Effect of UV Radiation and Salt Stress on the Accumulation of Economically Relevant Secondary Metabolites in Bell Pepper Plants

 ,
,
Abstract
1. Introduction
1.1. Green Biomass as a Source of Valuable Chemicals
1.2. Plant Stress as a Measure to Increase Leaf Secondary Metabolite Content
1.3. Non-Invasive Monitoring of Secondary Metabolites in Plant Leaves
1.4. Cynaroside and Graveobioside A
2. Material and Methods
2.1. Plant Material and Growth Conditions
2.2. Non-Destructive Recordings
2.3. Plant Harvest
2.4. Leaf Sample Preparation and Laboratory Analysis
2.5. Data Analysis and Statistics
3. Results
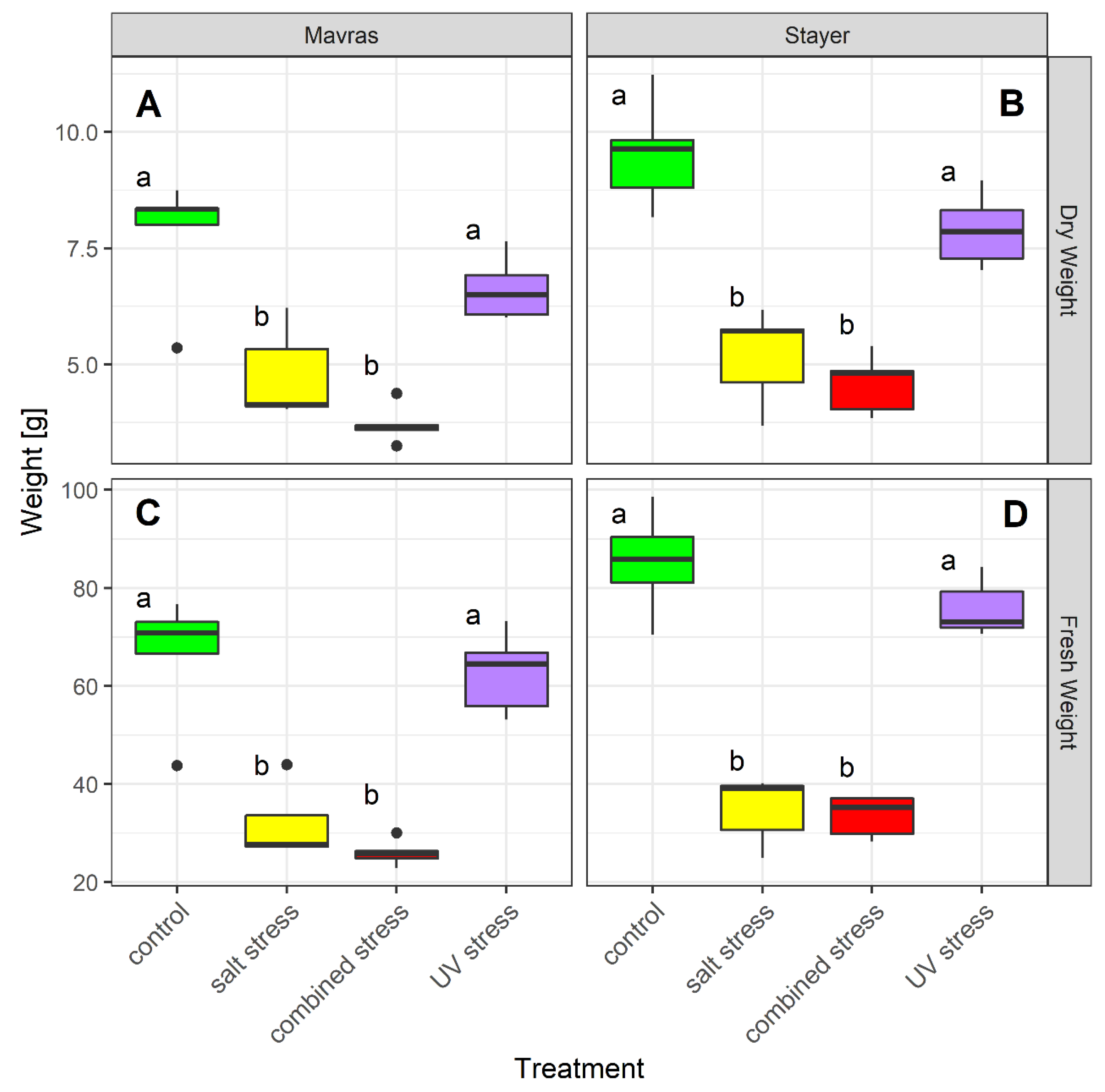
3.1. Stress-Related Effect Varies Among Secondary Metabolites and Cultivars
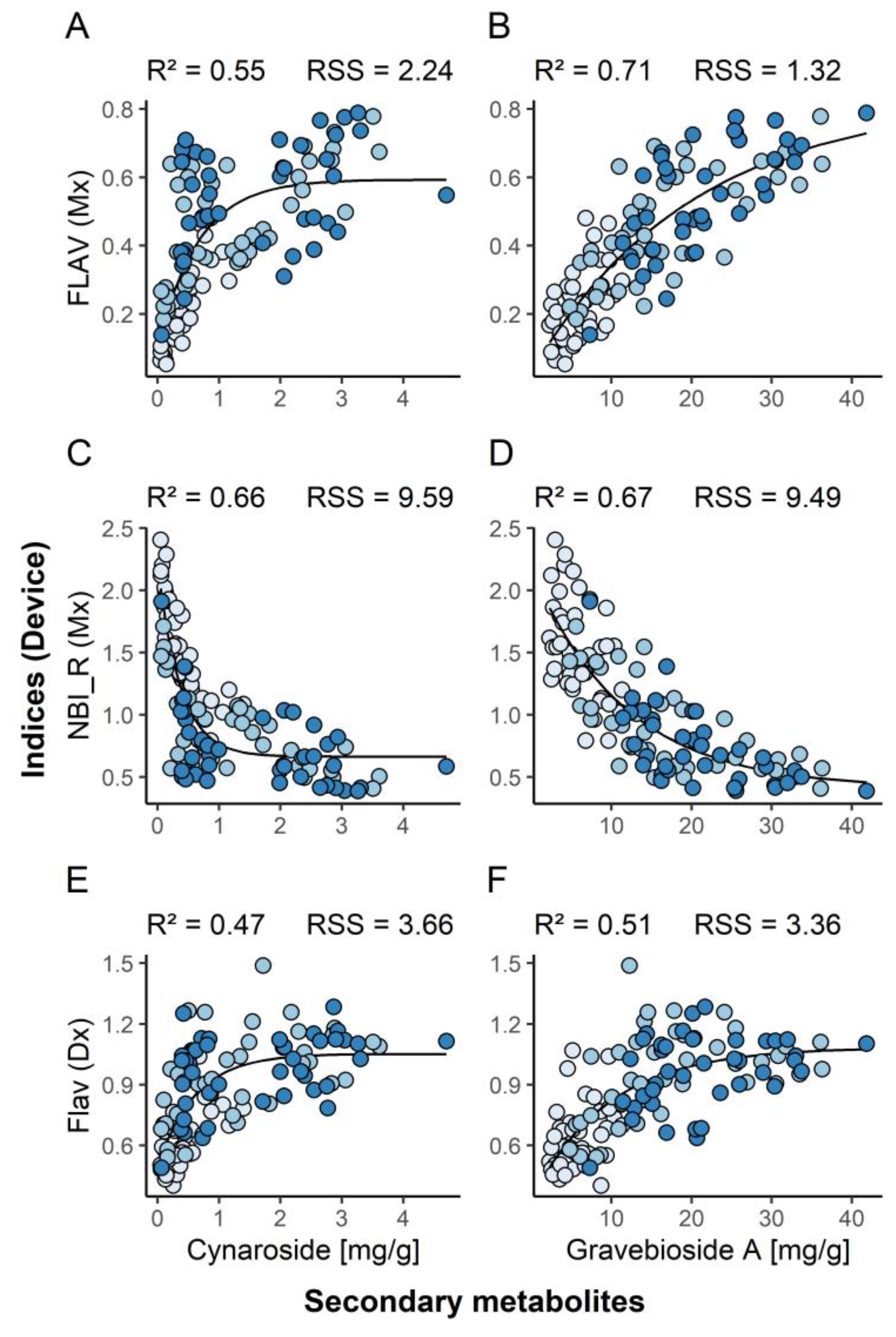
