
Retinoic Acid Signalling in the Pineal Gland Is Conserved across Mammalian Species and Its Transcriptional Activity Is Inhibited by Melatonin

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Preparation of Human Tissue
2.3. Ex Vivo Pineal Gland Culture
2.4. Cell Culture
2.5. PCR
2.6. Quantitative PCR
3. Results
3.1. Components Necessary for Retinoic Acid Signalling Are Expressed in the Human and Mouse Pineal Gland
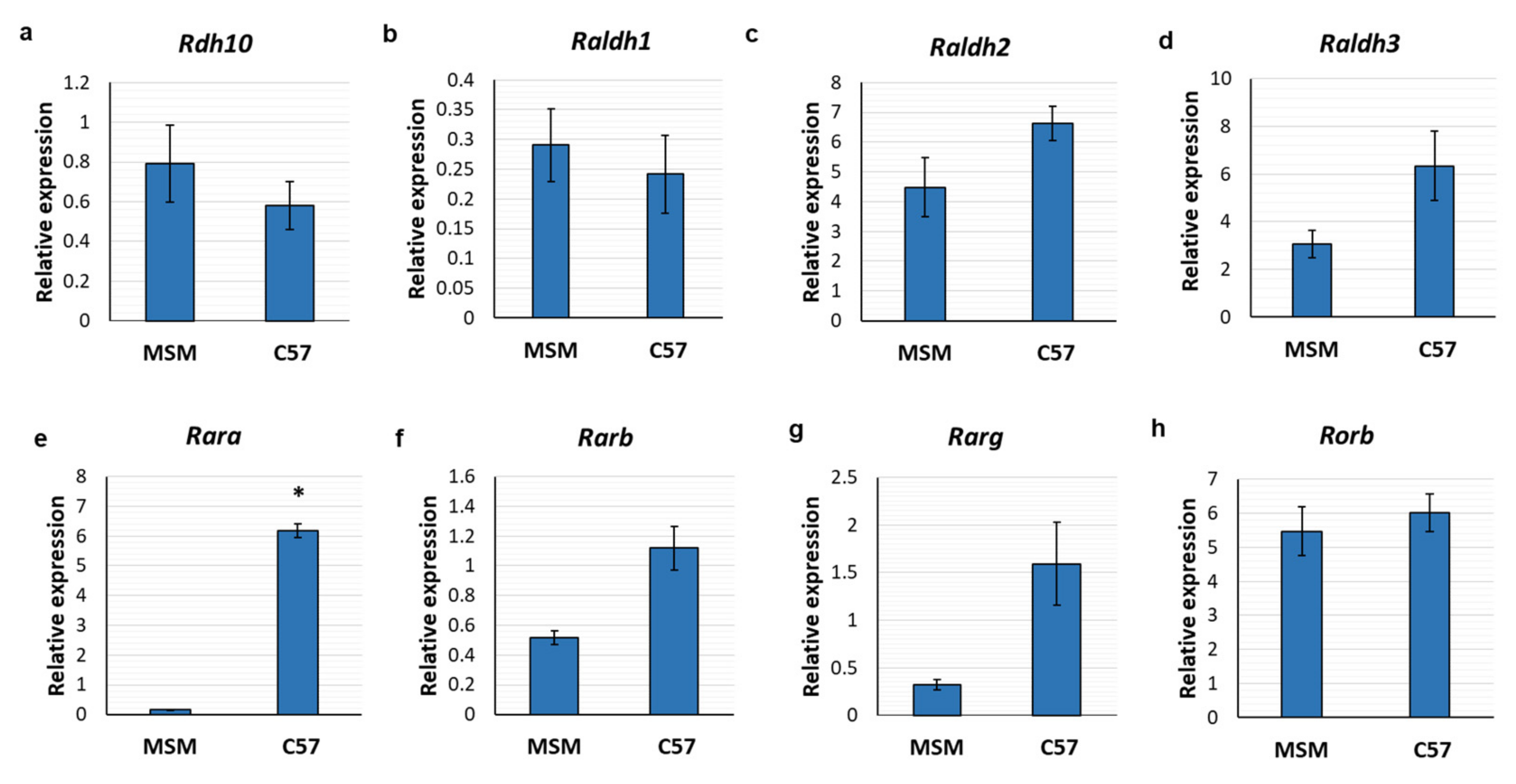
3.2. Retinoic Acid Receptors Are Differentially Expressed between Mouse Strains
3.3. Retinoic Acid and Pineal Clock Gene Expression
3.4. Melatonin Inhibits Induction of Gene Expression by Retinoic Acid
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AANAT | Arylalkylamine N-acetyltransferase |
| ASMT | Acetylserotonin O-methyltransferase |
| C57 | C57BL/6J |
| CAR | Cone arrestin |
| CBA | CBA/Ca |
| cDNA | Complementary deoxyribonucleic acid |
| Cry | Cryptochrome |
| DMSO | Dimethyl sulfoxide |
| ER | Oestrogen receptor |
| MEL | Melatonin |
| mRNA | Messenger ribonucleic acid |
| MSM | MSM/Ms |
| NE | Norepinephrine |
| qPCR | Quantitative polymerase chain reaction |
| RA | Retinoic acid |
| RALDH | Retinaldehyde dehydrogenase |
| RAR | Retinoic acid receptor |
| RARE | Retinoic acid response element |
| RNA | Ribonucleic acid |
| ROR | Retinoic acid receptor-related orphan receptor |
| RORE | ROR response element |
| SCN | Suprachiasmatic nucleus |
| SD | Sprague Dawley |
| SEM | Standard error of the mean |
| ZT | Zeitgeber time |
References
- Klein, D.C. Arylalkylamine N-acetyltransferase: “The timezyme”. J. Biol. Chem. 2007, 282, 4233–4237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, D.C.; Coon, S.L.; Roseboom, P.H.; Weller, J.L.; Bernard, M.; Gastel, J.A.; Zatz, M.; Iuvone, P.M.; Rodriguez, I.R.; Bégay, V.; et al. The melatonin rhythm-generating enzyme: Molecular regulation of serotonin N-acetyltransferase in the pineal gland. Recent Prog. Horm. Res. 1997, 52, 307–358. [Google Scholar] [PubMed]
- Coon, S.L.; Roseboom, P.H.; Baler, R.; Weller, J.L.; Namboodiri, M.; Koonin, E.; Klein, D.C. Pineal serotonin N-acetyltransferase: Expression cloning and molecular analysis. Science 1995, 270, 1681–1683. [Google Scholar] [CrossRef] [PubMed]
- Ganguly, S.; Gastel, J.A.; Weller, J.L.; Schwartz, C.; Jaffe, H.; Namboodiri, M.A.A.; Coon, S.L.; Hickman, A.B.; Rollag, M.; Obsil, T.; et al. Role of a pineal cAMP-operated arylalkylamine N-acetyltransferase/14-3-3-binding switch in melatonin synthesis. Proc. Natl. Acad. Sci. USA 2001, 98, 8083–8088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ackermann, K.; Bux, R.; Rüb, U.; Korf, H.W.; Kauert, G.; Stehle, J.H. Characterization of human melatonin synthesis using autoptic pineal tissue. Endocrinology 2006, 147, 3235–3242. [Google Scholar] [CrossRef] [Green Version]
