DNA Helicase-SSB Interactions Critical to the Regression and Restart of Stalled DNA Replication Forks in Escherichia coli


Abstract
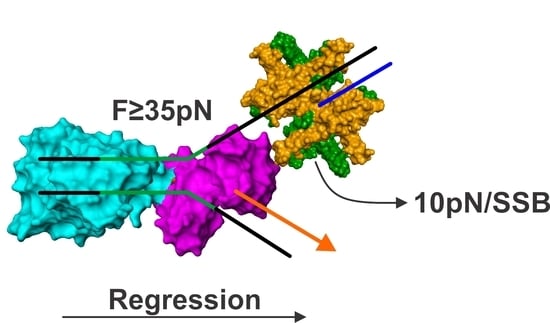
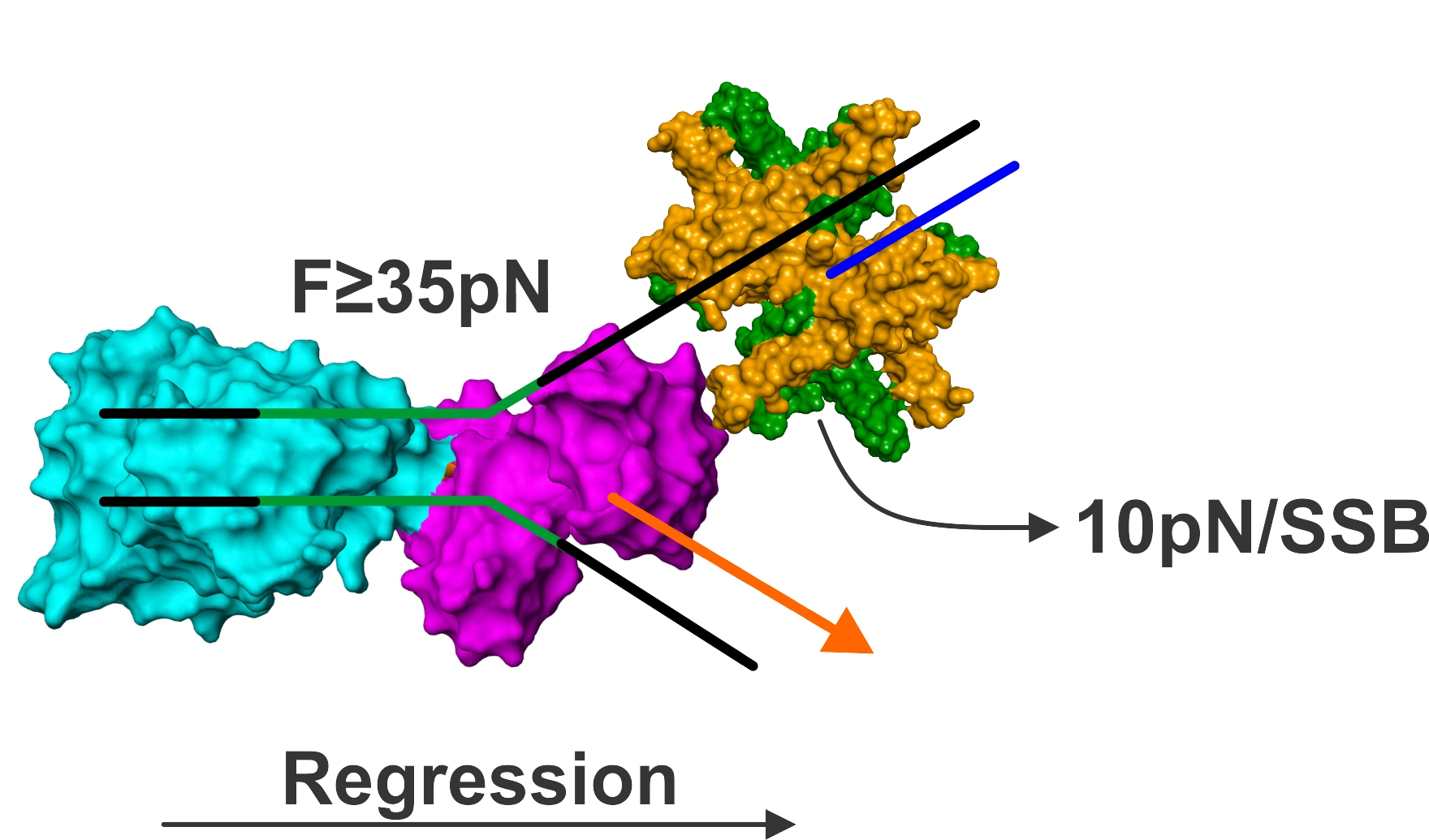{kind=link}
{kind=link}
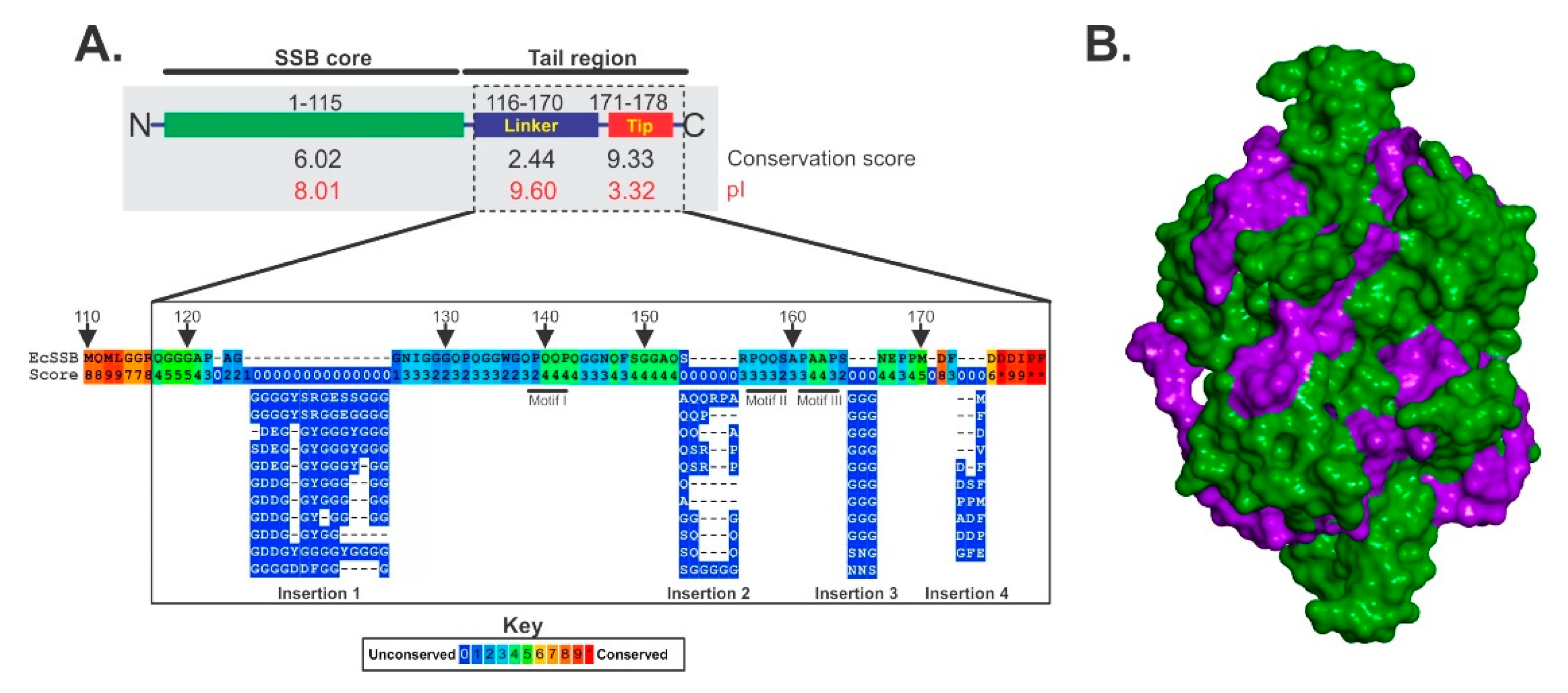{kind=link}
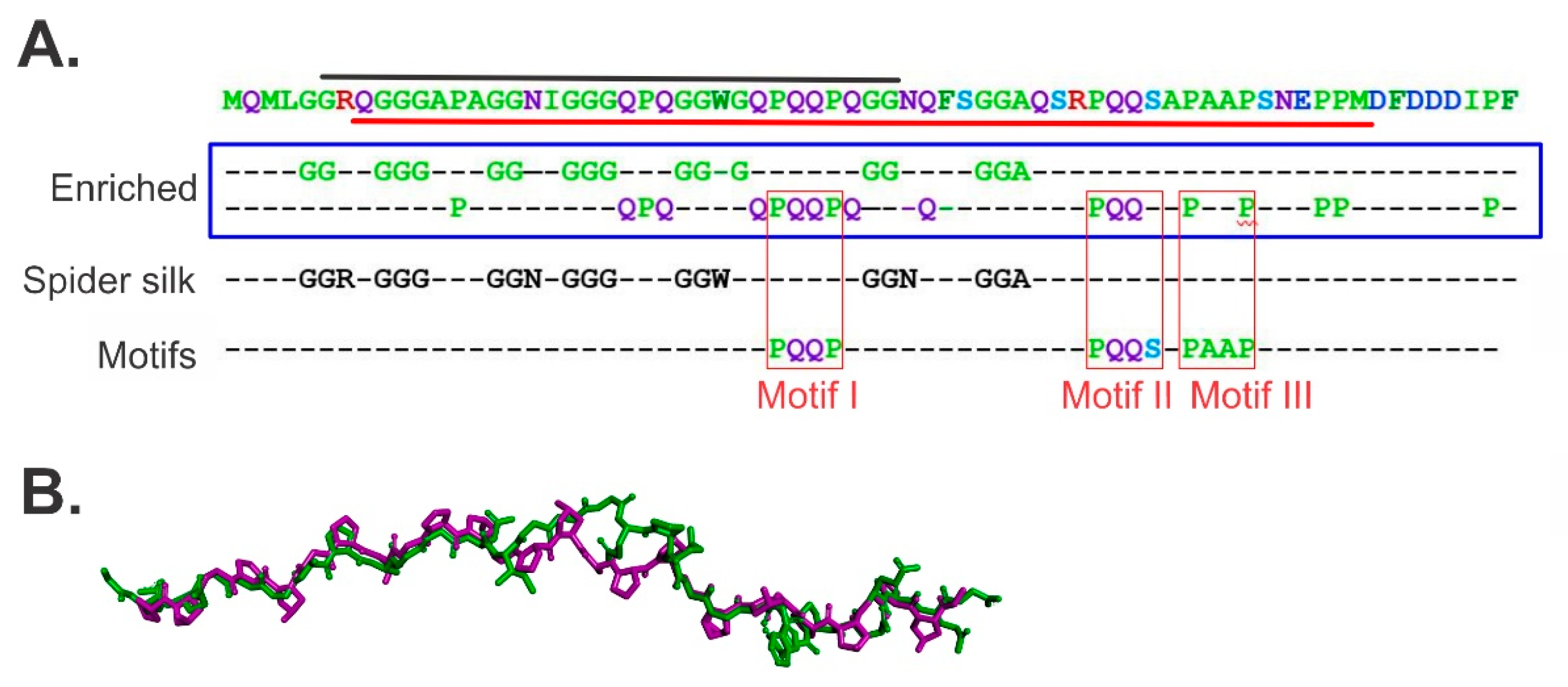{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Fork Regression Defined
3. The Protein Players
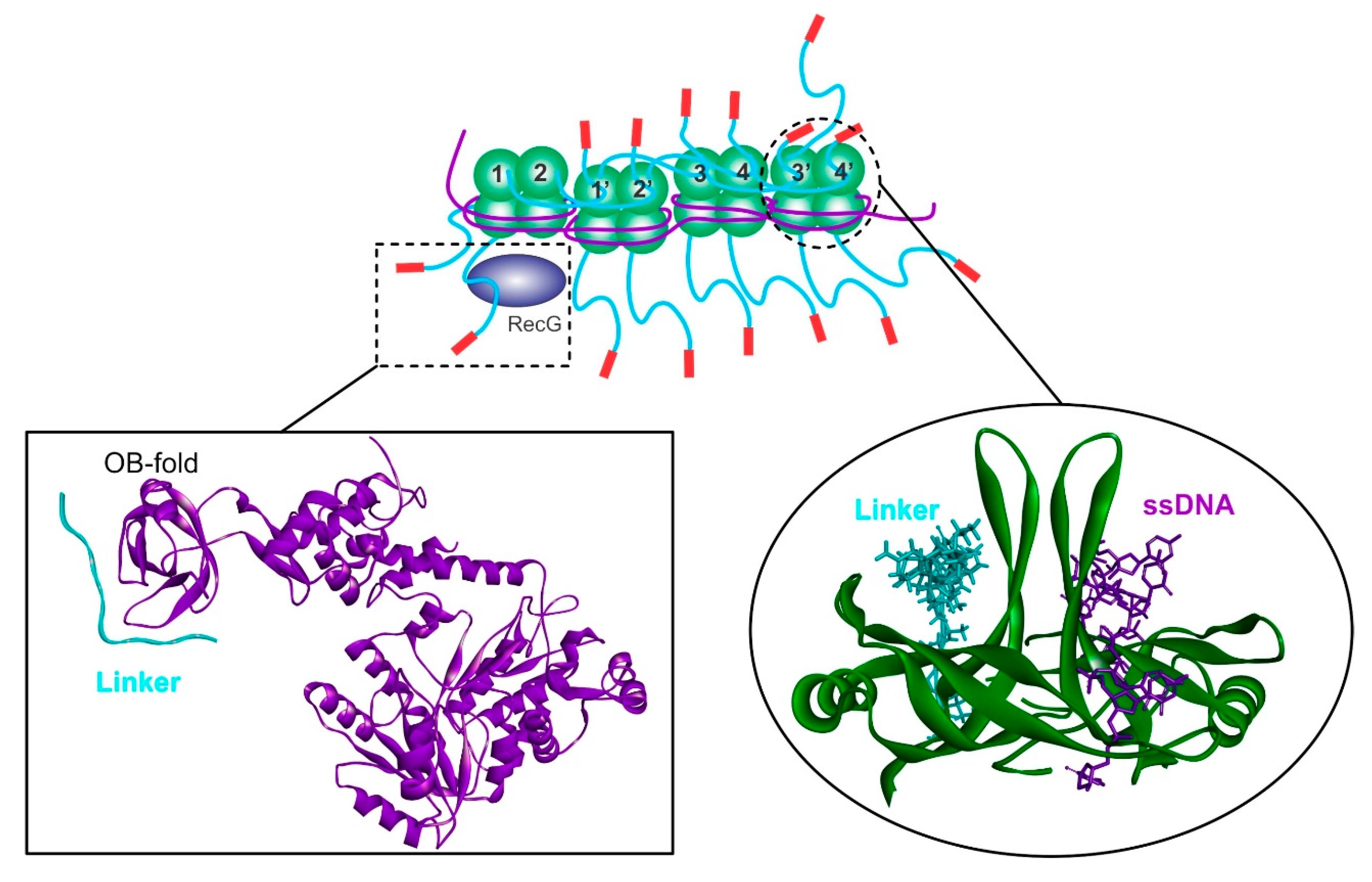
3.1. SSB—The Mediator of DNA Transactions at Forks
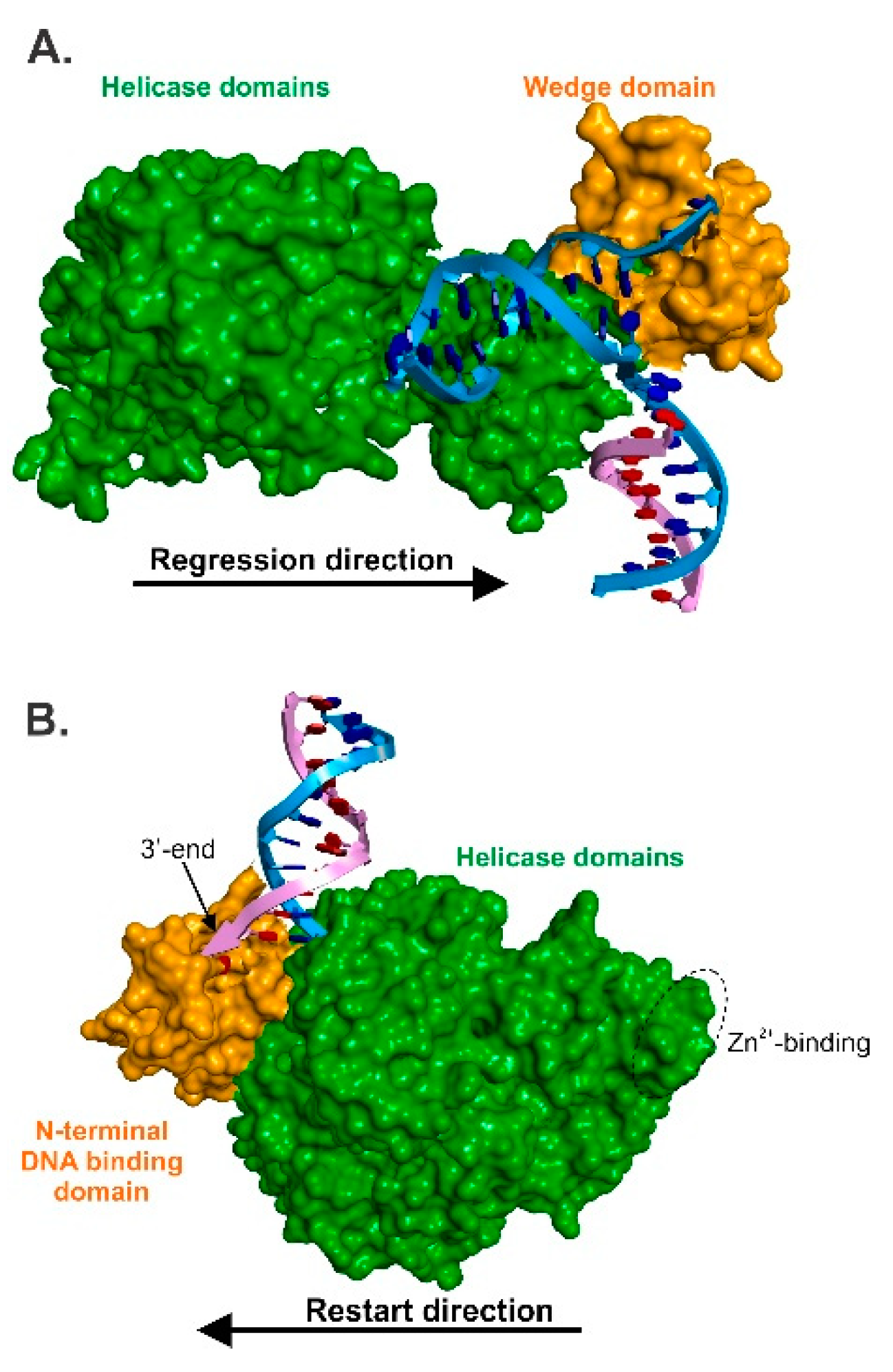
3.2. RecG—The Regression Beast
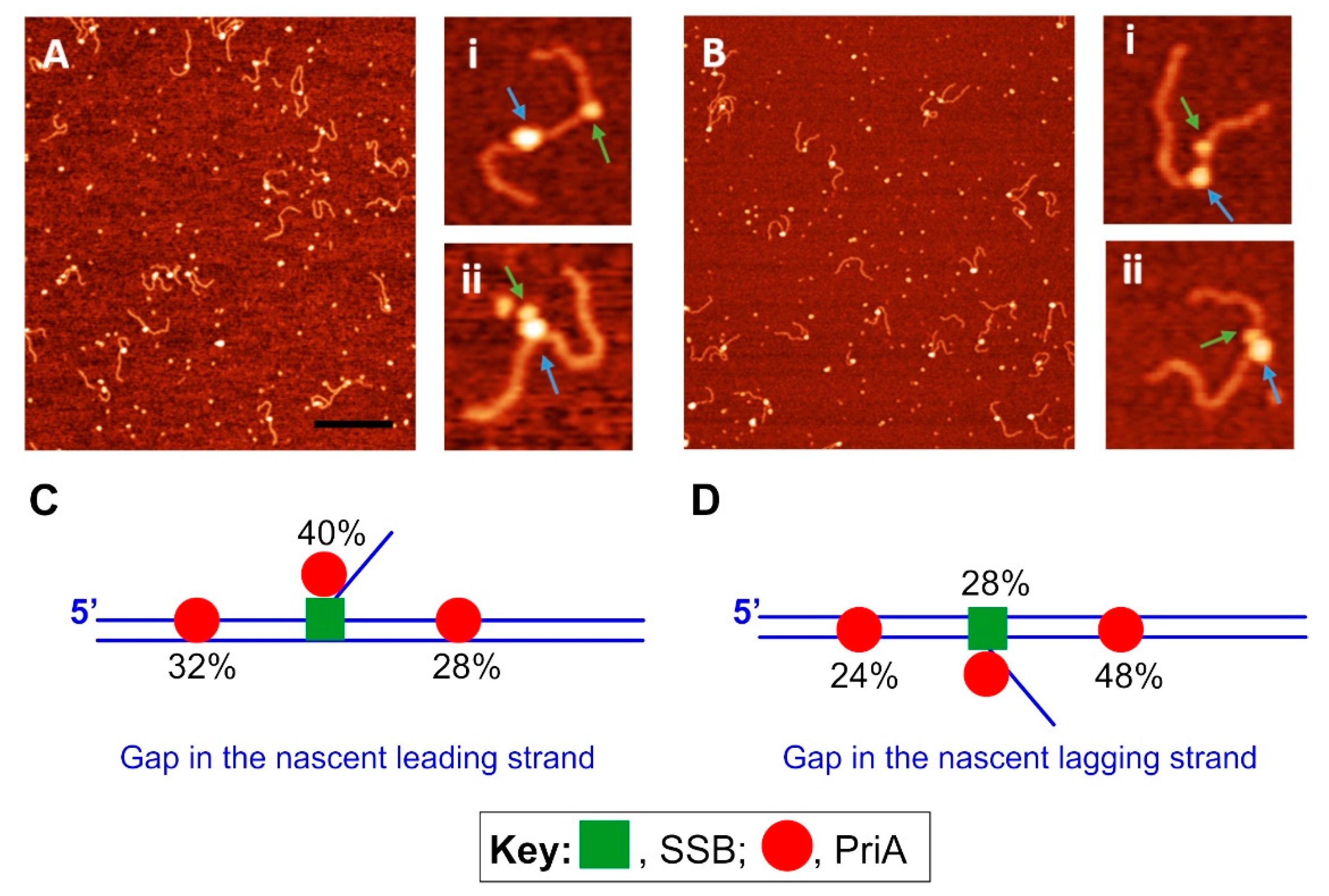
3.3. PriA—The Restart Specialist
4. SSB-DNA Helicase Interactions during Loading
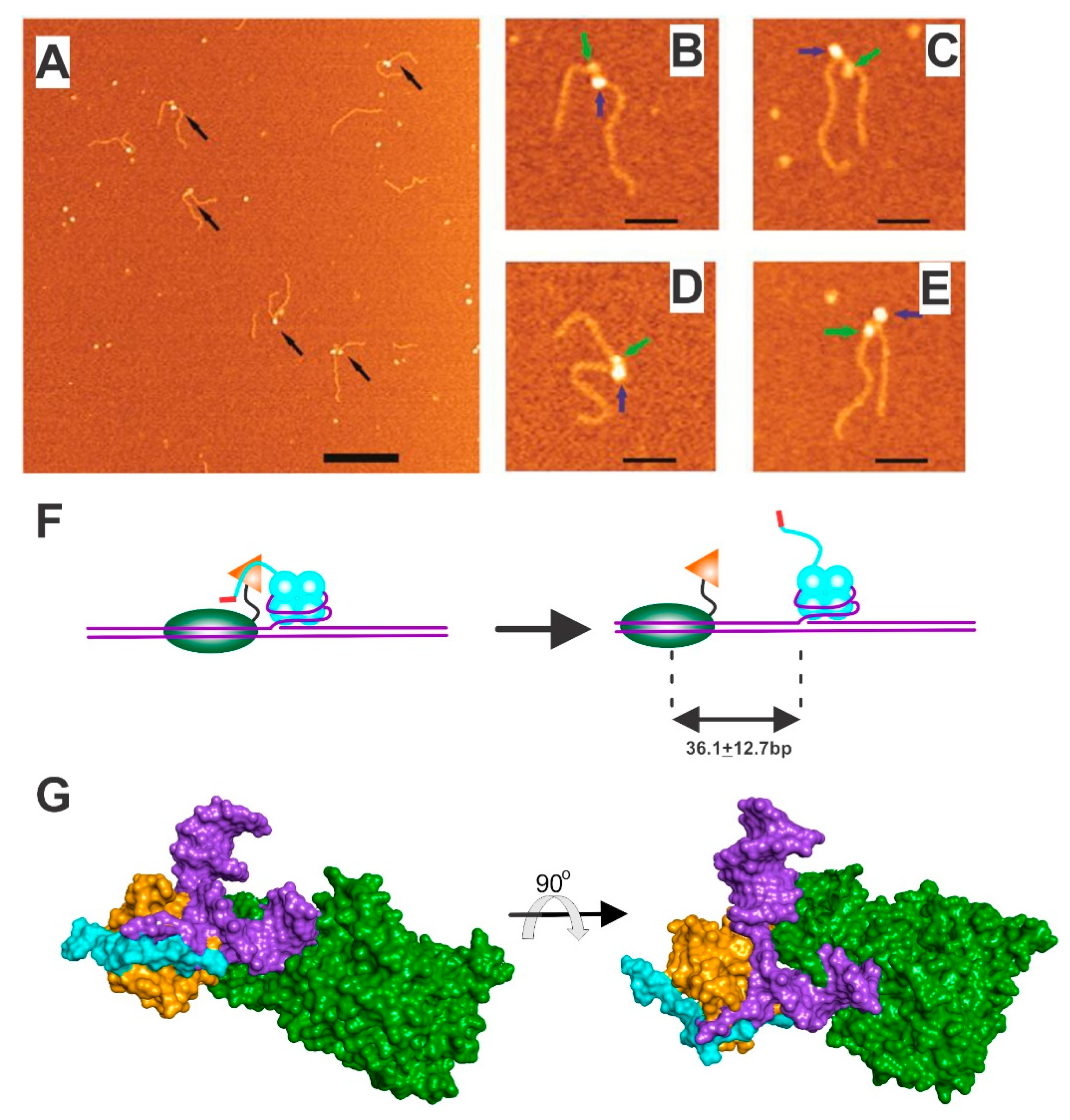
4.1. SSB-RecG
4.2. SSB-PriA
5. The Mechanics of Fork Regression by RecG
6. Summary
Funding
Conflicts of Interest
References
- Kogoma, T. Stable DNA replication: Interplay between DNA replication, homologous recombination, and transcription. Microbiol. Mol. Boil. Rev. 1997, 61, 212–238. [Google Scholar] [CrossRef]
- Kuzminov, A. Recombinational Repair of DNA Damage in Escherichia coli and Bacteriophage λ. Microbiol. Mol. Boil. Rev. 1999, 63, 751–813. [Google Scholar] [CrossRef]
- Kowalczykowski, S.C. Initiation of genetic recombination and recombination-dependent replication. Trends Biochem. Sci. 2000, 25, 156–165. [Google Scholar] [CrossRef]
- Cox, M. Recombinational DNA Repair of Damaged Replication Forks inEscherichia coli: Questions. Annu. Rev. Genet. 2001, 35, 53–82. [Google Scholar] [CrossRef]
- Kreuzer, K.N. Interplay between Dna Replication and Recombination in Prokaryotes. Annu. Rev. Microbiol. 2005, 59, 43–67. [Google Scholar] [CrossRef]
- McGlynn, P.; Lloyd, R.G. Recombinational repair and restart of damaged replication forks. Nat. Rev. Mol. Cell Boil. 2002, 3, 859–870. [Google Scholar] [CrossRef]
- Cox, M.M.; Goodman, M.F.; Kreuzer, K.N.; Sherratt, D.J.; Sandler, S.J.; Marians, K.J. The importance of repairing stalled replication forks. Nature 2000, 404, 37–41. [Google Scholar] [CrossRef]
- McGlynn, P.; Lloyd, R.G. Replicating past lesions in DNA. Mol. Cell 2002, 10, 700–701. [Google Scholar] [CrossRef]
- Marians, K.J. Mechanisms of replication fork restart inEscherichia coli. Philos. Trans. R. Soc. B: Boil. Sci. 2004, 359, 71–77. [Google Scholar] [CrossRef][Green Version]
- Mirkin, E.V.; Mirkin, S.M. Replication Fork Stalling at Natural Impediments. Microbiol. Mol. Boil. Rev. 2007, 71, 13–35. [Google Scholar] [CrossRef]
- Voineagu, I.; Narayanan, V.; Lobachev, K.S.; Mirkin, S.M. Replication stalling at unstable inverted repeats: Interplay between DNA hairpins and fork stabilizing proteins. Proc. Natl. Acad. Sci. USA 2008, 105, 9936–9941. [Google Scholar] [CrossRef] [PubMed]
- Gupta, M.K.; Guy, C.P.; Yeeles, J.T.P.; Atkinson, J.; Bell, H.; Lloyd, R.G.; Marians, K.J.; McGlynn, P. Protein–DNA complexes are the primary sources of replication fork pausing in Escherichia coli. Proc. Natl. Acad. Sci. USA 2013, 110, 7252–7257. [Google Scholar] [CrossRef]
- Michel, B.; Grompone, G.; Florès, M.-J.; Bidnenko, V. Multiple pathways process stalled replication forks. Proc. Natl. Acad. Sci. USA 2004, 101, 12783–12788. [Google Scholar] [CrossRef]
- McGlynn, P.; Lloyd, R.G. Genome stability and the processing of damaged replication forks by RecG. Trends Genet. 2002, 18, 413–419. [Google Scholar] [CrossRef]
- Marians, K.J. PriA-directed replication fork restart in Escherichia coli. Trends Biochem. Sci. 2000, 25, 185–189. [Google Scholar] [CrossRef]
- Marians, K.J. Replication and recombination intersect. Curr. Opin. Genet. Dev. 2000, 10, 151–156. [Google Scholar] [CrossRef]
