
Cellular, Molecular and Clinical Aspects of Aortic Aneurysm—Vascular Physiology and Pathophysiology
 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Epidemiology
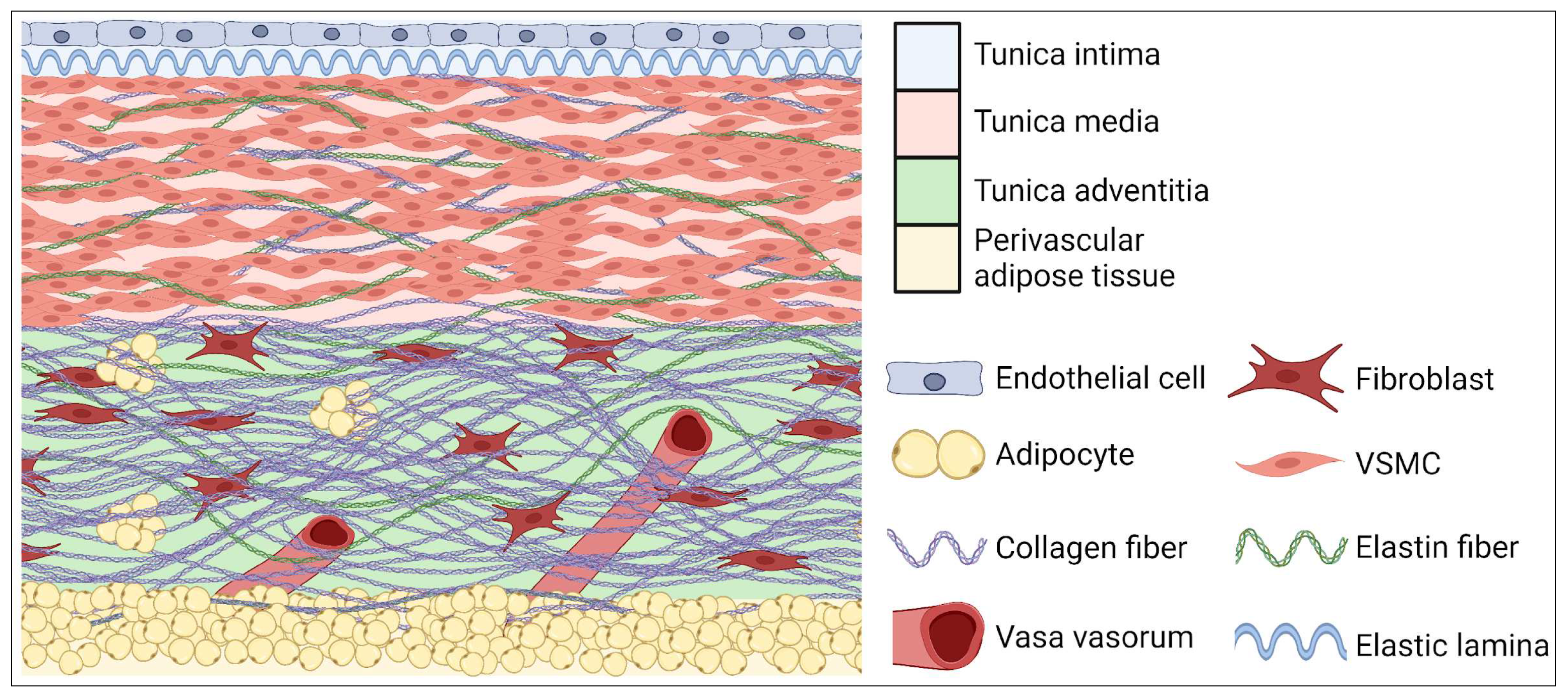
3. Anatomical Structure
4. Tissue and Cellular Structure and Main Abnormalities of Aorta
5. Abdominal Aorta Aneurysm—Cellular Physiology and Pathophysiology
6. Pathogenesis of Abdominal Aortic Aneurysm—Molecular Aspects
6.1. Inflammation—Markers
6.1.1. Chymase and Tryptase
6.1.2. Interleukin-6 (IL-6), Granulocyte Macrophage Colony-Stimulating Factor (GM-CSF) and Transforming Growth Factor Beta (TGF-β)
6.1.3. Monocyte Chemoattractant Protein-1 (MCP-1)
6.1.4. Reactive Oxygen Species (ROS)
6.1.5. Neutrophil Gelatinase-Associated Lipocalin (NGAL)
6.1.6. Interferon-γ (IFN-γ)
6.1.7. Homocysteine
6.1.8. Metalloproteinases (MMP)
6.1.9. Lysine Oxidase (LOX)
6.1.10. Osteoprotegerin (OPG)
6.1.11. Osteopontin (OPN)
6.1.12. Cathepsins
6.2. VSMCs
6.3. Hypertension and Abdominal Aortic Aneurysm
6.4. Genetic Factors
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Summers, K.L.; Kerut, E.K.; Sheahan, C.M.; Sheahan, M.G. Evaluating the Prevalence of Abdominal Aortic Aneurysms in the United States through a National Screening Database. J. Vasc. Surg. 2021, 73, 61–68. [Google Scholar] [CrossRef]
- Song, P.; He, Y.; Adeloye, D.; Zhu, Y.; Ye, X.; Yi, Q.; Rahimi, K.; Rudan, I. The Global and Regional Prevalence of Abdominal Aortic Aneurysms: A Systematic Review and Modeling Analysis. Ann. Surg. 2023, 277, 912–919. [Google Scholar] [CrossRef] [PubMed]
- Saw, S.T.; Leong, B.D.K.; Abdul Aziz, D.A. Early Detection of Undiagnosed Abdominal Aortic Aneurysm and Sub-Aneurysmal Aortic Dilatations in Patients with High-Risk Coronary Artery Disease: The Value of Targetted Screening Programme. Vasc. Health Risk Manag. 2020, 16, 215–229. [Google Scholar] [CrossRef] [PubMed]
- Koroleva, A.; Kodenko, M.; Leonov, D.; Kulberg, N.; Lisenko, N.; Grigorev, G.; Mokienko, O.; Vladzymyrskyy, A.; Morozov, S. The Effects of Hidden Aneurysms on the Posterior Flow: Computational Fluid Dynamic Study; Electrical and Power Engineering (REEPE): Moscow, Russia, 2021; Volume 3, pp. 1–4. [Google Scholar]
- Safiri, S.; Kolahi, A.-A.; Naghavi, M. Global, Regional and National Burden of Bladder Cancer and Its Attributable Risk Factors in 204 Countries and Territories, 1990–2019: A Systematic Analysis for the Global Burden of Disease Study 2019. BMJ Glob. Health 2021, 6, e004128. [Google Scholar] [CrossRef] [PubMed]
- Saracini, C.; Bolli, P.; Sticchi, E.; Pratesi, G.; Pulli, R.; Sofi, F.; Pratesi, C.; Gensini, G.F.; Abbate, R.; Giusti, B. Polymorphisms of Genes Involved in Extracellular Matrix Remodeling and Abdominal Aortic Aneurysm. J. Vasc. Surg. 2012, 55, 171–179.e2. [Google Scholar] [CrossRef] [PubMed]
- Jabłońska, A.; Zagrapan, B.; Neumayer, C.; Eilenberg, W.; Scheuba, A.; Brostjan, C.; Demyanets, S.; Klinger, M.; Nanobachvili, J.; Huk, I. Polymorphisms in the IL-6 and TNF-α Gene Are Associated with an Increased Risk of Abdominal Aortic Aneurysm. Int. J. Cardiol. 2021, 329, 192–197. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Kong, W. Cellular Signaling in Abdominal Aortic Aneurysm. Cell. Signal. 2020, 70, 109575. [Google Scholar] [CrossRef]
- Kugo, H.; Miyamoto, C.; Sawaragi, A.; Hoshino, K.; Hamatani, Y.; Matsumura, S.; Yoshioka, Y.; Moriyama, T.; Zaima, N. Sesame Extract Attenuates the Degradation of Collagen and Elastin Fibers in the Vascular Walls of Nicotine-Administered Mice. J. Oleo Sci. 2019, 68, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Xu, H.; Wen, H.; Wang, H.; Zhao, R.; Sun, Y.; Bai, C.; Ping, J.; Song, L.; Luo, M.; et al. Lysyl Hydroxylase 1 (LH1) Deficiency Promotes Angiotensin II (Ang II)-Induced Dissecting Abdominal Aortic Aneurysm. Theranostics 2021, 11, 9587–9604. [Google Scholar] [CrossRef]
- Lai, C.-H.; Chang, C.-W.; Lee, F.-T.; Kuo, C.-H.; Hsu, J.-H.; Liu, C.-P.; Wu, H.-L.; Yeh, J.-L. Targeting Vascular Smooth Muscle Cell Dysfunction with Xanthine Derivative KMUP-3 Inhibits Abdominal Aortic Aneurysm in Mice. Atherosclerosis 2020, 297, 16–24. [Google Scholar] [CrossRef]
- Lu, H.; Du, W.; Ren, L.; Hamblin, M.H.; Becker, R.C.; Chen, Y.E.; Fan, Y. Vascular Smooth Muscle Cells in Aortic Aneurysm: From Genetics to Mechanisms. J. Am. Heart Assoc. 2021, 10, e023601. [Google Scholar] [CrossRef] [PubMed]
- Mathur, A.; Mohan, V.; Ameta, D.; Gaurav, B.; Haranahalli, P. Aortic Aneurysm. J. Transl. Intern. Med. 2016, 4, 35–41. [Google Scholar] [CrossRef]
- Duncan, A.; Maslen, C.; Gibson, C.; Hartshorne, T.; Farooqi, A.; Saratzis, A.; Bown, M.J. Ultrasound Screening for Abdominal Aortic Aneurysm in High-Risk Women. Br. J. Surg. 2021, 108, 1192–1198. [Google Scholar] [CrossRef] [PubMed]
- van de Luijtgaarden, K.M.; Rouwet, E.V.; Hoeks, S.E.; Stolker, R.J.; Verhagen, H.J.; Majoor-Krakauer, D. Risk of Abdominal Aortic Aneurysm (AAA) among Male and Female Relatives of AAA Patients. Vasc. Med. 2017, 22, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.; Lu, Y.; Wei, J.; Wu, J.; Yang, J.; Cai, Z. Abdominal Aortic Aneurysm: Roles of Inflammatory Cells. Front. Immunol. 2021, 11, 609161. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Bu, X.; Wang, X.; Liu, J.; Ma, A.; Wang, T. Global Burden of Aortic Aneurysm and Attributable Risk Factors from 1990 to 2017. Glob. Heart 2021, 16, 35. [Google Scholar] [CrossRef] [PubMed]