- Borjigin, J.; Wang, M.M.; Snyder, S.H. Diurnal variation in mRNA encoding serotonin N-acetyltransferase in pineal gland. Nature 1995, 378, 783–785. [Google Scholar] [CrossRef]
- Goto, M.; Oshima, I.; Tomita, T.; Ebihara, S. Melatonin content of the pineal gland in different mouse strains. J. Pineal Res. 1989, 7, 195–204. [Google Scholar] [CrossRef]
- Ebihara, S.; Marks, T.; Hudson, D.J.; Menaker, M. Genetic control of melatonin synthesis in the pineal gland of the mouse. Science 1986, 231, 491–493. [Google Scholar] [CrossRef]
- Roseboom, P.H.; Namboodiri, M.; Zimonjic, D.; Popescu, N.; Rodriguez, I.; Gastel, J.; Klein, D.C. Natural melatonin “knockdown” in C57BL/6J mice: Rare mechanism truncates serotonin N-acetyltransferase. Mol. Brain Res. 1998, 63, 189–197. [Google Scholar] [CrossRef]
- Kasahara, T.; Abe, K.; Mekada, K.; Yoshiki, A.; Kato, T. Genetic variation of melatonin productivity in laboratory mice under domestication. Proc. Natl. Acad. Sci. USA 2010, 107, 6412–6417. [Google Scholar] [CrossRef]
- Namihira, M.; Honma, S.; Abe, H.; Tanahashi, Y.; Ikeda, M.; Honma, K.I. Daily variation and light responsiveness of mammalian clock gene, Clock and BMAL1, transcripts in the pineal body and different areas of brain in rats. Neurosci. Lett. 1999, 267, 69–72. [Google Scholar] [CrossRef]
- Fukuhara, C.; Dirden, J.C.; Tosini, G. Circadian expression of Period 1, Period 2, and arylalkylamine N-acetyltransferase mRNA in the rat pineal gland under different light conditions. Neurosci. Lett. 2000, 286, 167–170. [Google Scholar] [CrossRef]
- Nakamura, T.J.; Shinohara, K.; Funabashi, T.; Mitsushima, D.; Kimura, F. Circadian and photic regulation of cryptochrome mRNAs in the rat pineal gland. Neurosci. Res. 2001, 41, 25–32. [Google Scholar] [CrossRef]
- Dibner, C.; Schibler, U.; Albrecht, U. The mammalian circadian timing system: Organization and coordination of central and peripheral clocks. Annu. Rev. Physiol. 2010, 72, 517–549. [Google Scholar] [CrossRef] [Green Version]
- Fukuhara, C.; Yamazaki, S.; Liang, J. Pineal circadian clocks gate arylalkylamine N-acetyltransferase gene expression in the mouse pineal gland. J. Neurochem. 2005, 93, 156–162. [Google Scholar] [CrossRef]
- Simonneaux, V.; Poirel, V.J.; Garidou, M.L.; Nguyen, D.; Diaz-Rodriguez, E.; Pévet, P. Daily rhythm and regulation of clock gene expression in the rat pineal gland. Mol. Brain Res. 2004, 120, 164–172. [Google Scholar] [CrossRef]
- Karolczak, M.; Burbach, G.J.; Sties, G.; Korf, H.W.; Stehle, J.H. Clock gene mRNA and protein rhythms in the pineal gland of mice. Eur. J. Neurosci. 2004, 19, 3382–3388. [Google Scholar] [CrossRef]
