exercise in young and old men. J. Physiol. 2009, 587, 211–217. [Google Scholar] [CrossRef] [PubMed]
- McGlory, C.; Galloway, S.D.; Hamilton, D.L.; McClintock, C.; Breen, L.; Dick, J.R.; Bell, J.G.; Tipton, K.D. Temporal changes in human skeletal muscle and blood lipid composition with fish oil supplementation. Prostaglandins Leukot Essent Fatty Acids 2014, 90, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Lauretani, F.; Bandinelli, S.; Bartali, B.; Cherubini, A.; Iorio, A.D.; Ble, A.; Giacomini, V.; Corsi, A.M.; Guralnik, J.M.; Ferrucci, L. Omega-6 and omega-3 fatty acids predict accelerated decline of peripheral nerve function in older persons. Eur J. Neurol. 2007, 14, 801–808. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.X.; Leaf, A. Effects of long-chain polyunsaturated fatty acids on the contraction of neonatal rat cardiac myocytes. Proc. Natl. Acad. Sci. USA 1994, 91, 9886–9890. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, S.C.; Perez, M.R.; Hammond, K.E.; Sheader, E.A.; Negretti, N. Direct and indirect modulation of rat cardiac sarcoplasmic reticulum function by n-3 polyunsaturated fatty acids. J. Physiol. 2002, 538, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Lewis, E.J.; Radonic, P.W.; Wolever, T.M.; Wells, G.D. 21 Days of mammalian omega-3 fatty acid supplementation improves aspects of neuromuscular function and performance in male athletes compared to olive oil placebo. J. Int. Soc. Sports Nutr. 2015, 12, 28. [Google Scholar] [CrossRef] [PubMed]
- Mizunoya, W.; Iwamoto, Y.; Shirouchi, B.; Sato, M.; Komiya, Y.; Razin, F.R.; Tatsumi, R.; Sato, Y.; Nakamura, M.; Ikeuchi, Y. Dietary fat influences the expression of contractile and metabolic genes in rat skeletal muscle. PLoS ONE 2013, 8, e80152. [Google Scholar] [CrossRef] [PubMed]
- Leenders, M.; Verdijk, L.B.; van der Hoeven, L.; van Kranenburg, J.; Nilwik, R.; van Loon, L.J. Elderly men and women benefit equally from prolonged resistance-type exercise training. J. Gerontol. A Biol. Sci. Med. Sci. 2013, 68, 769–779. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Lee, M.; Park, Y. Synergistic attenuation of ovariectomy-induced bone loss by combined use of fish oil and 17beta-oestradiol. Br. J. Nutr. 2017, 117, 479–489. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.I.; Mittendorfer, B. Sexual dimorphism in skeletal muscle protein turnover. J. Appl. Physiol. 2016, 120, 674–682. [Google Scholar] [CrossRef] [PubMed]
- Egerman, M.A.; Cadena, S.M.; Gilbert, J.A.; Meyer, A.; Nelson, H.N.; Swalley, S.E.; Mallozzi, C.; Jacobi, C.; Jennings, L.L.; Clay, I.; et al. Gdf11 increases with age and inhibits skeletal muscle regeneration. Cell Metab. 2015, 22, 164–174. [Google Scholar] [CrossRef] [PubMed]
- Conboy, I.M.; Conboy, M.J.; Wagers, A.J.; Girma, E.R.; Weissman, I.L.; Rando, T.A. Rejuvenation of aged progenitor cells by exposure to a young systemic environment. Nature 2005, 433, 760–764. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C.; Bosco, N.; Bourdet-Sicard, R.; Capuron, L.; Delzenne, N.; Dore, J.; Franceschi, C.; Lehtinen, M.J.; Recker, T.; Salvioli, S.; et al. Health relevance of the modification of low grade inflammation in ageing (inflammageing) and the role of nutrition. Ageing Res. Rev. 2017, 40, 95–119. [Google Scholar] [CrossRef] [PubMed]
- Apolinario, L.M.; De Carvalho, S.C.; Santo Neto, H.; Marques, M.J. Long-term therapy with omega-3 ameliorates myonecrosis and benefits skeletal muscle regeneration in mdx mice. Anat. Rec. 2015, 298, 1589–1596. [Google Scholar] [CrossRef] [PubMed]