- Symonides, B.; Śliwczyński, A.; Gałązka, Z.; Pinkas, J.; Gaciong, Z. Geographic Disparities in the Application of Endovascular Repair of Unruptured Abdominal Aortic Aneurysm—Polish Population Analysis. Adv. Med. Sci. 2020, 65, 170–175. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Deng, H.; Zhou, Z.; Xiong, X.; Gao, L. Endothelium as a Potential Target for Treatment of Abdominal Aortic Aneurysm. Oxidative Med. Cell. Longev. 2018, 2018, e6306542. [Google Scholar] [CrossRef]
- Komutrattananont, P.; Mahakkanukrauh, P.; Das, S. Morphology of the Human Aorta and Age-Related Changes: Anatomical Facts. Anat. Cell Biol. 2019, 52, 109–114. [Google Scholar] [CrossRef]
- Stanek, A.; Brożyna-Tkaczyk, K.; Myśliński, W. The Role of Obesity-Induced Perivascular Adipose Tissue (PVAT) Dysfunction in Vascular Homeostasis. Nutrients 2021, 13, 3843. [Google Scholar] [CrossRef]
- Li, H.-F.; Liu, H.-T.; Chen, P.-Y.; Lin, H.; Tseng, T.-L. Role of PVAT in Obesity-Related Cardiovascular Disease through the Buffering Activity of ATF3. iScience 2022, 25, 105631. [Google Scholar] [CrossRef]
- Grewal, N.; Groot, A.C.G.; Lindeman, J.H.; Klautz, A.; Driessen, A.; Klautz, R.J.M.; Poelmann, R.E. Normal and Abnormal Development of the Aortic Valve and Ascending Aortic Wall: A Comprehensive Overview of the Embryology and Pathology of the Bicuspid Aortic Valve. Ann. Cardiothorac. Surg. 2022, 11, 38088–38388. [Google Scholar] [CrossRef]
- Pfaltzgraff, E.R.; Shelton, E.L.; Galindo, C.L.; Nelms, B.L.; Hooper, C.W.; Poole, S.D.; Labosky, P.A.; Bader, D.M.; Reese, J. Embryonic Domains of the Aorta Derived from Diverse Origins Exhibit Distinct Properties That Converge into a Common Phenotype in the Adult. J. Mol. Cell. Cardiol. 2014, 69, 88–96. [Google Scholar] [CrossRef]
- Quintana, R.A.; Taylor, W.R. Introduction to the Compendium on Aortic Aneurysms. Circ. Res. 2019, 124, 470–471. [Google Scholar] [CrossRef]
- Sen, I.; D’Oria, M.; Weiss, S.; Bower, T.C.; Oderich, G.S.; Kalra, M.; Colglazier, J.; DeMartino, R.R. Incidence and Natural History of Isolated Abdominal Aortic Dissection: A Population-Based Assessment from 1995 to 2015. J. Vasc. Surg. 2021, 73, 1198–1204.e1. [Google Scholar] [CrossRef]
- Joshi, N.V.; Elkhawad, M.; Forsythe, R.O.; McBride, O.M.B.; Rajani, N.K.; Tarkin, J.M.; Chowdhury, M.M.; Donoghue, E.; Robson, J.M.J.; Boyle, J.R.; et al. Greater Aortic Inflammation and Calcification in Abdominal Aortic Aneurysmal Disease than Atherosclerosis: A Prospective Matched Cohort Study. Open Heart 2020, 7, e001141. [Google Scholar] [CrossRef]
- Toghill, B.J.; Saratzis, A.; Bown, M.J. Abdominal Aortic Aneurysm—An Independent Disease to Atherosclerosis? Cardiovasc. Pathol. 2017, 27, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Wittig, C.; Szulcek, R. Extracellular Matrix Protein Ratios in the Human Heart and Vessels: How to Distinguish Pathological from Physiological Changes? Front. Physiol. 2021, 12, 708656. [Google Scholar] [CrossRef] [PubMed]
- Pernomian, L.; Tan, W.; McCarthy, C.; Wenceslau, C.F. Reprogramming Endothelial and Vascular Smooth Muscle Cells to Prevent and Treat Hypertension. Med. Hypotheses 2023, 179, 111162. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Jiang, Y.; Qu, Y.; Lv, J.; Zeng, H. sGC Agonist BAY1021189 Promotes Thoracic Aortic Dissection Formation by Accelerating Vascular Smooth Muscle Cell Phenotype Switch. Eur. J. Pharmacol. 2023, 952, 175789. [Google Scholar] [CrossRef] [PubMed]
- Kuzan, A.; Wisniewski, J.; Maksymowicz, K.; Kobielarz, M.; Gamian, A.; Chwilkowska, A. Relationship between Calcification, Atherosclerosis and Matrix Proteins in the Human Aorta. Folia Histochem. Cytobiol. 2021, 59, 8–21. [Google Scholar] [CrossRef] [PubMed]
- Neubauer, K.; Zieger, B. Endothelial Cells and Coagulation. Cell Tissue Res. 2022, 387, 391–398. [Google Scholar] [CrossRef] [PubMed]
- Li, M.-T.; Ke, J.; Guo, S.-F.; Shan, L.-L.; Gong, J.-H.; Qiao, T.-C.; Tian, H.-Y.; Wu, Y.; Peng, Z.-Y.; Zeng, X.-Q.; et al. Huzhangqingmaiyin Protected Vascular Endothelial Cells against Cerebral Small Vessel Disease through Inhibiting Inflammation. J. Ethnopharmacol. 2024, 318, 116905. [Google Scholar] [CrossRef] [PubMed]
- He, B.; Zhan, Y.; Cai, C.; Yu, D.; Wei, Q.; Quan, L.; Huang, D.; Liu, Y.; Li, Z.; Liu, L.; et al. Common Molecular Mechanism and Immune Infiltration Patterns of Thoracic and Abdominal Aortic Aneurysms. Front. Immunol. 2022, 13, 1030976. [Google Scholar] [CrossRef] [PubMed]
- Allaire, E.; Schneider, F.; Saucy, F.; Dai, J.; Cochennec, F.; Michineau, S.; Zidi, M.; Becquemin, J.-P.; Kirsch, M.; Gervais, M. New Insight in Aetiopathogenesis of Aortic Diseases. Eur. J. Vasc. Endovasc. Surg. 2009, 37, 531–537. [Google Scholar] [CrossRef] [PubMed]
- Kuivaniemi, H.; Ryer, E.J.; Elmore, J.R.; Tromp, G. Understanding the Pathogenesis of Abdominal Aortic Aneurysms. Expert Rev. Cardiovasc. Ther. 2015, 13, 975–987. [Google Scholar] [CrossRef] [PubMed]
- Yap, Z.J.; Sharif, M.; Bashir, M. Is There an Immunogenomic Difference between Thoracic and Abdominal Aortic Aneurysms? J. Card. Surg. 2021, 36, 1520–1530. [Google Scholar] [CrossRef]
- Wu, H.; Xie, C.; Wang, R.; Cheng, J.; Xu, Q.; Zhao, H. Comparative Analysis of Thoracic and Abdominal Aortic Aneurysms across the Segment and Species at the Single-Cell Level. Front. Pharmacol. 2023, 13, 1095757. [Google Scholar] [CrossRef]
- Patel, R.; Hall, S.; Lanford, H.; Ward, N.; Grespin, R.T.; Figueroa, M.; Mattia, V.; Xiong, Y.; Mukherjee, R.; Jones, J.; et al. Signaling through the IL-6-STAT3 Pathway Promotes Proteolytically-Active Macrophage Accumulation Necessary for Development of Small AAA. Vasc. Endovasc. Surg. 2023, 57, 433–444. [Google Scholar] [CrossRef]