3.2. Non-Invasive Monitoring of Secondary Metabolites
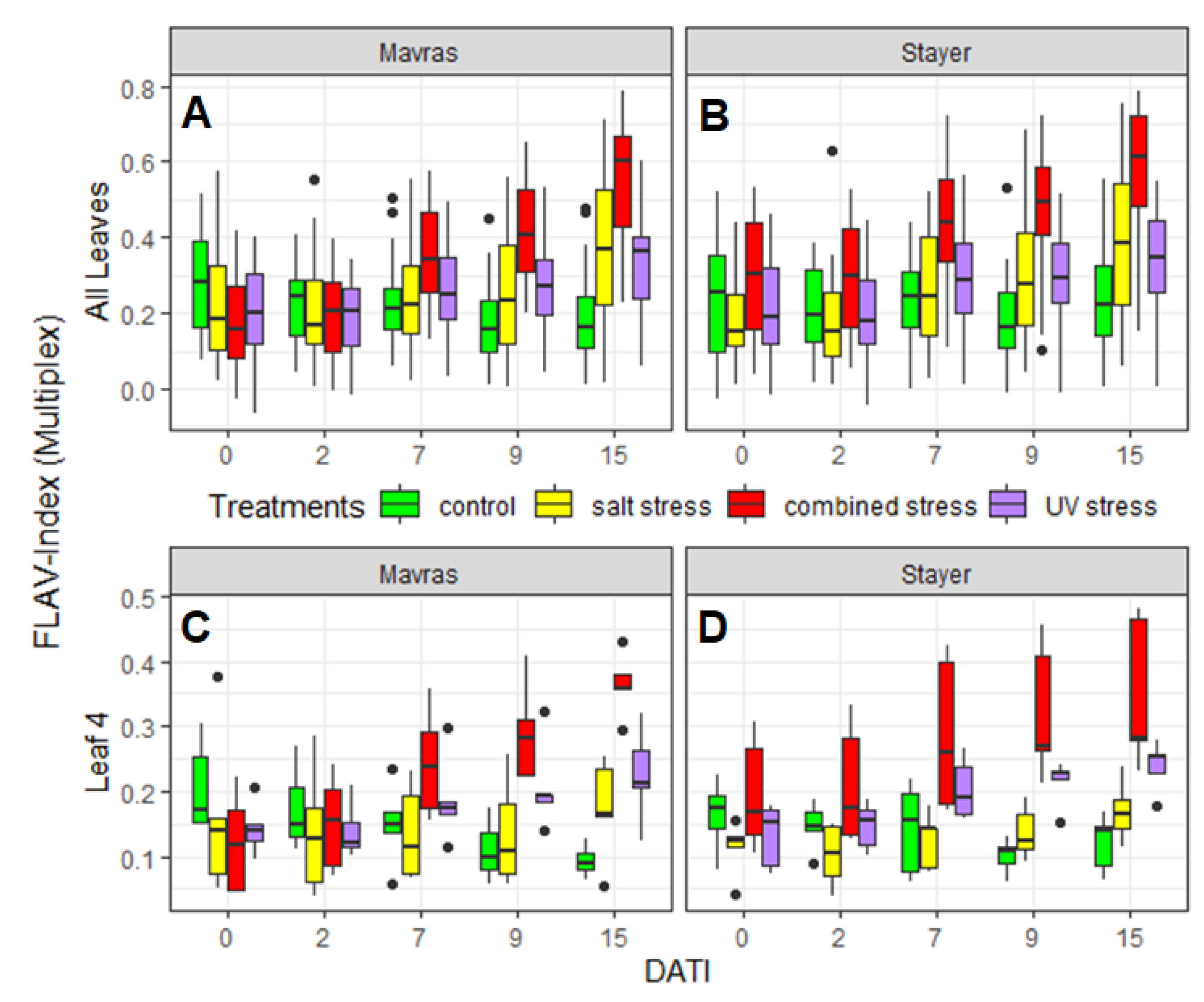
3.3. Spatial and Temporal Development of Secondary Metabolite Contents
4. Discussion
4.1. Stress-Related Effect Varies According to Secondary Metabolites and Cultivars
4.2. Non-Invasive Monitoring
4.3. Insights in Spatial and Temporal Accumulation of Secondary Metabolites
4.4. Implications and Future Challenges
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| DATI | Days after treatment inception |
| HPLC | High performance liquid chromatography |
| ROS | Reactive oxygen species |
| SM | Secondary metabolite |
References
- Wormit, A.; Bröring, S.; Carraresi, L.; Junker, L.; Jupke, A.; Lück, M.; Noga, G.; Reimer, J.J.; Schmittgen, S.; Thiele, B.; et al. TaReCa–Kaskadennutzung gartenbaulicher Biomasse für eine ressourceneffiziente Produktion von wertvollen bioaktiven Substanzen. Julius-Kühn-Arch. 2018, 460, 10–13. [Google Scholar]
- Junker-Frohn, L.V.; Lück, M.; Schmittgen, S.; Wensing, J.; Carraresi, L.; Thiele, B.; Groher, T.; Reimer, J.J.; Bröring, S.; Noga, G.; et al. Tomato’s Green Gold: Bioeconomy Potential of Residual Tomato Leaf Biomass as a Novel Source for the Secondary Metabolite Rutin. ACS Omega 2019, 4, 19071–19080. [Google Scholar] [CrossRef]
- European Commission A sustainable bioeconomy for Europe strengthening the connection between economy, society and the environment: updated bioeconomy strategy; European Commission: Brussels, Belgium, 2018; ISBN 978-92-79-94144-3.