- Courcelle, J.; Hanawalt, P.C. RecA-Dependent Recovery of Arrested DNA Replication Forks. Annu. Rev. Genet. 2003, 37, 611–646. [Google Scholar] [CrossRef]
- Henderson, M.L.; Kreuzer, K.N. Functions that Protect Escherichia coli from Tightly Bound DNA-Protein Complexes Created by Mutant EcoRII Methyltransferase. PLoS ONE 2015, 10, e0128092. [Google Scholar] [CrossRef]
- Heller, R.C.; Marians, K.J. Replication fork reactivation downstream of a blocked nascent leading strand. Nature 2006, 439, 557–562. [Google Scholar] [CrossRef]
- Heller, R.C.; Marians, K.J. Replisome assembly and the direct restart of stalled replication forks. Nat. Rev. Mol. Cell Boil. 2006, 7, 932–943. [Google Scholar] [CrossRef]
- Lusetti, S.L.; Cox, M. The Bacterial RecA Protein and the Recombinational DNA Repair of Stalled Replication Forks. Annu. Rev. Biochem. 2002, 71, 71–100. [Google Scholar] [CrossRef] [PubMed]
- Heller, R.C.; Marians, K.J. Unwinding of the Nascent Lagging Strand by Rep and PriA Enables the Direct Restart of Stalled Replication Forks. J. Boil. Chem. 2005, 280, 34143–34151. [Google Scholar] [CrossRef] [PubMed]
- Kuzminov, A. Collapse and repair of replication forks in Escherichia coli. Mol. Microbiol. 1995, 16, 373–384. [Google Scholar] [CrossRef]
- Manosas, M.; Perumal, S.K.; Bianco, P.R.; Ritort, F.; Benkovic, S.J.; Croquette, V. RecG and UvsW catalyse robust DNA rewinding critical for stalled DNA replication fork rescue. Nat. Commun. 2013, 4, 2368. [Google Scholar] [CrossRef] [PubMed]
- Bianco, P.R. Stalled replication fork rescue requires a novel DNA helicase. Methods 2016, 108, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Masai, H.; Asai, T.; Kubota, Y.; Arai, K.; Kogoma, T. Escherichia coli PriA protein is essential for inducible and constitutive stable DNA replication. EMBO J. 1994, 13, 5338–5345. [Google Scholar] [CrossRef] [PubMed]
- Gabbai, C.B.; Marians, K.J. Recruitment to stalled replication forks of the PriA DNA helicase and replisome-loading activities is essential for survival. DNA Repair 2010, 9, 202–209. [Google Scholar] [CrossRef]
- Sandler, S.J.; Marians, K.J. Role of PriA in Replication Fork Reactivation inEscherichia coli. J. Bacteriol. 2000, 182, 9–13. [Google Scholar] [CrossRef]
- Lewis, J.; Spenkelink, L.M.; Jergic, S.; Wood, E.A.; Monachino, E.; Horan, N.P.; Duderstadt, K.E.; Cox, M.M.; Robinson, A.; Dixon, N.E.; et al. Single-molecule visualization of fast polymerase turnover in the bacterial replisome. eLife 2017, 6, 6. [Google Scholar] [CrossRef]
- Jeiranian, H.A.; Schalow, B.J.; Courcelle, C.T.; Courcelle, J. Fate of the replisome following arrest by UV-induced DNA damage in Escherichia coli. Proc. Natl. Acad. Sci. USA 2013, 110, 11421–11426. [Google Scholar] [CrossRef]
- Yeeles, J.T.P.; Marians, K.J. The Escherichia coli Replisome Is Inherently DNA Damage Tolerant. Science 2011, 334, 235–238. [Google Scholar] [CrossRef]
- Postow, L.; Crisona, N.J.; Peter, B.J.; Hardy, C.D.; Cozzarelli, N.R. Topological challenges to DNA replication: Conformations at the fork. Proc. Natl. Acad. Sci. USA 2001, 98, 8219–8226. [Google Scholar] [CrossRef] [PubMed]
- Postow, L.; Ullsperger, C.; Keller, R.W.; Bustamante, C.; Vologodskii, A.V.; Cozzarelli, N.R. Positive Torsional Strain Causes the Formation of a Four-way Junction at Replication Forks. J. Boil. Chem. 2000, 276, 2790–2796. [Google Scholar] [CrossRef] [PubMed]
- McGlynn, P.; Lloyd, R.G. Modulation of RNA Polymerase by (p)ppGpp Reveals a RecG-Dependent Mechanism for Replication Fork Progression. Cell 2000, 101, 35–45. [Google Scholar] [CrossRef]
- Robu, M.E.; Inman, R.B.; Cox, M.M. Situational repair of replication forks: Roles of RecG and RecA proteins. J. Biol. Chem. 2004, 279, 10973–10981. [Google Scholar] [CrossRef]