- Wang, X.; He, B.; Deng, Y.; Liu, J.; Zhang, Z.; Sun, W.; Gao, Y.; Liu, X.; Zhen, Y.; Ye, Z.; et al. Identification of a Biomarker and Immune Infiltration in Perivascular Adipose Tissue of Abdominal Aortic Aneurysm. Front. Physiol. 2022, 13, 977910. [Google Scholar] [CrossRef]
- Meekel, J.P.; Dias-Neto, M.; Bogunovic, N.; Conceição, G.; Sousa-Mendes, C.; Stoll, G.R.; Leite-Moreira, A.; Huynh, J.; Micha, D.; Eringa, E.C.; et al. Inflammatory Gene Expression of Human Perivascular Adipose Tissue in Abdominal Aortic Aneurysms. Eur. J. Vasc. Endovasc. Surg. 2021, 61, 1008–1016. [Google Scholar] [CrossRef] [PubMed]
- Nosalski, R.; Guzik, T.J. Perivascular Adipose Tissue Inflammation in Vascular Disease. Br. J. Pharmacol. 2017, 174, 3496–3513. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Yang, Y.; Sun, Y.; Wei, G.; Zheng, H.; Chen, Y.; Cai, D.; Li, C.; Ma, Y.; Lin, Z.; et al. Circular RNA Cdyl Promotes Abdominal Aortic Aneurysm Formation by Inducing M1 Macrophage Polarization and M1-Type Inflammation. Mol. Ther. J. Am. Soc. Gene Ther. 2022, 30, 915–931. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhao, Y.; Jin, H.; Qi, X.; He, J.; Huang, J.; Ding, Y.; Chen, W.; Wu, C.; Ding, X.; et al. TROVE2 Strengthens the Anti-Inflammatory Effect via Macrophage Polarization by Estrogen Induction in Abdominal Aortic Aneurysm. Life Sci. 2020, 242, 117207. [Google Scholar] [CrossRef]
- Lu, S.; White, J.V.; Nwaneshiudu, I.; Nwaneshiudu, A.; Monos, D.S.; Solomides, C.C.; Oleszak, E.L.; Platsoucas, C.D. Human Abdominal Aortic Aneurysm (AAA): Evidence for an Autoimmune Antigen-Driven Disease. Autoimmun. Rev. 2022, 21, 103164. [Google Scholar] [CrossRef] [PubMed]
- Gao, R.; Liu, D.; Guo, W.; Ge, W.; Fan, T.; Li, B.; Gao, P.; Liu, B.; Zheng, Y.; Wang, J. Meprin-α (Mep1A) Enhances TNF-α Secretion by Mast Cells and Aggravates Abdominal Aortic Aneurysms. Br. J. Pharmacol. 2020, 177, 2872–2885. [Google Scholar] [CrossRef] [PubMed]
- Miao, T.; Wang, T.; Feng, T.; Yuan, D.; Guo, Q.; Xiong, F.; Yang, Y.; Liu, L.; He, Z.; Huang, B.; et al. Activated Invariant Natural Killer T Cells Infiltrate Aortic Tissue as Key Participants in Abdominal Aortic Aneurysm Pathology. Immunology 2021, 164, 792–802. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Yang, Z.; Wang, S.; Que, Y.; Shu, X.; Wu, F.; Liu, G.; Li, S.; Hu, P.; Chen, H.; et al. Substitution of SERCA2 Cys674 Accelerates Aortic Aneurysm by Inducing Endoplasmic Reticulum Stress and Promoting Cell Apoptosis. Br. J. Pharmacol. 2022, 179, 4423–4439. [Google Scholar] [CrossRef]
- Han, B.; Yang, M.; Liu, Q.; Wang, G.; Hou, J. Long Noncoding RNA SBF2-AS1 Promotes Abdominal Aortic Aneurysm Formation through the miRNA-520f-3p/SMARCD1 Axis. Dis. Markers 2022, 2022, 4782361. [Google Scholar] [CrossRef]
- Jing, J.; Chang, M.; Jiang, S.; Wang, T.; Sun, Q.; Yang, J.; Ma, C.; Li, T. Clinical Value of Serum miR-1-3p as a Potential Circulating Biomarker for Abdominal Aortic Aneurysm. Ann. Med. 2023, 55, 2260395. [Google Scholar] [CrossRef]
- Zhao, L.; Ouyang, Y.; Bai, Y.; Gong, J.; Liao, H. miR-155-5p Inhibits the Viability of Vascular Smooth Muscle Cell via Targeting FOS and ZIC3 to Promote Aneurysm Formation. Eur. J. Pharmacol. 2019, 853, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Fang, Z.-M.; Yi, X.; Wei, X.; Jiang, D.-S. The Interaction between Ferroptosis and Inflammatory Signaling Pathways. Cell Death Dis. 2023, 14, 205. [Google Scholar] [CrossRef] [PubMed]
- Papayannopoulos, V. Neutrophil Extracellular Traps in Immunity and Disease. Nat. Rev. Immunol. 2018, 18, 134–147. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Liu, Y.; Wang, Z.; Zhang, L.; Xu, Y.; Li, Y.; Zhang, L.; Wang, G.; Yang, S.; Xue, G. Mesenchymal Stem Cell-Derived Extracellular Vesicles Protect against Abdominal Aortic Aneurysm Formation by Inhibiting NET-Induced Ferroptosis. Exp. Mol. Med. 2023, 55, 939–951. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Cao, Y.; Cao, W.; Jia, Y.; Lu, N. The Application of Ferroptosis in Diseases. Pharmacol. Res. 2020, 159, 104919. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wang, J.; Cai, D.; Li, X.; Zhong, L.; He, X.; Lin, Z.; Lai, Y.; Zheng, H.; Zhou, Y.; et al. Reactive Oxygen Species-Induced Long Intergenic Noncoding RNA P21 Accelerates Abdominal Aortic Aneurysm Formation by Promoting Secretary Smooth Muscle Cell Phenotypes. J. Mol. Cell. Cardiol. 2023, 174, 63–76. [Google Scholar] [CrossRef] [PubMed]
- Chang, Z.; Zhao, G.; Zhao, Y.; Lu, H.; Xiong, W.; Liang, W.; Sun, J.; Wang, H.; Zhu, T.; Rom, O.; et al. BAF60a Deficiency in Vascular Smooth Muscle Cells Prevents Abdominal Aortic Aneurysm by Reducing Inflammation and Extracellular Matrix Degradation. Arterioscler. Thromb. Vasc. Biol. 2020, 40, 2494–2507. [Google Scholar] [CrossRef] [PubMed]
- Kostyunin, A.; Mukhamadiyarov, R.; Glushkova, T.; Bogdanov, L.; Shishkova, D.; Osyaev, N.; Ovcharenko, E.; Kutikhin, A. Ultrastructural Pathology of Atherosclerosis, Calcific Aortic Valve Disease, and Bioprosthetic Heart Valve Degeneration: Commonalities and Differences. Int. J. Mol. Sci. 2020, 21, 7434. [Google Scholar] [CrossRef]
- Jana, S.; Hu, M.; Shen, M.; Kassiri, Z. Extracellular Matrix, Regional Heterogeneity of the Aorta, and Aortic Aneurysm. Exp. Mol. Med. 2019, 51, 1–15. [Google Scholar] [CrossRef]
- Fu, Y.; Liu, H.; Li, K.; Wei, P.; Alam, N.; Deng, J.; Li, M.; Wu, H.; He, X.; Hou, H.; et al. C-Reactive Protein Deficiency Ameliorates Experimental Abdominal Aortic Aneurysms. Front. Immunol. 2023, 14, 1233807. [Google Scholar] [CrossRef]