- Carvalho Lemos, V.; Reimer, J.J.; Wormit, A. Color for Life: Biosynthesis and Distribution of Phenolic Compounds in Pepper (Capsicum annuum). Agriculture 2019, 9, 81. [Google Scholar] [CrossRef]
- Taylor, M.A.; Fraser, P.D. Solanesol: Added value from Solanaceous waste. Phytochemistry 2011, 72, 1323–1327. [Google Scholar] [CrossRef] [PubMed]
- Junker, L.V. Induktion des Sekundärmetabolismus in Tomatenpflanzen zur alternativen Verwendung der Blattbiomasse. DGG-Proc. Ger. Soc. Hortic. Sci. DGG 2017, 7, 1–5. [Google Scholar]
- Groher, T.; Schmittgen, S.; Noga, G.; Hunsche, M. Limitation of mineral supply as tool for the induction of secondary metabolites accumulation in tomato leaves. Plant Physiol. Biochem. 2018, 130, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Fini, A.; Brunetti, C.; Di Ferdinando, M.; Ferrini, F.; Tattini, M. Stress-induced flavonoid biosynthesis and the antioxidant machinery of plants. Plant Signal. Behav. 2011, 6, 709–711. [Google Scholar] [CrossRef]
- Müller, V.; Lankes, C.; Schmitz-Eiberger, M.; Noga, G.; Hunsche, M. Estimation of flavonoid and centelloside accumulation in leaves of Centella asiatica L. Urban by multiparametric fluorescence measurements. Environ. Exp. Bot. 2013, 93, 27–34. [Google Scholar] [CrossRef]
- Hoffmann, A.M.; Noga, G.; Hunsche, M. High blue light improves acclimation and photosynthetic recovery of pepper plants exposed to UV stress. Environ. Exp. Bot. 2015, 109, 254–263. [Google Scholar] [CrossRef]
- Escobar-Bravo, R.; Klinkhamer, P.G.L.; Leiss, K.A. Interactive Effects of UV-B Light with Abiotic Factors on Plant Growth and Chemistry, and Their Consequences for Defense against Arthropod Herbivores. Front. Plant Sci. 2017, 8, 278. [Google Scholar] [CrossRef]
- Acosta-Motos, J.; Ortuño, M.; Bernal-Vicente, A.; Diaz-Vivancos, P.; Sanchez-Blanco, M.; Hernandez, J. Plant Responses to Salt Stress: Adaptive Mechanisms. Agronomy 2017, 7, 18. [Google Scholar] [CrossRef]
- Atzori, G.; Mancuso, S.; Masi, E. Seawater potential use in soilless culture: A review. Sci. Hortic. 2019, 249, 199–207. [Google Scholar] [CrossRef]
- Hand, M.J.; Taffouo, V.D.; Nouck, A.E.; Nyemene, K.P.J.; Tonfack, B.; Meguekam, T.L.; Youmbi, E. Effects of Salt Stress on Plant Growth, Nutrient Partitioning, Chlorophyll Content, Leaf Relative Water Content, Accumulation of Osmolytes and Antioxidant Compounds in Pepper (Capsicum annuum L.) Cultivars. Not. Bot. Horti Agrobot. Cluj-Napoca 2017, 45, 481–490. [Google Scholar] [CrossRef]
- Agati, G.; Biricolti, S.; Guidi, L.; Ferrini, F.; Fini, A.; Tattini, M. The biosynthesis of flavonoids is enhanced similarly by UV radiation and root zone salinity in L. vulgare leaves. J. Plant Physiol. 2011, 168, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Gruda, N.S. Increasing Sustainability of Growing Media Constituents and Stand-Alone Substrates in Soilless Culture Systems. Agronomy 2019, 9, 298. [Google Scholar] [CrossRef]
