Telomere Maintenance Mechanisms in Cancer

 ,
,  ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Telomere Maintenance Mechanisms
2.1. Telomere Maintenance Mechanisms in Non-Malignant Cells
2.2. Telomere Maintenance Mechanisms in Tumour Cells
2.2.1. TERT Promoter Mutations
2.2.2. TERT and TERC Amplifications
2.2.3. TERT Germline Genetic Variations
2.2.4. TERT Rearrangements
2.2.5. Epigenetic Mechanisms
2.2.6. Alternative Lengthening of Telomeres
2.2.7. Non-Defined Telomere Maintenance Mechanism
3. Final Remarks
Supplementary Materials
Acknowledgments
Conflicts of Interest
References
- Blackburn, E.H.; Epel, E.S.; Lin, J. Human telomere biology: A contributory and interactive factor in aging, disease risks, and protection. Science 2015, 350, 1193–1198. [Google Scholar] [CrossRef] [PubMed]
- Arnoult, N.; Karlseder, J. Complex interactions between the DNA-damage response and mammalian telomeres. Nat. Struct. Mol. Biol. 2015, 22, 859–866. [Google Scholar] [CrossRef] [PubMed]
- Heidenreich, B.; Kumar, R. TERT promoter mutations in telomere biology. Mutat. Res. 2017, 771, 15–31. [Google Scholar] [CrossRef] [PubMed]
- Jafri, M.A.; Ansari, S.A.; Alqahtani, M.H.; Shay, J.W. Roles of telomeres and telomerase in cancer, and advances in telomerase-targeted therapies. Genome Med. 2016, 8, 69. [Google Scholar] [CrossRef] [PubMed]
- Naderlinger, E.; Holzmann, K. Epigenetic Regulation of Telomere Maintenance for Therapeutic Interventions in Gliomas. Genes 2017, 8, 145. [Google Scholar] [CrossRef] [PubMed]
- Akincilar, S.C.; Unal, B.; Tergaonkar, V. Reactivation of telomerase in cancer. Cell. Mol. Life Sci. 2016, 73, 1659–1670. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, R.J.; Karlseder, J. Telomeres: Protecting chromosomes against genome instability. Nat. Rev. Mol. Cell Biol. 2010, 11, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Nandakumar, J.; Cech, T.R. Finding the end: Recruitment of telomerase to telomeres. Nat. Rev. Mol. Cell Biol. 2013, 14, 69–82. [Google Scholar] [CrossRef] [PubMed]
- Blasco, M.A. Telomeres and human disease: Ageing, cancer and beyond. Nat. Rev. Genet. 2005, 6, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Li, S.; Stohr, B.A. The role of telomere biology in cancer. Annu. Rev. Pathol. 2013, 8, 49–78. [Google Scholar] [CrossRef] [PubMed]
- Sfeir, A.; de Lange, T. Removal of shelterin reveals the telomere end-protection problem. Science 2012, 336, 593–597. [Google Scholar] [CrossRef] [PubMed]
- De Lange, T. How telomeres solve the end-protection problem. Science 2009, 326, 948–952. [Google Scholar] [CrossRef] [PubMed]
- Erdel, F.; Kratz, K.; Willcox, S.; Griffith, J.D.; Greene, E.C.; de Lange, T. Telomere Recognition and Assembly Mechanism of Mammalian Shelterin. Cell Rep. 2017, 18, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Martinez, P.; Blasco, M.A. Telomeric and extra-telomeric roles for telomerase and the telomere-binding proteins. Nat. Rev. Cancer 2011, 11, 161–176. [Google Scholar] [CrossRef] [PubMed]
- Low, K.C.; Tergaonkar, V. Telomerase: Central regulator of all of the hallmarks of cancer. Trends Biochem. Sci. 2013, 38, 426–434. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, E.H.; Greider, C.W.; Szostak, J.W. Telomeres and telomerase: The path from maize, Tetrahymena and yeast to human cancer and aging. Nat. Med. 2006, 12, 1133. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Bryan, T.M.; Reddel, R.R. Increased copy number of the TERT and TERC telomerase subunit genes in cancer cells. Cancer Sci. 2008, 99, 1092–1099. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, J.C.; Cech, T.R. Human telomerase: Biogenesis, trafficking, recruitment, and activation. Genes Dev. 2015, 29, 1095–1105. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.B.; Graham, M.E.; Lovrecz, G.O.; Bache, N.; Robinson, P.J.; Reddel, R.R. Protein composition of catalytically active human telomerase from immortal cells. Science 2007, 315, 1850–1853. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Kim, N.K.; Feigon, J. Architecture of human telomerase RNA. Proc. Natl. Acad. Sci. USA 2011, 108, 20325–20332. [Google Scholar] [CrossRef] [PubMed]
- Holohan, B.; Wright, W.E.; Shay, J.W. Telomeropathies: An emerging spectrum disorder. J. Cell Biol. 2014, 205, 289–299. [Google Scholar] [CrossRef] [PubMed]
- Vinagre, J.; Pinto, V.; Celestino, R.; Reis, M.; Populo, H.; Boaventura, P.; Melo, M.; Catarino, T.; Lima, J.; Lopes, J.M.; et al. Telomerase promoter mutations in cancer: An emerging molecular biomarker? Virchows Arch. 2014, 465, 119–133. [Google Scholar] [CrossRef] [PubMed]
- Aubert, G. Telomere dynamics and aging. Prog. Mol. Biol. Transl. Sci. 2014, 125, 89–111. [Google Scholar] [PubMed]
- Podlevsky, J.D.; Chen, J.J. It all comes together at the ends: Telomerase structure, function, and biogenesis. Mutat. Res. 2012, 730, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Zvereva, M.I.; Shcherbakova, D.M.; Dontsova, O.A. Telomerase: Structure, functions, and activity regulation. Biochemistry 2011, 75, 1563–1583. [Google Scholar] [CrossRef]
- Bourgeron, T.; Xu, Z.; Doumic, M.; Teixeira, M.T. The asymmetry of telomere replication contributes to replicative senescence heterogeneity. Sci. Rep. 2015, 5, 15326. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Otin, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The hallmarks of aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef] [PubMed]
- Kyo, S.; Takakura, M.; Fujiwara, T.; Inoue, M. Understanding and exploiting hTERT promoter regulation for diagnosis and treatment of human cancers. Cancer Sci. 2008, 99, 1528–1538. [Google Scholar] [CrossRef] [PubMed]
- Gonzalo, S.; Blasco, M.A. Role of Rb family in the epigenetic definition of chromatin. Cell Cycle 2005, 4, 752–755. [Google Scholar] [CrossRef] [PubMed]
- Blasco, M.A. Telomere epigenetics: A higher-order control of telomere length in mammalian cells. Carcinogenesis 2004, 25, 1083–1087. [Google Scholar] [CrossRef] [PubMed]
- Gonzalo, S.; Jaco, I.; Fraga, M.F.; Chen, T.; Li, E.; Esteller, M.; Blasco, M.A. DNA methyltransferases control telomere length and telomere recombination in mammalian cells. Nat. Cell Biol. 2006, 8, 416–424. [Google Scholar] [CrossRef] [PubMed]
- García-Cao, M.; O’Sullivan, R.; Peters, A.H.; Jenuwein, T.; Blasco, M.A. Epigenetic regulation of telomere length in mammalian cells by the SUV39H1 and SUV39H2 histone methyltransferases. Nat. Genet. 2004, 36, 94. [Google Scholar] [CrossRef] [PubMed]
- Robin, J.D.; Ludlow, A.T.; Batten, K.; Magdinier, F.; Stadler, G.; Wagner, K.R.; Shay, J.W.; Wright, W.E. Telomere position effect: Regulation of gene expression with progressive telomere shortening over long distances. Genes Dev. 2014, 28, 2464–2476. [Google Scholar] [CrossRef] [PubMed]
- Cesare, A.J.; Reddel, R.R. Alternative lengthening of telomeres: Models, mechanisms and implications. Nat. Rev. Genet. 2010, 11, 319–330. [Google Scholar] [CrossRef] [PubMed]
- Bryan, T.M.; Englezou, A.; Dalla-Pozza, L.; Dunham, M.A.; Reddel, R.R. Evidence for an alternative mechanism for maintaining telomere length in human tumors and tumor-derived cell lines. Nat. Med. 1997, 3, 1271–1274. [Google Scholar] [CrossRef] [PubMed]
- Neumann, A.A.; Watson, C.M.; Noble, J.R.; Pickett, H.A.; Tam, P.P.; Reddel, R.R. Alternative lengthening of telomeres in normal mammalian somatic cells. Genes Dev. 2013, 27, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Novakovic, B.; Napier, C.E.; Vryer, R.; Dimitriadis, E.; Manuelpillai, U.; Sharkey, A.; Craig, J.M.; Reddel, R.R.; Saffery, R. DNA methylation mediated up-regulation of TERRA non-coding RNA is coincident with elongated telomeres in the human placenta. Mol. Hum. Reprod. 2016, 22, 791–799. [Google Scholar] [CrossRef] [PubMed]
- Slatter, T.L.; Tan, X.; Yuen, Y.C.; Gunningham, S.; Ma, S.S.; Daly, E.; Packer, S.; Devenish, C.; Royds, J.A.; Hung, N.A. The alternative lengthening of telomeres pathway may operate in non-neoplastic human cells. J. Pathol. 2012, 226, 509–518. [Google Scholar] [CrossRef] [PubMed]
- Rippe, K.; Luke, B. TERRA and the state of the telomere. Nat. Struct. Mol. Biol. 2015, 22, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Mason, P.J.; Perdigones, N. Telomere biology and translational research. Transl. Res. 2013, 162, 333–342. [Google Scholar] [CrossRef] [PubMed]
- Pestana, A.; Vinagre, J.; Sobrinho-Simoes, M.; Soares, P. TERT biology and function in cancer: Beyond immortalisation. J. Mol. Endocrinol. 2017, 58, R129–R146. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Colgin, L.M.; Reddel, R.R. Telomere maintenance mechanisms and cellular immortalization. Curr. Opin. Genet. Dev. 1999, 9, 97–103. [Google Scholar] [CrossRef]
- Cairney, C.J.; Keith, W.N. Telomerase redefined: Integrated regulation of HTR and hTERT for telomere maintenance and telomerase activity. Biochimie 2008, 90, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Barthel, F.P.; Wei, W.; Tang, M.; Martinez-Ledesma, E.; Hu, X.; Amin, S.B.; Akdemir, K.C.; Seth, S.; Song, X.; Wang, Q.; et al. Systematic analysis of telomere length and somatic alterations in 31 cancer types. Nat. Genet. 2017, 49, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Neidle, S. Quadruplex Nucleic Acids as Novel Therapeutic Targets. J. Med. Chem. 2016, 59, 5987–6011. [Google Scholar] [CrossRef] [PubMed]
- Jager, K.; Walter, M. Therapeutic targeting of telomerase. Genes 2016, 7, 39. [Google Scholar] [CrossRef] [PubMed]
- Ivancich, M.; Schrank, Z.; Wojdyla, L.; Leviskas, B.; Kuckovic, A.; Sanjali, A.; Puri, N. Treating cancer by targeting telomeres and telomerase. Antioxidants 2017, 6, 15. [Google Scholar] [CrossRef] [PubMed]
- Killela, P.J.; Reitman, Z.J.; Jiao, Y.; Bettegowda, C.; Agrawal, N.; Diaz, L.A., Jr.; Friedman, A.H.; Friedman, H.; Gallia, G.L.; Giovanella, B.C.; et al. TERT promoter mutations occur frequently in gliomas and a subset of tumors derived from cells with low rates of self-renewal. Proc. Natl. Acad. Sci. USA 2013, 110, 6021–6026. [Google Scholar] [CrossRef] [PubMed]
- Koelsche, C.; Sahm, F.; Capper, D.; Reuss, D.; Sturm, D.; Jones, D.T.; Kool, M.; Northcott, P.A.; Wiestler, B.; Bohmer, K.; et al. Distribution of TERT promoter mutations in pediatric and adult tumors of the nervous system. Acta Neuropathol. 2013, 126, 907–915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinagre, J.; Almeida, A.; Populo, H.; Batista, R.; Lyra, J.; Pinto, V.; Coelho, R.; Celestino, R.; Prazeres, H.; Lima, L.; et al. Frequency of TERT promoter mutations in human cancers. Nat. Commun. 2013, 4, 2185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, A.; Zheng, C.; Lindvall, C.; Hou, M.; Ekedahl, J.; Lewensohn, R.; Yan, Z.; Yang, X.; Henriksson, M.; Blennow, E.; et al. Frequent amplification of the telomerase reverse transcriptase gene in human tumors. Cancer Res. 2000, 60, 6230–6235. [Google Scholar] [PubMed]
- Saito, T.; Akaike, K.; Kurisaki-Arakawa, A.; Toda-Ishii, M.; Mukaihara, K.; Suehara, Y.; Takagi, T.; Kaneko, K.; Yao, T. TERT promoter mutations are rare in bone and soft tissue sarcomas of Japanese patients. Mol. Clin. Oncol. 2016, 4, 61–64. [Google Scholar] [CrossRef] [PubMed]
- Peifer, M.; Hertwig, F.; Roels, F.; Dreidax, D.; Gartlgruber, M.; Menon, R.; Kramer, A.; Roncaioli, J.L.; Sand, F.; Heuckmann, J.M.; et al. Telomerase activation by genomic rearrangements in high-risk neuroblastoma. Nature 2015, 526, 700–704. [Google Scholar] [CrossRef] [PubMed]
- Heaphy, C.M.; Subhawong, A.P.; Hong, S.M.; Goggins, M.G.; Montgomery, E.A.; Gabrielson, E.; Netto, G.J.; Epstein, J.I.; Lotan, T.L.; Westra, W.H.; et al. Prevalence of the alternative lengthening of telomeres telomere maintenance mechanism in human cancer subtypes. Am. J. Pathol. 2011, 179, 1608–1615. [Google Scholar] [CrossRef] [PubMed]
- Dilley, R.L.; Greenberg, R.A. ALTernative telomere maintenance and cancer. Trends Cancer 2015, 1, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Vinagre, J.; Nabais, J.; Pinheiro, J.; Batista, R.; Oliveira, R.C.; Goncalves, A.P.; Pestana, A.; Reis, M.; Mesquita, B.; Pinto, V.; et al. TERT promoter mutations in pancreatic endocrine tumours are rare and mainly found in tumours from patients with hereditary syndromes. Sci. Rep. 2016, 6, 29714. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, M.; Ogawa, R.; Yoshida, H.; Maeshima, A.; Kanai, Y.; Kinoshita, T.; Hiraoka, N.; Sekine, S. TERT promoter mutations are frequent and show association with MED12 mutations in phyllodes tumors of the breast. Br. J. Cancer 2015, 113, 1244–1248. [Google Scholar] [CrossRef] [PubMed]
- Nozad, S.; Sheehan, C.E.; Gay, L.M.; Elvin, J.A.; Vergilio, J.A.; Suh, J.; Ramkissoon, S.; Schrock, A.B.; Hirshfield, K.M.; Ali, N.; et al. Comprehensive genomic profiling of malignant phyllodes tumors of the breast. Breast Cancer Res. Treat. 2017, 162, 597–602. [Google Scholar] [CrossRef] [PubMed]
- Gay-Bellile, M.; Veronese, L.; Combes, P.; Eymard-Pierre, E.; Kwiatkowski, F.; Dauplat, M.M.; Cayre, A.; Privat, M.; Abrial, C.; Bignon, Y.J.; et al. TERT promoter status and gene copy number gains: Effect on TERT expression and association with prognosis in breast cancer. Oncotarget 2017, 8, 77540–77551. [Google Scholar] [CrossRef] [PubMed]
- Killela, P.J.; Pirozzi, C.J.; Healy, P.; Reitman, Z.J.; Lipp, E.; Rasheed, B.A.; Yang, R.; Diplas, B.H.; Wang, Z.; Greer, P.K.; et al. Mutations in IDH1, IDH2, and in the TERT promoter define clinically distinct subgroups of adult malignant gliomas. Oncotarget 2014, 5, 1515–1525. [Google Scholar] [CrossRef] [PubMed]
- Heidenreich, B.; Rachakonda, P.S.; Hosen, I.; Volz, F.; Hemminki, K.; Weyerbrock, A.; Kumar, R. TERT promoter mutations and telomere length in adult malignant gliomas and recurrences. Oncotarget 2015, 6, 10617–10633. [Google Scholar] [CrossRef] [PubMed]
- Pekmezci, M.; Rice, T.; Molinaro, A.M.; Walsh, K.M.; Decker, P.A.; Hansen, H.; Sicotte, H.; Kollmeyer, T.M.; McCoy, L.S.; Sarkar, G.; Perry, A.; Giannini, C.; et al. Adult infiltrating gliomas with WHO 2016 integrated diagnosis: Additional prognostic roles of ATRX and TERT. Acta Neuropathol. 2017, 133, 1001–1016. [Google Scholar] [CrossRef] [PubMed]
- Arita, H.; Narita, Y.; Fukushima, S.; Tateishi, K.; Matsushita, Y.; Yoshida, A.; Miyakita, Y.; Ohno, M.; Collins, V.P.; Kawahara, N.; et al. Upregulating mutations in the TERT promoter commonly occur in adult malignant gliomas and are strongly associated with total 1p19q loss. Acta Neuropathol. 2013, 126, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Simon, M.; Hosen, I.; Gousias, K.; Rachakonda, S.; Heidenreich, B.; Gessi, M.; Schramm, J.; Hemminki, K.; Waha, A.; Kumar, R. TERT promoter mutations: A novel independent prognostic factor in primary glioblastomas. Neuro-Oncology 2015, 17, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Wang, Y.; Liu, Y.; Liu, X.; Zhang, C.; Wang, L.; Li, S.; Ma, J.; Jiang, T. Brain regions associated with telomerase reverse transcriptase promoter mutations in primary glioblastomas. J. Neuro-Oncol. 2016, 128, 455–462. [Google Scholar] [CrossRef] [PubMed]
- You, H.; Wu, Y.; Chang, K.; Shi, X.; Chen, X.D.; Yan, W.; Li, R. Paradoxical prognostic impact of TERT promoter mutations in gliomas depends on different histological and genetic backgrounds. CNS Neurosci. Ther. 2017, 23, 790–797. [Google Scholar] [CrossRef] [PubMed]
- Mosrati, M.A.; Malmstrom, A.; Lysiak, M.; Krysztofiak, A.; Hallbeck, M.; Milos, P.; Hallbeck, A.L.; Bratthall, C.; Strandeus, M.; Stenmark-Askmalm, M.; et al. TERT promoter mutations and polymorphisms as prognostic factors in primary glioblastoma. Oncotarget 2015, 6, 16663–16673. [Google Scholar] [CrossRef] [PubMed]
- Nencha, U.; Rahimian, A.; Giry, M.; Sechi, A.; Mokhtari, K.; Polivka, M.; Schmitt, Y.; Di Stefano, A.L.; Alentorn, A.; Labussiere, M.; et al. TERT promoter mutations and rs2853669 polymorphism: Prognostic impact and interactions with common alterations in glioblastomas. J. Neuro-Oncol. 2016, 126, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wu, G.; Shan, Y.; Hartmann, C.; von Deimling, A.; Xing, M. Highly prevalent TERT promoter mutations in bladder cancer and glioblastoma. Cell Cycle 2013, 12, 1637–1638. [Google Scholar] [CrossRef] [PubMed]