- Nevado, R.M.; Hamczyk, M.R.; Gonzalo, P.; Andrés-Manzano, M.J.; Andrés, V. Premature Vascular Aging with Features of Plaque Vulnerability in an Atheroprone Mouse Model of Hutchinson–Gilford Progeria Syndrome with Ldlr Deficiency. Cells 2020, 9, 2252. [Google Scholar] [CrossRef] [PubMed]
- Salmasi, M.Y.; Morris-Rosendahl, D.; Jarral, O.A.; Rosendahl, U.; Asimakopoulos, G.; Raja, S.; Aragon-Martin, J.A.; Child, A.; Pepper, J.; Oo, A.; et al. Determining the Genetic Contribution in Patients with Non-Syndromic Ascending Thoracic Aortic Aneurysms: Correlation with Findings from Computational Pathology. Int. J. Cardiol. 2022, 366, 1–9. [Google Scholar] [CrossRef]
- Doherty, E.L.; Aw, W.Y.; Warren, E.C.; Hockenberry, M.; Whitworth, C.P.; Krohn, G.; Howell, S.; Diekman, B.O.; Legant, W.R.; Nia, H.T.; et al. Patient-Derived Extracellular Matrix Demonstrates Role of COL3A1 in Blood Vessel Mechanics. Acta Biomater. 2023, 166, 346–359. [Google Scholar] [CrossRef] [PubMed]
- Errichiello, E.; Malara, A.; Grimod, G.; Avolio, L.; Balduini, A.; Zuffardi, O. Low Penetrance COL5A1 Variants in a Young Patient with Intracranial Aneurysm and Very Mild Signs of Ehlers-Danlos Syndrome. Eur. J. Med. Genet. 2021, 64, 104099. [Google Scholar] [CrossRef] [PubMed]
- Deleeuw, V.; Carlson, E.; Renard, M.; Zientek, K.D.; Wilmarth, P.A.; Reddy, A.P.; Manalo, E.C.; Tufa, S.F.; Keene, D.R.; Olbinado, M.; et al. Unraveling the Role of TGFβ Signaling in Thoracic Aortic Aneurysm and Dissection Using Fbn1 Mutant Mouse Models. Matrix Biol. J. Int. Soc. Matrix Biol. 2023, 123, 17–33. [Google Scholar] [CrossRef]
- Watson, S.R.; Cooper, K.M.; Liu, P.; Gharraee, N.; Du, L.; Han, S.M.; Peña, E.A.; Sutton, M.A.; Eberth, J.F.; Lessner, S.M. Diet Alters Age-Related Remodeling of Aortic Collagen in Mice Susceptible to Atherosclerosis. Am. J. Physiol.-Heart Circ. Physiol. 2021, 320, H52–H65. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.M.; Douglas, G.; Bendall, J.K.; McNeill, E.; Crabtree, M.J.; Hale, A.B.; Mai, A.; Li, J.-M.; McAteer, M.A.; Schneider, J.E.; et al. Endothelial Cell–Specific Reactive Oxygen Species Production Increases Susceptibility to Aortic Dissection. Circulation 2014, 129, 2661–2672. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhao, X.; Zhao, G.; Guo, Y.; Lu, H.; Mu, W.; Zhong, J.; Garcia-Barrio, M.; Zhang, J.; Chen, Y.E.; et al. PRDM16 Deficiency in Vascular Smooth Muscle Cells Aggravates Abdominal Aortic Aneurysm. JCI Insight 2023, 8, e167041. [Google Scholar] [CrossRef]
- Wang, S.; Yuan, Q.; Zhao, W.; Zhou, W. Circular RNA RBM33 Contributes to Extracellular Matrix Degradation via miR-4268/EPHB2 Axis in Abdominal Aortic Aneurysm. PeerJ 2021, 9, e12232. [Google Scholar] [CrossRef]
- Teti, G.; Chiarini, F.; Mazzotti, E.; Ruggeri, A.; Carano, F.; Falconi, M. Cellular Senescence in Vascular Wall Mesenchymal Stromal Cells, a Possible Contribution to the Development of Aortic Aneurysm. Mech. Ageing Dev. 2021, 197, 111515. [Google Scholar] [CrossRef]
- Gadanec, L.K.; McSweeney, K.R.; Kubatka, P.; Caprnda, M.; Gaspar, L.; Prosecky, R.; Dragasek, J.; Kruzliak, P.; Apostolopoulos, V.; Zulli, A. Angiotensin II Constricts Mouse Iliac Arteries: Possible Mechanism for Aortic Aneurysms. Mol. Cell. Biochem. 2023. [Google Scholar] [CrossRef]
- Tomimori, Y.; Manno, A.; Tanaka, T.; Futamura-Takahashi, J.; Muto, T.; Nagahira, K. ASB17061, a Novel Chymase Inhibitor, Prevented the Development of Angiotensin II-Induced Abdominal Aortic Aneurysm in Apolipoprotein E-Deficient Mice. Eur. J. Pharmacol. 2019, 856, 172403. [Google Scholar] [CrossRef] [PubMed]
- Poto, R.; Patella, V.; Criscuolo, G.; Marone, G.; Coscioni, E.; Varricchi, G. Autoantibodies to IgE Can Induce the Release of Proinflammatory and Vasoactive Mediators from Human Cardiac Mast Cells. Clin. Exp. Med. 2023, 23, 1265–1276. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, M.R.; e Melo, R.G.; Silvestre, L.; e Fernandes, R.F.; Martins, C.; Pedro, L.M. A Literature Review on Pharmacologic Therapy for Abdominal Aortic Aneurysms. Angiol. Cir. Vasc. 2023, 19, 44–46. [Google Scholar] [CrossRef]
- Puchenkova, O.A.; Soldatov, V.O.; Belykh, A.E.; Bushueva, O.; Piavchenko, G.A.; Venediktov, A.A.; Shakhpazyan, N.K.; Deykin, A.V.; Korokin, M.V.; Pokrovskiy, M.V. Cytokines in Abdominal Aortic Aneurysm: Master Regulators With Clinical Application. Biomark. Insights 2022, 17, 11772719221095676. [Google Scholar] [CrossRef] [PubMed]
- Tingting, T.; Wenjing, F.; Qian, Z.; Hengquan, W.; Simin, Z.; Zhisheng, J.; Shunlin, Q. The TGF-β Pathway Plays a Key Role in Aortic Aneurysms. Clin. Chim. Acta Int. J. Clin. Chem. 2020, 501, 222–228. [Google Scholar] [CrossRef]
- Evers-van Gogh, I.J.A.; Oteng, A.-B.; Alex, S.; Hamers, N.; Catoire, M.; Stienstra, R.; Kalkhoven, E.; Kersten, S. Muscle-Specific Inflammation Induced by MCP-1 Overexpression Does Not Affect Whole-Body Insulin Sensitivity in Mice. Diabetologia 2016, 59, 624–633. [Google Scholar] [CrossRef]
- Lowis, C.; Ramara Winaya, A.; Kumari, P.; Rivera, C.F.; Vlahos, J.; Hermantara, R.; Pratama, M.Y.; Ramkhelawon, B. Mechanosignals in Abdominal Aortic Aneurysms. Front. Cardiovasc. Med. 2023, 9, 1021934. [Google Scholar] [CrossRef]
- Jin, Z.; Deng, H.; Xiong, S.; Gao, L. Perspective of SGLT2i in the Treatment of Abdominal Aortic Aneurysms. J. Cardiovasc. Pharmacol. 2023, 81, 241–247. [Google Scholar] [CrossRef]
- Salmon, M. NADPH Oxidases in Aortic Aneurysms. Antioxidants 2022, 11, 1830. [Google Scholar] [CrossRef]