- Robu, M.E.; Inman, R.B.; Cox, M. RecA protein promotes the regression of stalled replication forks in vitro. Proc. Natl. Acad. Sci. USA 2001, 98, 8211–8218. [Google Scholar] [CrossRef]
- Seigneur, M.; Bidnenko, V.; Ehrlich, S.; Michel, B. RuvAB Acts at Arrested Replication Forks. Cell 1998, 95, 419–430. [Google Scholar] [CrossRef]
- Gupta, S.; Yeeles, J.T.P.; Marians, K.J. Regression of Replication Forks Stalled by Leading-strand Template Damage. J. Boil. Chem. 2014, 289, 28388–28398. [Google Scholar] [CrossRef]
- Gupta, S.; Yeeles, J.T.P.; Marians, K.J. Regression of Replication Forks Stalled by Leading-strand Template Damage I-Both Recg and RuvAB Catalyze Regression, but RuvC Cleaves the Holliday Junctions FORMED BY RecG Preferentially. J. Biol. Chem. 2014, 289, 28376–28387. [Google Scholar] [CrossRef]
- Rosenberg, M.; Echols, H. Differential recognition of ultraviolet lesions by RecA protein. Possible mechanism for preferential targeting of SOS mutagenesis to (6-4) dipyrimidine sites. J. Boil. Chem. 1990, 265, 20641–20645. [Google Scholar]
- Wahab, S.A.; Choi, M.; Bianco, P.R. Characterization of the ATPase Activity of RecG and RuvAB Proteins on Model Fork Structures Reveals Insight into Stalled DNA Replication Fork Repair*. J. Boil. Chem. 2013, 288, 26397–26409. [Google Scholar] [CrossRef] [PubMed]
- Buss, J.A.; Kimura, Y.; Bianco, P.R. RecG interacts directly with SSB: Implications for stalled replication fork regression. Nucleic Acids Res. 2008, 36, 7029–7042. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Tan, H.Y.; Bianco, P.R.; Lyubchenko, Y.L. Remodeling of RecG Helicase at the DNA Replication Fork by SSB Protein. Sci. Rep. 2015, 5, 9625. [Google Scholar] [CrossRef] [PubMed]
- Bianco, P.R.; Lyubchenko, Y.L. SSB and the RecG DNA helicase: An intimate association to rescue a stalled replication fork. Protein Sci. 2017, 26, 638–649. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Marians, K.J. Purification and Characterization of DnaC810, a Primosomal Protein Capable of Bypassing PriA Function. J. Boil. Chem. 2000, 275, 8196–8205. [Google Scholar] [CrossRef]
- Yu, C.; Tan, H.Y.; Choi, M.; Stanenas, A.J.; Byrd, A.; Raney, K.D.; Cohan, C.S.; Bianco, P.R. SSB binds to the RecG and PriA helicases in vivo in the absence of DNA. Genes Cells 2016, 21, 163–184. [Google Scholar] [CrossRef]
- Cadman, C.J.; McGlynn, P. PriA helicase and SSB interact physically and functionally. Nucleic Acids Res. 2004, 32, 6378–6387. [Google Scholar] [CrossRef]
- Wang, Y.; Sun, Z.; Bianco, P.R.; Lyubchenko, Y.L. AFM characterization of the interaction of PriA helicase with stalled DNA replication forks. J. Biol. Chem. 2020, in press. [Google Scholar] [CrossRef]
- West, S. Processing of recombination intermediates by the ruvabc proteins. Annu. Rev. Genet. 1997, 31, 213–244. [Google Scholar] [CrossRef]
- Shereda, R.D.; Kozlov, A.G.; Lohman, T.M.; Cox, M.M.; Keck, J.L. SSB as an organizer/mobilizer of genome maintenance complexes. Crit. Rev. Biochem. Mol. Boil. 2008, 43, 289–318. [Google Scholar] [CrossRef]
- Chase, J.W.; Williams, K.R. Single-stranded DNA binding proteins required for DNA replication. Annu. Rev. Biochem. 1986, 55, 103–136. [Google Scholar] [CrossRef] [PubMed]
- Meyer, R.R.; Laine, P.S. The single-stranded DNA-binding protein of Escherichia coli. Microbiol. Rev. 1990, 54, 342–380. [Google Scholar] [CrossRef] [PubMed]
- Kowalczykowski, S.C.; A Dixon, D.; Eggleston, A.; Lauder, S.D.; Rehrauer, W.M. Biochemistry of homologous recombination in Escherichia coli. Microbiol. Rev. 1994, 58, 401–465. [Google Scholar] [CrossRef] [PubMed]
- Lohman, T.M.; E Ferrari, M. Escherichia coli Single-Stranded DNA-Binding Protein: Multiple DNA-Binding Modes and Cooperativities. Annu. Rev. Biochem. 1994, 63, 527–570. [Google Scholar] [CrossRef] [PubMed]