- Eckel-Passow, J.E.; Lachance, D.H.; Molinaro, A.M.; Walsh, K.M.; Decker, P.A.; Sicotte, H.; Pekmezci, M.; Rice, T.; Kosel, M.L.; Smirnov, I.V.; et al. Glioma groups based on 1p/19q, IDH, and TERT promoter mutations in tumors. N. Engl. J. Med. 2015, 372, 2499–2508. [Google Scholar] [CrossRef] [PubMed]
- Arita, H.; Narita, Y.; Takami, H.; Fukushima, S.; Matsushita, Y.; Yoshida, A.; Miyakita, Y.; Ohno, M.; Shibui, S.; Ichimura, K. TERT promoter mutations rather than methylation are the main mechanism for TERT upregulation in adult gliomas. Acta Neuropathol. 2013, 126, 939–941. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; Cai, J.; Yan, W.; Zhang, W.; Wang, Y.; Chen, B.; Li, G.; Li, S.; Wu, C.; Yao, K.; et al. Classification based on mutations of TERT promoter and IDH characterizes subtypes in grade II/III gliomas. Neuro-Oncology 2016, 18, 1099–1108. [Google Scholar] [CrossRef] [PubMed]
- Labussiere, M.; Di Stefano, A.L.; Gleize, V.; Boisselier, B.; Giry, M.; Mangesius, S.; Bruno, A.; Paterra, R.; Marie, Y.; Rahimian, A.; et al. TERT promoter mutations in gliomas, genetic associations and clinico-pathological correlations. Br. J. Cancer 2014, 111, 2024–2032. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.S.; Wang, Z.; He, X.J.; Diplas, B.H.; Yang, R.; Killela, P.J.; Meng, Q.; Ye, Z.Y.; Wang, W.; Jiang, X.T.; et al. Recurrent TERT promoter mutations identified in a large-scale study of multiple tumour types are associated with increased TERT expression and telomerase activation. Eur. J. Cancer 2015, 51, 969–976. [Google Scholar] [CrossRef] [PubMed]
- Batista, R.; Cruvinel-Carloni, A.; Vinagre, J.; Peixoto, J.; Catarino, T.A.; Campanella, N.C.; Menezes, W.; Becker, A.P.; de Almeida, G.C.; Matsushita, M.M.; et al. The prognostic impact of TERT promoter mutations in glioblastomas is modified by the rs2853669 single nucleotide polymorphism. Int. J. Cancer 2016, 139, 414–423. [Google Scholar] [CrossRef] [PubMed]
- Gao, K.; Li, G.; Qu, Y.; Wang, M.; Cui, B.; Ji, M.; Shi, B.; Hou, P. TERT promoter mutations and long telomere length predict poor survival and radiotherapy resistance in gliomas. Oncotarget 2016, 7, 8712–8725. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Koh, J.; Kim, S.I.; Won, J.K.; Park, C.K.; Choi, S.H.; Park, S.H. The frequency and prognostic effect of TERT promoter mutation in diffuse gliomas. Acta Neuropathol. Commun. 2017, 5, 62. [Google Scholar] [CrossRef] [PubMed]
- Arita, H.; Yamasaki, K.; Matsushita, Y.; Nakamura, T.; Shimokawa, A.; Takami, H.; Tanaka, S.; Mukasa, A.; Shirahata, M.; Shimizu, S.; et al. A combination of TERT promoter mutation and MGMT methylation status predicts clinically relevant subgroups of newly diagnosed glioblastomas. Acta Neuropathol. Commun. 2016, 4, 79. [Google Scholar] [CrossRef] [PubMed]
- Spiegl-Kreinecker, S.; Lotsch, D.; Ghanim, B.; Pirker, C.; Mohr, T.; Laaber, M.; Weis, S.; Olschowski, A.; Webersinke, G.; Pichler, J.; et al. Prognostic quality of activating TERT promoter mutations in glioblastoma: Interaction with the rs2853669 polymorphism and patient age at diagnosis. Neuro-Oncology 2015, 17, 1231–1240. [Google Scholar] [CrossRef] [PubMed]
- Nonoguchi, N.; Ohta, T.; Oh, J.E.; Kim, Y.H.; Kleihues, P.; Ohgaki, H. TERT promoter mutations in primary and secondary glioblastomas. Acta Neuropathol. 2013, 126, 931–937. [Google Scholar] [CrossRef] [PubMed]
- Jeong, D.E.; Woo, S.R.; Nam, H.; Nam, D.H.; Lee, J.H.; Joo, K.M. Preclinical and clinical implications of TERT promoter mutation in glioblastoma multiforme. Oncol. Lett. 2017, 14, 8213–8219. [Google Scholar] [CrossRef] [PubMed]
- Brennan, C.W.; Verhaak, R.G.; McKenna, A.; Campos, B.; Noushmehr, H.; Salama, S.R.; Zheng, S.; Chakravarty, D.; Sanborn, J.Z.; Berman, S.H.; et al. The somatic genomic landscape of glioblastoma. Cell 2013, 155, 462–477. [Google Scholar] [CrossRef] [PubMed]
- Remke, M.; Ramaswamy, V.; Peacock, J.; Shih, D.J.; Koelsche, C.; Northcott, P.A.; Hill, N.; Cavalli, F.M.; Kool, M.; Wang, X.; et al. TERT promoter mutations are highly recurrent in SHH subgroup medulloblastoma. Acta Neuropathol. 2013, 126, 917–929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kool, M.; Jones, D.T.; Jager, N.; Northcott, P.A.; Pugh, T.J.; Hovestadt, V.; Piro, R.M.; Esparza, L.A.; Markant, S.L.; Remke, M.; et al. Genome sequencing of SHH medulloblastoma predicts genotype-related response to smoothened inhibition. Cancer Cell 2014, 25, 393–405. [Google Scholar] [CrossRef] [PubMed]
- Lindsey, J.C.; Schwalbe, E.C.; Potluri, S.; Bailey, S.; Williamson, D.; Clifford, S.C. TERT promoter mutation and aberrant hypermethylation are associated with elevated expression in medulloblastoma and characterise the majority of non-infant SHH subgroup tumours. Acta Neuropathol. 2014, 127, 307–309. [Google Scholar] [CrossRef] [PubMed]
- Viana-Pereira, M.; Almeida, G.C.; Stavale, J.N.; Malheiro, S.; Clara, C.; Lobo, P.; Pimentel, J.; Reis, R.M. Study of hTERT and Histone 3 Mutations in Medulloblastoma. Pathobiology 2017, 84, 108–113. [Google Scholar] [CrossRef] [PubMed]
- Goutagny, S.; Nault, J.C.; Mallet, M.; Henin, D.; Rossi, J.Z.; Kalamarides, M. High incidence of activating TERT promoter mutations in meningiomas undergoing malignant progression. Brain Pathol. 2014, 24, 184–189. [Google Scholar] [CrossRef] [PubMed]
- Sahm, F.; Schrimpf, D.; Olar, A.; Koelsche, C.; Reuss, D.; Bissel, J.; Kratz, A.; Capper, D.; Schefzyk, S.; Hielscher, T.; et al. TERT Promoter Mutations and Risk of Recurrence in Meningioma. J. Natl. Cancer Inst. 2016, 108. [Google Scholar] [CrossRef] [PubMed]
- Koopmans, A.E.; Ober, K.; Dubbink, H.J.; Paridaens, D.; Naus, N.C.; Belunek, S.; Krist, B.; Post, E.; Zwarthoff, E.C.; de Klein, A. Prevalence and Implications of TERT Promoter Mutation in Uveal and Conjunctival Melanoma and in Benign and Premalignant Conjunctival Melanocytic Lesions. Investig. Ophthalmol. Vis. Sci. 2014, 55, 6024–6030. [Google Scholar] [CrossRef] [PubMed]
- Griewank, K.G.; Murali, R.; Schilling, B.; Scholz, S.; Sucker, A.; Song, M.; Susskind, D.; Grabellus, F.; Zimmer, L.; Hillen, U.; et al. TERT promoter mutations in ocular melanoma distinguish between conjunctival and uveal tumours. Br. J. Cancer 2013, 109, 497–501. [Google Scholar] [CrossRef] [PubMed]
- Griewank, K.G.; Murali, R.; Puig-Butille, J.A.; Schilling, B.; Livingstone, E.; Potrony, M.; Carrera, C.; Schimming, T.; Moller, I.; Schwamborn, M.; et al. TERT Promoter Mutation Status as an Independent Prognostic Factor in Cutaneous Melanoma. JNCI J. Natl. Cancer Inst. 2014, 106. [Google Scholar] [CrossRef] [PubMed]
- Scholz, S.L.; Thomasen, H.; Reis, H.; Moller, I.; Darawsha, R.; Muller, B.; Dekowski, D.; Sucker, A.; Schilling, B.; Schadendorf, D.; et al. Frequent TERT promoter mutations in ocular surface squamous neoplasia. Investig. Ophthalmol. Vis. Sci. 2015, 56, 5854–5861. [Google Scholar] [CrossRef] [PubMed]
- Dono, M.; Angelini, G.; Cecconi, M.; Amaro, A.; Esposito, A.I.; Mirisola, V.; Maric, I.; Lanza, F.; Nasciuti, F.; Viaggi, S.; et al. Mutation frequencies of GNAQ, GNA11, BAP1, SF3B1, EIF1AX and TERT in uveal melanoma: Detection of an activating mutation in the TERT gene promoter in a single case of uveal melanoma. Br. J. Cancer 2014, 110, 1058–1065. [Google Scholar] [CrossRef] [PubMed]
- Pilati, C.; Letouze, E.; Nault, J.C.; Imbeaud, S.; Boulai, A.; Calderaro, J.; Poussin, K.; Franconi, A.; Couchy, G.; Morcrette, G.; et al. Genomic profiling of hepatocellular adenomas reveals recurrent FRK-activating mutations and the mechanisms of malignant transformation. Cancer Cell 2014, 25, 428–441. [Google Scholar] [CrossRef] [PubMed]
- Nault, J.C.; Mallet, M.; Pilati, C.; Calderaro, J.; Bioulac-Sage, P.; Laurent, C.; Laurent, A.; Cherqui, D.; Balabaud, C.; Zucman-Rossi, J. High frequency of telomerase reverse-transcriptase promoter somatic mutations in hepatocellular carcinoma and preneoplastic lesions. Nat. Commun. 2013, 4, 2218. [Google Scholar] [CrossRef] [PubMed]
- Quaas, A.; Oldopp, T.; Harun, L.; Klingenfeld, C.; Krech, T.; Sauter, G.; Grob, T.J. Frequency of TERT promoter mutations in primary tumors of the liver. Virchows Arch. 2014, 465, 673–677. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Han, S.; Meng, L.; Li, Z.; Zhang, X.; Wu, A. TERT promoter mutations lead to high transcriptional activity under hypoxia and temozolomide treatment and predict poor prognosis in gliomas. PLoS ONE 2014, 9, e100297. [Google Scholar] [CrossRef] [PubMed]
- Totoki, Y.; Tatsuno, K.; Covington, K.R.; Ueda, H.; Creighton, C.J.; Kato, M.; Tsuji, S.; Donehower, L.A.; Slagle, B.L.; Nakamura, H.; et al. Trans-ancestry mutational landscape of hepatocellular carcinoma genomes. Nat. Genet. 2014, 46, 1267–1273. [Google Scholar] [CrossRef] [PubMed]
- Ally, A.; Balasundaram, M.; Carlsen, R.; Chuah, E.; Clarke, A.; Dhalla, N.; Holt, R.A.; Jones, S.J.; Lee, D.; Ma, Y.; et al. Comprehensive and integrative genomic characterization of hepatocellular carcinoma. Cell 2017, 169, 1327–1341. [Google Scholar] [CrossRef] [PubMed]
- Cevik, D.; Yildiz, G.; Ozturk, M. Common telomerase reverse transcriptase promoter mutations in hepatocellular carcinomas from different geographical locations. World J. Gastroenterol. 2015, 21, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, A.; Furuta, M.; Shiraishi, Y.; Gotoh, K.; Kawakami, Y.; Arihiro, K.; Nakamura, T.; Ueno, M.; Ariizumi, S.; Nguyen, H.H.; et al. Whole-genome mutational landscape of liver cancers displaying biliary phenotype reveals hepatitis impact and molecular diversity. Nat. Commun. 2015, 6, 6120. [Google Scholar] [CrossRef] [PubMed]
- Schulze, K.; Imbeaud, S.; Letouze, E.; Alexandrov, L.B.; Calderaro, J.; Rebouissou, S.; Couchy, G.; Meiller, C.; Shinde, J.; Soysouvanh, F.; et al. Exome sequencing of hepatocellular carcinomas identifies new mutational signatures and potential therapeutic targets. Nat. Genet. 2015, 47, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.E.; Chang, S.H.; Kim, W.Y.; Lim, S.D.; Kim, W.S.; Hwang, T.S.; Han, H.S. Frequent somatic TERT promoter mutations and CTNNB1 mutations in hepatocellular carcinoma. Oncotarget 2016, 7, 69267–69275. [Google Scholar] [CrossRef] [PubMed]
- Pezzuto, F.; Izzo, F.; Buonaguro, L.; Annunziata, C.; Tatangelo, F.; Botti, G.; Buonaguro, F.M.; Tornesello, M.L. Tumor specific mutations in TERT promoter and CTNNB1 gene in hepatitis B and hepatitis C related hepatocellular carcinoma. Oncotarget 2016, 7, 54253–54262. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Guo, X.; Chen, Y.; Chen, G.; Ma, Y.; Huang, K.; Zhang, Y.; Zhao, Q.; Winkler, C.A.; An, P.; et al. Telomerase reverse transcriptase promoter mutations in hepatitis B virus-associated hepatocellular carcinoma. Oncotarget 2016, 7, 27838–27847. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Zhou, W.; Li, C.; Yang, Y.; Shang, Y.K.; Chen, C.; Zhang, J.; Yao, R.; Wang, P.; Wen, W.; et al. Promoter mutations and cellular distribution of telomerase in non-clear cell and clear cell hepatocellular carcinoma. Oncotarget 2017, 8, 26288–26297. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.W.; Park, T.I.; Jang, S.Y.; Park, S.Y.; Park, W.J.; Jung, S.J.; Lee, J.H. Clinicopathological characteristics of TERT promoter mutation and telomere length in hepatocellular carcinoma. Medicine 2017, 96, e5766. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.T.; Cheng, G.H.; Yu, J.Y.; Zheng, S.Z.; Sun, C.; Sun, Q.; Li, K.L.; Lin, Z.M.; Liu, T.T.; Li, P.; et al. The TERT promoter mutation incidence is modified by germline TERT rs2736098 and rs2736100 polymorphisms in hepatocellular carcinoma. Oncotarget 2017, 8, 23120–23129. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.; Shi, L.; Wang, D.; Zhang, B.; Yang, Q.; Ji, M.; Shi, B.; Hou, P. Low frequency of TERT promoter mutations in a large cohort of gallbladder and gastric cancers. Int. J. Cancer 2014, 134, 2993–2994. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Gao, Y.; Chen, Z.; Hu, X.; Zhou, F.; He, J. Low frequency of TERT promoter somatic mutation in 313 sporadic esophageal squamous cell carcinomas. Int. J. Cancer 2014, 134, 493–494. [Google Scholar] [CrossRef] [PubMed]
- Van Nistelrooij, A.M.; Zwarthoff, E.C.; Post, E.; Lurkin, I.; van Marion, R.; Korpershoek, E.; Biermann, K.; Wijnhoven, B.P.; Dinjens, W.N. Absence of TERT promoter mutations in esophageal adenocarcinoma. Int. J. Cancer 2014, 134, 2014–2015. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Liang, X.; Bjorkholm, M.; Jia, J.; Xu, D. The absence of TERT promoter mutations in primary gastric cancer. Gene 2014, 540, 266–267. [Google Scholar] [CrossRef] [PubMed]
- Eichenmuller, M.; Trippel, F.; Kreuder, M.; Beck, A.; Schwarzmayr, T.; Haberle, B.; Cairo, S.; Leuschner, I.; von Schweinitz, D.; Strom, T.M.; et al. The genomic landscape of hepatoblastoma and their progenies with HCC-like features. J. Hepatol. 2014, 61, 1312–1320. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Liu, T.T.; Sofiadis, A.; Juhlin, C.C.; Zedenius, J.; Hoog, A.; Larsson, C.; Xu, D.W. TERT Promoter Mutation as an Early Genetic Event Activating Telomerase in Follicular Thyroid Adenoma (FTA) and Atypical FTA. Cancer 2014, 120, 2965–2979. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Bishop, J.; Shan, Y.; Pai, S.; Liu, D.; Murugan, A.K.; Sun, H.; El-Naggar, A.K.; Xing, M. Highly prevalent TERT promoter mutations in aggressive thyroid cancers. Endocr. Relat. Cancer 2013, 20, 603–610. [Google Scholar] [CrossRef] [PubMed]
- Xing, M.Z.; Liu, R.Y.; Liu, X.L.; Murugan, A.K.; Zhu, G.W.; Zeiger, M.A.; Pai, S.; Bishop, J. BRAFV600E and TERT promoter mutations cooperatively identify the most aggressive papillary thyroid cancer with highest recurrence. J. Clin. Oncol. 2014, 32, 2718. [Google Scholar] [CrossRef] [PubMed]
- Gandolfi, G.; Ragazzi, M.; Frasoldati, A.; Piana, S.; Ciarrocchi, A.; Sancisi, V. TERT promoter mutations are associated with distant metastases in papillary thyroid carcinoma. Eur. J. Endocrinol. 2015, 172, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Muzza, M.; Colombo, C.; Rossi, S.; Tosi, D.; Cirello, V.; Perrino, M.; De Leo, S.; Magnani, E.; Pignatti, E.; Vigo, B.; et al. Telomerase in differentiated thyroid cancer: Promoter mutations, expression and localization. Mol. Cell. Endocrinol. 2015, 399, 288–295. [Google Scholar] [CrossRef] [PubMed]
- Qasem, E.; Murugan, A.K.; Al-Hindi, H.; Xing, M.; Almohanna, M.; Alswailem, M.; Alzahrani, A.S. TERT promoter mutations in thyroid cancer: A report from a Middle Eastern population. Endocr. Relat. Cancer 2015, 22, 901–908. [Google Scholar] [CrossRef] [PubMed]
- Jeon, M.J.; Kim, W.G.; Sim, S.; Lim, S.; Kwon, H.; Kim, T.Y.; Shong, Y.K.; Kim, W.B. Low prevalence of somatic tert promoter mutations in classic papillary thyroid carcinoma. Endocrinol. Metab. 2016, 31, 100–104. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.E.; Hwang, T.S.; Choi, Y.-L.; Han, H.S.; Kim, W.S.; Jang, M.H.; Kim, S.K.; Yang, J.H. Prognostic significance of TERT promoter mutations in papillary thyroid carcinomas in a BRAFV600E mutation–prevalent population. Thyroid 2016, 26, 901–910. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Zhang, J.; Lu, J.; Gao, J.; Ren, X.; Teng, L.; Duan, H.; Lin, Y.; Li, X.; Zhang, B.; et al. BRAFV600E and TERT promoter mutations in papillary thyroid carcinoma in chinese patients. PLoS ONE 2016, 11, e0153319. [Google Scholar] [CrossRef]
- Bullock, M.; Ren, Y.; O’Neill, C.; Gill, A.; Aniss, A.; Sywak, M.; Sidhu, S.; Delbridge, L.; Learoyd, D.; de Vathaire, F.; et al. TERT promoter mutations are a major indicator of recurrence and death due to papillary thyroid carcinomas. Clin. Endocrinol. 2016, 85, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Melo, M.; da Rocha, A.G.; Vinagre, J.; Batista, R.; Peixoto, J.; Tavares, C.; Celestino, R.; Almeida, A.; Salgado, C.; Eloy, C.; et al. TERT promoter mutations are a major indicator of poor outcome in differentiated thyroid carcinomas. J. Clin. Endocrinol. Metab. 2014, 99, E754–E765. [Google Scholar] [CrossRef] [PubMed]
- Dettmer, M.S.; Schmitt, A.; Steinert, H.; Capper, D.; Moch, H.; Komminoth, P.; Perren, A. Tall cell papillary thyroid carcinoma: New diagnostic criteria and mutations in BRAF and TERT. Endocr. Relat. Cancer 2015, 22, 419–429. [Google Scholar] [CrossRef] [PubMed]