- Wu, T.; Li, N.; Zhang, Q.; Liu, R.; Zhao, H.; Fan, Z.; Zhuo, L.; Yang, Y.; Xu, Y. MKL1 Fuels ROS-Induced Proliferation of Vascular Smooth Muscle Cells by Modulating FOXM1 Transcription. Redox Biol. 2022, 59, 102586. [Google Scholar] [CrossRef]
- Groeneveld, M.E.; Struik, J.A.; Musters, R.J.P.; Tangelder, G.J.; Koolwijk, P.; Niessen, H.W.; Hoksbergen, A.W.J.; Wisselink, W.; Yeung, K.K. The Potential Role of Neutrophil Gelatinase-Associated Lipocalin in the Development of Abdominal Aortic Aneurysms. Ann. Vasc. Surg. 2019, 57, 210–219. [Google Scholar] [CrossRef] [PubMed]
- Klopf, J.; Brostjan, C.; Neumayer, C.; Eilenberg, W. Neutrophils as Regulators and Biomarkers of Cardiovascular Inflammation in the Context of Abdominal Aortic Aneurysms. Biomedicines 2021, 9, 1236. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Yan, H.; Cannon, J.L.; Springer, L.E.; Green, J.M.; Pham, C.T.N. CD43-Mediated IFN-γ Production by CD8+ T Cells Promotes Abdominal Aortic Aneurysm in Mice. J. Immunol. 2013, 190, 5078–5085. [Google Scholar] [CrossRef] [PubMed]
- Stepien, K.L.; Bajdak-Rusinek, K.; Fus-Kujawa, A.; Kuczmik, W.; Gawron, K. Role of Extracellular Matrix and Inflammation in Abdominal Aortic Aneurysm. Int. J. Mol. Sci. 2022, 23, 11078. [Google Scholar] [CrossRef] [PubMed]
- Iwańczyk, S.; Lehmann, T.; Grygier, M.; Woźniak, P.; Lesiak, M.; Araszkiewicz, A. Serum Matrix Metalloproteinase-8 Level in Patients with Coronary Artery Abnormal Dilatation. Pol. Arch. Intern. Med. 2022, 132, 16241. [Google Scholar] [CrossRef] [PubMed]
- Surma, S.; Stolorz, K.; Sierka, O.; Sieroń, A.; Lesiak, M. The Role of Selected Metalloproteinases and Some Genetic Factors in the Pathogenesis of Abdominal Aortic Aneurysm. Acta Angiol. 2021, 1, 22–31. [Google Scholar] [CrossRef]
- Berman, A.; Romary, D.; Kerr, K.; Gorazd, N.; Wigand, M.; Patnaik, S.; Finol, E.; Cox, A.; Goergen, C. Experimental Aortic Aneurysm Severity and Growth Depend on Topical Elastase Concentration and Lysyl Oxidase Inhibition. Sci. Rep. 2022, 12, 99. [Google Scholar] [CrossRef]
- Remus, E.W.; O’Donnell, R.E.; Rafferty, K.; Weiss, D.; Joseph, G.; Csiszar, K.; Fong, S.F.T.; Taylor, W.R. The Role of Lysyl Oxidase Family Members in the Stabilization of Abdominal Aortic Aneurysms. Am. J. Physiol.-Heart Circ. Physiol. 2012, 303, H1067–H1075. [Google Scholar] [CrossRef]
- Bumdelger, B.; Kokubo, H.; Kamata, R.; Fujii, M.; Yoshimura, K.; Aoki, H.; Orita, Y.; Ishida, T.; Ohtaki, M.; Nagao, M.; et al. Osteoprotegerin Prevents Development of Abdominal Aortic Aneurysms. PLoS ONE 2016, 11, e0147088. [Google Scholar] [CrossRef]
- Karasaki, K.; Kokubo, H.; Bumdelger, B.; Kaji, N.; Sakai, C.; Ishida, M.; Yoshizumi, M. Angiotensin II Type 1 Receptor Blocker Prevents Abdominal Aortic Aneurysm Progression in Osteoprotegerin-Deficient Mice via Upregulation of Angiotensin (1–7). J. Am. Heart Assoc. Cardiovasc. Cerebrovasc. Dis. 2023, 12, e027589. [Google Scholar] [CrossRef]
- Herrero-Cervera, A.; Espinós-Estévez, C.; Martín-Vañó, S.; Taberner-Cortés, A.; Aguilar-Ballester, M.; Vinué, Á.; Piqueras, L.; Martínez-Hervás, S.; González-Navarro, H. Dissecting Abdominal Aortic Aneurysm Is Aggravated by Genetic Inactivation of LIGHT (TNFSF14). Biomedicines 2021, 9, 1518. [Google Scholar] [CrossRef]
- Wang, S.K.; Green, L.A.; Gutwein, A.R.; Gupta, A.K.; Babbey, C.M.; Motaganahalli, R.L.; Fajardo, A.; Murphy, M.P. Osteopontin May Be a Driver of Abdominal Aortic Aneurysm Formation. J. Vasc. Surg. 2018, 68, 22S–29S. [Google Scholar] [CrossRef] [PubMed]
- Sajeesh, S.; Camardo, A.; Dahal, S.; Ramamurthi, A. Surface-Functionalized Stem Cell-Derived Extracellular Vesicles for Vascular Elastic Matrix Regenerative Repair. Mol. Pharm. 2023, 20, 2801–2813. [Google Scholar] [CrossRef]
- Zhang, X.; Luo, S.; Wang, M.; Shi, G.-P. Cysteinyl Cathepsins in Cardiovascular Diseases. Biochim. Biophys. Acta Proteins Proteom. 2020, 1868, 140360. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.H.; LeMaire, S.A.; Webb, N.R.; Cassis, L.A.; Daugherty, A.; Lu, H.S. Aortic Aneurysms and Dissections Series. Arterioscler. Thromb. Vasc. Biol. 2020, 40, e37–e46. [Google Scholar] [CrossRef] [PubMed]
- Golledge, J. Abdominal Aortic Aneurysm: Update on Pathogenesis and Medical Treatments. Nat. Rev. Cardiol. 2019, 16, 225–242. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Zhang, K.; Liu, Z.; Xu, Q.; You, B.; Li, C.; Cao, J.; Zhou, H.; Li, X.; Chen, J.; et al. VPO1 Modulates Vascular Smooth Muscle Cell Phenotypic Switch by Activating Extracellular Signal-Regulated Kinase 1/2 (ERK 1/2) in Abdominal Aortic Aneurysms. J. Am. Heart Assoc. 2018, 7, e010069. [Google Scholar] [CrossRef]
- Zalewski, D.; Chmiel, P.; Kołodziej, P.; Borowski, G.; Feldo, M.; Kocki, J.; Bogucka-Kocka, A. Dysregulations of Key Regulators of Angiogenesis and Inflammation in Abdominal Aortic Aneurysm. Int. J. Mol. Sci. 2023, 24, 12087. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Taghi Khani, A.; Sanchez Ortiz, A.; Swaminathan, S. GM-CSF: A Double-Edged Sword in Cancer Immunotherapy. Front. Immunol. 2022, 13, 901277. [Google Scholar] [CrossRef]
- Lazarus, H.M.; Ragsdale, C.E.; Gale, R.P.; Lyman, G.H. Sargramostim (Rhu GM-CSF) as Cancer Therapy (Systematic Review) and An Immunomodulator. A Drug Before Its Time? Front. Immunol. 2021, 12, 706186. [Google Scholar] [CrossRef]
- United States Food and Drug Administration. Executive Order 13944 List of Essential Medicines, Medical Countermeasures, and Critical Inputs; United States Food and Drug Administration: Silver Spring, MD, USA, 2023.