- Costes, A.; Lecointe, F.; McGovern, S.; Quevillon-Cheruel, S.; Polard, P. The C-terminal Domain of the Bacterial SSB Protein Acts as a DNA Maintenance Hub at Active Chromosome Replication Forks. PLoS Genet. 2010, 6, e1001238. [Google Scholar] [CrossRef]
- Sancar, A.; Williams, K.; Chase, J.W.; Rupp, W.D. Sequences of the ssb gene and protein. Proc. Natl. Acad. Sci. USA 1981, 78, 4274–4278. [Google Scholar] [CrossRef]
- Curth, U.; Genschel, J.; Urbanke, C.; Greipel, J. In Vitro and in Vivo Function of the C-Terminus of Escherichia coli Single-Stranded DNA Binding Protein. Nucleic Acids Res. 1996, 24, 2706–2711. [Google Scholar] [CrossRef]
- Raghunathan, S.; Kozlov, A.G.; Lohman, T.M.; Waksman, G. Structure of the DNA binding domain of E. coli SSB bound to ssDNA. Nat. Genet. 2000, 7, 648–652. [Google Scholar] [CrossRef]
- Kuznetsov, S.V.; Kozlov, A.G.; Lohman, T.M.; Ansari, A. Microsecond Dynamics of Protein–DNA Interactions: Direct Observation of the Wrapping/Unwrapping Kinetics of Single-stranded DNA around the E. coli SSB Tetramer. J. Mol. Boil. 2006, 359, 55–65. [Google Scholar] [CrossRef]
- Ding, W.; Tan, H.Y.; Zhang, J.X.; Wilczek, L.A.; Hsieh, K.R.; Mulkin, J.A.; Bianco, P.R. The mechanism of SSB-RecG binding: Implications for SSB interactome function. Protein Sci. 2020, in press. [Google Scholar] [CrossRef]
- Simossis, V.A.; Heringa, J. PRALINE: A multiple sequence alignment toolbox that integrates homology-extended and secondary structure information. Nucleic Acids Res. 2005, 33, W289–W294. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, M.R.; Gasteiger, E.; Bairoch, A.; Sanchez, J.-C.; Williams, K.L.; Appel, R.D.; Hochstrasser, D.F. Protein identification and analysis tools in the ExPASy server. Ther. Antib. 1999, 112, 531–552. [Google Scholar] [CrossRef]
- Kozlov, A.G.; Weiland, E.; Mittal, A.; Waldman, V.; Pappu, R.V.; Timothy, L.M. The Intrinsically Disordered C-terminal Tails of E. coli Single-Stranded DNA Binding Protein Regulate Cooperative Binding to Single-Stranded DNA. Biophys. J. 2015, 108, 389a. [Google Scholar] [CrossRef]
- Bianco, P.R. The tale of SSB. Prog. Biophys. Mol. Boil. 2016, 127, 111–118. [Google Scholar] [CrossRef]
- Kozlov, A.G.; Jezewska, M.J.; Bujalowski, W.; Lohman, T.M. Binding specificity of Escherichia coli single-stranded DNA binding protein for the chi subunit of DNA pol III holoenzyme and PriA helicase. Biochemistry 2010, 49, 3555–3566. [Google Scholar] [CrossRef]
- Sandigursky, M.; Mendez, F.; Bases, R.E.; Matsumoto, T.; Franklin, W.A. Protein-Protein Interactions between the Escherichia coli Single-Stranded DNA-Binding Protein and Exonuclease I. Radiat. Res. 1996, 145, 619. [Google Scholar] [CrossRef]
- Bianco, P.R.; Pottinger, S.; Tan, H.Y.; Nguyenduc, T.; Rex, K.; Varshney, U. The IDL of E. coli SSB links ssDNA and protein binding by mediating protein–protein interactions. Protein Sci. 2017, 26, 227–241. [Google Scholar] [CrossRef]
- Huang, Y.-H.; Huang, C.-Y. The glycine-rich flexible region in SSB is crucial for PriA stimulation. RSC Adv. 2018, 8, 35280–35288. [Google Scholar] [CrossRef]
- Nigam, R.; Mohan, M.; Shivange, G.; Dewangan, P.K.; Anindya, R. Escherichia coli AlkB interacts with single-stranded DNA binding protein SSB by an intrinsically disordered region of SSB. Mol. Boil. Rep. 2018, 45, 865–870. [Google Scholar] [CrossRef]
- Su, X.-C.; Wang, Y.; Yagi, H.; Shishmarev, D.; Mason, C.E.; Smith, P.J.; Vandevenne, M.; Dixon, N.E.; Otting, G. Bound or Free: Interaction of the C-terminal Domain of Escherichia coli Single-Stranded DNA-Binding Protein (SSB) with the Tetrameric Core of SSB. Biochemistry 2014, 53, 1925–1934. [Google Scholar] [CrossRef] [PubMed]