- Landa, I.; Ganly, I.; Chan, T.A.; Mitsutake, N.; Matsuse, M.; Ibrahimpasic, T.; Ghossein, R.A.; Fagin, J.A. Frequent somatic TERT promoter mutations in thyroid cancer: Higher prevalence in advanced forms of the disease. J. Clin. Endocrinol. Metab. 2013, 98, E1562–E1566. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Xing, M. Diagnostic and prognostic TERT promoter mutations in thyroid fine-needle aspiration biopsy. Endocr. Relat. Cancer 2014, 21, 825–830. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Wang, N.; Cao, J.; Sofiadis, A.; Dinets, A.; Zedenius, J.; Larsson, C.; Xu, D. The age- and shorter telomere-dependent TERT promoter mutation in follicular thyroid cell-derived carcinomas. Oncogene 2014, 33, 4978–4984. [Google Scholar] [CrossRef] [PubMed]
- George, J.R.; Henderson, Y.C.; Williams, M.D.; Roberts, D.B.; Hei, H.; Lai, S.Y.; Clayman, G.L. Association of TERT promoter mutation, but not BRAF mutation, with increased mortality in PTC. J. Clin. Endocrinol. Metab. 2015, 100, E1550–E1559. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Chen, E.; Dong, S.; Cai, Y.; Zhang, X.; Zhou, Y.; Zeng, R.; Yang, F.; Pan, C.; Liu, Y.; et al. BRAF and TERT promoter mutations in the aggressiveness of papillary thyroid carcinoma: A study of 653 patients. Oncotarget 2016, 7, 18346–18355. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.H.; Kim, Y.E.; Ahn, S.; Kim, J.Y.; Ki, C.S.; Oh, Y.L.; Kim, K.; Yun, J.W.; Park, W.Y.; Choe, J.H.; et al. TERT promoter mutations and long-term survival in patients with thyroid cancer. Endocr. Relat. Cancer 2016, 23, 813–823. [Google Scholar] [CrossRef] [PubMed]
- Landa, I.; Ibrahimpasic, T.; Boucai, L.; Sinha, R.; Knauf, J.A.; Shah, R.H.; Dogan, S.; Ricarte-Filho, J.C.; Krishnamoorthy, G.P.; Xu, B.; et al. Genomic and transcriptomic hallmarks of poorly differentiated and anaplastic thyroid cancers. J. Clin. Investig. 2016, 126, 1052–1066. [Google Scholar] [CrossRef] [PubMed]
- Myung, J.K.; Kwak, B.K.; Lim, J.A.; Lee, M.C.; Kim, M.J. TERT Promoter mutations and tumor persistence/recurrence in papillary thyroid cancer. Cancer Res. Treat. 2016, 48, 942–947. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.; Kim, T.H.; Kim, S.W.; Ki, C.S.; Jang, H.W.; Kim, J.S.; Kim, J.H.; Choe, J.H.; Shin, J.H.; Hahn, S.Y.; et al. Comprehensive screening for PD-L1 expression in thyroid cancer. Endocr. Relat. Cancer 2017, 24, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.H.; Ki, C.S.; Kim, H.S.; Kim, K.; Choe, J.H.; Kim, J.H.; Kim, J.S.; Oh, Y.L.; Hahn, S.Y.; Shin, J.H.; et al. Refining dynamic risk stratification and prognostic groups for differentiated thyroid cancer with TERT promoter mutations. J. Clin. Endocrinol. Metab. 2017, 102, 1757–1764. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.H.; Ki, C.S.; Hahn, S.Y.; Oh, Y.L.; Jang, H.W.; Kim, S.W.; Chung, J.H.; Shin, J.H. Ultrasonographic prediction of highly aggressive telomerase reverse transcriptase (TERT) promoter-mutated papillary thyroid cancer. Endocrine 2017, 57, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Melo, M.; Gaspar da Rocha, A.; Batista, R.; Vinagre, J.; Martins, M.J.; Costa, G.; Ribeiro, C.; Carrilho, F.; Leite, V.; Lobo, C.; et al. TERT, BRAF, and NRAS in primary thyroid cancer and metastatic disease. J. Clin. Endocrinol. Metab. 2017, 102, 1898–1907. [Google Scholar] [CrossRef] [PubMed]
- Boaventura, P.; Batista, R.; Pestana, A.; Reis, M.; Mendes, A.; Eloy, C.; Sobrinho-Simoes, M.; Soares, P. TERT promoter mutations: A genetic signature of benign and malignant thyroid tumours occurring in the context of tinea capitis irradiation. Eur. J. Endocrinol. 2017, 176, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Chindris, A.M.; Casler, J.D.; Bernet, V.J.; Rivera, M.; Thomas, C.; Kachergus, J.M.; Necela, B.M.; Hay, I.D.; Westphal, S.A.; Grant, C.S.; et al. Clinical and molecular features of Hurthle cell carcinoma of the thyroid. J. Clin. Endocrinol. Metab. 2015, 100, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Liu, R.; Qu, S.; Zhu, G.; Bishop, J.; Liu, X.; Sun, H.; Shan, Z.; Wang, E.; Luo, Y.; et al. Association of TERT promoter mutation 1,295,228 C>T with BRAFV600E mutation, older patient age, and distant metastasis in anaplastic thyroid cancer. J. Clin. Endocrinol. Metab. 2015, 100, E632–E637. [Google Scholar] [CrossRef] [PubMed]
- Oishi, N.; Kondo, T.; Ebina, A.; Sato, Y.; Akaishi, J.; Hino, R.; Yamamoto, N.; Mochizuki, K.; Nakazawa, T.; Yokomichi, H.; et al. Molecular alterations of coexisting thyroid papillary carcinoma and anaplastic carcinoma: Identification of TERT mutation as an independent risk factor for transformation. Mod. Pathol. 2017, 30, 1527–1537. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Li, J.; Li, X.; Liang, Z.; Gao, W.; Liang, J.; Cheng, S.; Lin, Y. TERT promoter mutation predicts radioiodine-refractory character in distant metastatic differentiated thyroid cancer. J. Nucl. Med. 2017, 58, 258–265. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.T.; Brown, T.C.; Juhlin, C.C.; Andreasson, A.; Wang, N.; Backdahl, M.; Healy, J.M.; Prasad, M.L.; Korah, R.; Carling, T.; et al. The activating TERT promoter mutation C228T is recurrent in subsets of adrenal tumors. Endocr. Relat. Cancer 2014, 21, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Martins, C.S.; de Castro, M.; Calado, R.T. Absence of TERT promoter mutations in pituitary adenomas. J. Endocrinol. Investig. 2016, 39, 933–934. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.L.; Qu, S.; Liu, R.Y.; Sheng, C.J.; Shi, X.G.; Zhu, G.W.; Murugan, A.K.; Guan, H.X.; Yu, H.Y.; Wang, Y.G.; et al. TERT Promoter Mutations and Their Association with BRAFV600E Mutation and Aggressive Clinicopathological Characteristics of Thyroid Cancer. J. Clin. Endocrinol. Metab. 2014, 99, E1130–E1136. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Kim, Y.; Jeon, S.; Bae, J.S.; Jung, S.L.; Jung, C.K. Cytologic, clinicopathologic, and molecular features of papillary thyroid carcinoma with prominent hobnail features: 10 case reports and systematic literature review. Int. J. Clin. Exp. Pathol. 2015, 8, 7988–7997. [Google Scholar] [PubMed]
- De Biase, D.; Gandolfi, G.; Ragazzi, M.; Eszlinger, M.; Sancisi, V.; Gugnoni, M.; Visani, M.; Pession, A.; Casadei, G.; Durante, C.; et al. TERT promoter mutations in papillary thyroid microcarcinomas. Thyroid 2015, 25, 1013–1019. [Google Scholar] [CrossRef] [PubMed]
- Yabuta, T.; Matsuse, M.; Hirokawa, M.; Yamashita, S.; Mitsutake, N.; Miyauchi, A. TERT promoter mutations were not found in papillary thyroid microcarcinomas that showed disease progression on active surveillance. Thyroid 2017, 27, 1206–1207. [Google Scholar] [CrossRef] [PubMed]
- Alzahrani, A.S.; Qasem, E.; Murugan, A.K.; Al-Hindi, H.N.; AlKhafaji, D.; Almohanna, M.; Xing, M.; Alhomaidah, D.; AlSwailem, M. Uncommon TERT promoter mutations in pediatric thyroid cancer. Thyroid 2016, 26, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Onder, S.; Ozturk, S.S.; Yegen, G.; Sormaz, I.C.; Yilmaz, I.; Poyrazoglu, S.; Sanli, Y.; Senyurek, Y.G.; Kapran, Y.; Mete, O. Classic architecture with multicentricity and local recurrence, and absence of TERT Promoter mutations are correlates of BRAFV600E harboring pediatric papillary thyroid carcinomas. Endocr. Pathol. 2016, 27, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.C.; Ayhan, A.; Maeda, D.; Kim, K.R.; Clarke, B.A.; Shaw, P.; Chui, M.H.; Rosen, B.; Shih, I.M.; Wang, T.L. Frequent somatic mutations of the telomerase reverse transcriptase promoter in ovarian clear cell carcinoma but not in other major types of gynaecological malignancy. J. Pathol. 2014, 232, 473–481. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.N.; Chiang, Y.C.; Cheng, W.F.; Chen, C.A.; Lin, M.C.; Kuo, K.T. Molecular alterations in endometrial and ovarian clear cell carcinomas: Clinical impacts of telomerase reverse transcriptase promoter mutation. Mod. Pathol. 2015, 28, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Cheng, K.A.; Kurtis, B.; Babayeva, S.; Zhuge, J.; Tantchou, I.; Cai, D.; Lafaro, R.J.; Fallon, J.T.; Zhong, M. Heterogeneity of TERT promoter mutations status in squamous cell carcinomas of different anatomical sites. Ann. Diagn. Pathol. 2015, 19, 146–148. [Google Scholar] [CrossRef] [PubMed]
- Vinothkumar, V.; Arunkumar, G.; Revathidevi, S.; Arun, K.; Manikandan, M.; Kuha, A.; Rao, M.; Rajkumar, K.S.; Ajay, C.; Rajaraman, R. TERT promoter hot spot mutations are frequent in Indian cervical and oral squamous cell carcinomas. Tumor Biol. 2016, 37, 7907–7913. [Google Scholar] [CrossRef] [PubMed]
- Dias-Santagata, D.; Selim, M.A.; Su, Y.; Peng, Y.; Vollmer, R.; Chlopik, A.; Tell-Marti, G.; Paral, K.M.; Shalin, S.C.; Shea, C.R.; et al. KIT mutations and CD117 overexpression are markers of better progression-free survival in vulvar melanomas. Br. J. Dermatol. 2017, 177, 1376–1384. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.; Dang, S.; Wu, K.; Shao, Y.; Yang, Q.; Ji, M.; Shi, B.; Hou, P. TERT promoter mutations predict worse survival in laryngeal cancer patients. Int. J. Cancer 2014, 135, 1008–1010. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.P.; Wang, C.I.; Pickering, C.R.; Huang, Y.; Tsai, C.N.; Tsang, N.M.; Kao, H.K.; Cheng, M.H.; Myers, J.N. Prevalence of promoter mutations in the TERT gene in oral cavity squamous cell carcinoma. Head Neck 2017, 39, 1131–1137. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Ahn, D.; Sohn, J.H.; Kim, Y.H.; Lee, J.H.; Lee, H. TERT Promoter Mutation and Telomere Length in Salivary Gland Tumors. Pathol. Oncol. Res. 2017. [Google Scholar] [CrossRef]
- Lam, G.; Xian, R.R.; Li, Y.; Burns, K.H.; Beemon, K.L. Lack of TERT Promoter Mutations in Human B-Cell Non-Hodgkin Lymphoma. Genes 2016, 7, 93. [Google Scholar] [CrossRef] [PubMed]
- Panero, J.; Alves-Paiva, R.M.; Roisman, A.; Santana-Lemos, B.A.; Falcao, R.P.; Oliveira, G.; Martins, D.; Stanganelli, C.; Slavutsky, I.; Calado, R.T. Acquired TERT promoter mutations stimulate TERT transcription in mantle cell lymphoma. Am. J. Hematol. 2016, 91, 481–485. [Google Scholar] [CrossRef] [PubMed]
- Mosrati, M.A.; Willander, K.; Falk, I.J.; Hermanson, M.; Hoglund, M.; Stockelberg, D.; Wei, Y.; Lotfi, K.; Soderkvist, P. Association between TERT promoter polymorphisms and acute myeloid leukemia risk and prognosis. Oncotarget 2015, 6, 25109–25120. [Google Scholar] [CrossRef] [PubMed]
- Tallet, A.; Nault, J.C.; Renier, A.; Hysi, I.; Galateau-Salle, F.; Cazes, A.; Copin, M.C.; Hofman, P.; Andujar, P.; Le Pimpec-Barthes, F.; et al. Overexpression and promoter mutation of the TERT gene in malignant pleural mesothelioma. Oncogene 2014, 33, 3748–3752. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Gong, R.; Wang, R.; Pan, Y.; Cai, D.; Pan, B.; Li, Y.; Xiang, J.; Li, H.; Zhang, J.; et al. Recurrent TERT promoter mutations in non-small cell lung cancers. Lung Cancer 2014, 86, 369–373. [Google Scholar] [CrossRef] [PubMed]
- Yuan, P.; Cao, J.L.; Abuduwufuer, A.; Wang, L.M.; Yuan, X.S.; Lv, W.; Hu, J. Clinical Characteristics and Prognostic Significance of TERT Promoter Mutations in Cancer: A Cohort Study and a Meta-Analysis. PLoS ONE 2016, 11, e0146803. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Hao, L.; Li, Y.; Wang, S.; Chen, H.; Zhang, L.; Ke, B.; Yin, Y.; Suo, H.; Sun, B.; et al. Prognostic value analysis of mutational and clinicopathological factors in non-small cell lung cancer. PLoS ONE 2014, 9, e107276. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.J.; Kim, D.S.; Park, W.J.; Lee, H.; Choi, I.J.; Park, J.Y.; Lee, J.H. Mutation of the TERT promoter leads to poor prognosis of patients with non-small cell lung cancer. Oncol. Lett. 2017, 14, 1609–1614. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Zhuge, J.; Bezerra, S.M.; Faraj, S.F.; Munari, E.; Fallon, J.T.; Yang, X.J.; Argani, P.; Netto, G.J.; Zhong, M. High frequency of TERT promoter mutation in small cell carcinoma of bladder, but not in small cell carcinoma of other origins. J. Hematol. Oncol. 2014, 7, 47. [Google Scholar] [CrossRef] [PubMed]
- Lindner, S.; Bachmann, H.S.; Odersky, A.; Schaefers, S.; Klein-Hitpass, L.; Hero, B.; Fischer, M.; Eggert, A.; Schramm, A.; Schulte, J.H. Absence of telomerase reverse transcriptase promoter mutations in neuroblastoma. Biomed. Rep. 2015, 3, 443–446. [Google Scholar] [CrossRef] [PubMed]
- Griewank, K.G.; Murali, R.; Schilling, B.; Schimming, T.; Moller, I.; Moll, I.; Schwamborn, M.; Sucker, A.; Zimmer, L.; Schadendorf, D.; et al. TERT promoter mutations are frequent in cutaneous basal cell carcinoma and squamous cell carcinoma. PLoS ONE 2013, 8, e80354. [Google Scholar] [CrossRef] [PubMed]
- Populo, H.; Boaventura, P.; Vinagre, J.; Batista, R.; Mendes, A.; Caldas, R.; Pardal, J.; Azevedo, F.; Honavar, M.; Guimaraes, I.; et al. TERT promoter mutations in skin cancer: The effects of sun exposure and X-irradiation. J. Investig. Dermatol. 2014, 134, 2251–2257. [Google Scholar] [CrossRef] [PubMed]
- Scott, G.A.; Laughlin, T.S.; Rothberg, P.G. Mutations of the TERT promoter are common in basal cell carcinoma and squamous cell carcinoma. Mod. Pathol. 2014, 27, 516–523. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.Y.; Liao, S.L.; Hong, J.B.; Chu, C.Y.; Sheen, Y.S.; Jhuang, J.Y.; Tsai, J.H.; Liau, J.Y. TERT promoter mutations in periocular carcinomas: Implications of ultraviolet light in pathogenesis. Br. J. Ophthalmol. 2016, 100, 274–277. [Google Scholar] [CrossRef] [PubMed]
- Macerola, E.; Loggini, B.; Giannini, R.; Garavello, G.; Giordano, M.; Proietti, A.; Niccoli, C.; Basolo, F.; Fontanini, G. Coexistence of TERT promoter and BRAF mutations in cutaneous melanoma is associated with more clinicopathological features of aggressiveness. Virchows Arch. 2015, 467, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Nagore, E.; Heidenreich, B.; Rachakonda, S.; Garcia-Casado, Z.; Requena, C.; Soriano, V.; Frank, C.; Traves, V.; Quecedo, E.; Sanjuan-Gimenez, J.; et al. TERT promoter mutations in melanoma survival. Int. J. Cancer 2016, 139, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Liau, J.Y.; Tsai, J.H.; Jeng, Y.M.; Chu, C.Y.; Kuo, K.T.; Liang, C.W. TERT promoter mutation is uncommon in acral lentiginous melanoma. J. Cutan. Pathol. 2014, 41, 504–508. [Google Scholar] [CrossRef] [PubMed]
- Vazquez, V.d.L.; Vicente, A.L.; Carloni, A.; Berardinelli, G.; Soares, P.; Scapulatempo, C.; Martinho, O.; Reis, R.M. Molecular profiling, including TERT promoter mutations, of acral lentiginous melanomas. Melanoma Res. 2016, 26, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Kong, Y.; Chi, Z.; Sheng, X.; Cui, C.; Wang, X.; Mao, L.; Tang, B.; Li, S.; Lian, B.; et al. MAPK pathway and tert promoter gene mutation pattern and its prognostic value in melanoma patients: A retrospective study of 2793 cases. Clin. Cancer Res. 2017, 23, 6120–6127. [Google Scholar] [CrossRef] [PubMed]
- Roh, M.R.; Park, K.H.; Chung, K.Y.; Shin, S.J.; Rha, S.Y.; Tsao, H. Telomerase reverse transcriptase (TERT) promoter mutations in Korean melanoma patients. Am. J. Cancer Res. 2017, 7, 134–138. [Google Scholar] [PubMed]
- Yang, S.; Leone, D.; Frydenlund, N.; Hoang, M.; Deng, A.; Hernandez-Perez, M.; Biswas, A.; Singh, R.; Yaar, R.; Mahalingam, M. Frequency of telomerase reverse transcripter promoter mutations in desmoplastic melanoma subtypes: Analyses of 76 cases. Melanoma Res. 2016, 26, 361–366. [Google Scholar] [CrossRef] [PubMed]
- Shain, A.H.; Garrido, M.; Botton, T.; Talevich, E.; Yeh, I.; Sanborn, J.Z.; Chung, J.; Wang, N.J.; Kakavand, H.; Mann, G.J.; et al. Exome sequencing of desmoplastic melanoma identifies recurrent NFKBIE promoter mutations and diverse activating mutations in the MAPK pathway. Nat. Genet. 2015, 47, 1194–1199. [Google Scholar] [CrossRef] [PubMed]
- Egberts, F.; Kruger, S.; Behrens, H.M.; Bergner, I.; Papaspyrou, G.; Werner, J.A.; Alkatout, I.; Haag, J.; Hauschild, A.; Rocken, C. Melanomas of unknown primary frequently harbor TERT-promoter mutations. Melanoma Res. 2014, 24, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Barnhill, R.L.; Dummer, R.; Dalton, J.; Wu, J.; Pappo, A.; Bahrami, A. TERT Promoter Mutations Are Predictive of Aggressive Clinical Behavior in Patients with Spitzoid Melanocytic Neoplasms. Sci. Rep. 2015, 5, 11200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egberts, F.; Bohne, A.S.; Kruger, S.; Hedderich, J.; Rompel, R.; Haag, J.; Rocken, C.; Hauschild, A. Varying Mutational Alterations in Multiple Primary Melanomas. J. Mol. Diagn. 2016, 18, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Ekedahl, H.; Lauss, M.; Olsson, H.; Griewank, K.G.; Schadendorf, D.; Ingvar, C.; Jonsson, G. High TERT promoter mutation frequency in non-acral cutaneous metastatic melanoma. Pigment Cell Melanoma Res. 2016, 29, 598–600. [Google Scholar] [CrossRef] [PubMed]