- Mackey, A.L.; Rasmussen, L.K.; Kadi, F.; Schjerling, P.; Helmark, I.C.; Ponsot, E.; Aagaard, P.; Durigan, J.L.; Kjaer, M. Activation of satellite cells and the regeneration of human skeletal muscle are expedited by ingestion of nonsteroidal anti-inflammatory medication. FASEB J. 2016, 30, 2266–2281. [Google Scholar] [CrossRef] [PubMed]
- Landi, F.; Marzetti, E.; Liperoti, R.; Pahor, M.; Russo, A.; Martone, A.M.; Colloca, G.; Capoluongo, E.; Bernabei, R. Nonsteroidal anti-inflammatory drug (NSAID) use and sarcopenia in older people: Results from the ilsirente study. J. Am. Med. Direct. Assoc. 2013, 14, 626. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Liang, X.; Shan, T.; Jiang, Q.; Deng, C.; Zheng, R.; Kuang, S. Mtor is necessary for proper satellite cell activity and skeletal muscle regeneration. Biochem. Biophys. Res. Commun. 2015, 463, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Kamolrat, T.; Gray, S.R. The effect of eicosapentaenoic and docosahexaenoic acid on protein synthesis and breakdown in murine C2C12 myotubes. Biochem. Biophys. Res. Commun. 2013, 432, 593–598. [Google Scholar] [CrossRef] [PubMed]
- Bodine, S.C.; Stitt, T.N.; Gonzalez, M.; Kline, W.O.; Stover, G.L.; Bauerlein, R.; Zlotchenko, E.; Scrimgeour, A.; Lawrence, J.C.; Glass, D.J.; et al. Akt/mTOR pathway is a crucial regulator of skeletal muscle hypertrophy and can prevent muscle atrophy in vivo. Nat. Cell Biol. 2001, 3, 1014–1019. [Google Scholar] [CrossRef] [PubMed]
- Martin, T.D.; Dennis, M.D.; Gordon, B.S.; Kimball, S.R.; Jefferson, L.S. Mtorc1 and jnk coordinate phosphorylation of the p70s6k1 autoinhibitory domain in skeletal muscle following functional overloading. Am. J. Physiol. Endocrinol. Metab. 2014, 306, E1397–E1405. [Google Scholar] [CrossRef] [PubMed]
- Bennett, A.M.; Tonks, N.K. Regulation of distinct stages of skeletal muscle differentiation by mitogen-activated protein kinases. Science 1997, 278, 1288–1291. [Google Scholar] [CrossRef] [PubMed]
- Kramer, H.F.; Goodyear, L.J. Exercise, mapk, and nf-kappab signaling in skeletal muscle. J. Appl. Physiol. 2007, 103, 388–395. [Google Scholar] [CrossRef] [PubMed]
- Volonte, D.; Liu, Y.; Galbiati, F. The modulation of caveolin-1 expression controls satellite cell activation during muscle repair. FASEB J. 2005, 19, 237–239. [Google Scholar] [CrossRef] [PubMed]
- Kook, S.H.; Son, Y.O.; Choi, K.C.; Lee, H.J.; Chung, W.T.; Hwang, I.H.; Lee, J.C. Cyclic mechanical stress suppresses myogenic differentiation of adult bovine satellite cells through activation of extracellular signal-regulated kinase. Mol. Cell. Biochem. 2008, 309, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.Y.; Yao, K.; Bode, A.M.; Bergen, H.R., III; Madden, B.J.; Oh, S.M.; Ermakova, S.; Kang, B.S.; Choi, H.S.; Shim, J.H.; et al. RSK2 mediates muscle cell differentiation through regulation of NFAT3. J. Biol. Chem. 2007, 282, 8380–8392. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Johnson, S.E. ERK2 is required for efficient terminal differentiation of skeletal myoblasts. Biochem. Biophys. Res. Commun. 2006, 345, 1425–1433. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Zheng, Y.; Zhang, Y.; Zhao, J.; Chang, F.; Lu, T.; Zhang, R.; Li, Q.; Hu, X.; Li, N. Different effects of omega-3 fatty acids on the cell cycle in c2c12 myoblast proliferation. Mol. Cell. Biochem. 2012, 367, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Day, K.; Shefer, G.; Shearer, A.; Yablonka-Reuveni, Z. The depletion of skeletal muscle satellite cells with age is concomitant with reduced capacity of single progenitors to produce reserve progeny. Dev. Biol. 2010, 340, 330–343. [Google Scholar] [CrossRef] [PubMed]