- Andrade-Silva, J.; Cipolla-Neto, J.; Peliciari-Garcia, R.A. The in vitro maintenance of clock genes expression within the rat pineal gland under standard and norepinephrine-synchronized stimulation. Neurosci. Res. 2014, 81–82, 1–10. [Google Scholar] [CrossRef]
- Wongchitrat, P.; Felder-Schmittbuhl, M.P.; Phansuwan-Pujito, P.; Pévet, P.; Simonneaux, V. Endogenous rhythmicity of Bmal1 and Rev-erbα in the hamster pineal gland is not driven by norepinephrine. Eur. J. Neurosci. 2009, 29, 2009–2016. [Google Scholar] [CrossRef]
- Wongchitrat, P.; Felder-Schmittbuhl, M.P.; Govitrapong, P.; Phansuwan-Pujito, P.; Simonneaux, V. A noradrenergic sensitive endogenous clock is present in the rat pineal gland. Neuroendocrinology 2011, 94, 75–83. [Google Scholar] [CrossRef]
- Ghyselinck, N.B.; Duester, G. Retinoic acid signaling pathways. Development 2019, 146, dev167502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ransom, J.; Morgan, P.J.; McCaffery, P.J.; Stoney, P.N. The rhythm of retinoids in the brain. J. Neurochem. 2014, 129, 366–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shearer, K.D.; Stoney, P.N.; Morgan, P.J.; McCaffery, P.J. A vitamin for the brain. Trends Neurosci. 2012, 35, 733–741. [Google Scholar] [CrossRef] [PubMed]
- Goodman, T.; Crandall, J.E.; Nanescu, S.E.; Quadro, L.; Shearer, K.; Ross, A.; Mccaffery, P. Patterning of retinoic acid signaling and cell proliferation in the hippocampus. Hippocampus 2012, 22, 2171–2183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashton, A.; Stoney, P.N.; Ransom, J.; McCaffery, P. Rhythmic Diurnal Synthesis and Signaling of Retinoic Acid in the Rat Pineal Gland and its Action to Rapidly Down-regulate ERK Phosphorylation. Mol. Neurobiol. 2018, 55, 8219–8235. [Google Scholar] [CrossRef] [Green Version]
- Shirai, H.; Oishi, K.; Ishida, N. Bidirectional CLOCK/BMAL1-dependent circadian gene regulation by retinoic acid in vitro. Biochem. Biophys. Res. Commun. 2006, 351, 387–391. [Google Scholar] [CrossRef]
- McNamara, P.; Seo, S.; Rudic, R.; Sehgal, A.; Chakravarti, D.; FitzGerald, G. Regulation of CLOCK and MOP4 by nuclear hormone receptors in the vasculature: A humoral mechanism to reset a peripheral clock. Cell 2001, 105, 877–889. [Google Scholar] [CrossRef] [Green Version]
- Sherman, H.; Gutman, R.; Chapnik, N.; Meylan, J.; le Coutre, J.; Froy, O. All-trans retinoic acid modifies the expression of clock and disease marker genes. J. Nutr. Biochem. 2012, 23, 209–217. [Google Scholar] [CrossRef]
- Nakahata, Y.; Akashi, M.; Trcka, D.; Yasuda, A.; Takumi, T. The in vitro real-time oscillation monitoring system identifies potential entrainment factors for circadian clocks. BMC Mol. Biol. 2006, 7, 5. [Google Scholar] [CrossRef] [Green Version]