- Williams, K.R.; Spicer, E.K.; Lopresti, M.B.; A Guggenheimer, R.; Chase, J.W. Limited proteolysis studies on the Escherichia coli single-stranded DNA binding protein. Evidence for a functionally homologous domain in both the Escherichia coli and T4 DNA binding proteins. J. Boil. Chem. 1983, 258, 3346–3355. [Google Scholar]
- Brown, A.M.; Zondlo, N.J. A Propensity Scale for Type II Polyproline Helices (PPII): Aromatic Amino Acids in Proline-Rich Sequences Strongly Disfavor PPII Due to Proline–Aromatic Interactions. Biochemistry 2012, 51, 5041–5051. [Google Scholar] [CrossRef] [PubMed]
- Meirson, T.; Bomze, D.; Kahlon, L.; Gil-Henn, H.; Samson, A.O. A helical lock and key model of polyproline II conformation with SH3. Bioinformatics 2019, 36, 154–159. [Google Scholar] [CrossRef] [PubMed]
- Kurochkina, N.; Guha, U. SH3 domains: Modules of protein–protein interactions. Biophys. Rev. 2012, 5, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Saksela, K.; Permi, P. SH3 domain ligand binding: What’s the consensus and where’s the specificity? FEBS Lett. 2012, 586, 2609–2614. [Google Scholar] [CrossRef] [PubMed]
- Kay, B.K. SH3 domains come of age. FEBS Lett. 2012, 586, 2606–2608. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, V.; Kishan, K.V.R. Functional evolution of two subtly different (similar) folds. BMC Struct. Boil. 2001, 1, 5. [Google Scholar]
- Cheng, K.; Xu, H.; Chen, X.; Wang, L.; Tian, B.; Zhao, Y.; Hua, Y. Structural basis for DNA 5′-end resection by RecJ. eLife 2016, 5, 14294. [Google Scholar] [CrossRef]
- Wakamatsu, T.; Kitamura, Y.; Kotera, Y.; Nakagawa, N.; Kuramitsu, S.; Masui, R. Structure of RecJ Exonuclease Defines Its Specificity for Single-stranded DNA*. J. Boil. Chem. 2010, 285, 9762–9769. [Google Scholar] [CrossRef]
- Singleton, M.; Scaife, S.; Wigley, D.B. Structural Analysis of DNA Replication Fork Reversal by RecG. Cell 2001, 107, 79–89. [Google Scholar] [CrossRef]
- Ryzhikov, M.; Koroleva, O.; Postnov, D.; Tran, A.; Korolev, S. Mechanism of RecO recruitment to DNA by single-stranded DNA binding protein. Nucleic Acids Res. 2011, 39, 6305–6314. [Google Scholar] [CrossRef] [PubMed]
- Windgassen, T.A.; Leroux, M.; Sandler, S.J.; Keck, J.L. Function of a strand-separation pin element in the PriA DNA replication restart helicase. J. Boil. Chem. 2018, 294, 2801–2814. [Google Scholar] [CrossRef] [PubMed]
- Bharti, S.K.; Rex, K.; Sreedhar, P.; Krishnan, N.; Varshney, U. Chimeras of Escherichia coli and Mycobacterium tuberculosis Single-Stranded DNA Binding Proteins: Characterization and Function in Escherichia coli. PLoS ONE 2011, 6, 27216. [Google Scholar] [CrossRef] [PubMed]
- Storm, P.; Hoekstra, W.; De Haan, P.; Verhoef, C. Genetic recombination in Escherichia coliIV. Isolation and characterization of recombinaion-deficient mutants of Escherichia coli K12. Mutat. Res. Mol. Mech. Mutagen. 1971, 13, 9–17. [Google Scholar] [CrossRef][Green Version]
- Benson, F.E.; Collier, S.; Lloyd, R.G. Evidence of abortive recombination in ruv mutants of Escherichia coli K12. Mol. Genet. Genom. 1991, 225, 266–272. [Google Scholar] [CrossRef]
- Lloyd, R.G. Conjugational recombination in resolvase-deficient ruvC mutants of Escherichia coli K-12 depends on recG. J. Bacteriol. 1991, 173, 5414–5418. [Google Scholar] [CrossRef]
- Kalman, M.; Murphy, H.; Cashel, M. The nucleotide sequence of recG, the distal spo operon gene in Escherichia coli K-12. Gene 1992, 110, 95–99. [Google Scholar] [CrossRef]
- Singleton, M.; Dillingham, M.; Wigley, D.B. Structure and Mechanism of Helicases and Nucleic Acid Translocases. Annu. Rev. Biochem. 2007, 76, 23–50. [Google Scholar] [CrossRef]
- Vincent, S.D.; Mahdi, A.A.; Lloyd, R.G. The RecG Branch Migration Protein of Escherichia coli Dissociates R-loops. J. Mol. Boil. 1996, 264, 713–721. [Google Scholar] [CrossRef]