- Nagore, E.; Heidenreich, B.; Requena, C.; Garcia-Casado, Z.; Martorell-Calatayud, A.; Pont-Sanjuan, V.; Jimenez-Sanchez, A.I.; Kumar, R. TERT promoter mutations associate with fast-growing melanoma. Pigment Cell Melanoma Res. 2016, 29, 236–238. [Google Scholar] [CrossRef] [PubMed]
- Hayward, N.K.; Wilmott, J.S.; Waddell, N.; Johansson, P.A.; Field, M.A.; Nones, K.; Patch, A.M.; Kakavand, H.; Alexandrov, L.B.; Burke, H.; et al. Whole-genome landscapes of major melanoma subtypes. Nature 2017, 545, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Ofner, R.; Ritter, C.; Heidenreich, B.; Kumar, R.; Ugurel, S.; Schrama, D.; Becker, J.C. Distribution of TERT promoter mutations in primary and metastatic melanomas in Austrian patients. J. Cancer Res. Clin. Oncol. 2017, 143, 613–617. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.W.; Hodis, E.; Xu, M.J.; Kryukov, G.V.; Chin, L.; Garraway, L.A. Highly recurrent TERT promoter mutations in human melanoma. Science 2013, 339, 957–959. [Google Scholar] [CrossRef] [PubMed]
- Horn, S.; Figl, A.; Rachakonda, P.S.; Fischer, C.; Sucker, A.; Gast, A.; Kadel, S.; Moll, I.; Nagore, E.; Hemminki, K.; et al. TERT promoter mutations in familial and sporadic melanoma. Science 2013, 339, 959–961. [Google Scholar] [CrossRef] [PubMed]
- Griewank, K.G.; Schilling, B.; Murali, R.; Bielefeld, N.; Schwamborn, M.; Sucker, A.; Zimmer, L.; Hillen, U.; Schaller, J.; Brenn, T.; et al. TERT promoter mutations are frequent in atypical fibroxanthomas and pleomorphic dermal sarcomas. Mod. Pathol. 2014, 27, 502–508. [Google Scholar] [CrossRef] [PubMed]
- Xie, H.; Liu, T.; Wang, N.; Bjornhagen, V.; Hoog, A.; Larsson, C.; Lui, W.O.; Xu, D. TERT promoter mutations and gene amplification: Promoting TERT expression in Merkel cell carcinoma. Oncotarget 2014, 5, 10048–10057. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.J.; Yang, S.; Succaria, F.; Bhawan, J. Telomerase (TERT) promoter mutations are infrequent in Merkel cell carcinomas. J. Am. Acad. Dermatol. 2015, 73, 864–865. [Google Scholar] [CrossRef] [PubMed]
- Koelsche, C.; Renner, M.; Hartmann, W.; Brandt, R.; Lehner, B.; Waldburger, N.; Alldinger, I.; Schmitt, T.; Egerer, G.; Penzel, R.; et al. TERT promoter hotspot mutations are recurrent in myxoid liposarcomas but rare in other soft tissue sarcoma entities. J. Exp. Clin. Cancer Res. 2014, 33, 33. [Google Scholar] [CrossRef] [PubMed]
- Bahrami, A.; Lee, S.; Schaefer, I.M.; Boland, J.M.; Patton, K.T.; Pounds, S.; Fletcher, C.D. TERT promoter mutations and prognosis in solitary fibrous tumor. Mod. Pathol. 2016, 29, 1511–1522. [Google Scholar] [CrossRef] [PubMed]
- Dubbink, H.J.; Bakels, H.; Post, E.; Zwarthoff, E.C.; Verdijk, R.M. TERT promoter mutations and BRAF mutations are rare in sporadic, and TERT promoter mutations are absent in NF1-related malignant peripheral nerve sheath tumors. J. Neuro-Oncol. 2014, 120, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Campanella, N.C.; Celestino, R.; Pestana, A.; Scapulatempo-Neto, C.; de Oliveira, A.T.; Brito, M.J.; Gouveia, A.; Lopes, J.M.; Guimaraes, D.P.; Soares, P.; et al. Low frequency of TERT promoter mutations in gastrointestinal stromal tumors (GISTs). Eur. J. Hum. Genet. 2015, 23, 877–879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosen, I.; Rachakonda, P.S.; Heidenreich, B.; Sitaram, R.T.; Ljungberg, B.; Roos, G.; Hemminki, K.; Kumar, R. TERT promoter mutations in clear cell renal cell carcinoma. Int. J. Cancer 2015, 136, 2448–2452. [Google Scholar] [CrossRef] [PubMed]
- Casuscelli, J.; Becerra, M.F.; Manley, B.J.; Zabor, E.C.; Reznik, E.; Redzematovic, A.; Arcila, M.E.; Tennenbaum, D.M.; Ghanaat, M.; Kashan, M.; et al. Characterization and Impact of TERT Promoter Region Mutations on Clinical Outcome in Renal Cell Carcinoma. Eur. Urol. Focus 2017. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Liu, T.; Liu, L.; Liu, J.; Liu, C.; Wang, C.; Ge, N.; Ren, H.; Yan, K.; Hu, S.; et al. TERT promoter mutations in renal cell carcinomas and upper tract urothelial carcinomas. Oncotarget 2014, 5, 1829–1836. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Huang, P.; Li, C.; Huang, Y.; Li, X.; Wang, Y.; Chen, C.; Lv, Z.; Tang, A.; Sun, X.; et al. Telomerase reverse transcriptase gene promoter mutations help discern the origin of urogenital tumors: A genomic and molecular study. Eur. Urol. 2014, 65, 274–277. [Google Scholar] [CrossRef] [PubMed]
- Allory, Y.; Beukers, W.; Sagrera, A.; Flandez, M.; Marques, M.; Marquez, M.; van der Keur, K.A.; Dyrskjot, L.; Lurkin, I.; Vermeij, M.; et al. Telomerase reverse transcriptase promoter mutations in bladder cancer: High frequency across stages, detection in urine, and lack of association with outcome. Eur. Urol. 2014, 65, 360–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hurst, C.D.; Platt, F.M.; Knowles, M.A. Comprehensive mutation analysis of the TERT promoter in bladder cancer and detection of mutations in voided urine. Eur. Urol. 2014, 65, 367–369. [Google Scholar] [CrossRef] [PubMed]
- Rachakonda, P.S.; Hosen, I.; de Verdier, P.J.; Fallah, M.; Heidenreich, B.; Ryk, C.; Wiklund, N.P.; Steineck, G.; Schadendorf, D.; Hemminki, K.; et al. TERT promoter mutations in bladder cancer affect patient survival and disease recurrence through modification by a common polymorphism. Proc. Natl. Acad. Sci. USA 2013, 110, 17426–17431. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Liu, T.; Liu, C.; Meng, Y.; Yuan, X.; Liu, L.; Ge, N.; Liu, J.; Wang, C.; Ren, H. TERT promoter mutations and TERT mRNA but not FGFR3 mutations are urinary biomarkers in Han Chinese patients with urothelial bladder cancer. Oncologist 2015, 20, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Descotes, F.; Kara, N.; Decaussin-Petrucci, M.; Piaton, E.; Geiguer, F.; Rodriguez-Lafrasse, C.; Terrier, J.E.; Lopez, J.; Ruffion, A. Non-invasive prediction of recurrence in bladder cancer by detecting somatic TERT promoter mutations in urine. Br. J. Cancer 2017, 117, 583–587. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.T.; Liu, C.; Wang, K.; Liu, L.; Liu, T.T.; Ge, N.; Kong, F.; Yang, L.; Bjorkholm, M.; Fan, Y.D.; et al. The genetic difference between Western and Chinese urothelial cell carcinomas: Infrequent FGFR3 mutation in Han Chinese patients. Oncotarget 2016, 7, 25826–25835. [Google Scholar] [CrossRef] [PubMed]
- Pietzak, E.J.; Bagrodia, A.; Cha, E.K.; Drill, E.N.; Iyer, G.; Isharwal, S.; Ostrovnaya, I.; Baez, P.; Li, Q.; Berger, M.F.; et al. Next-generation sequencing of nonmuscle invasive bladder cancer reveals potential biomarkers and rational therapeutic targets. Eur. Urol. 2017, 72, 952–959. [Google Scholar] [CrossRef] [PubMed]
- Kinde, I.; Munari, E.; Faraj, S.F.; Hruban, R.H.; Schoenberg, M.; Bivalacqua, T.; Allaf, M.; Springer, S.; Wang, Y.; Diaz, L.A., Jr.; et al. TERT promoter mutations occur early in urothelial neoplasia and are biomarkers of early disease and disease recurrence in urine. Cancer Res. 2013, 73, 7162–7167. [Google Scholar] [CrossRef] [PubMed]
- Vail, E.; Zheng, X.; Zhou, M.; Yang, X.; Fallon, J.T.; Epstein, J.I.; Zhong, M. Telomerase reverse transcriptase promoter mutations in glandular lesions of the urinary bladder. Ann. Diagn. Pathol. 2015, 19, 301–305. [Google Scholar] [CrossRef] [PubMed]
- Brown, N.A.; Lew, M.; Weigelin, H.C.; Weizer, A.Z.; Montgomery, J.S.; Betz, B.L.; Mehra, R. Comparative study of TERT promoter mutation status within spatially, temporally and morphologically distinct components of urothelial carcinoma. Histopathology 2018, 72, 354–356. [Google Scholar] [CrossRef] [PubMed]
- Cowan, M.; Springer, S.; Nguyen, D.; Taheri, D.; Guner, G.; Rodriguez, M.A.; Wang, Y.; Kinde, I.; VandenBussche, C.J.; Olson, M.T.; et al. High prevalence of TERT promoter mutations in primary squamous cell carcinoma of the urinary bladder. Mod. Pathol. 2016, 29, 511–515. [Google Scholar] [CrossRef] [PubMed]
- Cowan, M.L.; Springer, S.; Nguyen, D.; Taheri, D.; Guner, G.; Mendoza Rodriguez, M.A.; Wang, Y.; Kinde, I.; Del Carmen Rodriguez Pena, M.; VandenBussche, C.J.; et al. Detection of TERT promoter mutations in primary adenocarcinoma of the urinary bladder. Hum. Pathol. 2016, 53, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez Pena, M.D.C.; Tregnago, A.C.; Eich, M.L.; Springer, S.; Wang, Y.; Taheri, D.; Ertoy, D.; Fujita, K.; Bezerra, S.M.; Cunha, I.W.; et al. Spectrum of genetic mutations in de novo PUNLMP of the urinary bladder. Virchows Arch. 2017, 471, 761–767. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Liu, T.T.; Ge, N.; Liu, L.; Yuan, X.T.; Liu, J.K.; Kong, F.; Wang, C.; Ren, H.B.; Yan, K.Q.; et al. TERT promoter mutations are associated with distant metastases in upper tract urothelial carcinomas and serve as urinary biomarkers detected by a sensitive castPCR. Oncotarget 2014, 5, 12428–12439. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Lopez-Beltran, A.; Osunkoya, A.O.; Wang, M.; Zhang, S.; Davidson, D.D.; Emerson, R.E.; Williamson, S.R.; Tan, P.H.; Kaimakliotis, H.Z.; et al. TERT promoter mutation status in sarcomatoid urothelial carcinomas of the upper urinary tract. Future Oncol. 2017, 13, 705–714. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.; Taheri, D.; Springer, S.; Cowan, M.; Guner, G.; Mendoza Rodriguez, M.A.; Wang, Y.; Kinde, I.; VandenBussche, C.J.; Olson, M.T.; et al. High prevalence of TERT promoter mutations in micropapillary urothelial carcinoma. Virchows Arch. 2016, 469, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Kurtis, B.; Zhuge, J.; Ojaimi, C.; Ye, F.; Cai, D.; Zhang, D.; Fallon, J.T.; Zhong, M. Recurrent TERT promoter mutations in urothelial carcinoma and potential clinical applications. Ann. Diagn. Pathol. 2016, 21, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Priemer, D.S.; Wang, M.; Zhang, S.; Lopez-Beltran, A.; Kouba, E.; Montironi, R.; Davidson, D.D.; MacLennan, G.T.; Wang, L.; Osunkoya, A.O.; et al. Small-cell carcinomas of the urinary bladder and prostate: TERT promoter mutation status differentiates sites of malignancy and provides evidence of common clonality between small-cell carcinoma of the urinary bladder and urothelial carcinoma. Eur. Urol. Focus 2017. [Google Scholar] [CrossRef] [PubMed]
- Lack, E.E. Tumours of the adrenal glands and extraadrenal paraganglia; American Registry of Pathology in collaboration with the Armed Forces Institute of Pathology: Washington, DC, USA, 2007. [Google Scholar]
- Heidenreich, B.; Nagore, E.; Rachakonda, P.S.; Garcia-Casado, Z.; Requena, C.; Traves, V.; Becker, J.; Soufir, N.; Hemminki, K.; Kumar, R. Telomerase reverse transcriptase promoter mutations in primary cutaneous melanoma. Nat. Commun. 2014, 5, 3401. [Google Scholar] [CrossRef] [PubMed]
- Hosen, I.; Rachakonda, P.S.; Heidenreich, B.; de Verdier, P.J.; Ryk, C.; Steineck, G.; Hemminki, K.; Kumar, R. Mutations in TERT promoter and FGFR3 and telomere length in bladder cancer. Int. J. Cancer 2015, 137, 1621–1629. [Google Scholar] [CrossRef] [PubMed]
- Piscuoglio, S.; Ng, C.K.Y.; Murray, M.; Burke, K.A.; Edelweiss, M.; Geyer, F.C.; Macedo, G.S.; Inagaki, A.; Papanastasiou, A.D.; Martelotto, L.G.; et al. Massively parallel sequencing of phyllodes tumours of the breast reveals actionable mutations, and TERT promoter hotspot mutations and TERT gene amplification as likely drivers of progression. J. Pathol. 2016, 238, 508–518. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Wang, Y.; Kratz, J.; Brat, D.J.; Robitaille, Y.; Moghrabi, A.; Perlman, E.J.; Dang, C.V.; Burger, P.C.; Eberhart, C.G. hTERT gene amplification and increased mRNA expression in central nervous system embryonal tumors. Am. J. Pathol. 2003, 162, 1763–1769. [Google Scholar] [CrossRef]
- Palmqvist, R.; Zhang, A.; Xu, D.; Golovleva, I.; Norrback, K.F.; Gruber, A.; Oberg, A.; Stenling, R.; Roos, G. hTERT gene copy number is not associated with hTERT RNA expression or telomerase activity in colorectal cancer. Int. J. Cancer 2005, 116, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Takuma, Y.; Nouso, K.; Kobayashi, Y.; Nakamura, S.; Tanaka, H.; Matsumoto, E.; Fujikawa, T.; Suzuki, M.; Hanafusa, T.; Shiratori, Y. Telomerase reverse transcriptase gene amplification in hepatocellular carcinoma. J. Gastroenterol. Hepatol. 2004, 19, 1300–1304. [Google Scholar] [CrossRef] [PubMed]
- Visnovsky, J.; Kudela, E.; Farkasova, A.; Balharek, T.; Krkoska, M.; Danko, J. Amplification of TERT and TERC genes in cervical intraepithelial neoplasia and cervical cancer. Neuroendocrinol. Lett. 2014, 35, 518–522. [Google Scholar] [PubMed]
- Freier, K.; Pungs, S.; Flechtenmacher, C.; Bosch, F.X.; Lichter, P.; Joos, S.; Hofele, C. Frequent high telomerase reverse transcriptase expression in primary oral squamous cell carcinoma. J. Oral. Pathol. Med. 2007, 36, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.Q.; Cutz, J.C.; Liu, N.; Lau, D.; Shepherd, F.A.; Squire, J.A.; Tsao, M.S. Amplification of telomerase (hTERT) gene is a poor prognostic marker in non-small-cell lung cancer. Br. J. Cancer 2006, 94, 1452–1459. [Google Scholar] [CrossRef] [PubMed]
- Penzo, M.; Ludovini, V.; Trere, D.; Siggillino, A.; Vannucci, J.; Bellezza, G.; Crino, L.; Montanaro, L. Dyskerin and TERC expression may condition survival in lung cancer patients. Oncotarget 2015, 6, 21755–21760. [Google Scholar] [CrossRef] [PubMed]
- Diaz, A.; Puig-Butille, J.A.; Munoz, C.; Costa, D.; Diez, A.; Garcia-Herrera, A.; Carrera, C.; Badenas, C.; Sole, F.; Malvehy, J.; Puig, S.; Alos, L. TERT gene amplification is associated with poor outcome in acral lentiginous melanoma. J. Am. Acad. Dermatol. 2014, 71, 839–841. [Google Scholar] [CrossRef] [PubMed]
- Puig-Butillé, J.A.; Badenas, C.; Ogbah, Z.; Carrera, C.; Aguilera, P.; Malvehy, J.; Puig, S. Genetic alterations in RAS-regulated pathway in acral lentiginous melanoma. Exp. Dermatol. 2013, 22, 148–150. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Opresko, P.; Pappo, A.; Kirkwood, J.M.; Bahrami, A. Association of TERT promoter mutations with telomerase expression in melanoma. Pigment Cell Melanoma Res. 2016, 29, 391–393. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, Y.; Chochi, Y.; Matsuyama, H.; Eguchi, S.; Kawauchi, S.; Furuya, T.; Oga, A.; Kang, J.J.; Naito, K.; Sasaki, K. Gain of 5p15.33 is associated with progression of bladder cancer. Oncology 2007, 72, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Sugita, M.; Tanaka, N.; Davidson, S.; Sekiya, S.; Varella-Garcia, M.; West, J.; Drabkin, H.A.; Gemmill, R.M. Molecular definition of a small amplification domain within 3q26 in tumors of cervix, ovary, and lung. Cancer Genet. Cytogenet. 2000, 117, 9–18. [Google Scholar] [CrossRef]
- Liu, H.Q.; Liu, S.L.; Wang, H.; Xie, X.Y.; Chen, X.L.; Zhang, X.M.; Zhang, Y.C. Genomic amplification of the human telomerase gene (hTERC) associated with human papillomavirus is related to the progression of uterine cervical dysplasia to invasive cancer. Diagn. Pathol. 2012, 7, 147. [Google Scholar] [CrossRef] [PubMed]
- Andersson, S.; Sowjanya, P.; Wangsa, D.; Hjerpe, A.; Johansson, B.; Auer, G.; Gravitt, P.E.; Larsson, C.; Wallin, K.L.; Ried, T.; et al. Detection of genomic amplification of the human telomerase gene TERC, a potential marker for triage of women with HPV-positive, abnormal Pap smears. Am. J. Pathol. 2009, 175, 1831–1847. [Google Scholar] [CrossRef] [PubMed]
- Soder, A.I.; Hoare, S.F.; Muir, S.; Going, J.J.; Parkinson, E.K.; Keith, W.N. Amplification, increased dosage and in situ expression of the telomerase RNA gene in human cancer. Oncogene 1997, 14, 1013–1021. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.-C.; Shyong, W.-Y.; Chang, M.-S.; Chen, Y.-J.; Lin, C.-H.; Huang, Z.-D.; Hsu, M.-T.; Chen, M.-L. Frequent gain of copy number on the long arm of chromosome 3 in human cervical adenocarcinoma. Cancer Genet. Cytogenet. 2001, 131, 48–53. [Google Scholar] [CrossRef]