- He, W.A.; Berardi, E.; Cardillo, V.M.; Acharyya, S.; Aulino, P.; Thomas-Ahner, J.; Wang, J.; Bloomston, M.; Muscarella, P.; Nau, P.; et al. Nf-kappab-mediated pax7 dysregulation in the muscle microenvironment promotes cancer cachexia. J. Clin. Investig. 2013, 123, 4821–4835. [Google Scholar] [CrossRef] [PubMed]
- Murphy, L.O.; Blenis, J. Mapk signal specificity: The right place at the right time. Trends Biochem. Sci. 2006, 31, 268–275. [Google Scholar] [CrossRef] [PubMed]
- Saini, A.; Sharples, A.P.; Al-Shanti, N.; Stewart, C.E. Omega-3 fatty acid epa improves regenerative capacity of mouse skeletal muscle cells exposed to saturated fat and inflammation. Biogerontology 2017, 18, 109–129. [Google Scholar] [CrossRef] [PubMed]
- Gillespie, M.A.; Le Grand, F.; Scimè, A.; Kuang, S.; von Maltzahn, J.; Seale, V.; Cuenda, A.; Ranish, J.A.; Rudnicki, M.A. P38-γ–dependent gene silencing restricts entry into the myogenic differentiation program. J. Cell Biol. 2009, 187, 991–1005. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Yang, R.; Martinod, K.; Kasuga, K.; Pillai, P.S.; Porter, T.F.; Oh, S.F.; Spite, M. Maresins: Novel macrophage mediators with potent antiinflammatory and proresolving actions. J. Exp. Med. 2009, 206, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Gura, K.M.; Kim, S.; Arsenault, D.A.; Bistrian, B.R.; Puder, M. Current clinical applications of omega-6 and omega-3 fatty acids. Nutr. Clin. Pract. 2006, 21, 323–341. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Zhou, Y.; Hu, X.; Peng, X.; Wei, H.; Peng, J.; Jiang, S. Activation of PPARγ2 by ppargamma1 through a functional ppre in transdifferentiation of myoblasts to adipocytes induced by EPA. Cell Cycle 2015, 14, 1830–1841. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Tachibana, H.; Morinaga, Y.; Fujimura, Y.; Yamada, K. Modulation of proliferation and differentiation of c2c12 skeletal muscle cells by fatty acids. Life Sci. 2009, 84, 415–420. [Google Scholar] [CrossRef] [PubMed]
- Smith, H.J.; Lorite, M.J.; Tisdale, M.J. Effect of a cancer cachectic factor on protein synthesis/degradation in murine C2C12 myoblasts: Modulation by eicosapentaenoic acid. Cancer Res. 1999, 59, 5507–5513. [Google Scholar] [PubMed]
- Lee, M.S.; Kim, I.H.; Kim, Y. Effects of eicosapentaenoic acid and docosahexaenoic acid on uncoupling protein 3 gene expression in C(2)C(12) muscle cells. Nutrients 2013, 5, 1660–1671. [Google Scholar] [CrossRef] [PubMed]
- Woodworth-Hobbs, M.E.; Hudson, M.B.; Rahnert, J.A.; Zheng, B.; Franch, H.A.; Price, S.R. Docosahexaenoic acid prevents palmitate-induced activation of proteolytic systems in c2c12 myotubes. J. Nutr. Biochem. 2014, 25, 868–874. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.C.; Chen, P.Y.; Wu, Y.L.; Chen, C.W.; Chen, H.W.; Lii, C.K.; Sun, H.L.; Liu, K.L. Long-chain polyunsaturated fatty acids amend palmitate-induced inflammation and insulin resistance in mouse C2C12 myotubes. Food Funct. 2016, 7, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Lalia, A.Z.; Dasari, S.; Robinson, M.M.; Abid, H.; Morse, D.M.; Klaus, K.A.; Lanza, I.R. Influence of omega-3 fatty acids on skeletal muscle protein metabolism and mitochondrial bioenergetics in older adults. Aging 2017, 9, 1096–1115. [Google Scholar] [CrossRef] [PubMed]