- Cerovic, Z.G.; Samson, G.; Morales, F.; Tremblay, N.; Moya, I. Ultraviolet-induced fluorescence for plant monitoring: present state and prospects. Agronomie 1999, 19, 543–578. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Keydan, G.P.; Merzlyak, M.N. Three-band model for noninvasive estimation of chlorophyll, carotenoids, and anthocyanin contents in higher plant leaves. Geophys. Res. Lett. 2006, 33, L11402. [Google Scholar] [CrossRef]
- Groher, T.; Schmittgen, S.; Fiebig, A.; Noga, G.; Hunsche, M. Suitability of fluorescence indices for the estimation of fruit maturity compounds in tomato fruits. J. Sci. Food Agric. 2018, 98, 5656–5665. [Google Scholar] [CrossRef]
- Groher, T.; Röhlen-Schmittgen, S.; Fiebig, A.; Noga, G.; Hunsche, M. Influence of supplementary LED lighting on physiological and biochemical parameters of tomato (Solanum lycopersicum L.) leaves. Sci. Hortic. 2019, 250, 154–158. [Google Scholar] [CrossRef]
- Sun, X.; Sun, G.; Wang, M.; Xiao, J.; Sun, X. Protective effects of cynaroside against H2O2-induced apoptosis in H9c2 cardiomyoblasts. J. Cell. Biochem. 2011, 112, 2019–2029. [Google Scholar] [CrossRef]
- Chen, J.; Yang, T.; Huang, X.; Ma, Y.; Zhang, X.; Shen, Y.; Geng, C.; Lu, C. Application of Graveobioside A in the preparation of medicines or health foods against hyperuricemia and gout 2016. Patent CN105560262A, 11 May 2016. [Google Scholar]
- Lin, L.-Z.; Lu, S.; Harnly, J.M. Detection and Quantification of Glycosylated Flavonoid Malonates in Celery, Chinese Celery, and Celery Seed by LC-DAD-ESI/MS. J. Agric. Food Chem. 2007, 55, 1321–1326. [Google Scholar] [CrossRef] [PubMed]
- Kashiwagi, T.; Horibata, Y.; Mekuria, D.B.; Tebayashi, S.; Kim, C.-S. Ovipositional Deterrent in the Sweet Pepper, Capsicum annuum, at the Mature Stage against Liriomyza trifolii (Burgess). Biosci. Biotechnol. Biochem. 2005, 69, 1831–1835. [Google Scholar] [CrossRef] [PubMed]
- Castillo-Sánchez, L.E.; Jiménez-Osornio, J.J.; América, M. Secondary Metabolites of the Annonaceae, Solanaceae and Meliaceae Families used as Biological Control of Insects. Trop. Subtrop. Agroecosyst. 2010, 12, 445–462. [Google Scholar]
- Villaverde, J.J.; Sandín-España, P.; Sevilla-Morán, B.; López-Goti, C.; Alonso-Prados, J.L. Biopesticides from Natural Products: Current Development, Legislative Framework, and Future Trends. BioResources 2016, 11, 5618–5640. [Google Scholar] [CrossRef]
- Leufen, G.; Noga, G.; Hunsche, M. Physiological response of sugar beet (Betavulgaris) genotypes to a temporary water deficit, as evaluated with a multiparameter fluorescence sensor. Acta Physiol. Plant. 2013, 35, 1763–1774. [Google Scholar] [CrossRef]
- Goulas, Y.; Cerovic, Z.G.; Cartelat, A.; Moya, I. Dualex: A new instrument for field measurements of epidermal ultraviolet absorbance by chlorophyll fluorescence. Appl. Opt. 2004, 43, 4488. [Google Scholar] [CrossRef] [PubMed]
- Cerovic, Z.G.; Moise, N.; Agati, G.; Latouche, G.; Ben Ghozlen, N.; Meyer, S. New portable optical sensors for the assessment of winegrape phenolic maturity based on berry fluorescence. J. Food Compos. Anal. 2008, 21, 650–654. [Google Scholar] [CrossRef]
- Schmittgen, S.; Ellenberger, J. A dataset on secondary metabolites in bell pepper leaves. 2019. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. Available online: https://www.R-project.org/ (accessed on 16 January 2020).