- Sharples, G.J.; Ingleston, S.M.; Lloyd, R.G. Holliday Junction Processing in Bacteria: Insights from the Evolutionary Conservation of RuvABC, RecG, and RusA. J. Bacteriol. 1999, 181, 5543–5550. [Google Scholar] [CrossRef]
- Whitby, M.C.; Vincent, S.; Lloyd, R. Branch migration of Holliday junctions: Identification of RecG protein as a junction specific DNA helicase. EMBO J. 1994, 13, 5220–5228. [Google Scholar] [CrossRef]
- Peter, M.; Lloyd, R.G. RecG helicase activity at three- and four-strand DNA structures. Nucleic Acids Res. 1999, 27, 3049–3056. [Google Scholar] [CrossRef] [PubMed][Green Version]
- McGlynn, P. Characterisation of the catalytically active form of RecG helicase. Nucleic Acids Res. 2000, 28, 2324–2332. [Google Scholar] [CrossRef] [PubMed]
- Slocum, S.L.; Buss, J.A.; Kimura, Y.; Bianco, P.R. Characterization of the ATPase Activity of the Escherichia coli RecG Protein Reveals that the Preferred Cofactor is Negatively Supercoiled DNA. J. Mol. Boil. 2007, 367, 647–664. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Peter, B.J.; Ullsperger, C.; Hiasa, H.; Marians, K.J.; Cozzarelli, N.R. The Structure of Supercoiled Intermediates in DNA Replication. Cell 1998, 94, 819–827. [Google Scholar] [CrossRef]
- McGlynn, P.; Lloyd, R.G.; Marians, K.J. Formation of Holliday junctions by regression of nascent DNA in intermediates containing stalled replication forks: RecG stimulates regression even when the DNA is negatively supercoiled. Proc. Natl. Acad. Sci. USA 2001, 98, 8235–8240. [Google Scholar] [CrossRef]
- Lecointe, F.; Sérèna, C.; Velten, M.; Costes, A.; McGovern, S.; Meile, J.-C.; Errington, J.; Ehrlich, S.D.; Noirot, P.; Polard, P. Anticipating chromosomal replication fork arrest: SSB targets repair DNA helicases to active forks. EMBO J. 2007, 26, 4239–4251. [Google Scholar] [CrossRef]
- Wen, Q.; Mahdi, A.A.; Briggs, G.S.; Sharples, G.J.; Lloyd, R.G. Conservation of RecG activity from pathogens to hyperthermophiles. DNA Repair 2005, 4, 23–31. [Google Scholar] [CrossRef]
- Mahdi, A.A.; Briggs, G.S.; Sharples, G.J.; Wen, Q.; Lloyd, R.G. A model for dsDNA translocation revealed by a structural motif common to RecG and Mfd proteins. EMBO J. 2003, 22, 724–734. [Google Scholar] [CrossRef]
- Sun, Z.; Hashemi, M.; Warren, G.; Bianco, P.R.; Lyubchenko, Y.L. Dynamics of the Interaction of RecG Protein with Stalled Replication Forks. Biochemistry 2018, 57, 1967–1976. [Google Scholar] [CrossRef]
- Briggs, G.S.; Mahdi, A.A.; Wen, Q.; Lloyd, R.G. DNA Binding by the Substrate Specificity (Wedge) Domain of RecG Helicase Suggests a Role in Processivity. J. Boil. Chem. 2005, 280, 13921–13927. [Google Scholar] [CrossRef] [PubMed]
- Mahdi, A.A.; McGlynn, P.; Levett, S.D.; Lloyd, R.G. DNA Binding and Helicase Domains of the Escherichia coli Recombination Protein RecG. Nucleic Acids Res. 1997, 25, 3875–3880. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Marians, K.J. PriA: At the crossroads of DNA replication and recombination. In Progress in Nucleic Acid Research and Molecular Biology; Academic Press: Cambridge, MA, USA, 1999; Volume 63, pp. 39–67. [Google Scholar] [CrossRef]
- Masai, H. DnaA- and PriA-dependent primosomes Two distinct replication complexes for replication of Escherichia coli chromosome. Front. Biosci. 1996, 1, d48–d58. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.H.; Masai, H.; Allen, G.C.; Kornberg, A. The priA gene encoding the primosomal replicative n’ protein of Escherichia coli. Proc. Natl. Acad. Sci. USA 1990, 87, 4620–4624. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.M.; Nakai, H. PriA and phage T4 gp59: Factors that promote DNA replication on forked DNA substrates microreview. Mol. Microbiol. 2000, 36, 519–527. [Google Scholar] [CrossRef] [PubMed]