- Navigatore-Fonzo, L.S.; Golini, R.L.; Ponce, I.T.; Delgado, S.M.; Plateo-Pignatari, M.G.; Gimenez, M.S.; Anzulovich, A.C. Retinoic acid receptors move in time with the clock in the hippocampus. Effect of a vitamin-A-deficient diet. J. Nutr. Biochem. 2013, 24, 859–867. [Google Scholar] [CrossRef]
- Golini, R.S.; Delgado, S.M.; Navigatore Fonzo, L.S.; Ponce, I.T.; Lacoste, M.G.; Anzulovich, A.C. Daily patterns of clock and cognition-related factors are modified in the hippocampus of vitamin A-deficient rats. Hippocampus 2012, 22, 1720–1732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navigatore-Fonzo, L.S.; Delgado, S.M.; Golini, R.S.; Anzulovich, A.C. Circadian rhythms of locomotor activity and hippocampal clock genes expression are dampened in vitamin A-deficient rats. Nutr. Res. 2014, 34, 326–335. [Google Scholar] [CrossRef] [PubMed]
- Navigatore Fonzo, L.S.; Golini, R.S.; Delgado, S.M.; Ponce, I.T.; Bonomi, M.R.; Rezza, I.G.; Gimenez, M.S.; Anzulovich, A.C. Temporal patterns of lipoperoxidation and antioxidant enzymes are modified in the hippocampus of vitamin A-deficient rats. Hippocampus 2009, 19, 869–880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, M.J.; Coon, S.L.; Carter, D.A.; Humphries, A.; Kim, J.S.; Shi, Q.; Gaildrat, P.; Morin, F.; Ganguly, S.; Hogenesch, J.B.; et al. Night/Day Changes in Pineal Expression of >600 Genes: Central Role of Adrenergic/cAMP signaling. J. Biol. Chem. 2009, 284, 7606–7622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vivien-Roels, B.; Malan, A.; Rettori, M.C.; Delagrange, P.; Jeanniot, J.-P.; Pévet, P. Daily Variations in Pineal Melatonin Concentrations in Inbred and Outbred Mice. J. Biol. Rhythms. 1998, 13, 403–409. [Google Scholar] [CrossRef]
- Moriwaki, K.; Miyashita, N.; Mita, A.; Gotoh, H.; Tsuchiya, K.; Kato, H.; Mekada, K.; Noro, C.; Oota, S.; Yoshiki, A.; et al. Unique inbred strain MSM/Ms established from the Japanese wild mouse. Exp. Anim. 2009, 58, 123–134. [Google Scholar] [CrossRef] [Green Version]
- De Thé, H.; Vivanco-Ruiz, M.M.; Tiollais, P.; Stunnenberg, H.; Dejean, A. Identification of a retinoic acid responsive element in the retinoic acid receptor β gene. Nature 1990, 343, 177–180. [Google Scholar] [CrossRef]
- Teplitzky, S.R.; Kiefer, T.L.; Cheng, Q.; Dwivedi, P.D.; Moroz, K.; Myers, L.; Anderson, M.B.; Collins, A.; Dai, J.; Yuan, L.; et al. Chemoprevention of NMU-induced rat mammary carcinoma with the combination of melatonin and 9-cis-retinoic acid. Cancer Lett. 2001, 168, 155–163. [Google Scholar] [CrossRef]