- RStudio Team. RStudio: Integrated Development Environment for R; Boston, MA, USA, 2016. [Google Scholar]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer International Publishing A&G: Cham, Switzerland, 2016. [Google Scholar]
- Mudrić, S.Ž.; Gašić, U.M.; Dramićanin, A.M.; Ćirić, I.Ž.; Milojković-Opsenica, D.M.; Popović-Đorđević, J.B.; Momirović, N.M.; Tešić, Ž.L. The polyphenolics and carbohydrates as indicators of botanical and geographical origin of Serbian autochthonous clones of red spice paprika. Food Chem. 2017, 217, 705–715. [Google Scholar] [CrossRef]
- Kim, W.-R.; Kim, E.O.; Kang, K.; Oidovsambuu, S.; Jung, S.H.; Kim, B.S.; Nho, C.W.; Um, B.-H. Antioxidant Activity of Phenolics in Leaves of Three Red Pepper (Capsicum annuum) Cultivars. J. Agric. Food Chem. 2014, 62, 850–859. [Google Scholar] [CrossRef]
- Ramakrishna, A.; Ravishankar, G.A. Influence of abiotic stress signals on secondary metabolites in plants. Plant Signal. Behav. 2011, 6, 1720–1731. [Google Scholar] [PubMed]
- Hernandez, J.A.; Almansa, M.S. Short-term effects of salt stress on antioxidant systems and leaf water relations of pea leaves. Physiol. Plant. 2002, 115, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Jansen, M.A.K.; Hectors, K.; O’Brien, N.M.; Guisez, Y.; Potters, G. Plant stress and human health: Do human consumers benefit from UV-B acclimated crops? Plant Sci. 2008, 175, 449–458. [Google Scholar] [CrossRef]
- Barnes, P.W.; Searles, P.S.; Ballaré, C.L.; Ryel, R.J.; Caldwell, M.M. Non-invasive measurements of leaf epidermal transmittance of UV radiation using chlorophyll fluorescence: field and laboratory studies. Physiol. Plant. 2000, 109, 274–283. [Google Scholar] [CrossRef]
- Marenco, R.A.; Antezana-Vera, S.A.; Nascimento, H.C.S. Relationship between specific leaf area, leaf thickness, leaf water content and SPAD-502 readings in six Amazonian tree species. Photosynthetica 2009, 47, 184–190. [Google Scholar] [CrossRef]
- Padilla, F.M.; de Souza, R.; Peña-Fleitas, M.T.; Gallardo, M.; Giménez, C.; Thompson, R.B. Different Responses of Various Chlorophyll Meters to Increasing Nitrogen Supply in Sweet Pepper. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef]
- Close, D.C.; McArthur, C. Rethinking the role of many plant phenolics–protection from photodamage not herbivores? Oikos 2002, 99, 166–172. [Google Scholar] [CrossRef]
- Close, D.C.; Beadle, C.L. The Ecophysiology of Foliar Anthocyanin. Bot. Rev. 2003, 69, 149–161. [Google Scholar] [CrossRef]
- Riederer, M.; Muller, C. Annual Plant Reviews, Biology of the Plant Cuticle; John Wiley & Sons: Hoboken, NJ, USA, 2008; ISBN 978-1-4051-7157-1. [Google Scholar]
- Parihar, P.; Singh, S.; Singh, R.; Singh, V.P.; Prasad, S.M. Effect of salinity stress on plants and its tolerance strategies: A review. Environ. Sci. Pollut. Res. 2015, 22, 4056–4075. [Google Scholar] [CrossRef]
- Costa, J.M.; Ortuño, M.F.; Chaves, M.M. Deficit Irrigation as a Strategy to Save Water: Physiology and Potential Application to Horticulture. J. Integr. Plant Biol. 2007, 49, 1421–1434. [Google Scholar] [CrossRef]
- Sgherri, C.; Navari-Izzo, F.; Pardossi, A.; Soressi, G.P.; Izzo, R. The Influence of Diluted Seawater and Ripening Stage on the Content of Antioxidants in Fruits of Different Tomato Genotypes. J. Agric. Food Chem. 2007, 55, 2452–2458. [Google Scholar] [CrossRef] [PubMed]