- Mihalik, N.E.; Wen, S.; Driesschaert, B.; Eubank, T.D. Formulation and in Vitro Characterization of PLGA/PLGA-PEG Nanoparticles Loaded with Murine Granulocyte-Macrophage Colony-Stimulating Factor. AAPS PharmSciTech 2021, 22, 191. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.; Postigo-Fernandez, J.; Kim, K.; Zhu, C.; Yu, J.; Meroni, M.; Mayfield, B.; Bartolomé, A.; Dapito, D.H.; Ferrante, A.W.; et al. Notch-Mediated Hepatocyte MCP-1 Secretion Causes Liver Fibrosis. JCI Insight 2023, 8, e165369. [Google Scholar] [CrossRef] [PubMed]
- Pulito-Cueto, V.; Remuzgo-Martínez, S.; Genre, F.; Atienza-Mateo, B.; Mora-Cuesta, V.M.; Iturbe-Fernández, D.; Lera-Gómez, L.; Sebastián Mora-Gil, M.; Prieto-Peña, D.; Portilla, V.; et al. Elevated VCAM-1, MCP-1 and ADMA Serum Levels Related to Pulmonary Fibrosis of Interstitial Lung Disease Associated with Rheumatoid Arthritis. Front. Mol. Biosci. 2022, 9, 1056121. [Google Scholar] [CrossRef] [PubMed]
- Angelova, P.R.; Dinkova-Kostova, A.T.; Abramov, A.Y. Assessment of ROS Production in the Mitochondria of Live Cells. Methods Mol. Biol. Clifton NJ 2021, 2202, 33–42. [Google Scholar] [CrossRef]
- Lennicke, C.; Cochemé, H.M. Redox Metabolism: ROS as Specific Molecular Regulators of Cell Signaling and Function. Mol. Cell 2021, 81, 3691–3707. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Yang, Q.; Fang, G.; Li, B.-S.; Wu, D.-B.; Guo, W.-J.; Hong, S.-S.; Hong, L. Collagen Metabolic Disorder Induced by Oxidative Stress in Human Uterosacral Ligament-derived Fibroblasts: A Possible Pathophysiological Mechanism in Pelvic Organ Prolapse. Mol. Med. Rep. 2016, 13, 2999–3008. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Fu, Y.; Xiao, H.; Song, Y.; Chen, R.; Shen, J.; An, X.; Shen, Q.; Li, Z.; Zhang, Y. Cardiac Fibrosis Alleviated by Exercise Training Is AMPK-Dependent. PLoS ONE 2015, 10, e0129971. [Google Scholar] [CrossRef]
- Spada, S.; Tocci, A.; Di Modugno, F.; Nisticò, P. Fibronectin as a Multiregulatory Molecule Crucial in Tumor Matrisome: From Structural and Functional Features to Clinical Practice in Oncology. J. Exp. Clin. Cancer Res. 2021, 40, 102. [Google Scholar] [CrossRef]
- Xu, Y.; Yang, S.; Xue, G. The Role of Long Non-Coding RNA in Abdominal Aortic Aneurysm. Front. Genet. 2023, 14, 1153899. [Google Scholar] [CrossRef]
- Guo, F.; Wang, E.; Yang, Y.; Mao, Y.; Liu, C.; Bu, W.; Li, P.; Zhao, L.; Jin, Q.; Liu, B.; et al. A Natural Biomineral for Enhancing the Biomineralization and Cell Response of 3D Printed Polylactic Acid Bone Scaffolds. Int. J. Biol. Macromol. 2023, 242, 124728. [Google Scholar] [CrossRef] [PubMed]
- Lee, L.Y.-H.; Oldham, W.M.; He, H.; Wang, R.; Mulhern, R.; Handy, D.E.; Loscalzo, J. Interferon-γ Impairs Human Coronary Artery Endothelial Glucose Metabolism by Tryptophan Catabolism and Activates Fatty Acid Oxidation. Circulation 2021, 144, 1612–1628. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Jiang, Z.; Wang, S.; Zhang, P. IFN-γ Restores the Impaired Function of RNase L and Induces Mitochondria-Mediated Apoptosis in Lung Cancer. Cell Death Dis. 2019, 10, 642. [Google Scholar] [CrossRef] [PubMed]
- Park, S.-H.; Hwang, J.-S.; Oh, S.-H.; Shin, Y.-J. MiR-302a Regenerates Human Corneal Endothelial Cells against IFN-γ-Induced Cell Death. Cells 2022, 12, 36. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wei, G.; Cheng, S.; Wang, D.; Ma, J.; Xin, S. Circulatory CD4-Positive T-Lymphocyte Imbalance Mediated by Homocysteine-Induced AIM2 and NLRP1 Inflammasome Upregulation and Activation Is Associated with Human Abdominal Aortic Aneurysm. J. Vasc. Res. 2020, 57, 276–290. [Google Scholar] [CrossRef] [PubMed]
- Miao, Y.; Zhao, Y.; Han, L.; Ma, X.; Deng, J.; Yang, J.; Lü, S.; Shao, F.; Kong, W.; Wang, W.; et al. NSun2 Regulates Aneurysm Formation by Promoting Autotaxin Expression and T Cell Recruitment. Cell. Mol. Life Sci. CMLS 2021, 78, 1709–1727. [Google Scholar] [CrossRef] [PubMed]
- Murali Krishna, S.; Morton, S.K.; Li, J.; Golledge, J. Risk Factors and Mouse Models of Abdominal Aortic Aneurysm Rupture. Int. J. Mol. Sci. 2020, 21, 7250. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Meng, X.; Guo, Z. Elastin Structure, Synthesis, Regulatory Mechanism and Relationship With Cardiovascular Diseases. Front. Cell Dev. Biol. 2021, 9, 596702. [Google Scholar] [CrossRef]
- Kobielarz, M. Effect of Collagen Fibres and Elastic Lamellae Content on the Mechanical Behaviour of Abdominal Aortic Aneurysms. Acta Bioeng. Biomech. 2020, 22, 9–21. [Google Scholar] [CrossRef]
- Quintana, R.A.; Taylor, W.R. Cellular Mechanisms of Aortic Aneurysm Formation. Circ. Res. 2019, 124, 607–618. [Google Scholar] [CrossRef]
- Pasta, S.; Agnese, V.; Gallo, A.; Cosentino, F.; Di Giuseppe, M.; Gentile, G.; Raffa, G.M.; Maalouf, J.F.; Michelena, H.I.; Bellavia, D.; et al. Shear Stress and Aortic Strain Associations With Biomarkers of Ascending Thoracic Aortic Aneurysm. Ann. Thorac. Surg. 2020, 110, 1595–1604. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, W.; Li, L.; Khalil, R.A. MMPs and ADAMs/ADAMTS Inhibition Therapy of Abdominal Aortic Aneurysm. Life Sci. 2020, 253, 117659. [Google Scholar] [CrossRef] [PubMed]
- Steffensen, L.B.; Stubbe, J.; Lindholt, J.S.; Beck, H.C.; Overgaard, M.; Bloksgaard, M.; Genovese, F.; Holm Nielsen, S.; Tha, M.L.T.; Bang-Moeller, S.K.; et al. Basement Membrane Collagen IV Deficiency Promotes Abdominal Aortic Aneurysm Formation. Sci. Rep. 2021, 11, 12903. [Google Scholar] [CrossRef] [PubMed]
- Su, G.-J.; Gao, J.; Wu, C.-W.; Zou, J.-F.; Zhang, D.; Zhu, D.-L.; Liu, J.; Zhang, J.-H.; Huang, X.-J. Serum Levels of MMP-8 and MMP-9 as Markers in Chronic Subdural Hematoma. J. Clin. Med. 2022, 11, 902. [Google Scholar] [CrossRef] [PubMed]
- Bararu Bojan (Bararu), I.; Pleșoianu, C.E.; Badulescu, O.V.; Vladeanu, M.C.; Badescu, M.C.; Iliescu, D.; Bojan, A.; Ciocoiu, M. Molecular and Cellular Mechanisms Involved in Aortic Wall Aneurysm Development. Diagnostics 2023, 13, 253. [Google Scholar] [CrossRef] [PubMed]
- Migacz, M.; Janoska-Gawrońska, A.; Holecki, M.; Chudek, J. The Role of Osteoprotegerin in the Development, Progression and Management of Abdominal Aortic Aneurysms. Open Med. 2020, 15, 457–463. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhang, Y.; Song, W.; Sun, Y.; Jiang, Y. Osteopontin N-Terminal Function in an Abdominal Aortic Aneurysm From Apolipoprotein E-Deficient Mice. Front. Cell Dev. Biol. 2021, 9, 681790. [Google Scholar] [CrossRef]
- Cao, G.; Xuan, X.; Hu, J.; Zhang, R.; Jin, H.; Dong, H. How Vascular Smooth Muscle Cell Phenotype Switching Contributes to Vascular Disease. Cell Commun. Signal. 2022, 20, 180. [Google Scholar] [CrossRef]