- Nowfar, S.; Teplitzky, S.R.; Melancon, K.; Kiefer, T.L.; Cheng, Q.; Dwivedi, P.D.; Bischoff, E.D.; Moroz, K.; Anderson, M.B.; Dai, J.; et al. Tumor prevention by 9-cis-retinoic acid in the N-nitroso-N-methylurea model of mammary carcinogenesis is potentiated by the pineal hormone melatonin. Breast Cancer Res. Treat. 2002, 72, 33–43. [Google Scholar] [CrossRef]
- Li, A.; Zhu, X.; Brown, B.; Craft, C.M. Melatonin enhances retinoic acid induction of cone arrestin gene expression in retinoblastoma cells. Adv. Exp. Med. Biol. 2003, 533, 361–368. [Google Scholar] [CrossRef]
- Kiefer, T.L.; Lai, L.; Yuan, L.; Dong, C.; Burow, M.E.; Hill, S.M. Differential regulation of estrogen receptor alpha, glucocorticoid receptor and retinoic acid receptor alpha transcriptional activity by melatonin is mediated via different G proteins. J. Pineal Res. 2005, 38, 231–239. [Google Scholar] [CrossRef]
- Loudig, O.; Babichuk, C.; White, J.; Abu-Abed, S.; Mueller, C.; Petkovich, M. Cytochrome P450RAI(CYP26) promoter: A distinct composite retinoic acid response element underlies the complex regulation of retinoic acid metabolism. Mol. Endocrinol. 2000, 14, 1483–1497. [Google Scholar] [CrossRef]
- Gastel, J.A.; Roseboom, P.H.; Rinaldi, P.A.; Weller, J.L.; Klein, D.C. Melatonin production: Proteasomal proteolysis in serotonin N-acetyltransferase regulation. Science 1998, 279, 1358–1360. [Google Scholar] [CrossRef]
- Ganguly, S.; Weller, J.L.; Ho, A.; Chemineau, P.; Malpaux, B.; Klein, D.C. Melatonin synthesis: 14-3-3-dependent activation and inhibition of arylalkylamine N-acetyltransferase mediated by phosphoserine-205. Proc. Natl. Acad. Sci. USA 2005, 102, 1222–1227. [Google Scholar] [CrossRef] [Green Version]
- Maronde, E.; Saade, A.; Ackermann, K.; Goubran-Botros, H.; Pagan, C.; Bux, R.; Bourgeron, T.; Dehghani, F.; Stehle, J.H. Dynamics in enzymatic protein complexes offer a novel principle for the regulation of melatonin synthesis in the human pineal gland. J. Pineal Res. 2011, 51, 145–155. [Google Scholar] [CrossRef]
- Maghsoodi, B.; Poon, M.M.; Nam, C.I.; Aoto, J.; Ting, P.; Chen, L. Retinoic acid regulates RARα-mediated control of translation in dendritic RNA granules during homeostatic synaptic plasticity. Proc. Natl. Acad. Sci. USA 2008, 105, 16015–16020. [Google Scholar] [CrossRef] [Green Version]
- Kambhampati, S.; Li, Y.; Verma, A.; Sassano, A.; Majchrzak, B.; Deb, D.K.; Parmar, S.; Giafis, N.; Kalvakolanu, D.V.; Rahman, A.; et al. Activation of protein kinase C by all-trans-retinoic acid. J. Biol. Chem. 2003, 278, 32544–32551. [Google Scholar] [CrossRef] [Green Version]
- Masiá, S.; Alvarez, S.; de Lera, A.R.; Barettino, D. Rapid, nongenomic actions of retinoic acid on phosphatidylinositol-3-kinase signaling pathway mediated by the retinoic acid receptor. Mol. Endocrinol. 2007, 21, 2391–2402. [Google Scholar] [CrossRef]