- Schiaffino, S.; Bormioli, S.P.; Aloisi, M. The fate of newly formed satellite cells during compensatory muscle hypertrophy. Virchows Arch. B Cell Pathol. 1976, 21, 113–118. [Google Scholar] [PubMed]
- Moss, F.P.; Leblond, C.P. Nature of dividing nuclei in skeletal muscle of growing rats. J. Cell Biol. 1970, 44, 459–461. [Google Scholar] [CrossRef] [PubMed]
- Yoshino, J.; Smith, G.I.; Kelly, S.C.; Julliand, S.; Reeds, D.N.; Mittendorfer, B. Effect of dietary n-3 pufa supplementation on the muscle transcriptome in older adults. Physiol. Rep. 2016, 4, e12785. [Google Scholar] [CrossRef] [PubMed]
- Pawar, A.; Jump, D.B. Unsaturated fatty acid regulation of peroxisome proliferator-activated receptor alpha activity in rat primary hepatocytes. J. Biol. Chem. 2003, 278, 35931–35939. [Google Scholar] [CrossRef] [PubMed]
- Kishida, E.; Tajiri, M.; Masuzawa, Y. Docosahexaenoic acid enrichment can reduce l929 cell necrosis induced by tumor necrosis factor. Biochim. Biophys. Acta 2006, 1761, 454–462. [Google Scholar] [CrossRef] [PubMed]
- Lo, C.J.; Chiu, K.C.; Fu, M.; Chu, A.; Helton, S. Fish oil modulates macrophage p44/p42 mitogen-activated protein kinase activity induced by lipopolysaccharide. JPEN J. Parenter Enter. Nutr. 2000, 24, 159–163. [Google Scholar] [CrossRef] [PubMed]
- Weldon, S.M.; Mullen, A.C.; Loscher, C.E.; Hurley, L.A.; Roche, H.M. Docosahexaenoic acid induces an anti-inflammatory profile in lipopolysaccharide-stimulated human thp-1 macrophages more effectively than eicosapentaenoic acid. J. Nutr. Biochem. 2007, 18, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Mullen, A.; Loscher, C.E.; Roche, H.M. Anti-inflammatory effects of epa and dha are dependent upon time and dose-response elements associated with lps stimulation in thp-1-derived macrophages. J. Nutr. Biochem. 2010, 21, 444–450. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Palmitate (PAL) and Tumour Necrosis Factor-alpha (TNF-α) elicit lipotoxic and cytotoxic deleterious effects on satellite cells during various stages of myogenesis. N-3 PUFAs inhibit the negative effects of PAL and TNF-α by activating anti-inflammatory pathways within the cell thereby promoting differentiation. In isolation, it is currently unknown whether n-3 PUFAs are capable of modulating the expressions of key myogenic transcriptional regulatory factors (MRFs) Pax7, MyoD, and Myogenin that regulate myogenesis. Solid line: represents established role/pathway that n-3 PUFAs modulate during myogenic differentiation. Dotted line: limited or evidence no supporting the role of Omega-3 polyunsaturated fatty acids (n-3 PUFAs) during myogenesis. Solid line: substantial evidence for n-3 PUFAs effecting myogenesis via various pathways.
Figure 1.
Palmitate (PAL) and Tumour Necrosis Factor-alpha (TNF-α) elicit lipotoxic and cytotoxic deleterious effects on satellite cells during various stages of myogenesis. N-3 PUFAs inhibit the negative effects of PAL and TNF-α by activating anti-inflammatory pathways within the cell thereby promoting differentiation. In isolation, it is currently unknown whether n-3 PUFAs are capable of modulating the expressions of key myogenic transcriptional regulatory factors (MRFs) Pax7, MyoD, and Myogenin that regulate myogenesis. Solid line: represents established role/pathway that n-3 PUFAs modulate during myogenic differentiation. Dotted line: limited or evidence no supporting the role of Omega-3 polyunsaturated fatty acids (n-3 PUFAs) during myogenesis. Solid line: substantial evidence for n-3 PUFAs effecting myogenesis via various pathways.