- Nurse, P.; Zavitz, K.H.; Marians, K.J. Inactivation of the Escherichia coli priA DNA replication protein induces the SOS response. J. Bacteriol. 1991, 173, 6686–6693. [Google Scholar] [CrossRef]
- Lee, E.H.; Kornberg, A. Replication deficiencies in priA mutants of Escherichia coli lacking the primosomal replication n’ protein. Proc. Natl. Acad. Sci. USA 1991, 88, 3029–3032. [Google Scholar] [CrossRef]
- Sandler, S.J.; Samra, H.S.; Clark, A.J. Differential Suppression of Pria2::Kan Phenotypes in Escherichia coli K-12 by Mutations in Pria, Lexa, and Dnac. Genetics 1996, 143, 5–13. [Google Scholar]
- Kogoma, T.; Cadwell, G.W.; Barnard, K.G.; Asai, T. The DNA replication priming protein, PriA, is required for homologous recombination and double-strand break repair. J. Bacteriol. 1996, 178, 1258–1264. [Google Scholar] [CrossRef]
- Heller, R.C.; Marians, K.J. Non-replicative helicases at the replication fork. DNA Repair 2007, 6, 945–952. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Masai, H. Stabilization of a Stalled Replication Fork by Concerted Actions of Two Helicases. J. Boil. Chem. 2005, 281, 3484–3493. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Mizukoshi, T.; Taniyama, C.; Kohda, D.; Arai, K.-I.; Masai, H. DNA Binding of PriA Protein Requires Cooperation of the N-terminal D-loop/Arrested-fork Binding and C-terminal Helicase Domains. J. Boil. Chem. 2002, 277, 38062–38071. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.-W.; North, S.H.; Nakai, H. Properties of the PriA Helicase Domain and Its Role in Binding PriA to Specific DNA Structures. J. Boil. Chem. 2004, 279, 38503–38512. [Google Scholar] [CrossRef]
- Windgassen, T.; Leroux, M.; Satyshur, K.; Sandler, S.J.; Keck, J.L. Structure-specific DNA replication-fork recognition directs helicase and replication restart activities of the PriA helicase. Proc. Natl. Acad. Sci. USA 2018, 115, E9075–E9084. [Google Scholar] [CrossRef]
- Zavitz, K.H.; Marians, K.J. Helicase-deficient cysteine to glycine substitution mutants of Escherichia coli replication protein PriA retain single-stranded DNA-dependent ATPase activity. Zn2+ stimulation of mutant PriA helicase and primosome assembly activities. J. Boil. Chem. 1993, 268, 4337–4346. [Google Scholar]
- Masai, H.; Deneke, J.; Furui, Y.; Tanaka, T.; Arai, K.I. Escherichia coli and Bacillus subtilis PriA proteins essential for recombination-dependent DNA replication: Involvement of ATPase/helicase activity of PriA for inducible stable DNA replication. Biochimie 1999, 81, 847–857. [Google Scholar] [CrossRef]
- Liu, J.; Nurse, P.; Marians, K.J. The ordered assembly of the phiX174-type primosome. III. PriB facilitates complex formation between PriA and DnaT. J. Boil. Chem. 1996, 271, 15656–15661. [Google Scholar] [CrossRef]
- Nurse, P.; Liu, J.; Marians, K.J. Two modes of PriA binding to DNA. J. Boil. Chem. 1999, 274, 25026–25032. [Google Scholar] [CrossRef]
- Jones, J.M.; Nakai, H. Duplex opening by primosome protein PriA for replisome assembly on a recombination intermediate. J. Mol. Boil. 1999, 289, 503–515. [Google Scholar] [CrossRef]
- McGlynn, P.; A Al-Deib, A.; Liu, J.; Marians, K.J.; Lloyd, R.G. The DNA replication protein PriA and the recombination protein RecG bind D-loops. J. Mol. Boil. 1997, 270, 212–221. [Google Scholar] [CrossRef] [PubMed]
- Mizukoshi, T.; Tanaka, T.; Arai, K.-I.; Kohda, D.; Masai, H. A Critical Role of the 3′ Terminus of Nascent DNA Chains in Recognition of Stalled Replication Forks. J. Boil. Chem. 2003, 278, 42234–42239. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.S.; Marians, K.J. Escherichia coli replication factor Y, a component of the primosome, can act as a DNA helicase. Proc. Natl. Acad. Sci. USA 1987, 84, 8345–8349. [Google Scholar] [CrossRef] [PubMed]