- Ross, A.W.; Webster, C.A.; Mercer, J.G.; Moar, K.M.; Ebling, F.J.; Schuhler, S.; Barrett, P.; Morgan, P.J. Photoperiodic Regulation of Hypothalamic Retinoid Signaling: Association of Retinoid X Receptor γ with Body Weight. Endocrinology 2004, 145, 13–20. [Google Scholar] [CrossRef] [Green Version]
- Helfer, G.; Ross, A.W.; Russell, L.; Thomson, L.M.; Shearer, K.D.; Goodman, T.H.; McCaffery, P.J.; Morgan, P.J. Photoperiod regulates vitamin A and Wnt/β-catenin signaling in F344 rats. Endocrinology 2012, 153, 815–824. [Google Scholar] [CrossRef]
- Von Gall, C.; Lewy, A.; Schomerus, C.; Vivien-Roels, B.; Pevet, P.; Korf, H.-W.; Stehle, J.H. Transcription factor dynamics and neuroendocrine signalling in the mouse pineal gland: A comparative analysis of melatonin-deficient C57BL mice and melatonin-proficient C3H mice. Eur. J. Neurosci. 2000, 12, 964–972. [Google Scholar] [CrossRef] [PubMed]
- Conti, A.; Maestroni, G.J. HPLC validation of a circadian melatonin rhythm in the pineal gland of inbred mice. J. Pineal Res. 1996, 20, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Eck-Enriquez, K.; Kiefer, T.; Spriggs, L.; Hill, S. Pathways through which a regimen of melatonin and retinoic acid induces apoptosis in MCF-7 human breast cancer cells. Breast Cancer Res. Treat. 2000, 61, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Lai, L.; Yuan, L.; Chen, Q.; Dong, C.; Mao, L.; Rowan, B.; Frasch, T.; Hill, S.M. The Gαi and Gαq proteins mediate the effects of melatonin on steroid/thyroid hormone receptor transcriptional activity and breast cancer cell proliferation. J. Pineal Res. 2008, 45, 476–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiefer, T.; Ram, P.T.; Yuan, L.; Hill, S.M. Melatonin inhibits estrogen receptor transactivation and cAMP levels in breast cancer cells. Breast Cancer Res. Treat. 2002, 71, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Wiesenberg, I.; Missbach, M.; Kahlen, J.P.; Schräder, M.; Carlberg, C. Transcriptional activation of the nuclear receptor RZRα by the pineal gland hormone melatonin and identification of CGP 52608 as a synthetic ligand. Nucleic Acids Res. 1995, 23, 327–333. [Google Scholar] [CrossRef] [Green Version]
- Dai, J.; Ram, P.T.; Yuan, L.; Spriggs, L.L. ; Hill SM Transcriptional repression of RORα activity in human breast cancer cells by melatonin. Mol. Cell Endocrinol. 2001, 176, 111–120. [Google Scholar] [CrossRef]
- Lardone, P.J.; Guerrero, J.M.; Fernández-Santos, J.M.; Rubio, A.; Martín-Lacave, I.; Carrillo-Vico, A. Melatonin synthesized by T lymphocytes as a ligand of the retinoic acid-related orphan receptor. J. Pineal Res. 2011, 51, 454–462. [Google Scholar] [CrossRef]