- Andersson, S.; Wallin, K.L.; Hellstrom, A.C.; Morrison, L.E.; Hjerpe, A.; Auer, G.; Ried, T.; Larsson, C.; Heselmeyer-Haddad, K. Frequent gain of the human telomerase gene TERC at 3q26 in cervical adenocarcinomas. Br. J. Cancer 2006, 95, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Jin, G.; Xu, L.; Shu, Y.; Tian, T.; Liang, J.; Xu, Y.; Wang, F.; Chen, J.; Dai, J.; Hu, Z.; et al. Common genetic variants on 5p15.33 contribute to risk of lung adenocarcinoma in a Chinese population. Carcinogenesis 2009, 30, 987–990. [Google Scholar] [CrossRef] [PubMed]
- Landi, M.T.; Chatterjee, N.; Yu, K.; Goldin, L.R.; Goldstein, A.M.; Rotunno, M.; Mirabello, L.; Jacobs, K.; Wheeler, W.; Yeager, M.; et al. A genome-wide association study of lung cancer identifies a region of chromosome 5p15 associated with risk for adenocarcinoma. Am. J. Hum. Genet. 2009, 85, 679–691. [Google Scholar] [CrossRef] [PubMed]
- Shete, S.; Hosking, F.J.; Robertson, L.B.; Dobbins, S.E.; Sanson, M.; Malmer, B.; Simon, M.; Marie, Y.; Boisselier, B.; Delattre, J.Y.; et al. Genome-wide association study identifies five susceptibility loci for glioma. Nat. Genet. 2009, 41, 899–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Broderick, P.; Matakidou, A.; Eisen, T.; Houlston, R.S. Role of 5p15.33 (TERT-CLPTM1L), 6p21.33 and 15q25.1 (CHRNA5-CHRNA3) variation and lung cancer risk in never-smokers. Carcinogenesis 2010, 31, 234–238. [Google Scholar] [CrossRef] [PubMed]
- Hsiung, C.A.; Lan, Q.; Hong, Y.C.; Chen, C.J.; Hosgood, H.D.; Chang, I.S.; Chatterjee, N.; Brennan, P.; Wu, C.; Zheng, W.; et al. The 5p15.33 locus is associated with risk of lung adenocarcinoma in never-smoking females in Asia. PLoS Genet. 2010, 6, e1001051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miki, D.; Kubo, M.; Takahashi, A.; Yoon, K.A.; Kim, J.; Lee, G.K.; Zo, J.I.; Lee, J.S.; Hosono, N.; Morizono, T.; et al. Variation in TP63 is associated with lung adenocarcinoma susceptibility in Japanese and Korean populations. Nat. Genet. 2010, 42, 893–896. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Chen, Y.; Zhao, Y.; Fan, W.; Zhou, K.; Liu, Y.; Zhou, L.; Mao, Y.; Wei, Q.; Xu, J.; et al. Association of sequence variants on chromosomes 20, 11, and 5 (20q13.33, 11q23.3, and 5p15.33) with glioma susceptibility in a Chinese population. Am. J. Epidemiol. 2011, 173, 915–922. [Google Scholar] [CrossRef] [PubMed]
- Bae, E.Y.; Lee, S.Y.; Kang, B.K.; Lee, E.J.; Choi, Y.Y.; Kang, H.G.; Choi, J.E.; Jeon, H.S.; Lee, W.K.; Kam, S. Replication of results of genome-wide association studies on lung cancer susceptibility loci in a Korean population. Respirology 2012, 17, 699–706. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.F.; Cai, S.; Chen, Q.G.; Ni, Z.H.; Tang, J.H.; Xu, D.W.; Wang, X.B. Multiple variants of TERT and CLPTM1L constitute risk factors for lung adenocarcinoma. Genet. Mol. Res. 2012, 11, 370–378. [Google Scholar] [CrossRef] [PubMed]
- Di Stefano, A.L.; Enciso-Mora, V.; Marie, Y.; Desestret, V.; Labussière, M.; Boisselier, B.; Mokhtari, K.; Idbaih, A.; Hoang-Xuan, K.; Delattre, J.-Y. Association between glioma susceptibility loci and tumour pathology defines specific molecular etiologies. Neuro-Oncology 2012, 15, 542–547. [Google Scholar] [CrossRef] [PubMed]
- Kinnersley, B.; Migliorini, G.; Broderick, P.; Whiffin, N.; Dobbins, S.E.; Casey, G.; Hopper, J.; Sieber, O.; Lipton, L.; Kerr, D.J.; et al. The TERT variant rs2736100 is associated with colorectal cancer risk. Br. J. Cancer 2012, 107, 1001–1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Wu, J.; Hu, L.; Ding, C.; Kan, Y.; Shen, Y.; Chen, X.; Shen, H.; Guo, X.; Hu, Z. Common genetic variants in TERT contribute to risk of cervical cancer in a Chinese population. Mol. Carcinog. 2012, 51 (Suppl. 1), E118–E122. [Google Scholar] [CrossRef] [PubMed]
- Lan, Q.; Cawthon, R.; Gao, Y.; Hu, W.; Hosgood, H.D., 3rd; Barone-Adesi, F.; Ji, B.T.; Bassig, B.; Chow, W.H.; Shu, X.; et al. Longer telomere length in peripheral white blood cells is associated with risk of lung cancer and the rs2736100 (CLPTM1L-TERT) polymorphism in a prospective cohort study among women in China. PLoS ONE 2013, 8, e59230. [Google Scholar] [CrossRef] [PubMed]
- Myneni, A.A.; Chang, S.C.; Niu, R.; Liu, L.; Ochs-Balcom, H.M.; Li, Y.; Zhang, C.; Zhao, B.; Shi, J.; Han, X.; et al. Genetic polymorphisms of TERT and CLPTM1L and risk of lung cancer—A case-control study in a Chinese population. Lung Cancer 2013, 80, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Safaeian, M.; Rajaraman, P.; Hartge, P.; Yeager, M.; Linet, M.; Butler, M.A.; Ruder, A.M.; Purdue, M.P.; Hsing, A.; Beane-Freeman, L.; et al. Joint effects between five identified risk variants, allergy, and autoimmune conditions on glioma risk. Cancer Causes Control 2013, 24, 1885–1891. [Google Scholar] [CrossRef] [PubMed]
- Sheng, X.; Tong, N.; Tao, G.; Luo, D.; Wang, M.; Fang, Y.; Li, J.; Xu, M.; Zhang, Z.; Wu, D. TERT polymorphisms modify the risk of acute lymphoblastic leukemia in Chinese children. Carcinogenesis 2013, 34, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Jager, R.; Harutyunyan, A.S.; Rumi, E.; Pietra, D.; Berg, T.; Olcaydu, D.; Houlston, R.S.; Cazzola, M.; Kralovics, R. Common germline variation at the TERT locus contributes to familial clustering of myeloproliferative neoplasms. Am. J. Hematol. 2014, 89, 1107–1110. [Google Scholar] [CrossRef] [PubMed]
- Oddsson, A.; Kristinsson, S.Y.; Helgason, H.; Gudbjartsson, D.F.; Masson, G.; Sigurdsson, A.; Jonasdottir, A.; Jonasdottir, A.; Steingrimsdottir, H.; Vidarsson, B.; et al. The germline sequence variant rs2736100_C in TERT associates with myeloproliferative neoplasms. Leukemia 2014, 28, 1371–1374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, M.M.; Zhang, Y.; Shen, L.; Ren, Y.W.; Li, X.L.; Yin, Z.H.; Zhou, B.S. Genetic variations in TERT-CLPTM1L genes and risk of lung cancer in a Chinese population. Asian Pac. J. Cancer Prev. 2014, 15, 2809–2813. [Google Scholar] [CrossRef] [PubMed]
- Campa, D.; Rizzato, C.; Stolzenberg-Solomon, R.; Pacetti, P.; Vodicka, P.; Cleary, S.P.; Capurso, G.; Bueno-de-Mesquita, H.B.; Werner, J.; Gazouli, M.; et al. TERT gene harbors multiple variants associated with pancreatic cancer susceptibility. Int. J. Cancer 2015, 137, 2175–2183. [Google Scholar] [CrossRef] [PubMed]
- Krahling, T.; Balassa, K.; Kiss, K.P.; Bors, A.; Batai, A.; Halm, G.; Egyed, M.; Fekete, S.; Remenyi, P.; Masszi, T. Co-occurrence of myeloproliferative neoplasms and solid tumors is attributed to a synergism between cytoreductive therapy and the common TERT polymorphism rs2736100. Cancer Epidemiol. Prev. Biomark. 2015. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.G.; Ma, L.; Cen, Q.H.; Huang, J.S.; Zhang, J.X.; Zhang, J.J. Association of genetic polymorphisms in TERT-CLPTM1L with lung cancer in a Chinese population. Genet. Mol. Res. 2015, 14, 4469–4476. [Google Scholar] [CrossRef] [PubMed]
- Tapper, W.; Jones, A.V.; Kralovics, R.; Harutyunyan, A.S.; Zoi, K.; Leung, W.; Godfrey, A.L.; Guglielmelli, P.; Callaway, A.; Ward, D.; et al. Genetic variation at MECOM, TERT, JAK2 and HBS1L-MYB predisposes to myeloproliferative neoplasms. Nat. Commun. 2015, 6, 6691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, R.; Cao, L.; Pu, H.; Wang, H.; Zheng, Y.; Niu, X.; Weng, X.; Zhang, H.; Favus, M.; Zhang, L.; et al. TERT Polymorphism rs2736100-C is associated with EGFR mutation-positive non-small cell lung cancer. Clin. Cancer Res. 2015, 21, 5173–5180. [Google Scholar] [CrossRef] [PubMed]
- Bayram, S.; Ulger, Y.; Sumbul, A.T.; Kaya, B.Y.; Genc, A.; Rencuzogullari, E.; Dadas, E. Polymorphisms in human telomerase reverse transcriptase (hTERT) gene and susceptibility to gastric cancer in a Turkish population: Hospital-based case-control study. Gene 2016, 585, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Carkic, J.; Nikolic, N.; Radojevic-Skodric, S.; Kuzmanovic-Pficer, J.; Brajovic, G.; Antunovic, M.; Milasin, J.; Popovic, B. The role of TERT-CLPTM1L SNPs, hTERT expression and telomere length in the pathogenesis of oral squamous cell carcinoma. J. Oral. Sci 2016, 58, 449–458. [Google Scholar] [CrossRef] [PubMed]
- Dahlstrom, J.; Liu, T.; Yuan, X.; Saft, L.; Ghaderi, M.; Wei, Y.B.; Lavebratt, C.; Li, P.; Zheng, C.; Bjorkholm, M.; et al. TERT rs2736100 genotypes are associated with differential risk of myeloproliferative neoplasms in Swedish and Chinese male patient populations. Ann. Hematol. 2016, 95, 1825–1832. [Google Scholar] [CrossRef] [PubMed]
- Ge, M.; Shi, M.; An, C.; Yang, W.; Nie, X.; Zhang, J.; Lv, Z.; Li, J.; Zhou, L.; Du, Z.; et al. Functional evaluation of TERT-CLPTM1L genetic variants associated with susceptibility of papillary thyroid carcinoma. Sci. Rep. 2016, 6, 26037. [Google Scholar] [CrossRef] [PubMed]
- Trifa, A.P.; Bănescu, C.; Tevet, M.; Bojan, A.; Dima, D.; Urian, L.; Török-Vistai, T.; Popov, V.M.; Zdrenghea, M.; Petrov, L. TERT rs2736100 A> C SNP and JAK2 46/1 haplotype significantly contribute to the occurrence of JAK2V617F and CALR mutated myeloproliferative neoplasms—A multicentric study on 529 patients. Br. J. Haematol. 2016, 174, 218–226. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.; Prescott, J.; Yuan, C.; Zhang, M.; Kraft, P.; Babic, A.; Morales-Oyarvide, V.; Qian, Z.R.; Buring, J.E.; Cochrane, B.B.; et al. Leucocyte telomere length, genetic variants at the TERT gene region and risk of pancreatic cancer. Gut 2017, 66, 1116–1122. [Google Scholar] [CrossRef] [PubMed]
- Rafnar, T.; Sulem, P.; Stacey, S.N.; Geller, F.; Gudmundsson, J.; Sigurdsson, A.; Jakobsdottir, M.; Helgadottir, H.; Thorlacius, S.; Aben, K.K. Sequence variants at the TERT-CLPTM1L locus associate with many cancer types. Nat. Genet. 2009, 41, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Thakur, A.; Liang, Y.; Zhang, S.; Wang, T.; Chen, T.; Meng, J.; Wang, L.; Wu, F.; Jin, T.; et al. Polymorphisms in the TERT gene are associated with lung cancer risk in the Chinese Han population. Eur. J. Cancer Prev. 2014, 23, 497–501. [Google Scholar] [CrossRef] [PubMed]
- Su, L.Y.; Li, X.L.; Shen, L.; Zhang, Y.; Zhao, M.M.; Yin, Z.H.; Su, H.Y.; Zhou, B.S. Polymorphisms of TERT and CLPTM1L and the risk of hepatocellular carcinoma in Chinese males. Asian Pac. J. Cancer Prev. 2014, 15, 8197–8201. [Google Scholar] [CrossRef] [PubMed]
- Jannuzzi, A.T.; Karaman, E.; Oztas, E.; Yanar, H.T.; Ozhan, G. Telomerase Reverse Transcriptase (TERT) Gene Variations and Susceptibility of Colorectal Cancer. Genet. Test. Mol. Biomark. 2015, 19, 692–697. [Google Scholar] [CrossRef] [PubMed]
- Yoo, S.S.; Do, S.K.; Choi, J.E.; Lee, S.Y.; Lee, J.; Cha, S.I.; Kim, C.H.; Park, J.Y. TERT Polymorphism rs2853669 Influences on Lung Cancer Risk in the Korean Population. J. Korean Med. Sci. 2015, 30, 1423–1428. [Google Scholar] [CrossRef] [PubMed]
- Oztas, E.; Kara, H.; Kara, Z.P.; Aydogan, M.U.; Uras, C.; Ozhan, G. Association Between Human Telomerase Reverse Transcriptase Gene Variations and Risk of Developing Breast Cancer. Genet. Test. Mol. Biomark. 2016, 20, 459–464. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, X.; Zhang, H.; Zhai, Y.; Wang, Z.; Li, P.; Yu, L.; Xia, X.; Zhang, Y.; Zeng, Y.; et al. Common variations in TERT-CLPTM1L locus are reproducibly associated with the risk of nasopharyngeal carcinoma in Chinese populations. Oncotarget 2016, 7, 759–770. [Google Scholar] [CrossRef] [PubMed]
- Egan, K.M.; Thompson, R.C.; Nabors, L.; Olson, J.J.; Brat, D.J.; LaRocca, R.V.; Brem, S.; Moots, P.L.; Madden, M.H.; Browning, J.E. Cancer susceptibility variants and the risk of adult glioma in a US case-control study. J. Neuro-Oncol. 2011, 104, 535–542. [Google Scholar] [CrossRef] [PubMed]
- Nan, H.; Qureshi, A.A.; Prescott, J.; de Vivo, I.; Han, J. Genetic variants in telomere-maintaining genes and skin cancer risk. Hum. Genet. 2011, 129, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Jin, T.B.; Wei, X.B.; He, S.M.; Liang, H.J.; Yang, H.X.; Cui, Y.; Chen, C.; Cai, L.B.; Gao, G.D. Selected polymorphisms of GSTP1 and TERT were associated with glioma risk in Han Chinese. Cancer Epidemiol. 2012, 36, 525–527. [Google Scholar] [CrossRef] [PubMed]
- Terry, K.L.; Tworoger, S.S.; Vitonis, A.F.; Wong, J.; Titus-Ernstoff, L.; De Vivo, I.; Cramer, D.W. Telomere length and genetic variation in telomere maintenance genes in relation to ovarian cancer risk. Cancer Epidemiol. Biomark. Prev. 2012, 21, 504–512. [Google Scholar] [CrossRef] [PubMed]
- Jin, T.B.; Zhang, J.Y.; Li, G.; Du, S.L.; Geng, T.T.; Gao, J.; Liu, Q.P.; Gao, G.D.; Kang, L.L.; Chen, C.; et al. RTEL1 and TERT polymorphisms are associated with astrocytoma risk in the Chinese Han population. Tumor Biol. 2013, 34, 3659–3666. [Google Scholar] [CrossRef] [PubMed]
- Kote-Jarai, Z.; Saunders, E.J.; Leongamornlert, D.A.; Tymrakiewicz, M.; Dadaev, T.; Jugurnauth-Little, S.; Ross-Adams, H.; Al Olama, A.A.; Benlloch, S.; Halim, S.; et al. Fine-mapping identifies multiple prostate cancer risk loci at 5p15, one of which associates with TERT expression. Hum. Mol. Genet. 2013, 22, 2520–2528. [Google Scholar] [CrossRef] [PubMed]
- Pellatt, A.J.; Wolff, R.K.; Torres-Mejia, G.; John, E.M.; Herrick, J.S.; Lundgreen, A.; Baumgartner, K.B.; Giuliano, A.R.; Hines, L.M.; Fejerman, L. Telomere length, telomere-related genes, and breast cancer risk: The breast cancer health disparities study. Genes Chromosomes Cancer 2013, 52, 595–609. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Li, C.; Yang, L.; Zhang, X.; Zhao, X.; Song, X.; Li, X.; Wang, J.; Qian, J.; Yang, Y.; et al. Significant association of 5p15.33 (TERT-CLPTM1L genes) with lung cancer in Chinese Han population. Exp. Lung Res. 2013, 39, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Park, S.L.; Fesinmeyer, M.D.; Timofeeva, M.; Caberto, C.P.; Kocarnik, J.M.; Han, Y.; Love, S.A.; Young, A.; Dumitrescu, L.; Lin, Y.; et al. Pleiotropic associations of risk variants identified for other cancers with lung cancer risk: The PAGE and TRICL consortia. J. Natl. Cancer Inst. 2014, 106. [Google Scholar] [CrossRef] [PubMed]
- Duan, X.; Cao, W.; Wang, L.; Liu, S.; Liu, Z.; Zhang, B.; Yang, H.; Feng, T.; Zhang, J.; Zhang, X.; et al. Genetic variants in TERT are associated with risk of gastric cancer in a Chinese Han population. Oncotarget 2016, 7, 82727–82732. [Google Scholar] [CrossRef] [PubMed]
- Savage, S.A.; Chanock, S.J.; Lissowska, J.; Brinton, L.A.; Richesson, D.; Peplonska, B.; Bardin-Mikolajczak, A.; Zatonski, W.; Szeszenia-Dabrowska, N.; Garcia-Closas, M. Genetic variation in five genes important in telomere biology and risk for breast cancer. Br. J. Cancer 2007, 97, 832–836. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Gammon, M.D.; Wu, H.C.; Terry, M.B.; Wang, Q.; Bradshaw, P.T.; Teitelbaum, S.L.; Neugut, A.I.; Santella, R.M. Multiple genetic variants in telomere pathway genes and breast cancer risk. Cancer Epidemiol. Biomark. Prev. 2010, 19, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Zhong, R.; Liu, L.; Zou, L.; Zhu, Y.; Chen, W.; Zhu, B.; Shen, N.; Rui, R.; Long, L.; Ke, J. Genetic variations in TERT-CLPTM1L locus are associated with risk of lung cancer in chinese population. Mol. Carcinog. 2013, 52 (Suppl. 1), 118–126. [Google Scholar] [CrossRef] [PubMed]
- Shadrina, A.S.; Boyarskikh, U.A.; Oskina, N.A.; Sinkina, T.V.; Lazarev, A.F.; Petrova, V.D.; Filipenko, M.L. TERT polymorphisms rs2853669 and rs7726159 influence on prostate cancer risk in Russian population. Tumor Biol. 2015, 36, 841–847. [Google Scholar] [CrossRef] [PubMed]
- Ko, E.; Seo, H.W.; Jung, E.S.; Kim, B.H.; Jung, G. The TERT promoter SNP rs2853669 decreases E2F1 transcription factor binding and increases mortality and recurrence risks in liver cancer. Oncotarget 2016, 7, 684–699. [Google Scholar] [CrossRef] [PubMed]
- Pellegrini, C.; Maturo, M.G.; Martorelli, C.; Suppa, M.; Antonini, A.; Kostaki, D.; Verna, L.; Landi, M.T.; Peris, K.; Fargnoli, M.C. Characterization of melanoma susceptibility genes in high-risk patients from Central Italy. Melanoma Res. 2017, 27, 258–267. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.E.; Kang, H.G.; Jang, J.S.; Choi, Y.Y.; Kim, M.J.; Kim, J.S.; Jeon, H.S.; Lee, W.K.; Cha, S.I.; Kim, C.H.; et al. Polymorphisms in telomere maintenance genes and risk of lung cancer. Cancer Epidemiol. Biomark. Prev. 2009, 18, 2773–2781. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, F.J.; Vizcaino, M.A.; Blakeley, J.; Heaphy, C.M. Frequent alternative lengthening of telomeres and ATRX loss in adult NF1-associated diffuse and high-grade astrocytomas. Acta Neuropathol. 2016, 132, 761–763. [Google Scholar] [CrossRef] [PubMed]