- Rombouts, K.B.; van Merrienboer, T.A.R.; Ket, J.C.F.; Bogunovic, N.; van der Velden, J.; Yeung, K.K. The Role of Vascular Smooth Muscle Cells in the Development of Aortic Aneurysms and Dissections. Eur. J. Clin. Investig. 2022, 52, e13697. [Google Scholar] [CrossRef]
- Lv, P.; Yin, Y.-J.; Kong, P.; Cao, L.; Xi, H.; Wang, N.; Yang, H.-C.; Lv, Y.-H.; Chen, N.; Wang, R.; et al. SM22α Loss Contributes to Apoptosis of Vascular Smooth Muscle Cells via Macrophage-Derived circRasGEF1B. Oxidative Med. Cell. Longev. 2021, 2021, 5564884. [Google Scholar] [CrossRef]
- Petsophonsakul, P.; Furmanik, M.; Forsythe, R.; Dweck, M.; Schurink, G.W.; Natour, E.; Reutelingsperger, C.; Jacobs, M.; Mees, B.; Schurgers, L. Role of Vascular Smooth Muscle Cell Phenotypic Switching and Calcification in Aortic Aneurysm Formation. Arterioscler. Thromb. Vasc. Biol. 2019, 39, 1351–1368. [Google Scholar] [CrossRef]
- Lin, J.; Chen, S.; Yao, Y.; Yan, M. Status of Diagnosis and Therapy of Abdominal Aortic Aneurysms. Front. Cardiovasc. Med. 2023, 10, 1199804. [Google Scholar] [CrossRef]
- Kobeissi, E.; Hibino, M.; Pan, H.; Aune, D. Blood Pressure, Hypertension and the Risk of Abdominal Aortic Aneurysms: A Systematic Review and Meta-Analysis of Cohort Studies. Eur. J. Epidemiol. 2019, 34, 547–555. [Google Scholar] [CrossRef]
- Wu, M.; Zhang, S.; Zhang, W.; Zhou, Y.; Guo, Z.; Fang, Y.; Yang, Y.; Shen, Z.; Lian, D.; Shen, A.; et al. Qingda Granule Ameliorates Vascular Remodeling and Phenotypic Transformation of Adventitial Fibroblasts via Suppressing the TGF-Β1/Smad2/3 Pathway. J. Ethnopharmacol. 2023, 313, 116535. [Google Scholar] [CrossRef]
- Yu, N.; Shen, A.; Chu, J.; Huang, Y.; Zhang, L.; Lin, S.; Cai, Q.; Sankararaman, S.; Sferra, T.J.; Chen, Y.; et al. Qingda Granule Inhibits Angiotensin Ⅱ Induced VSMCs Proliferation through MAPK and PI3K/AKT Pathways. J. Ethnopharmacol. 2020, 258, 112767. [Google Scholar] [CrossRef]
- Dai, Z.; Zhu, M.M.; Peng, Y.; Jin, H.; Machireddy, N.; Qian, Z.; Zhang, X.; Zhao, Y.-Y. Endothelial and Smooth Muscle Cell Interaction via FoxM1 Signaling Mediates Vascular Remodeling and Pulmonary Hypertension. Am. J. Respir. Crit. Care Med. 2018, 198, 788–802. [Google Scholar] [CrossRef]
- Xu, H.; Wang, C.; Song, T.-T.; Liu, Y.; Dong, C.-W. Effects of Ziyin Qianyang Formula on Renal Fibrosis through the TGF-Β1/Smads Signaling Pathway in Spontaneously Hypertensive Rats. Evid.-Based Complement. Altern. Med. ECAM 2022, 2022, 6088673. [Google Scholar] [CrossRef]
- Hinterseher, I.; Tromp, G.; Kuivaniemi, H. Genes and Abdominal Aortic Aneurysm. Ann. Vasc. Surg. 2011, 25, 388–412. [Google Scholar] [CrossRef]
- Acharya, M.; Maselli, D.; Mariscalco, G. Genetic Screening in Heritable Thoracic Aortic Disease—Rationale, Potentials and Pitfalls. Indian J. Thorac. Cardiovasc. Surg. 2022, 38, 24–35. [Google Scholar] [CrossRef]
- Rodrigues Bento, J.; Meester, J.; Luyckx, I.; Peeters, S.; Verstraeten, A.; Loeys, B. The Genetics and Typical Traits of Thoracic Aortic Aneurysm and Dissection. Annu. Rev. Genom. Hum. Genet. 2022, 23, 223–253. [Google Scholar] [CrossRef]
- Hannuksela, M.; Stattin, E.-L.; Johansson, B.; Carlberg, B. Screening for Familial Thoracic Aortic Aneurysms with Aortic Imaging Does Not Detect All Potential Carriers of the Disease. AORTA 2015, 3, 1–8. [Google Scholar] [CrossRef]
- Ostberg, N.P.; Zafar, M.A.; Ziganshin, B.A.; Elefteriades, J.A. The Genetics of Thoracic Aortic Aneurysms and Dissection: A Clinical Perspective. Biomolecules 2020, 10, 182. [Google Scholar] [CrossRef]
- Proost, D.; Vandeweyer, G.; Meester, J.A.N.; Salemink, S.; Kempers, M.; Ingram, C.; Peeters, N.; Saenen, J.; Vrints, C.; Lacro, R.V.; et al. Performant Mutation Identification Using Targeted Next-Generation Sequencing of 14 Thoracic Aortic Aneurysm Genes. Hum. Mutat. 2015, 36, 808–814. [Google Scholar] [CrossRef]
- van de Luijtgaarden, K.M.; Heijsman, D.; Maugeri, A.; Weiss, M.M.; Verhagen, H.J.M.; IJpma, A.; Brüggenwirth, H.T.; Majoor-Krakauer, D. First Genetic Analysis of Aneurysm Genes in Familial and Sporadic Abdominal Aortic Aneurysm. Hum. Genet. 2015, 134, 881–893. [Google Scholar] [CrossRef]
- Renard, M.; Francis, C.; Ghosh, R.; Scott, A.F.; Witmer, P.D.; Adès, L.C.; Andelfinger, G.U.; Arnaud, P.; Boileau, C.; Callewaert, B.L.; et al. Clinical Validity of Genes for Heritable Thoracic Aortic Aneurysm and Dissection. J. Am. Coll. Cardiol. 2018, 72, 605–615. [Google Scholar] [CrossRef]
- Milewicz, D.M.; Braverman, A.C.; De Backer, J.; Morris, S.A.; Boileau, C.; Maumenee, I.H.; Jondeau, G.; Evangelista, A.; Pyeritz, R.E. Marfan Syndrome. Nat. Rev. Dis. Primers 2021, 7, 64. [Google Scholar] [CrossRef]
- Isselbacher, E.M.; Lino Cardenas, C.L.; Lindsay, M.E. Hereditary Influence in Thoracic Aortic Aneurysm and Dissection. Circulation 2016, 133, 2516–2528. [Google Scholar] [CrossRef]
- De Cario, R.; Sticchi, E.; Lucarini, L.; Attanasio, M.; Nistri, S.; Marcucci, R.; Pepe, G.; Giusti, B. Role of TGFBR1 and TGFBR2 Genetic Variants in Marfan Syndrome. J. Vasc. Surg. 2018, 68, 225–233.e5. [Google Scholar] [CrossRef]
- Gouda, P.; Kay, R.; Habib, M.; Aziz, A.; Aziza, E.; Welsh, R. Clinical Features and Complications of Loeys-Dietz Syndrome: A Systematic Review. Int. J. Cardiol. 2022, 362, 158–167. [Google Scholar] [CrossRef]
- Dittman, J.M.; Saldana-Ruiz, N.; Newhall, K.; Byers, P.H.; Starnes, B.W.; Shalhub, S. Open Repair of Abdominal Aortic Aneurysms in Patients with Vascular Ehlers-Danlos Syndrome. J. Vasc. Surg. Cases Innov. Tech. 2023, 9, 101194. [Google Scholar] [CrossRef]
- Faggion Vinholo, T.; Zafar, M.A.; Ziganshin, B.A.; Elefteriades, J.A. Nonsyndromic Thoracic Aortic Aneurysms and Dissections—Is Screening Possible? Semin. Thorac. Cardiovasc. Surg. 2019, 31, 628–634. [Google Scholar] [CrossRef]
- Renard, M.; Callewaert, B.; Baetens, M.; Campens, L.; MacDermot, K.; Fryns, J.-P.; Bonduelle, M.; Dietz, H.C.; Gaspar, I.M.; Cavaco, D.; et al. Novel MYH11 and ACTA2 Mutations Reveal a Role for Enhanced TGFβ Signaling in FTAAD. Int. J. Cardiol. 2013, 165, 314–321. [Google Scholar] [CrossRef]
- Salmasi, M.Y.; Alwis, S.; Cyclewala, S.; Jarral, O.A.; Mohamed, H.; Mozalbat, D.; Nienaber, C.A.; Athanasiou, T.; Morris-Rosendahl, D.; Moore, J., Jr.; et al. The Genetic Basis of Thoracic Aortic Disease: The Future of Aneurysm Classification? Hell. J. Cardiol. 2023, 69, 41–50. [Google Scholar] [CrossRef]
- Hiratzka, L.F.; Bakris, G.L.; Beckman, J.A.; Bersin, R.M.; Carr, V.F.; Casey, D.E.; Eagle, K.A.; Hermann, L.K.; Isselbacher, E.M.; Kazerooni, E.A.; et al. 2010 ACCF/AHA/AATS/ACR/ASA/SCA/SCAI/SIR/STS/SVM Guidelines for the Diagnosis and Management of Patients With Thoracic Aortic Disease. Circulation 2010, 121, e266–e369. [Google Scholar] [CrossRef]