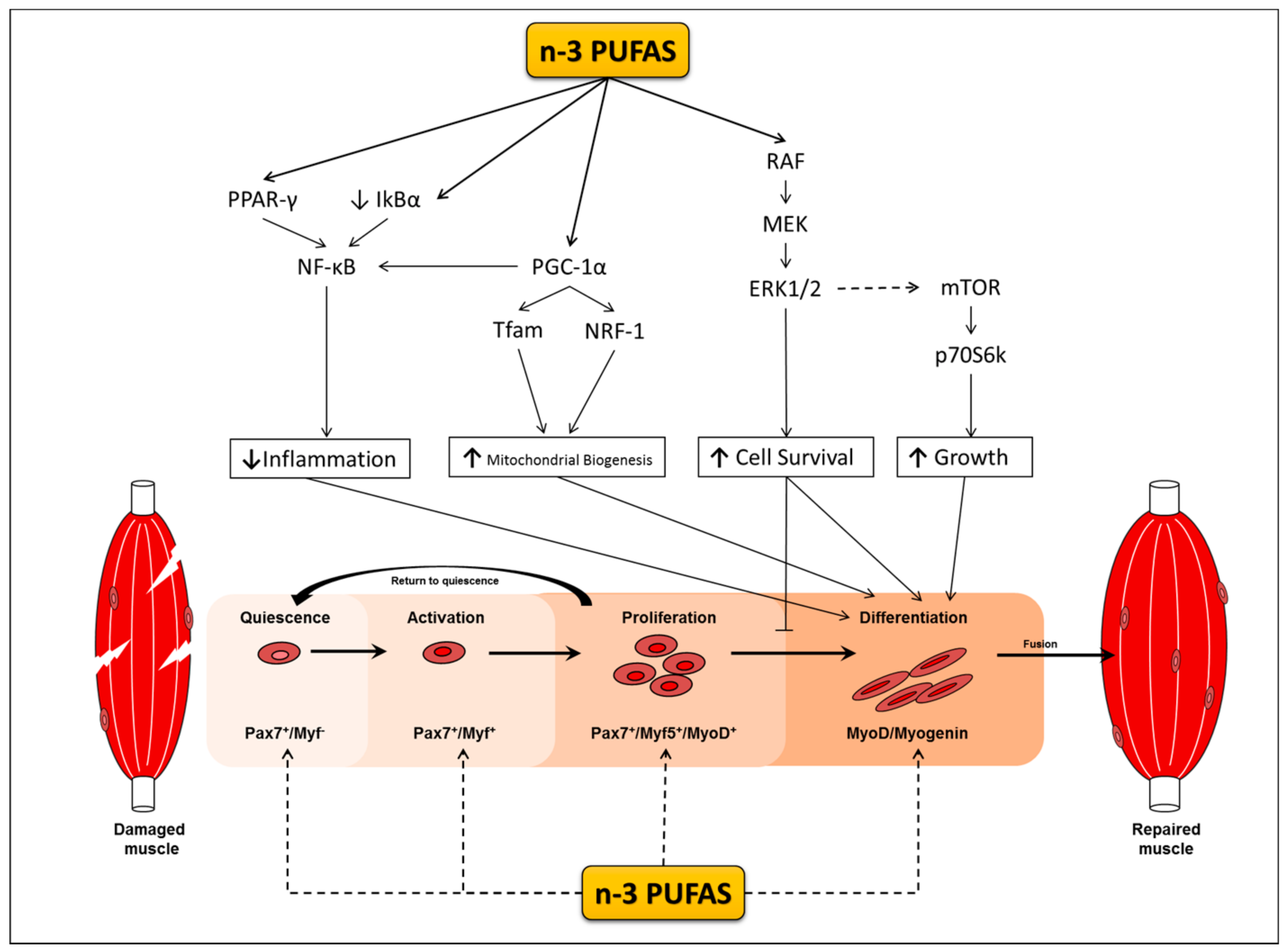

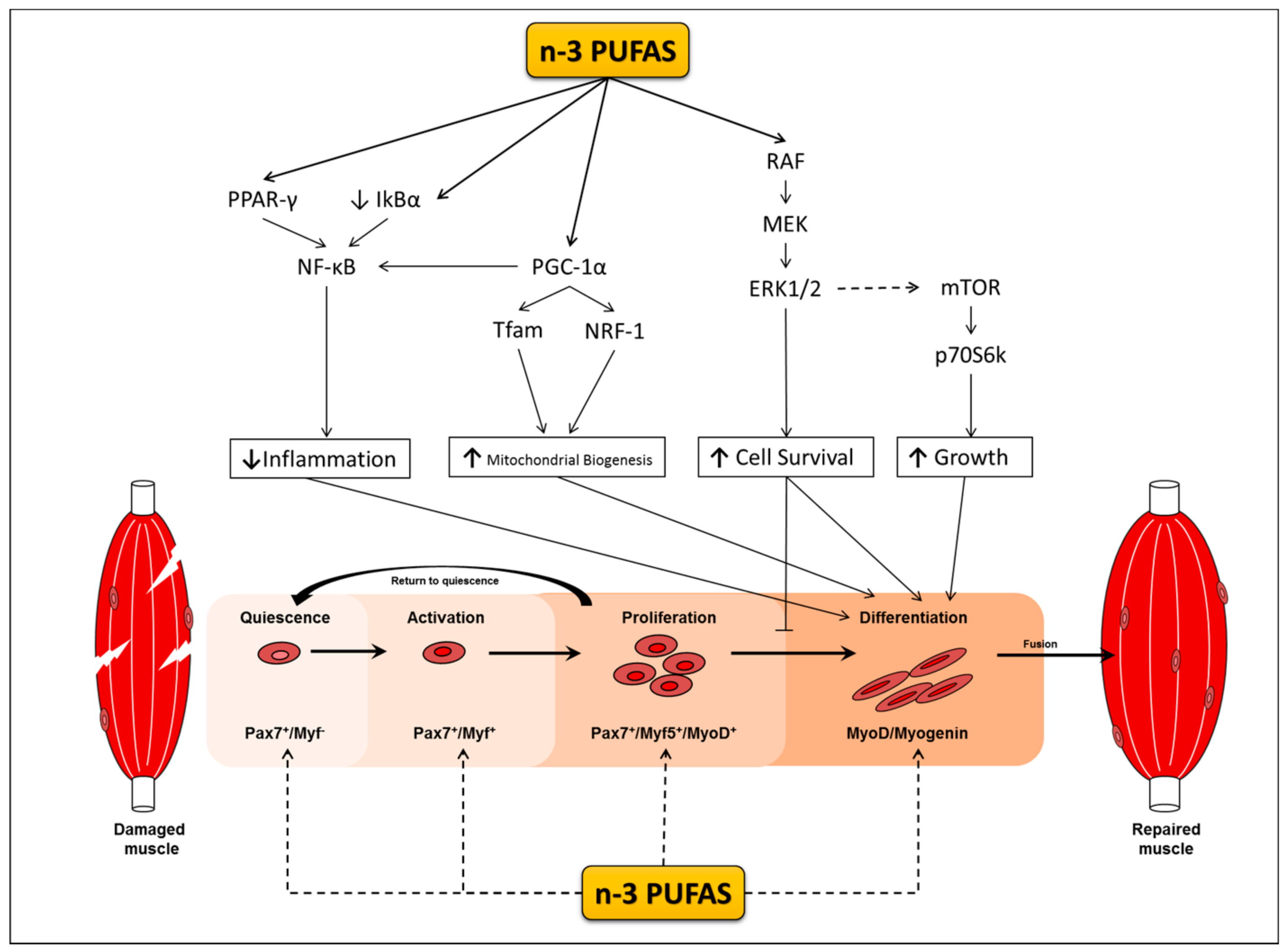{kind=link}
Table 1.
Summary of studies investigating n-3 PUFAs in proliferating and differentiating myoblasts.