- Abedalthagafi, M.; Phillips, J.J.; Kim, G.E.; Mueller, S.; Haas-Kogen, D.A.; Marshall, R.E.; Croul, S.E.; Santi, M.R.; Cheng, J.; Zhou, S.M.; et al. The alternative lengthening of telomere phenotype is significantly associated with loss of ATRX expression in high-grade pediatric and adult astrocytomas: A multi-institutional study of 214 astrocytomas. Mod. Pathol. 2013, 26, 1425–1432. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.J.; Hakin-Smith, V.; Teo, M.; Xinarianos, G.E.; Jellinek, D.A.; Carroll, T.; McDowell, D.; MacFarlane, M.R.; Boet, R.; Baguley, B.C.; et al. Association of mutant TP53 with alternative lengthening of telomeres and favorable prognosis in glioma. Cancer Res. 2006, 66, 6473–6476. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.N.; Heaphy, C.M.; Wilde, R.F.; Orr, B.A.; Odia, Y.; Eberhart, C.G.; Meeker, A.K.; Rodriguez, F.J. Molecular and Morphologic Correlates of the Alternative Lengthening of Telomeres Phenotype in High-Grade Astrocytomas. Brain Pathol. 2013, 23, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Henson, J.D.; Hannay, J.A.; McCarthy, S.W.; Royds, J.A.; Yeager, T.R.; Robinson, R.A.; Wharton, S.B.; Jellinek, D.A.; Arbuckle, S.M.; Yoo, J.Y.; et al. A robust assay for alternative lengthening of telomeres in tumors shows the significance of alternative lengthening of telomeres in sarcomas and astrocytomas. Clin. Cancer Res. 2005, 11, 217–225. [Google Scholar] [PubMed]
- Mangerel, J.; Price, A.; Castelo-Branco, P.; Brzezinski, J.; Buczkowicz, P.; Rakopoulos, P.; Merino, D.; Baskin, B.; Wasserman, J.; Mistry, M.; et al. Alternative lengthening of telomeres is enriched in, and impacts survival of TP53 mutant pediatric malignant brain tumors. Acta Neuropathol. 2014, 128, 853–862. [Google Scholar] [CrossRef] [PubMed]
- McDonald, K.L.; McDonnell, J.; Muntoni, A.; Henson, J.D.; Hegi, M.E.; von Deimling, A.; Wheeler, H.R.; Cook, R.J.; Biggs, M.T.; Little, N.S.; et al. Presence of alternative lengthening of telomeres mechanism in patients with glioblastoma identifies a less aggressive tumor type with longer survival. J. Neuropathol. Exp. Neurol. 2010, 69, 729–736. [Google Scholar] [CrossRef] [PubMed]
- Fogli, A.; Demattei, M.V.; Corset, L.; Vaurs-Barriere, C.; Chautard, E.; Biau, J.; Kemeny, J.L.; Godfraind, C.; Pereira, B.; Khalil, T.; et al. Detection of the alternative lengthening of telomeres pathway in malignant gliomas for improved molecular diagnosis. J. Neuro-Oncol. 2017, 135, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Hakin-Smith, V.; Jellinek, D.A.; Levy, D.; Carroll, T.; Teo, M.; Timperley, W.R.; McKay, M.J.; Reddel, R.R.; Royds, J.A. Alternative lengthening of telomeres and survival in patients with glioblastoma multiforme. Lancet 2003, 361, 836–838. [Google Scholar] [CrossRef]
- Wood, L.D.; Heaphy, C.M.; Daniel, H.D.; Naini, B.V.; Lassman, C.R.; Arroyo, M.R.; Kamel, I.R.; Cosgrove, D.P.; Boitnott, J.K.; Meeker, A.K.; et al. Chromophobe hepatocellular carcinoma with abrupt anaplasia: A proposal for a new subtype of hepatocellular carcinoma with unique morphological and molecular features. Mod. Pathol. 2013, 26, 1586–1593. [Google Scholar] [CrossRef] [PubMed]
- Omori, Y.; Nakayama, F.; Li, D.; Kanemitsu, K.; Semba, S.; Ito, A.; Yokozaki, H. Alternative lengthening of telomeres frequently occurs in mismatch repair system-deficient gastric carcinoma. Cancer Sci. 2009, 100, 413–418. [Google Scholar] [CrossRef] [PubMed]
- Dogeas, E.; Karagkounis, G.; Heaphy, C.M.; Hirose, K.; Pawlik, T.M.; Wolfgang, C.L.; Meeker, A.; Hruban, R.H.; Cameron, J.L.; Choti, M.A. Alternative lengthening of telomeres predicts site of origin in neuroendocrine tumor liver metastases. J. Am. Coll. Surg. 2014, 218, 628–635. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Xu, D.; Sofiadis, A.; Hoog, A.; Vukojevic, V.; Backdahl, M.; Zedenius, J.; Larsson, C. Telomerase-dependent and independent telomere maintenance and its clinical implications in medullary thyroid carcinoma. J. Clin. Endocrinol. Metab. 2014, 99, E1571–E1579. [Google Scholar] [CrossRef] [PubMed]
- Else, T.; Giordano, T.J.; Hammer, G.D. Evaluation of telomere length maintenance mechanisms in adrenocortical carcinoma. J. Clin. Endocrinol. Metab. 2008, 93, 1442–1449. [Google Scholar] [CrossRef] [PubMed]
- Marinoni, I.; Kurrer, A.S.; Vassella, E.; Dettmer, M.; Rudolph, T.; Banz, V.; Hunger, F.; Pasquinelli, S.; Speel, E.J.; Perren, A. Loss of DAXX and ATRX are associated with chromosome instability and reduced survival of patients with pancreatic neuroendocrine tumors. Gastroenterology 2014, 146, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Lee, H.S.; Nam, K.H.; Choi, J.; Kim, W.H. Telomere length abnormalities and telomerase RNA component expression in gastroenteropancreatic neuroendocrine tumors. Anticancer Res. 2015, 35, 3501–3510. [Google Scholar] [PubMed]
- Singhi, A.D.; Liu, T.C.; Roncaioli, J.L.; Cao, D.; Zeh, H.J.; Zureikat, A.H.; Tsung, A.; Marsh, J.W.; Lee, K.K.; Hogg, M.E.; et al. Alternative Lengthening of Telomeres and Loss of DAXX/ATRX Expression Predicts Metastatic Disease and Poor Survival in Patients with Pancreatic Neuroendocrine Tumors. Clin. Cancer Res. 2017, 23, 600–609. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Brosnan-Cashman, J.A.; An, S.; Kim, S.J.; Song, K.B.; Kim, M.S.; Kim, M.J.; Hwang, D.W.; Meeker, A.K.; Yu, E.; et al. Alternative Lengthening of Telomeres in Primary Pancreatic Neuroendocrine Tumors Is Associated with Aggressive Clinical Behavior and Poor Survival. Clin. Cancer Res. 2017, 23, 1598–1606. [Google Scholar] [CrossRef] [PubMed]
- VandenBussche, C.J.; Allison, D.B.; Graham, M.K.; Charu, V.; Lennon, A.M.; Wolfgang, C.L.; Hruban, R.H.; Heaphy, C.M. Alternative lengthening of telomeres and ATRX/DAXX loss can be reliably detected in FNAs of pancreatic neuroendocrine tumors. Cancer Cytopathol. 2017, 125, 544–551. [Google Scholar] [CrossRef] [PubMed]
- Heaphy, C.M.; de Wilde, R.F.; Jiao, Y.; Klein, A.P.; Edil, B.H.; Shi, C.; Bettegowda, C.; Rodriguez, F.J.; Eberhart, C.G.; Hebbar, S.; et al. Altered telomeres in tumors with ATRX and DAXX mutations. Science 2011, 333, 425. [Google Scholar] [CrossRef] [PubMed]
- Villa, R.; Daidone, M.G.; Motta, R.; Venturini, L.; De Marco, C.; Vannelli, A.; Kusamura, S.; Baratti, D.; Deraco, M.; Costa, A.; et al. Multiple mechanisms of telomere maintenance exist and differentially affect clinical outcome in diffuse malignant peritoneal mesothelioma. Clin. Cancer Res. 2008, 14, 4134–4140. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-K.; Park, N.-H.; Lee, H. Prognostic Value of Alternative Lengthening of Telomeres-Associated Biomarkers in Uterine Sarcoma and Uterine Carcinosarcoma. Int. J. Gynecol. Cancer 2012, 22, 434–441. [Google Scholar] [CrossRef] [PubMed]
- Pezzolo, A.; Pistorio, A.; Gambini, C.; Haupt, R.; Ferraro, M.; Erminio, G.; De Bernardi, B.; Garaventa, A.; Pistoia, V. Intratumoral diversity of telomere length in individual neuroblastoma tumors. Oncotarget 2015, 6, 7493–7503. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.E.; Varkonyi, R.J.; Schwalm, J.; Cragle, R.; Klein-Szanto, A.; Patchefsky, A.; Cukierman, E.; von Mehren, M.; Broccoli, D. Multiple mechanisms of telomere maintenance exist in liposarcomas. Clin. Cancer Res. 2005, 11, 5347–5355. [Google Scholar] [CrossRef] [PubMed]
- Costa, A.; Daidone, M.G.; Daprai, L.; Villa, R.; Cantu, S.; Pilotti, S.; Mariani, L.; Gronchi, A.; Henson, J.D.; Reddel, R.R.; et al. Telomere maintenance mechanisms in liposarcomas: Association with histologic subtypes and disease progression. Cancer Res. 2006, 66, 8918–8924. [Google Scholar] [CrossRef] [PubMed]
- Venturini, L.; Motta, R.; Gronchi, A.; Daidone, M.; Zaffaroni, N. Prognostic relevance of ALT-associated markers in liposarcoma: A comparative analysis. BMC Cancer 2010, 10, 254. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.C.; Jeng, Y.M.; Liau, J.Y.; Tsai, J.H.; Hsu, H.H.; Yang, C.Y. Alternative lengthening of telomeres and loss of ATRX are frequent events in pleomorphic and dedifferentiated liposarcomas. Mod. Pathol. 2015, 28, 1064–1073. [Google Scholar] [CrossRef] [PubMed]
- Liau, J.Y.; Lee, J.C.; Tsai, J.H.; Yang, C.Y.; Liu, T.L.; Ke, Z.L.; Hsu, H.H.; Jeng, Y.M. Comprehensive screening of alternative lengthening of telomeres phenotype and loss of ATRX expression in sarcomas. Mod. Pathol. 2015, 28, 1545–1554. [Google Scholar] [CrossRef] [PubMed]
- Liau, J.Y.; Tsai, J.H.; Jeng, Y.M.; Lee, J.C.; Hsu, H.H.; Yang, C.Y. Leiomyosarcoma with Alternative Lengthening of Telomeres Is Associated with Aggressive Histologic Features, Loss of ATRX Expression, and Poor Clinical Outcome. Am. J. Surg. Pathol. 2015, 39, 236–244. [Google Scholar] [CrossRef] [PubMed]
- Ohali, A.; Avigad, S.; Naumov, I.; Goshen, Y.; Ash, S.; Yaniv, I. Different telomere maintenance mechanisms in alveolar and embryonal rhabdomyosarcoma. Genes Chromosomes Cancer 2008, 47, 965–970. [Google Scholar] [CrossRef] [PubMed]
- Liau, J.Y.; Tsai, J.H.; Yang, C.Y.; Lee, J.C.; Liang, C.W.; Hsu, H.H.; Jeng, Y.M. Alternative lengthening of telomeres phenotype in malignant vascular tumors is highly associated with loss of ATRX expression and is frequently observed in hepatic angiosarcomas. Hum. Pathol. 2015, 46, 1360–1366. [Google Scholar] [CrossRef] [PubMed]
- Venturini, L.; Daidone, M.G.; Motta, R.; Cimino-Reale, G.; Hoare, S.F.; Gronchi, A.; Folini, M.; Keith, W.N.; Zaffaroni, N. Telomere maintenance mechanisms in malignant peripheral nerve sheath tumors: Expression and prognostic relevance. Neuro-Oncology 2012, 14, 736–744. [Google Scholar] [CrossRef] [PubMed]
- Ulaner, G.A.; Huang, H.Y.; Otero, J.; Zhao, Z.Q.; Ben-Porat, L.; Satagopan, J.M.; Gorlick, R.; Meyers, P.; Healey, J.H.; Huvos, A.G.; et al. Absence of a telomere maintenance mechanism as a favorable prognostic factor in patients with osteosarcoma. Cancer Res. 2003, 63, 1759–1763. [Google Scholar] [PubMed]
- Sanders, R.P.; Drissi, R.; Billups, C.A.; Daw, N.C.; Valentine, M.B.; Dome, J.S. Telomerase expression predicts unfavorable outcome in osteosarcoma. J. Clin. Oncol. 2004, 22, 3790–3797. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, T.; Shay, J.W.; Wright, W.E.; Hiyama, E.; Shimose, S.; Kubo, T.; Sugita, T.; Yasunaga, Y.; Ochi, M. Telomere-maintenance mechanisms in soft-tissue malignant fibrous histiocytomas. J. Bone Joint Surg. Am. 2009, 91, 928–937. [Google Scholar] [CrossRef] [PubMed]
- Venturini, L.; Daidone, M.G.; Motta, R.; Collini, P.; Spreafico, F.; Terenziani, M.; Piva, L.; Radice, P.; Perotti, D.; Zaffaroni, N. Telomere maintenance in Wilms tumors: First evidence for the presence of alternative lengthening of telomeres mechanism. Genes Chromosomes Cancer 2011, 50, 823–829. [Google Scholar] [CrossRef] [PubMed]
- Nault, J.C.; Calderaro, J.; Di Tommaso, L.; Balabaud, C.; Zafrani, E.S.; Bioulac-Sage, P.; Roncalli, M.; Zucman-Rossi, J. Telomerase reverse transcriptase promoter mutation is an early somatic genetic alteration in the transformation of premalignant nodules in hepatocellular carcinoma on cirrhosis. Hepatology 2014, 60, 1983–1992. [Google Scholar] [CrossRef] [PubMed]
- Pinyol, R.; Tovar, V.; Llovet, J.M. TERT promoter mutations: Gatekeeper and driver of hepatocellular carcinoma. J. Hepatol. 2014, 61, 685–687. [Google Scholar] [CrossRef] [PubMed]
- Bae, J.S.; Kim, Y.; Jeon, S.; Kim, S.H.; Kim, T.J.; Lee, S.; Kim, M.H.; Lim, D.J.; Lee, Y.S.; Jung, C.K. Clinical utility of TERT promoter mutations and ALK rearrangement in thyroid cancer patients with a high prevalence of the BRAFV600E mutation. Diagn. Pathol. 2016, 11, 21. [Google Scholar] [CrossRef] [PubMed]
- Scarpa, A.; Chang, D.K.; Nones, K.; Corbo, V.; Patch, A.M.; Bailey, P.; Lawlor, R.T.; Johns, A.L.; Miller, D.K.; Mafficini, A.; et al. Whole-genome landscape of pancreatic neuroendocrine tumours. Nature 2017, 543, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Tian, D.; Chen, Q.; Zhang, S.; Liu, B.; Ji, B. Telomerase reverse transcriptase (TERT) rs2736100 polymorphism contributes to increased risk of glioma: Evidence from a meta-analysis. Int. J. Clin. Exp. Med. 2015, 8, 422–430. [Google Scholar] [PubMed]
- Cao, J.L.; Yuan, P.; Abuduwufuer, A.; Lv, W.; Yang, Y.H.; Hu, J. Association between the TERT Genetic Polymorphism rs2853676 and Cancer Risk: Meta-Analysis of 76,108 Cases and 134,215 Controls. PLoS ONE 2015, 10, e0128829. [Google Scholar] [CrossRef] [PubMed]
- Schwartzentruber, J.; Korshunov, A.; Liu, X.Y.; Jones, D.T.; Pfaff, E.; Jacob, K.; Sturm, D.; Fontebasso, A.M.; Quang, D.A.; Tonjes, M.; et al. Driver mutations in histone H3.3 and chromatin remodelling genes in paediatric glioblastoma. Nature 2012, 482, 226–231. [Google Scholar] [CrossRef] [PubMed]
- Castelo-Branco, P.; Choufani, S.; Mack, S.; Gallagher, D.; Zhang, C.; Lipman, T.; Zhukova, N.; Walker, E.J.; Martin, D.; Merino, D.; et al. Methylation of the TERT promoter and risk stratification of childhood brain tumours: An integrative genomic and molecular study. Lancet Oncol. 2013, 14, 534–542. [Google Scholar] [CrossRef]
- Hertwig, F.; Peifer, M.; Fischer, M. Telomere maintenance is pivotal for high-risk neuroblastoma. Cell Cycle 2016, 15, 311–312. [Google Scholar] [CrossRef] [PubMed]
- Kawashima, M.; Kojima, M.; Ueda, Y.; Kurihara, S.; Hiyama, E. Telomere biology including TERT rearrangements in neuroblastoma: A useful indicator for surgical treatments. J. Pediatr. Surg. 2016, 51, 2080–2085. [Google Scholar] [CrossRef] [PubMed]
- Duan, X.F.; Zhao, Q. TERT-mediated and ATRX-mediated Telomere Maintenance and Neuroblastoma. J. Pediatr. Hematol. Oncol. 2018, 40, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Onitake, Y.; Hiyama, E.; Kamei, N.; Yamaoka, H.; Sueda, T.; Hiyama, K. Telomere biology in neuroblastoma: Telomere binding proteins and alternative strengthening of telomeres. J.Pediatr. Surg. 2009, 44, 2258–2266. [Google Scholar] [CrossRef] [PubMed]
- Oikawa, T.; Yamada, T. Molecular biology of the ETS family of transcription factors. Gene 2003, 303, 11–34. [Google Scholar] [CrossRef]
- Borah, S.; Xi, L.; Zaug, A.J.; Powell, N.M.; Dancik, G.M.; Cohen, S.B.; Costello, J.C.; Theodorescu, D.; Cech, T.R. TERT promoter mutations and telomerase reactivation in urothelial cancer. Science 2015, 347, 1006–1010. [Google Scholar]
- Xi, L.H.; Schmidt, J.C.; Zaug, A.J.; Ascarrunz, D.R.; Cech, T.R. A novel two-step genome editing strategy with CRISPR-Cas9 provides new insights into telomerase action and TERT gene expression. Genome Biol. 2015, 16, 231. [Google Scholar] [CrossRef] [PubMed]
- Heidenreich, B.; Rachakonda, P.S.; Hemminki, K.; Kumar, R. TERT promoter mutations in cancer development. Curr. Opin. Genet. Dev. 2014, 24, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Labussiere, M.; Boisselier, B.; Mokhtari, K.; Di Stefano, A.L.; Rahimian, A.; Rossetto, M.; Ciccarino, P.; Saulnier, O.; Paterra, R.; Marie, Y.; et al. Combined analysis of TERT, EGFR, and IDH status defines distinct prognostic glioblastoma classes. Neurology 2014, 83, 1200–1206. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Qi, C.; Maling, G.; Xiang, W.; Yanhui, L.; Ruofei, L.; Yunhe, M.; Jiewen, L.; Qing, M. TERT mutation in glioma: Frequency, prognosis and risk. J. Clin. Neurosci. 2016, 26, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Nault, J.C.; Zucman-Rossi, J. TERT promoter mutations in primary liver tumors. Clin. Res. Hepatol. Gastroenterol. 2016, 40, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.L.; Jeng, Y.M.; Chang, C.N.; Lee, H.J.; Hsu, H.C.; Lai, P.L.; Yuan, R.H. TERT promoter mutation in resectable hepatocellular carcinomas: A strong association with hepatitis C infection and absence of hepatitis B infection. Int. J. Surg. 2014, 12, 659–665. [Google Scholar] [CrossRef] [PubMed]
- Pezzuto, F.; Buonaguro, L.; Buonaguro, F.M.; Tornesello, M.L. Frequency and geographic distribution of TERT promoter mutations in primary hepatocellular carcinoma. Infect. Agents Cancer 2017, 12, 27. [Google Scholar] [CrossRef] [PubMed]