- Sgherri, C.; Kadlecová, Z.; Pardossi, A.; Navari-Izzo, F.; Izzo, R. Irrigation with Diluted Seawater Improves the Nutritional Value of Cherry Tomatoes. J. Agric. Food Chem. 2008, 56, 3391–3397. [Google Scholar] [CrossRef] [PubMed]
- Neocleous, D.; Nikolaou, G. Antioxidant Seasonal Changes in Soilless Greenhouse Sweet Peppers. Agronomy 2019, 9, 730. [Google Scholar] [CrossRef]
- Sun, M.; Gu, X.; Fu, H.; Zhang, L.; Chen, R.; Cui, L.; Zheng, L.; Zhang, D.; Tian, J. Change of secondary metabolites in leaves of Ginkgo biloba L. in response to UV-B induction. Innov. Food Sci. Emerg. Technol. 2010, 11, 672–676. [Google Scholar] [CrossRef]
- Gogo, E.O.; Förster, N.; Dannehl, D.; Frommherz, L.; Trierweiler, B.; Opiyo, A.M.; Ulrichs, C.; Huyskens-Keil, S. Postharvest UV-C application to improve health promoting secondary plant compound pattern in vegetable amaranth. Innov. Food Sci. Emerg. Technol. 2017, 45, 426–437. [Google Scholar] [CrossRef]
- Brugger, A.; Behmann, J.; Paulus, S.; Luigs, H.-G.; Kuska, M.T.; Schramowski, P.; Kersting, K.; Steiner, U.; Mahlein, A.-K. Extending Hyperspectral Imaging for Plant Phenotyping to the UV-Range. Remote Sens. 2019, 11, 1401. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | Cultivar | Treatment | Cultivar × Treatment |
|---|---|---|---|
| Cynaroside | 0.179 | <2 × 10−16 | 0.917 |
| Graveobioside A | 0.055 | 1.25 × 10−5 | 0.141 |
| Dry Weight | 0.00082 | 3.8 × 10−12 | 0.426 |
| Fresh Weight | 0.00017 | 1.15 × 10−15 | 0.146 |
| Leaf | Cultivar | Treatment | DATI | Treatment × DATI |
|---|---|---|---|---|
| All leaves | Mavras | 0.085 | 0.027 | <2 × 10−16 |
| Stayer | 0.079 | 0.509 | 2.17 × 10−6 | |
| Leaf 4 | Mavras | 0.00011 | 0.055 | 0.00027 |
| Stayer | 8.37 × 10−12 | 0.00484 | 0.081 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ellenberger, J.; Siefen, N.; Krefting, P.; Schulze Lutum, J.-B.; Pfarr, D.; Remmel, M.; Schröder, L.; Röhlen-Schmittgen, S. Effect of UV Radiation and Salt Stress on the Accumulation of Economically Relevant Secondary Metabolites in Bell Pepper Plants. Agronomy 2020, 10, 142. https://doi.org/10.3390/agronomy10010142
Ellenberger J, Siefen N, Krefting P, Schulze Lutum J-B, Pfarr D, Remmel M, Schröder L, Röhlen-Schmittgen S. Effect of UV Radiation and Salt Stress on the Accumulation of Economically Relevant Secondary Metabolites in Bell Pepper Plants. Agronomy. 2020; 10(1):142. https://doi.org/10.3390/agronomy10010142
Chicago/Turabian StyleEllenberger, Jan, Nils Siefen, Priska Krefting, Jan-Bernd Schulze Lutum, Daniel Pfarr, Maja Remmel, Lukas Schröder, and Simone Röhlen-Schmittgen. 2020. "Effect of UV Radiation and Salt Stress on the Accumulation of Economically Relevant Secondary Metabolites in Bell Pepper Plants" Agronomy 10, no. 1: 142. https://doi.org/10.3390/agronomy10010142
APA StyleEllenberger, J., Siefen, N., Krefting, P., Schulze Lutum, J.-B., Pfarr, D., Remmel, M., Schröder, L., & Röhlen-Schmittgen, S. (2020). Effect of UV Radiation and Salt Stress on the Accumulation of Economically Relevant Secondary Metabolites in Bell Pepper Plants. Agronomy, 10(1), 142. https://doi.org/10.3390/agronomy10010142