- Schrader, M.; Danielsson, C.; Wiesenberg, I.; Carlberg, C. Identification of Natural Monomeric Response Elements of the Nuclear Receptor RZR/ROR. J. Biol. Chem. 1996, 271, 19732–19736. [Google Scholar] [CrossRef] [Green Version]
- Tini, M.; Fraser, R.A.; Giguere, V. Functional interactions between retinoic acid receptor-related orphan nuclear receptor (RORα) and the retinoic acid receptors in the regulation of the γF-crystallin promoter. J. Biol. Chem. 1995, 270, 20156–20161. [Google Scholar] [CrossRef]
- Davis, K.; Lazar, M. Induction of retinoic acid receptor-β by retinoic acid is cell specific. Endocrinology 1993, 132, 1469–1474. [Google Scholar] [CrossRef]
- Folkers, G.E.; van der Burg, B.; van der Saag, P.T. Promoter architecture, cofactors, and orphan receptors contribute to cell-specific activation of the retinoic acid receptor β2 promoter. J. Biol. Chem. 1998, 273, 32200–32212. [Google Scholar] [CrossRef] [Green Version]
- Balmer, J.E.; Blomhoff, R. Gene expression regulation by retinoic acid. J. Lipid Res. 2002, 43, 1773–1808. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.; Casadesus, G.; Zhu, X.; Lee, H.; Perry, G.; Smith, M.A.; Gustaw-Rothenberg, K.; Lerner, A. All-trans-retinoic acid as a novel therapeutic strategy for Alzheimer’s Disease. Expert Rev. Neurother. 2009, 9, 1615–1621. [Google Scholar] [CrossRef] [Green Version]
- Rosales-Corral, S.A.; Acuña-Castroviejo, D.; Coto-Montes, A.; Boga, J.A.; Manchester, L.C.; Fuentes-Broto, L.; Korkmaz, A.; Ma, S.; Tan, D.X.; Reiter, R.J. Alzheimer’s disease: Pathological mechanisms and the beneficial role of melatonin. J. Pineal Res. 2012, 52, 167–202. [Google Scholar] [CrossRef]
- Ceinos, R.M.; Chansard, M.; Revel, F.; Calgari, C.; Míguez, J.M.; Simonneaux, V. Analysis of adrenergic regulation of melatonin synthesis in Siberian hamster pineal emphasizes the role of HIOMT. NeuroSignals 2004, 13, 308–317. [Google Scholar] [CrossRef]
- Cawley, S.; Bekiranov, S.; Ng, H.H.; Kapranov, P.; Sekinger, E.A.; Kampa, D.; Piccolboni, A.; Sementchenko, V.; Cheng, J.; Williams, A.J.; et al. Unbiased mapping of transcription factor binding sites along human chromosomes 21 and 22 points to widespread regulation of noncoding RNAs. Cell 2004, 116, 499–509. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | RefSeq Code | Product Size (bp) | Forward Primer (5’-3’) | Reverse Primer (5’-3’) | |
|---|---|---|---|---|---|
| A | Raldh1 | NM_013467.3 | 196 | ACGTGGAAGAAGGGGACAAGGCTG | GCAAAGACTTTCCCACCATTGAGTGCC |
| Raldh2 | NM_009022.4 | 198 | CAAGGAGGCTGGCTTTCCACCC | GGGCTCTTCCCTCCGAGTTCCA | |
| Raldh3 | NM_053080.3 | 150 | TCAAAGAGGTCGGGTTCCCTCCG | AGGCGGCTTCTCTGACCAGCT | |