- Melo, M.; da Rocha, A.G.; Vinagre, J.; Sobrinho-Simões, M.; Soares, P. Coexistence of TERT promoter and BRAF mutations in papillary thyroid carcinoma: Added value in patient prognosis? J. Clin. Oncol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Stoehr, R.; Taubert, H.; Zinnall, U.; Giedl, J.; Gaisa, N.T.; Burger, M.; Ruemmele, P.; Hurst, C.D.; Knowles, M.A.; Wullich, B.; et al. Frequency of TERT Promoter Mutations in Prostate Cancer. Pathobiology 2015, 82, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Chiba, K.; Lorbeer, F.K.; Shain, A.H.; McSwiggen, D.T.; Schruf, E.; Oh, A.; Ryu, J.; Darzacq, X.; Bastian, B.C.; Hockemeyer, D. Mutations in the promoter of the telomerase gene TERT contribute to tumorigenesis by a two-step mechanism. Science 2017, 357, 1416–1420. [Google Scholar] [CrossRef] [PubMed]
- Stratton, M.R.; Campbell, P.J.; Futreal, P.A. The cancer genome. Nature 2009, 458, 719. [Google Scholar] [CrossRef] [PubMed]
- Beroukhim, R.; Mermel, C.H.; Porter, D.; Wei, G.; Raychaudhuri, S.; Donovan, J.; Barretina, J.; Boehm, J.S.; Dobson, J.; Urashima, M. The landscape of somatic copy-number alteration across human cancers. Nature 2010, 463, 899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, T.-M.; Xi, R.; Luquette, L.J.; Park, R.W.; Johnson, M.D.; Park, P.J. Functional genomic analysis of chromosomal aberrations in a compendium of 8000 cancer genomes. Genome Res. 2013, 23, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Zack, T.I.; Schumacher, S.E.; Carter, S.L.; Cherniack, A.D.; Saksena, G.; Tabak, B.; Lawrence, M.S.; Zhang, C.-Z.; Wala, J.; Mermel, C.H. Pan-cancer patterns of somatic copy number alteration. Nat. Genet. 2013, 45, 1134. [Google Scholar] [CrossRef] [PubMed]
- Bryce, L.A.; Morrisont, N.; Hoare, S.F.; Muir, S.; Keith, W.N. Mapping of the gene for the human telomerase reverse transcriptase, hTERT, to chromosome 5p15. 33 by fluorescence in situ hybridization. Neoplasia 2000, 2, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Rooney, P.H.; Murray, G.I.; Stevenson, D.A.; Haites, N.E.; Cassidy, J.; McLeod, H.L. Comparative genomic hybridization and chromosomal instability in solid tumours. Br. J. Cancer 1999, 80, 862–873. [Google Scholar] [CrossRef] [PubMed]
- Santarius, T.; Shipley, J.; Brewer, D.; Stratton, M.R.; Cooper, C.S. A census of amplified and overexpressed human cancer genes. Nat. Rev. Cancer 2010, 10, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Knuutila, S.; Bjorkqvist, A.M.; Autio, K.; Tarkkanen, M.; Wolf, M.; Monni, O.; Szymanska, J.; Larramendy, M.L.; Tapper, J.; Pere, H.; et al. DNA copy number amplifications in human neoplasms: Review of comparative genomic hybridization studies. Am. J. Pathol. 1998, 152, 1107–1123. [Google Scholar] [PubMed]
- Iwabuchi, H.; Sakamoto, M.; Sakunaga, H.; Ma, Y.-Y.; Carcangiu, M.L.; Pinkel, D.; Yang-Feng, T.L.; Gray, J.W. Genetic analysis of benign, low-grade, and high-grade ovarian tumors. Cancer Res. 1995, 55, 6172–6180. [Google Scholar] [PubMed]
- Arnold, N.; Hägele, L.; Walz, L.; Schempp, W.; Pfisterer, J.; Bauknecht, T.; Kiechle, M. Overrepresentation of 3q and 8q material and loss of 18q material are recurrent findings in advanced human ovarian cancer. Genes Chromosomes Cancer 1996, 16, 46–54. [Google Scholar] [CrossRef]
- Balsara, B.R.; Sonoda, G.; Du Manoir, S.; Siegfried, J.M.; Gabrielson, E.; Testa, J.R. Comparative genomic hybridization analysis detects frequent, often high-level, overrepresentation of DNA sequences at 3q, 5p, 7p, and 8q in human non-small cell lung carcinomas. Cancer Res. 1997, 57, 2116–2120. [Google Scholar] [PubMed]
- Heselmeyer, K.; Schröck, E.; Du Manoir, S.; Blegen, H.; Shah, K.; Steinbeck, R.; Auer, G.; Ried, T. Gain of chromosome 3q defines the transition from severe dysplasia to invasive carcinoma of the uterine cervix. Proc. Natl. Acad. Sci. USA 1996, 93, 479–484. [Google Scholar] [CrossRef] [PubMed]
- Heselmeyer-Haddad, K.; Macville, M.; Schröck, E.; Blegen, H.; Hellström, A.-C.; Shah, K.; Auer, G.; Ried, T. Advanced-stage cervical carcinomas are defined by a recurrent pattern of chromosomal aberrations revealing high genetic instability and a consistent gain of chromosome arm 3q. Genes Chromosom. Cancer 1997, 19, 233–240. [Google Scholar] [CrossRef]
- Ried, T.; Petersen, I.; Holtgreve-Grez, H.; Speicher, M.R.; Schröck, E.; du Manoir, S.; Cremer, T. Mapping of multiple DNA gains and losses in primary small cell lung carcinomas by comparative genomic hybridization. Cancer Res. 1994, 54, 1801–1806. [Google Scholar] [PubMed]
- Levin, N.A.; Brzoska, P.; Gupta, N.; Minna, J.D.; Gray, J.W.; Christman, M.F. Identification of frequent novel genetic alterations in small cell lung carcinoma. Cancer Res. 1994, 54, 5086–5091. [Google Scholar] [PubMed]
- Levin, N.A.; Bnorka, P.M.; Warnock, M.L.; Gray, J.W.; Christman, M.F. Identification of novel regions of altered DNA copy number in small cell lung tumors. Genes Chromosomes Cancer 1995, 13, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Brass, N.; Ukena, I.; Remberger, K.; Mack, U.; Sybrecht, G.; Meese, E. DNA amplification on chromosome 3q26.1-q26.3 in squamous cell carcinoma of the lung detected by reverse chromosome painting. Eur. J. Cancer 1996, 32, 1205–1208. [Google Scholar] [CrossRef]
- Brzoska, P.M.; Levin, N.A.; Fu, K.K.; Kaplan, M.J.; Singer, M.I.; Gray, J.W.; Christman, M.F. Frequent novel DNA copy number increase in squamous cell head and neck tumors. Cancer Res. 1995, 55, 3055–3059. [Google Scholar] [PubMed]
- Speicher, M.R.; Howe, C.; Crotty, P.; du Manoir, S.; Costa, J.; Ward, D.C. Comparative genomic hybridization detects novel deletions and amplifications in head and neck squamous cell carcinomas. Cancer Res. 1995, 55, 1010–1013. [Google Scholar] [PubMed]
- Bockmühl, U.; Schwendel, A.; Dietel, M.; Petersen, I. Distinct patterns of chromosomal alterations in high-and low-grade head and neck squamous cell carcinomas. Cancer Res. 1996, 56, 5325–5329. [Google Scholar] [PubMed]
- Dierlamm, J.; Rosenberg, C.; Stul, M.; Pittaluga, S.; Wlodarska, I.; Michaux, L.; Dehaen, M.; Verhoef, G.; Thomas, J.; De Kelver, W. Characteristic pattern of chromosomal gains and losses in marginal zone B cell lymphoma detected by comparative genomic hybridization. Leukemia 1997, 11, 747. [Google Scholar] [CrossRef] [PubMed]
- Björkqvist, A.-M.; Husgafvel-Pursiainen, K.; Anttila, S.; Karjalainen, A.; Tammilehto, L.; Mattson, K.; Vainio, H.; Knuutila, S. DNA gains in 3q occur frequently in squamous cell carcinoma of the lung, but not in adenocarcinoma. Genes Chromosomes Cancer 1998, 22, 79–82. [Google Scholar] [CrossRef]
- Tarkkanen, M.; Elomaa, I.; Blomqvist, C.; Kivioja, A.H.; Kellokumpu-Lehtinen, P.; Bohling, T.; Valle, J.; Knuutila, S. DNA sequence copy number increase at 8q: A potential new prognostic marker in high-grade osteosarcoma. Int. J. Cancer 1999, 84, 114–121. [Google Scholar] [CrossRef]
- Larramendy, M.L.; Tarkkanen, M.; Blomqvist, C.; Virolainen, M.; Wiklund, T.; Asko-Seljavaara, S.; Elomaa, I.; Knuutila, S. Comparative genomic hybridization of malignant fibrous histiocytoma reveals a novel prognostic marker. Am. J. Pathol. 1997, 151, 1153–1161. [Google Scholar] [PubMed]
- El-Rifai, W.; Sarlomo-Rikala, M.; Miettinen, M.; Knuutila, S.; Andersson, L.C. DNA copy number losses in chromosome 14: An early change in gastrointestinal stromal tumors. Cancer Res. 1996, 56, 3230–3233. [Google Scholar] [PubMed]
- Zhang, A.; Zheng, C.; Hou, M.; Lindvall, C.; Wallin, K.L.; Ångström, T.; Yang, X.; Hellström, A.C.; Blennow, E.; Björkholm, M. Amplification of the telomerase reverse transcriptase (hTERT) gene in cervical carcinomas. Genes Chromosomes Cancer 2002, 34, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Flacco, A.; Ludovini, V.; Bianconi, F.; Ragusa, M.; Bellezza, G.; Tofanetti, F.R.; Pistola, L.; Siggillino, A.; Vannucci, J.; Cagini, L.; et al. MYC and human telomerase gene (TERC) copy number gain in early-stage non-small cell lung cancer. Am. J. Clin. Oncol. 2015, 38, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Hwang, K.T.; Han, W.; Cho, J.; Lee, J.W.; Ko, E.; Kim, E.K.; Jung, S.Y.; Jeong, E.M.; Bae, J.Y.; Kang, J.J.; et al. Genomic copy number alterations as predictive markers of systemic recurrence in breast cancer. Int. J. Cancer 2008, 123, 1807–1815. [Google Scholar] [CrossRef] [PubMed]
- Heselmeyer-Haddad, K.; Janz, V.; Castle, P.E.; Chaudhri, N.; White, N.; Wilber, K.; Morrison, L.E.; Auer, G.; Burroughs, F.H.; Sherman, M.E.; et al. Detection of genomic amplification of the human telomerase gene (TERC) in cytologic specimens as a genetic test for the diagnosis of cervical dysplasia. Am. J. Pathol. 2003, 163, 1405–1416. [Google Scholar] [CrossRef]
- Heselmeyer-Haddad, K.; Sommerfeld, K.; White, N.M.; Chaudhri, N.; Morrison, L.E.; Palanisamy, N.; Wang, Z.Y.; Auer, G.; Steinberg, W.; Ried, T. Genomic amplification of the human telomerase gene (TERC) in pap smears predicts the development of cervical cancer. Am. J. Pathol. 2005, 166, 1229–1238. [Google Scholar] [CrossRef]
- Baird, D.M. Variation at the TERT locus and predisposition for cancer. Exp. Rev. Mol. Med. 2010, 12, e16. [Google Scholar] [CrossRef] [PubMed]
- McKay, J.D.; Hung, R.J.; Gaborieau, V.; Boffetta, P.; Chabrier, A.; Byrnes, G.; Zaridze, D.; Mukeria, A.; Szeszenia-Dabrowska, N.; Lissowska, J.; et al. Lung cancer susceptibility locus at 5p15.33. Nat. Genet. 2008, 40, 1404–1406. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.M.; Zhang, X.Y.; Jin, B. TERT genetic polymorphism rs2736100 was associated with lung cancer: A meta-analysis based on 14,492 subjects. Genet. Test. Mol. Biomark. 2013, 17, 937–941. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Jiao, S. Increased lung cancer risk associated with the TERT rs2736100 polymorphism: An updated meta-analysis. Tumor Biol. 2014, 35, 5763–5769. [Google Scholar] [CrossRef] [PubMed]
- Choi, B.J.; Yoon, J.H.; Kim, O.; Choi, W.S.; Nam, S.W.; Lee, J.Y.; Park, W.S. Influence of the hTERT rs2736100 polymorphism on telomere length in gastric cancer. World J. Gastroenterol. 2015, 21, 9328–9336. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.T.; Meng, Y.; Li, P.; Ge, N.; Kong, F.; Yang, L.; Bjorkholm, M.; Zhao, S.T.; Xu, D.W. The association between the TERT rs2736100 AC genotype and reduced risk of upper tract urothelial carcinomas in a Han Chinese population. Oncotarget 2016, 7, 31972–31979. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Yu, H.; Sun, J.; Qi, J.; Liu, Q.; Li, R.; Zheng, S.L.; Xu, J.; Kang, J. Association of genetic polymorphisms in the telomerase reverse transcriptase gene with prostate cancer aggressiveness. Mol. Med. Rep. 2015, 12, 489–497. [Google Scholar] [CrossRef] [PubMed]
- Zou, P.; Gu, A.; Ji, G.; Zhao, L.; Zhao, P.; Lu, A. The TERT rs2736100 polymorphism and cancer risk: A meta-analysis based on 25 case-control studies. BMC Cancer 2012, 12, 7. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Yin, Z.; Wu, W.; Li, X.; Zhou, B. Genetic variants in TERT-CLPTM1L genetic region associated with several types of cancer: A meta-analysis. Gene 2013, 526, 390–399. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Li, G.; Wei, S.; Niu, J.; Wang, L.E.; Sturgis, E.M.; Wei, Q. Genetic variations in TERT-CLPTM1L genes and risk of squamous cell carcinoma of the head and neck. Carcinogenesis 2010, 31, 1977–1981. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Qiao, N.; Wang, Y.; Jiang, M.; Wang, S.; Wang, C.; Hu, L. Association between the telomerase reverse transcriptase (TERT) rs2736098 polymorphism and cancer risk: Evidence from a case-control study of non-small-cell lung cancer and a meta-analysis. PLoS ONE 2013, 8, e76372. [Google Scholar] [CrossRef] [PubMed]
- Prescott, J.; McGrath, M.; Lee, I.M.; Buring, J.E.; De Vivo, I. Telomere length and genetic analyses in population-based studies of endometrial cancer risk. Cancer 2010, 116, 4275–4282. [Google Scholar] [CrossRef] [PubMed]
- Hofer, P.; Baierl, A.; Bernhart, K.; Leeb, G.; Mach, K.; Micksche, M.; Gsur, A. Association of genetic variants of human telomerase with colorectal polyps and colorectal cancer risk. Mol. Carcinog. 2012, 51 (Suppl. 1), E176–E182. [Google Scholar] [CrossRef] [PubMed]
- Pellatt, A.J.; Wolff, R.K.; Herrick, J.; Lundgreen, A.; Slattery, M.L. TERT’s role in colorectal carcinogenesis. Mol. Carcinog. 2013, 52, 507–513. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.L.; Ogundiran, T.O.; Adebamowo, C.; Nathanson, K.L.; Domchek, S.M.; Rebbeck, T.R.; Simon, M.S.; John, E.M.; Hennis, A.; Nemesure, B.; et al. Lack of association between common single nucleotide polymorphisms in the TERT-CLPTM1L locus and breast cancer in women of African ancestry. Breast Cancer Res. Treat. 2012, 132, 341–345. [Google Scholar] [CrossRef] [PubMed]
- Varadi, V.; Brendle, A.; Grzybowska, E.; Johansson, R.; Enquist, K.; Butkiewicz, D.; Pamula-Pilat, J.; Pekala, W.; Hemminki, K.; Lenner, P.; et al. A functional promoter polymorphism in the TERT gene does not affect inherited susceptibility to breast cancer. Cancer Genet. Cytogenet. 2009, 190, 71–74. [Google Scholar] [CrossRef] [PubMed]
- Park, C.K.; Lee, S.H.; Kim, J.Y.; Kim, J.E.; Kim, T.M.; Lee, S.T.; Choi, S.H.; Park, S.H.; Kim, I.H. Expression level of hTERT is regulated by somatic mutation and common single nucleotide polymorphism at promoter region in glioblastoma. Oncotarget 2014, 5, 3399–3407. [Google Scholar] [CrossRef] [PubMed]
- Shen, N.; Lu, Y.J.; Wang, X.; Peng, J.; Zhu, Y.W.; Cheng, L.M. Association between rs2853669 in TERT gene and the risk and prognosis of human cancer: A systematic review and meta-analysis. Oncotarget 2017, 8, 50864–50872. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Rozadilla, C.; Kartsonaki, C.; Woolley, C.; McClellan, M.; Whittington, D.; Horgan, G.; Leedham, S.; Kriaucionis, S.; East, J.; Tomlinson, I. Telomere length and genetics are independent colorectal tumour risk factors in an evaluation of biomarkers in normal bowel. Br. J. Cancer 2018, 118, 727–732. [Google Scholar] [CrossRef] [PubMed]
- Stephens, P.J.; McBride, D.J.; Lin, M.-L.; Varela, I.; Pleasance, E.D.; Simpson, J.T.; Stebbings, L.A.; Leroy, C.; Edkins, S.; Mudie, L.J. Complex landscapes of somatic rearrangement in human breast cancer genomes. Nature 2009, 462, 1005. [Google Scholar] [CrossRef] [PubMed]
- Valentijn, L.J.; Koster, J.; Zwijnenburg, D.A.; Hasselt, N.E.; van Sluis, P.; Volckmann, R.; van Noesel, M.M.; George, R.E.; Tytgat, G.A.; Molenaar, J.J.; et al. TERT rearrangements are frequent in neuroblastoma and identify aggressive tumors. Nat. Genet. 2015, 47, 1411–1414. [Google Scholar] [CrossRef] [PubMed]
- Schwab, M.; Alitalo, K.; Klempnauer, K.-H.; Varmus, H.E.; Bishop, J.M.; Gilbert, F.; Brodeur, G.; Goldstein, M.; Trent, J. Amplified DNA with limited homology to myc cellular oncogene is shared by human neuroblastoma cell lines and a neuroblastoma tumour. Nature 1983, 305, 245. [Google Scholar] [CrossRef] [PubMed]
- Poremba, C.; Hero, B.; Heine, B.; Scheel, C.; Schaefer, K.-L.; Christiansen, H.; Berthold, F.; Kneif, S.; Stein, H.; Juergens, H. Telomerase is a strong indicator for assessing the proneness to progression in neuroblastomas. Med. Pediatr. Oncol. 2000, 35, 651–655. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, S.; Popova, E.Y.; Grigoryev, S.A.; Zhu, J. Rearrangement of upstream sequences of the hTERT gene during cellular immortalization. Genes Chromosomes Cancer 2009, 48, 963–974. [Google Scholar] [CrossRef] [PubMed]
- Nik-Zainal, S.; Alexandrov, L.B.; Wedge, D.C.; Van Loo, P.; Greenman, C.D.; Raine, K.; Jones, D.; Hinton, J.; Marshall, J.; Stebbings, L.A. Mutational processes molding the genomes of 21 breast cancers. Cell 2012, 149, 979–993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexandrov, L.B.; Nik-Zainal, S.; Wedge, D.C.; Aparicio, S.A.; Behjati, S.; Biankin, A.V.; Bignell, G.R.; Bolli, N.; Borg, A.; Børresen-Dale, A.-L. Signatures of mutational processes in human cancer. Nature 2013, 500, 415. [Google Scholar] [CrossRef] [PubMed]
- Ohali, A.; Avigad, S.; Ash, S.; Goshen, Y.; Luria, D.; Feinmesser, M.; Zaizov, R.; Yaniv, I. Telomere length is a prognostic factor in neuroblastoma. Cancer 2006, 107, 1391–1399. [Google Scholar] [CrossRef] [PubMed]
- Jeison, M.; Yaniv, I.; Ash, S. Genetic stratification of neuroblastoma for treatment tailoring. Future Oncol. 2011, 7, 1087–1099. [Google Scholar] [CrossRef] [PubMed]