| Source | Treatment | Cell Line | Outcome |
|---|---|---|---|
| Smith, et al. [141] | 50 μM EPA 1–10 nM PIF | C2C12 | EPA ↓ protein degradation in PIF treated myoblasts |
| Magee, et al. [87] | 50 μM EPA 20 ng/mL TNF-α | C2C12 | EPA ↓ the negative effects TNF-α via ↓ NF-κB expression. EPA ↑ PPARγ expression |
| Lee, et al. [140] | 1–10 μM DHA 48 h 1–10 μM EPA 48 h | C2C12 | DHA ↑ cell proliferation at 10µM concentration only. EPA has no effect on proliferation between 0 and10 µM. EPA and DHA were no investigated in the context of differentiation in this study |
| Magee, et al. [88] | 50 μM EPA 24 h 20 TNF-α ng/mL 24 h | C2C12 | EPA ↓ the negative effects TNFα has on cell death, apoptosis and myotube formation by ↑ PPARγ expression via the NfKB signaling pathway. |
| Peng, et al. [131] | 10–100 μM DHA 24 h 10–100 μM EPA 24 h | C2C12 | EPA and DHA ↓ cell proliferation in dose dependent manner DHA and EPA ↓ protein levels of Cyclin D and Cyclin E and CDK2. EPA and DHA ↓ ERK1/2 protein expression |
| Briolay, et al. [79] | 20 μM DHA 72 h 20 μM EPA 72 h | L6 | Treating cells with EPA and DHA ↑ fusion index. This is accompanied by ↑ in p70S6K1, and membrane bund protein caveolin-3 that is associated with fusion. |
| Luo, et al. [139] | 400–600 μM EPA 10 days | C2C12 | EPA in high doses of 400–600 µM induces transdifferentiation of myoblasts to adipocytes by ↓ Wnt/β-catanin signalling through PPARγ |
| Saini, et al. [135] | 50 μM EPA 36 h & 72 h 750 μM PAL 36 h & 72 h | C2 | EPA ↓ palmitate induced cell death via affecting the MAPK pathway. EPA can protect cells from MAPK induced cell death and myogenic suppression but not against of Id3/caspase induced apoptosis EPA ↑ MyoD |
Abbreviations: Eicosapentaenoic acid (EPA), Docosahexanenoic acid (DHA) Alpha linolenic acid (ALA), Proteolysis inducing factor (PIF), tumour necrosis factor-alpha (TNF- α), nuclear factor kappa-light-chain-enhancer of activated B cells (NF-kB), Peroxisome proliferator activated-receptor gamma (PPARγ), Cyclin dependany kinase 2 (CDK2), Palmitate (PAL). ↑/↓ indicates the trend.
Table 2.
Summary of studies investigating the effects of PUFAs in full differentiated myotubes.
| Publication | Treatment | Outcome |
|---|---|---|
| Bryner, et al. [96] | 100 μM DHA 500 μM PAL | DHA attenuates ↓ tube size by PAL |
| Wang, et.al [69] | 300–700 μM DHA 300–700 μM EPA | EPA and DHA ↓ total protein degradation in a dose response manner DHA and EPA ↓ protein degradation by regulating the IkBα/NFκB signalling DHA is more effective in preventing degradation than EPA |
| Kamolrat and Gray [122] | 50 μM DHA 50 μM EPA | EPA ↑ MPS and ↓ MPB DHA has no effect on MPS or MPB at 50 μM |
| Lee, et al. [142] | 1–50 μM DHA, 1–50 μM EPA | DHA and EPA both ↑ UCP3 AICAR synergistically ↑ UCP3 activity when co-treated with DHA or EPA |
| Woodworth-Hobbs, et al. [143] | 100 μM DHA 500 μM PAL | DHA ↓ proteolysis by palmitate in a time dependent manner and counteracts the effects of PAL by restoring Akt activity, ↓ FOXO activity ↓ and atrogin expression. |
| Lee, et al. [91] | 1–50 μM DHA 1–50 μM EPA | EPA and DHA ↑ PGC-1α, NRF-1 and Tfam gene expression and ↑ PGC-1α promoter activity |
| Chen, et al. [144] | 50 μM DHA 50 μM EPA, 50 μM AA 750 μM PAL | The negative effects of PAL on AMPK phosphorylation GLUT4 mRNA expression and basal glucose uptake were ↓ by AA, DHA and EPA. The expression of genes associated with protein degradation was ↓ by EPA, DHA and AA |
| Pinel, et al. [95] | 50 μM DHA 50 μM EPA 50 μM ALA 500 μM PAL | EPA and DHA ↑ membrane fluidity which may improve glucose uptake.EPA and DHA ↑ key gene involved in β-oxidation. |
Abbreviations: Eicosapentaenoic acid (EPA), Docosahexanenoic acid (DHA) ), tumour necrosis factor-alpha (TNF-α), nuclear factor kappa-light-chain-enhancer of activated B cells (NF-kB), Peroxisome proliferator activated-receptor gamma (PPARγ), Palmitate (PAL), Muscle protein synthesis (MPS) Muscle protein breakdown (MPB), Uncoupling protein-3 (UCP3), 5-Aminoimidazole-4-carboxamide ribonucleotide (AICAR), AMP-dependent protein kinase (AMPK), Forkhead box O (FOXO), Peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PGC-1α). ↑/↓ indicates the trend.
Table 3.
Summary of human studies that have investigated the effects of n-3 PUFA supplementation combined with resistance exercise and/or protein supplementation on muscle anabolism and muscle function.
Table 3.