| Rara | NM_009024.2 | 247 | GAGGGCTGTAAGGGCTTCTTCCG | TGAGCTCGCCCACCTCAGGC | |
| Rarb | NM_011243.1 | 134 | ACACCACGAATTCCAGCGCTGAC | CAGACCTGTGAAGCCCGGCA | |
| Rarg | NM_011244.4, NM_001042727.2 | 218 | CCTGTGAAGGCTGCAAGGGCT | GTCGGGCGAGCCCTCCTCTT | |
| Rdh10 | NM_133832.3 | 241 | GCTGGAGTTGAGGATTACTGTGCCAG | GCTGGTCAGTGAGGATGGCCC | |
| Rorb | NM_001043354.2 | 136 | CGGCCACATCATGCGAGCACA | GGCATTGTTCTGCTGGCTCCTCC | |
| Aanat | NM_009591.3 | 131 | TCCGGCACTTCCTAACCCTGTGT | CCTGTGTAGTGTCAGCGACTCCTGA | |
| Actb | NM_007393.5 | 112 | CCACACCCGCCACCAGTTCG | TACAGCCCGGGGAGCATCGT | |
| B | Raldh1 | NM_013467.3 | 196 | ACGTGGAAGAAGGGGACAAGGCTG | GCAAAGACTTTCCCACCATTGAGTGCC |
| Raldh2 | NM_009022.4 | 198 | CAAGGAGGCTGGCTTTCCACCC | GGGCTCTTCCCTCCGAGTTCCA | |
| Raldh3 | NM_053080.3 | 150 | TCAAAGAGGTCGGGTTCCCTCCG | AGGCGGCTTCTCTGACCAGCT | |
| Rara | NM_009024.2 | 247 | GAGGGCTGTAAGGGCTTCTTCCG | TGAGCTCGCCCACCTCAGGC | |
| Rarb | NM_011243.1 | 134 | ACACCACGAATTCCAGCGCTGAC | CAGACCTGTGAAGCCCGGCA | |
| Rarg | NM_011244.4, NM_001042727.2 | 218 | CCTGTGAAGGCTGCAAGGGCT | GTCGGGCGAGCCCTCCTCTT | |
| Rdh10 | NM_133832.3 | 241 | GCTGGAGTTGAGGATTACTGTGCCAG | GCTGGTCAGTGAGGATGGCCC | |
| Rorb | NM_001043354.2 | 136 | CGGCCACATCATGCGAGCACA | GGCATTGTTCTGCTGGCTCCTCC | |
| Aanat | NM_009591.3 | 131 | TCCGGCACTTCCTAACCCTGTGT | CCTGTGTAGTGTCAGCGACTCCTGA | |
| Actb | NM_007393.5 | 112 | CCACACCCGCCACCAGTTCG | TACAGCCCGGGGAGCATCGT | |
| C | Per1 | NM_001034125.1 | 137 | CCAGTGGTGGGAGGCACCCT | ATGATGTCCGACTCCGGGGGC |
| Per2 | NM_031678.1 | 105 | AAAACTGCTCCACGGGGCGG | CGTCAGGGCTGGGGTGAGTG | |
| Cry1 | NM_198750.2 | 81 | CTGACCCGCGGCGACCTATG | GCTCCAGTCGGCGTCAAGCA | |
| Bmal1 | NM_024362.2 | 135 | CGGGCGACTGCACTCACACA | GCCAAAATAGCCGTCGCCCTCT | |
| Gapdh | NM_017008.4 | 119 | GGGCTCTCTGCTCCTCCCTGT | CAGGCGTCCGATACGGCCAAA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ashton, A.; Clark, J.; Fedo, J.; Sementilli, A.; Fragoso, Y.D.; McCaffery, P. Retinoic Acid Signalling in the Pineal Gland Is Conserved across Mammalian Species and Its Transcriptional Activity Is Inhibited by Melatonin. Cells 2023, 12, 286. https://doi.org/10.3390/cells12020286
Ashton A, Clark J, Fedo J, Sementilli A, Fragoso YD, McCaffery P. Retinoic Acid Signalling in the Pineal Gland Is Conserved across Mammalian Species and Its Transcriptional Activity Is Inhibited by Melatonin. Cells. 2023; 12(2):286. https://doi.org/10.3390/cells12020286
Chicago/Turabian StyleAshton, Anna, Jason Clark, Julia Fedo, Angelo Sementilli, Yara D. Fragoso, and Peter McCaffery. 2023. "Retinoic Acid Signalling in the Pineal Gland Is Conserved across Mammalian Species and Its Transcriptional Activity Is Inhibited by Melatonin" Cells 12, no. 2: 286. https://doi.org/10.3390/cells12020286