- Lasken, R.S.; Kornberg, A. The primosomal protein n’ of Escherichia coli is a DNA helicase. J. Boil. Chem. 1988, 263, 5512–5518. [Google Scholar]
- Tanaka, T.; Taniyama, C.; Arai, K.-I.; Masai, H. ATPase/helicase motif mutants of Escherichia coli PriA protein essential for recombination-dependent DNA replication. Genes Cells 2003, 8, 251–261. [Google Scholar] [CrossRef]
- Liu, J.; Marians, K.J. PriA-directed Assembly of a Primosome on D Loop DNA. J. Boil. Chem. 1999, 274, 25033–25041. [Google Scholar] [CrossRef]
- McGlynn, P.; Lloyd, R.G. Rescue of stalled replication forks by RecG: Simultaneous translocation on the leading and lagging strand templates supports an active DNA unwinding model of fork reversal and Holliday junction formation. Proc. Natl. Acad. Sci. USA 2001, 98, 8227–8234. [Google Scholar] [CrossRef]
- Gregg, A.V.; McGlynn, P.; Jaktaji, R.P.; Lloyd, R.G. Direct Rescue of Stalled DNA Replication Forks via the Combined Action of PriA and RecG Helicase Activities. Mol. Cell 2002, 9, 241–251. [Google Scholar] [CrossRef]
- Sasaki, K.; Ose, T.; Okamoto, N.; Maenaka, K.; Tanaka, T.; Masai, H.; Saito, M.; Shirai, T.; Kohda, D. Structural basis of the 3′-end recognition of a leading strand in stalled replication forks by PriA. EMBO J. 2007, 26, 2584–2593. [Google Scholar] [CrossRef]
- Tanaka, T.; Mizukoshi, T.; Sasaki, K.; Kohda, D.; Masai, H. Escherichia coli PriA Protein, Two Modes of DNA Binding and Activation of ATP Hydrolysis. J. Boil. Chem. 2007, 282, 19917–19927. [Google Scholar] [CrossRef]
- Bhattacharyya, B.; George, N.P.; Thurmes, T.M.; Zhou, R.; Jani, N.; Wessel, S.R.; Sandler, S.J.; Ha, T.; Keck, J.L. Structural mechanisms of PriA-mediated DNA replication restart. Proc. Natl. Acad. Sci. USA 2013, 111, 1373–1378. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.M.; Nakai, H. Escherichia coli PriA helicase: Fork binding orients the helicase to unwind the lagging strand side of arrested replication forks11Edited by M. Gottesman. J. Mol. Boil. 2001, 312, 935–947. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Senac, M.D.M.; Webb, M. Mechanism of Translocation and Kinetics of DNA Unwinding by the Helicase RecG†. Biochemistry 2005, 44, 16967–16976. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.; Kozlov, A.G.; Roy, R.; Zhang, J.; Korolev, S.; Lohman, T.M.; Ha, T. SSB Functions as a Sliding Platform that Migrates on DNA via Reptation. Cell 2011, 146, 485. [Google Scholar] [CrossRef]
- Tan, H.Y.; Wilczek, L.A.; Pottinger, S.; Manosas, M.; Yu, C.; Nguyenduc, T.; Bianco, P.R. The intrinsically disordered linker of E. coli SSB is critical for the release from single-stranded DNA. Protein Sci. 2017, 26, 700–717. [Google Scholar] [CrossRef] [PubMed]








© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bianco, P.R. DNA Helicase-SSB Interactions Critical to the Regression and Restart of Stalled DNA Replication Forks in Escherichia coli. Genes 2020, 11, 471. https://doi.org/10.3390/genes11050471
Bianco PR. DNA Helicase-SSB Interactions Critical to the Regression and Restart of Stalled DNA Replication Forks in Escherichia coli. Genes. 2020; 11(5):471. https://doi.org/10.3390/genes11050471
Chicago/Turabian StyleBianco, Piero R. 2020. "DNA Helicase-SSB Interactions Critical to the Regression and Restart of Stalled DNA Replication Forks in Escherichia coli" Genes 11, no. 5: 471. https://doi.org/10.3390/genes11050471
APA StyleBianco, P. R. (2020). DNA Helicase-SSB Interactions Critical to the Regression and Restart of Stalled DNA Replication Forks in Escherichia coli. Genes, 11(5), 471. https://doi.org/10.3390/genes11050471