Summary of human studies that have investigated the effects of n-3 PUFA supplementation combined with resistance exercise and/or protein supplementation on muscle anabolism and muscle function.
| Publication | Dose | Duration | Age | Sex | N | REX | Protein | Outcomes |
|---|---|---|---|---|---|---|---|---|
| Smith, et al. [33] | 1.86 g/day EPA 1.5 g/day DHA | 8 weeks | 39.7 ± 1.7 | M | 5 | - | Insulin clamp Amino Acid clamp | Hyperaminoacidemia-hyperinsulinemia induced MPS ↑ after supplementation and ↑ p-mTORSer2448 p-p70S6k1Thr389. |
| F | 4 | |||||||
| Smith, et al. [47] | 1.86 g/day EPA 1.5 g/day DHA | 8 weeks | 71 ± 1 | M | 5 | - | Insulin clamp Amino Acid clamp | Hyperaminoacidemia-hyperinsulinemia induced MPS ↑ after supplementation and ↑ p-mTORSer2448 p-p70S6k1Thr389. |
| F | 3 | |||||||
| Rodacki, et al. [34] | 0.4 g/day EPA 0.3 g/day DHA | 90 days | 64 ± 1.4 | F | 45 | 3x/week | - | Fish Oil supplementation and strength training results in ↑ improvements in peak torque and rate of torque development than strength training alone. |
| 150 days | 64 ± 1.4 | F | 45 | 3x/week | ||||
| McGlory, et al. [106] | 3.5 g/ay EPA 0.9 g/day DHA | 4 weeks | 21.3 ± 3 | M | 10 | - | - | EPA and DHA in the blood ↑ from week 0 to 2 and ↔ from 2 to 4 weeks. Levels of EPA and DHA in skeletal muscle are ↑ from 0 to 2 weeks and further ↑ 2 to 4 weeks. |
| Smith, et al. [32] | 1.86 g/day EPA 1.5 g/day DHA | 26 weeks | 68 ± 5 | M | 10 | - | - | ↑ thigh muscle volume, ↑ hand grip strength and ↑1RM |
| F | 19 | |||||||
| McGlory, et al. [46] | 3.5 g/day EPA 0.9 g/day DHA 0.1 g/day DPA | 8 weeks | 24 ± 0 | M | 9 | Acute bout | 30 g Whey | Compared to placebo no change in myofibrillar MPS following an acute bout of REX and protein ingestion ↓ p70S6K1 and Akt anabolic kinases following REX and protein ingestion. |
| Da Boit, et al. [78] | 2.1 g/day EPA 0.6 g/day DHA | 18 weeks | 70.6 ± 4.5 | M | 27 | 2x/week | - | ↑ Maximal isometric torque and ↑ muscle quality in women after exercise training in n-3 PUFA group than in the placebo group, with no such differences in men |
| 70.7 ± 3.3 | F | 23 | ||||||
| Lalia, et al. [145] | 0.675 g/day EPA 0.3 g/day DHA | 16 weeks | 27 ± 5 | M/F | 12 | Acute bout | - | Myofibrillar MPS ↑ in both young and old after exercise and supplementation whereas mitoMPS and sarcoMPS ↑ in the older participants |
| 76 ± 5 | M/F | 12 |
Abbreviations: Eicosapentaenoic acid (EPA), Docosahexanenoic acid (DHA), Docosapentanoeic acid (DPA), Resistance exercise (REX), Muscle protein synthesis (MPS) Muscle protein breakdown (MPB), 1 Repitition Max (1RM),Male (M), Female (F). ↑/↓ indicates the trend.
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tachtsis, B.; Camera, D.; Lacham-Kaplan, O. Potential Roles of n-3 PUFAs during Skeletal Muscle Growth and Regeneration. Nutrients 2018, 10, 309. https://doi.org/10.3390/nu10030309
AMA Style
Tachtsis B, Camera D, Lacham-Kaplan O. Potential Roles of n-3 PUFAs during Skeletal Muscle Growth and Regeneration. Nutrients. 2018; 10(3):309. https://doi.org/10.3390/nu10030309
Chicago/Turabian StyleTachtsis, Bill, Donny Camera, and Orly Lacham-Kaplan. 2018. "Potential Roles of n-3 PUFAs during Skeletal Muscle Growth and Regeneration" Nutrients 10, no. 3: 309. https://doi.org/10.3390/nu10030309
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.