- Negishi, K.; Aizawa, K.; Shindo, T.; Suzuki, T.; Sakurai, T.; Saito, Y.; Miyakawa, T.; Tanokura, M.; Kataoka, Y.; Maeda, M.; et al. An Myh11 Single Lysine Deletion Causes Aortic Dissection by Reducing Aortic Structural Integrity and Contractility. Sci. Rep. 2022, 12, 8844. [Google Scholar] [CrossRef]
- Wallace, S.E.; Regalado, E.S.; Gong, L.; Janda, A.L.; Guo, D.; Russo, C.F.; Kulmacz, R.J.; Hanna, N.; Jondeau, G.; Boileau, C.; et al. MYLK Pathogenic Variants Aortic Disease Presentation, Pregnancy Risk, and Characterization of Pathogenic Missense Variants. Genet. Med. 2019, 21, 144–151. [Google Scholar] [CrossRef]
- Linné, A.; Lindström, D.; Hultgren, R. High Prevalence of Abdominal Aortic Aneurysms in Brothers and Sisters of Patients despite a Low Prevalence in the Population. J. Vasc. Surg. 2012, 56, 305–310. [Google Scholar] [CrossRef]
- Sakalihasan, N.; Defraigne, J.-O.; Kerstenne, M.-A.; Cheramy-Bien, J.-P.; Smelser, D.T.; Tromp, G.; Kuivaniemi, H. Family Members of Patients with Abdominal Aortic Aneurysms Are at Increased Risk for Aneurysms: Analysis of 618 Probands and Their Families from the Liège AAA Family Study. Ann. Vasc. Surg. 2014, 28, 787–797. [Google Scholar] [CrossRef]
- Duan, X.-Y.; Guo, D.; Regalado, E.S.; Shen, H.; Coselli, J.S.; Estrera, A.L.; Safi, H.J.; Bamshad, M.J.; Nickerson, D.A.; LeMaire, S.A.; et al. SMAD4 Rare Variants in Individuals and Families with Thoracic Aortic Aneurysms and Dissections. Eur. J. Hum. Genet. 2019, 27, 1054–1060. [Google Scholar] [CrossRef]
- Da Ros, F.; Carnevale, R.; Cifelli, G.; Bizzotto, D.; Casaburo, M.; Perrotta, M.; Carnevale, L.; Vinciguerra, I.; Fardella, S.; Iacobucci, R.; et al. Targeting Interleukin-1β Protects from Aortic Aneurysms Induced by Disrupted Transforming Growth Factor β Signaling. Immunity 2017, 47, 959–973.e9. [Google Scholar] [CrossRef]
- Zhang, F.; Li, K.; Zhang, W.; Zhao, Z.; Chang, F.; Du, J.; Zhang, X.; Bao, K.; Zhang, C.; Shi, L.; et al. Ganglioside GM3 Protects Against Abdominal Aortic Aneurysm by Suppressing Ferroptosis in Vascular Smooth Muscle Cells. Circulation 2023. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Jiao, X.; Yang, Y.; Lv, Q.; Du, Z.; Li, L.; Hu, C.; Du, Y.; Zhang, J.; Li, F.; et al. ANGPTL8 Deletion Attenuates Abdominal Aortic Aneurysm Formation in ApoE−/− Mice. Clin. Sci. 2023, 137, 979–993. [Google Scholar] [CrossRef] [PubMed]
- Niu, X.; Wang, B. A Network Medical Framework Based on Inflammatory Genes to Identify Drug Candidates for Abdominal Aortic Aneurysms. Curr. Mol. Pharmacol. 2023, 17, e170523216998. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
| Factor | Molecular Interactions | Pathophysiological Significance | References |
|---|---|---|---|
| Chymase | Transform pro-MMP-9 to active form; induce TNF and ANG-II expression | Chymase inhibitors suppress the accumulation of vascular tumor cells, reduce mean blood pressure and total renal blood flow on murine model | [72,73] |
| Tryptase | Activate PAR-2 and iPLA2; induce LTC4, PGD2 expression | Tryptase is upregulated in several CVDs, inducing the fibrosis, inflammation, calcification and permeability of the vessels | [74,75] |
| IL-6 | Stimulate GM-CSF and STAT3 expression | IL-6 signaling accumulates proteolytically-active macrophages, inducing vascular inflammation | [40,76] |
| GM-CSF | Upregulate MCP-1 expression | A deficiency of GM-CSF inhibits AAA and leads to a decrease in inflammatory monocytes, as well as the activation of M1 macrophage | [22] |
| TGF-β | Upregulate ERK expression | Accelerates AAA progression by widening the artery wall, though VSMC proliferation | [77] |
| MCP-1 | Upregulate CCR2, Lyz1, Cd52, Cd180 and other inflammatory/immune cell markers | MCP-1 pathways contribute to T cell receptor signaling, chemokine signaling, phagocytosis and inflammatory-related pathways | [78,79] |
| ROS | Upregulate MKL1, E2F1, FOXM1, ICAM-1 and plenty of cardiovascular pathophysiological factors | ROS neutralization reduces inflammatory cell infiltration, VSMC migration, phenotype switching and apoptosis | [80,81,82] |
| NGAL | Upregulate Cas-3 and protect MMP-9 against degradation | NGAL blood concentration indicates vessel wall deterioration. Its level is even higher in patients with ruptured AAA | [83,84] |
| IFN-γ | Upregulate CXCL10, MMP-2 and MMP-9 | Plays as an inflammatory factor, enhances endothelium adhesion and macrophage recruitment | [76,85] |
| HCY | Activate Ang-II receptor, upregulate ATX (or ENPP2) and IL-αL, -β1, -β2 and -β7 | Induces blood clotting, the degradation of elastin in the elastic membrane and the processes of fibrosis and calcification | [48,86] |
| MMPs | Can affect the expression of other types of MMPs. Some of them are engaged in increasing TNF and TGF-β levels. | MMPs are directly responsible for the degradation of ECM components and thus the pathophysiology of AAA | [87,88] |
| LOX | Expression of LOX is related with ANG-II induced AAA; its activity is inhibited by β-APN. | The role of LOX is to enhance the cross-linking of collagen and elastin, contributing to the integrity and stabilization of a healthy vascular wall. | [89,90] |
| OPG | OPG deficiency enhances the trail-induced Jnk-MMP9 pathway, controls RANK/RANKL interactions | A decreased level of OPG expression constitutes an AAA marker. | [75,91,92] |
| OPN | Inhibit the expression of TR1 lymphocytes, chemoattract the inflammatory cells | It is strictly related to VSMC phenotype switch, supporting calcification, hyperchondroplasia and aneurysm development | [86,93,94] |
| Cathepsins | Its expression is regulated by TNFα and INF-γ; regulates the distribution of ERL chemokines | Regulates the chemotaxis of macrophages, influencing the development of AAA and promoting angiogenesis | [95,96] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Domagała, D.; Data, K.; Szyller, H.; Farzaneh, M.; Mozdziak, P.; Woźniak, S.; Zabel, M.; Dzięgiel, P.; Kempisty, B. Cellular, Molecular and Clinical Aspects of Aortic Aneurysm—Vascular Physiology and Pathophysiology. Cells 2024, 13, 274. https://doi.org/10.3390/cells13030274
Domagała D, Data K, Szyller H, Farzaneh M, Mozdziak P, Woźniak S, Zabel M, Dzięgiel P, Kempisty B. Cellular, Molecular and Clinical Aspects of Aortic Aneurysm—Vascular Physiology and Pathophysiology. Cells. 2024; 13(3):274. https://doi.org/10.3390/cells13030274
Chicago/Turabian StyleDomagała, Dominika, Krzysztof Data, Hubert Szyller, Maryam Farzaneh, Paul Mozdziak, Sławomir Woźniak, Maciej Zabel, Piotr Dzięgiel, and Bartosz Kempisty. 2024. "Cellular, Molecular and Clinical Aspects of Aortic Aneurysm—Vascular Physiology and Pathophysiology" Cells 13, no. 3: 274. https://doi.org/10.3390/cells13030274