- Cahill, D.P.; Louis, D.N.; Cairncross, J.G. Molecular background of oligodendroglioma: 1p/19q, IDH, TERT, CIC and FUBP1. CNS Oncol. 2015, 4, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Handy, D.E.; Castro, R.; Loscalzo, J. Epigenetic modifications: Basic mechanisms and role in cardiovascular disease. Circulation 2011, 123, 2145–2156. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.; Rao, C.M. Epigenetics in cancer: Fundamentals and Beyond. Pharmacol. Ther. 2017, 173, 118–134. [Google Scholar] [CrossRef] [PubMed]
- Falus, A.; Marton, I.; Borbényi, E.; Tahy, Á.; Karádi, P.; Aradi, J.; Stauder, A.; Kopp, M. A challenging epigenetic message: Telomerase activity is associated with complex changes in lifestyle. Cell Biol. Int. 2011, 35, 1079–1083. [Google Scholar] [CrossRef] [PubMed]
- Entringer, S.; Epel, E.S.; Kumsta, R.; Lin, J.; Hellhammer, D.H.; Blackburn, E.H.; Wüst, S.; Wadhwa, P.D. Stress exposure in intrauterine life is associated with shorter telomere length in young adulthood. Proc. Natl. Acad. Sci. USA 2011, 108, E513–E518. [Google Scholar] [CrossRef] [PubMed]
- Price, L.H.; Kao, H.-T.; Burgers, D.E.; Carpenter, L.L.; Tyrka, A.R. Telomeres and early-life stress: An overview. Biol. Psychiatry 2013, 73, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Epel, E.; Blackburn, E. Telomeres and lifestyle factors: Roles in cellular aging. Mutat. Res. Fundam. Mol. Mech. Mutagen. 2012, 730, 85–89. [Google Scholar] [CrossRef] [PubMed]
- Lewis, K.A.; Tollefsbol, T.O. Regulation of the Telomerase Reverse Transcriptase Subunit through Epigenetic Mechanisms. Front. Genet. 2016, 7, 83. [Google Scholar] [CrossRef] [PubMed]
- Hardy, T.M.; Tollefsbol, T.O. Epigenetic diet: Impact on the epigenome and cancer. Epigenomics 2011, 3, 503–518. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.A.; Takai, D. The role of DNA methylation in mammalian epigenetics. Science 2001, 293, 1068–1070. [Google Scholar] [CrossRef] [PubMed]
- Feng, W.; Han, Z.; Zhu, R.; Liu, P.; Liu, S. Association of p16 gene methylation with prostate cancer risk: A meta-analysis. J. BUON 2015, 20, 1074–1080. [Google Scholar] [PubMed]
- Herman, J.G.; Umar, A.; Polyak, K.; Graff, J.R.; Ahuja, N.; Issa, J.-P.J.; Markowitz, S.; Willson, J.K.; Hamilton, S.R.; Kinzler, K.W. Incidence and functional consequences of hMLH1 promoter hypermethylation in colorectal carcinoma. Proc. Natl. Acad. Sci. USA 1998, 95, 6870–6875. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yao, X.; Wang, Y.; Hu, F.; Wang, F.; Jiang, L.; Liu, Y.; Wang, D.; Sun, G.; Zhao, Y. MLH1 promoter methylation frequency in colorectal cancer patients and related clinicopathological and molecular features. PLoS ONE 2013, 8, e59064. [Google Scholar] [CrossRef] [PubMed]
- Szyf, M.; Pakneshan, P.; Rabbani, S.A. DNA methylation and breast cancer. Biochem. Pharmacol. 2004, 68, 1187–1197. [Google Scholar] [CrossRef] [PubMed]
- Dessain, S.K.; Yu, H.-Y.; Reddel, R.R.; Beijersbergen, R.L.; Weinberg, R.A. Methylation of the human telomerase gene CpG island. Cancer Res. 2000, 60, 537–541. [Google Scholar] [PubMed]
- Guilleret, I.; Benhattar, J. Demethylation of the human telomerase catalytic subunit (hTERT) gene promoter reduced hTERT expression and telomerase activity and shortened telomeres. Exp. Cell Res. 2003, 289, 326–334. [Google Scholar] [CrossRef]
- Guilleret, I.; Yan, P.; Grange, F.; Braunschweig, R.; Bosman, F.T.; Benhattar, J. Hypermethylation of the human telomerase catalytic subunit (hTERT) gene correlates with telomerase activity. Int. J. Cancer 2002, 101, 335–341. [Google Scholar] [CrossRef] [PubMed]
- Gigek, C.O.; Leal, M.F.; Silva, P.N.O.; Lisboa, L.C.F.; Lima, E.M.; Calcagno, D.Q.; Assumpcao, P.P.; Burbano, R.R.; Smith, M.D.A.C. hTERT methylation and expression in gastric cancer. Biomarkers 2009, 14, 630–636. [Google Scholar] [CrossRef] [PubMed]
- Widschwendter, A.; Müller, H.M.; Hubalek, M.M.; Wiedemair, A.; Fiegl, H.; Goebel, G.; Mueller-Holzner, E.; Marth, C.; Widschwendter, M. Methylation status and expression of human telomerase reverse transcriptase in ovarian and cervical cancer. Gynecol. Oncol. 2004, 93, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Oikonomou, P.; Messinis, I.; Tsezou, A. DNA methylation is not likely to be responsible for hTERT expression in premalignant cervical lesions. Exp. Biol. Med. 2007, 232, 881–886. [Google Scholar]
- Fan, Y.; Lee, S.; Wu, G.; Easton, J.; Yergeau, D.; Dummer, R.; Vogel, P.; Kirkwood, J.M.; Barnhill, R.L.; Pappo, A.; et al. Telomerase Expression by Aberrant Methylation of the TERT Promoter in Melanoma Arising in Giant Congenital Nevi. J. Investig. Dermatol. 2016, 136, 339–342. [Google Scholar] [CrossRef] [PubMed]
- Renaud, S.; Loukinov, D.; Abdullaev, Z.; Guilleret, I.; Bosman, F.T.; Lobanenkov, V.; Benhattar, J. Dual role of DNA methylation inside and outside of CTCF-binding regions in the transcriptional regulation of the telomerase hTERT gene. Nucleic Acids Res. 2007, 35, 1245–1256. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Katzenellenbogen, R.A.; Grandori, C.; Galloway, D.A. An unbiased in vivo screen reveals multiple transcription factors that control HPV E6-regulated hTERT in keratinocytes. Virology 2013, 446, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Valls-Bautista, C.; Bougel, S.; Piñol-Felis, C.; Viñas-Salas, J.; Benhattar, J. hTERT methylation is necessary but not sufficient for telomerase activity in colorectal cells. Oncol. Lett. 2011, 2, 1257–1260. [Google Scholar] [CrossRef] [PubMed]
- Bechter, O.E.; Eisterer, W.; Dlaska, M.; Kühr, T.; Thaler, J. CpG island methylation of the hTERT promoter is associated with lower telomerase activity in B-cell lymphocytic leukemia. Exp. Hematol. 2002, 30, 26–33. [Google Scholar] [CrossRef]
- Choi, J.-H.; Park, S.H.; Park, J.; Park, B.G.; Cha, S.-J.; Kong, K.-H.; Lee, K.-H.; Park, A.J. Site-specific methylation of CpG nucleotides in the hTERT promoter region can control the expression of hTERT during malignant progression of colorectal carcinoma. Biochem. Biophys. Res. Commun. 2007, 361, 615–620. [Google Scholar] [CrossRef] [PubMed]
- Kumari, A.; Srinivasan, R.; Vasishta, R.K.; Wig, J.D. Positive regulation of human telomerase reverse transcriptase gene expression and telomerase activity by DNA methylation in pancreatic cancer. Ann. Surg. Oncol. 2009, 16, 1051–1059. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Petko, Z.; Dzieciatkowski, S.; Lin, L.; Ghiassi, M.; Stain, S.; Chapman, W.C.; Washington, M.K.; Willis, J.; Markowitz, S.D. CpG island methylation of genes accumulates during the adenoma progression step of the multistep pathogenesis of colorectal cancer. Genes Chromosomes Cancer 2006, 45, 781–789. [Google Scholar] [CrossRef] [PubMed]
- Iliopoulos, D.; Satra, M.; Drakaki, A.; Poultsides, G.A.; Tsezou, A. Epigenetic regulation of hTERT promoter in hepatocellular carcinomas. Int. J. Oncol. 2009, 34, 391–399. [Google Scholar] [PubMed]
- Castelo-Branco, P.; Leao, R.; Lipman, T.; Campbell, B.; Lee, D.; Price, A.; Zhang, C.; Heidari, A.; Stephens, D.; Boerno, S.; et al. A cancer specific hypermethylation signature of the TERT promoter predicts biochemical relapse in prostate cancer: A retrospective cohort study. Oncotarget 2016, 7, 57726–57736. [Google Scholar] [CrossRef] [PubMed]
- Faleiro, I.; Apolonio, J.D.; Price, A.J.; De Mello, R.A.; Roberto, V.P.; Tabori, U.; Castelo-Branco, P. The TERT hypermethylated oncologic region predicts recurrence and survival in pancreatic cancer. Future Oncol. 2017, 13, 2045–2051. [Google Scholar] [CrossRef] [PubMed]
- Lundblad, V.; Blackburn, E.H. An alternative pathway for yeast telomere maintenance rescues est1− senescence. Cell 1993, 73, 347–360. [Google Scholar] [CrossRef]
- Bryan, T.M.; Englezou, A.; Gupta, J.; Bacchetti, S.; Reddel, R.R. Telomere Elongation in Immortal Human-Cells without Detectable Telomerase Activity. EMBO J. 1995, 14, 4240–4248. [Google Scholar] [PubMed]
- Dunham, M.A.; Neumann, A.A.; Fasching, C.L.; Reddel, R.R. Telomere maintenance by recombination in human cells. Nat. Genet. 2000, 26, 447–450. [Google Scholar] [CrossRef] [PubMed]
- Dilley, R.L.; Verma, P.; Cho, N.W.; Winters, H.D.; Wondisford, A.R.; Greenberg, R.A. Break-induced telomere synthesis underlies alternative telomere maintenance. Nature 2016, 539, 54–58. [Google Scholar] [CrossRef] [PubMed]
- Bryan, T.M.; Reddel, R.R. Telomere dynamics and telomerase activity in in vitro immortalised human cells. Eur. J. Cancer 1997, 33, 767–773. [Google Scholar] [CrossRef]
- Perrem, K.; Colgin, L.M.; Neumann, A.A.; Yeager, T.R.; Reddel, R.R. Coexistence of alternative lengthening of telomeres and telomerase in hTERT-transfected GM847 cells. Mol. Cell. Biol. 2001, 21, 3862–3875. [Google Scholar] [CrossRef] [PubMed]
- Yeager, T.R.; Neumann, A.A.; Englezou, A.; Huschtscha, L.I.; Noble, J.R.; Reddel, R.R. Telomerase-negative immortalized human cells contain a novel type of promyelocytic leukemia (PML) body. Cancer Res. 1999, 59, 4175–4179. [Google Scholar] [PubMed]
- Osterwald, S.; Deeg, K.I.; Chung, I.; Parisotto, D.; Worz, S.; Rohr, K.; Erfle, H.; Rippe, K. PML induces compaction, TRF2 depletion and DNA damage signaling at telomeres and promotes their alternative lengthening. J. Cell Sci. 2015, 128, 1887–1900. [Google Scholar] [CrossRef] [PubMed]
- Cesare, A.J.; Griffith, J.D. Telomeric DNA in ALT cells is characterized by free telomeric circles and heterogeneous T-loops. Mol. Cell. Biol. 2004, 24, 9948–9957. [Google Scholar] [CrossRef] [PubMed]
- Henson, J.D.; Reddel, R.R. Assaying and investigating Alternative Lengthening of Telomeres activity in human cells and cancers. FEBS Lett. 2010, 584, 3800–3811. [Google Scholar] [CrossRef] [PubMed]
- Amorim, J.P.; Santos, G.; Vinagre, J.; Soares, P. The Role of ATRX in the Alternative Lengthening of Telomeres (ALT) Phenotype. Genes 2016, 7, 66. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Y.; Shi, C.; Edil, B.H.; de Wilde, R.F.; Klimstra, D.S.; Maitra, A.; Schulick, R.D.; Tang, L.H.; Wolfgang, C.L.; Choti, M.A.; et al. DAXX/ATRX, MEN1, and mTOR pathway genes are frequently altered in pancreatic neuroendocrine tumors. Science 2011, 331, 1199–1203. [Google Scholar] [CrossRef] [PubMed]
- Sampl, S.; Pramhas, S.; Stern, C.; Preusser, M.; Marosi, C.; Holzmann, K. Expression of telomeres in astrocytoma WHO grade 2 to 4: TERRA level correlates with telomere length, telomerase activity, and advanced clinical grade. Transl. Oncol. 2012, 5, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Hung, N.; Chen, Y.J.; Taha, A.; Olivecrona, M.; Boet, R.; Wiles, A.; Warr, T.; Shaw, A.; Eiholzer, R.; Baguley, B.C.; Eccles, M.R.; Braithwaite, A.W.; Macfarlane, M.; Royds, J.A.; Slatter, T. Increased paired box transcription factor 8 has a survival function in glioma. BMC Cancer 2014, 14, 159. [Google Scholar] [CrossRef] [PubMed]
- Hung, N.A.; Eiholzer, R.A.; Kirs, S.; Zhou, J.; Ward-Hartstonge, K.; Wiles, A.K.; Frampton, C.M.; Taha, A.; Royds, J.A.; Slatter, T.L. Telomere profiles and tumor-associated macrophages with different immune signatures affect prognosis in glioblastoma. Mod. Pathol. 2016, 29, 212–226. [Google Scholar] [CrossRef] [PubMed]
- Ulaner, G.A.; Hoffman, A.R.; Otero, J.; Huang, H.Y.; Zhao, Z.Q.; Mazumdar, M.; Gorlick, R.; Meyers, P.; Healey, J.H.; Ladanyi, M. Divergent patterns of telomere maintenance mechanisms among human sarcomas: Sharply contrasting prevalence of the alternative lengthening of telomeres mechanism in Ewing’s sarcomas and osteosarcomas. Genes Chromosomes Cancer 2004, 41, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Gagos, S.; Papaioannou, G.; Chiourea, M.; Merk-Loretti, S.; Jefford, C.E.; Mikou, P.; Irminger-Finger, I.; Liossi, A.; Blouin, J.L.; Dahoun, S. Unusually stable abnormal karyotype in a highly aggressive melanoma negative for telomerase activity. Mol. Cytogenet. 2008, 1, 20. [Google Scholar] [CrossRef] [PubMed]
- Viceconte, N.; Dheur, M.S.; Majerova, E.; Pierreux, C.E.; Baurain, J.F.; van Baren, N.; Decottignies, A. Highly aggressive metastatic melanoma cells unable to maintain telomere length. Cell Rep. 2017, 19, 2529–2543. [Google Scholar] [CrossRef] [PubMed]
- Royds, J.A.; Al Nadaf, S.; Wiles, A.K.; Chen, Y.J.; Ahn, A.; Shaw, A.; Bowie, S.; Lam, F.; Baguley, B.C.; Braithwaite, A.W.; et al. The CDKN2A G500 allele is more frequent in GBM patients with no defined telomere maintenance mechanism tumors and is associated with poorer survival. PLoS ONE 2011, 6, e26737. [Google Scholar] [CrossRef] [PubMed]
- Schoeftner, S.; Blasco, M.A. Developmentally regulated transcription of mammalian telomeres by DNA-dependent RNA polymerase II. Nat. Cell Biol. 2008, 10, 228. [Google Scholar] [CrossRef] [PubMed]
- Luke, B.; Lingner, J. TERRA: Telomeric repeat-containing RNA. EMBO J. 2009, 28, 2503–2510. [Google Scholar] [CrossRef] [PubMed]
- Azzalin, C.M.; Reichenbach, P.; Khoriauli, L.; Giulotto, E.; Lingner, J. Telomeric repeat–containing RNA and RNA surveillance factors at mammalian chromosome ends. Science 2007, 318, 798–801. [Google Scholar] [CrossRef] [PubMed]
- MacNeil, D.E.; Bensoussan, H.J.; Autexier, C. Telomerase Regulation from Beginning to the End. Genes 2016, 7, 64. [Google Scholar] [CrossRef] [PubMed]
- Morris, K.V.; Mattick, J.S. The rise of regulatory RNA. Nat. Rev. Genet. 2014, 15, 423. [Google Scholar] [CrossRef] [PubMed]
- Montero, J.J.; Lopez de Silanes, I.; Grana, O.; Blasco, M.A. Telomeric RNAs are essential to maintain telomeres. Nat. Commun. 2016, 7, 12534. [Google Scholar] [CrossRef] [PubMed]
- Azhibek, D.; Skvortsov, D.; Andreeva, A.; Zatsepin, T.; Arutyunyan, A.; Zvereva, M.; Dontsova, O. TERRA mimicking ssRNAs prevail over the DNA substrate for telomerase in vitro due to interactions with the alternative binding site. J. Mol. Recognit. 2016, 29, 242–247. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.; Norseen, J.; Wiedmer, A.; Riethman, H.; Lieberman, P.M. TERRA RNA binding to TRF2 facilitates heterochromatin formation and ORC recruitment at telomeres. Mol. Cell 2009, 35, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Cusanelli, E.; Chartrand, P. Telomeric repeat-containing RNA TERRA: A noncoding RNA connecting telomere biology to genome integrity. Front. Genet. 2015, 6, 143. [Google Scholar] [CrossRef] [PubMed]
- Redon, S.; Reichenbach, P.; Lingner, J. The non-coding RNA TERRA is a natural ligand and direct inhibitor of human telomerase. Nucleic Acids Res. 2010, 38, 5797–5806. [Google Scholar] [CrossRef] [PubMed]
- De Silanes, I.L.; Grana, O.; De Bonis, M.L.; Dominguez, O.; Pisano, D.G.; Blasco, M.A. Identification of TERRA locus unveils a telomere protection role through association to nearly all chromosomes. Nat. Commun. 2014, 5, 4723. [Google Scholar] [CrossRef] [PubMed]
- Flynn, R.L.; Cox, K.E.; Jeitany, M.; Wakimoto, H.; Bryll, A.R.; Ganem, N.J.; Bersani, F.; Pineda, J.R.; Suva, M.L.; Benes, C.H.; et al. Alternative lengthening of telomeres renders cancer cells hypersensitive to ATR inhibitors. Science 2015, 347, 273–277. [Google Scholar] [CrossRef] [PubMed]
- Arora, R.; Lee, Y.; Wischnewski, H.; Brun, C.M.; Schwarz, T.; Azzalin, C.M. RNaseH1 regulates TERRA-telomeric DNA hybrids and telomere maintenance in ALT tumour cells. Nat. Commun. 2014, 5, 5220. [Google Scholar] [CrossRef] [PubMed]
- Arnoult, N.; Van Beneden, A.; Decottignies, A. Telomere length regulates TERRA levels through increased trimethylation of telomeric H3K9 and HP1alpha. Nat. Struct. Mol. Biol. 2012, 19, 948–956. [Google Scholar] [CrossRef] [PubMed]
- Porro, A.; Feuerhahn, S.; Delafontaine, J.; Riethman, H.; Rougemont, J.; Lingner, J. Functional characterization of the TERRA transcriptome at damaged telomeres. Nat. Commun. 2014, 5, 5379. [Google Scholar] [CrossRef] [PubMed]
- Cusanelli, E.; Romero, C.A.P.; Chartrand, P. Telomeric noncoding RNA TERRA is induced by telomere shortening to nucleate telomerase molecules at short telomeres. Mol. Cell 2013, 51, 780–791. [Google Scholar] [CrossRef] [PubMed]
- Ng, L.J.; Cropley, J.E.; Pickett, H.A.; Reddel, R.R.; Suter, C.M. Telomerase activity is associated with an increase in DNA methylation at the proximal subtelomere and a reduction in telomeric transcription. Nucleic Acids Res. 2009, 37, 1152–1159. [Google Scholar] [CrossRef] [PubMed]
- Lovejoy, C.A.; Li, W.; Reisenweber, S.; Thongthip, S.; Bruno, J.; de Lange, T.; De, S.; Petrini, J.H.; Sung, P.A.; Jasin, M.; et al. Loss of ATRX, genome instability, and an altered DNA damage response are hallmarks of the alternative lengthening of telomeres pathway. PLoS Genet. 2012, 8, e1002772. [Google Scholar] [CrossRef] [PubMed]



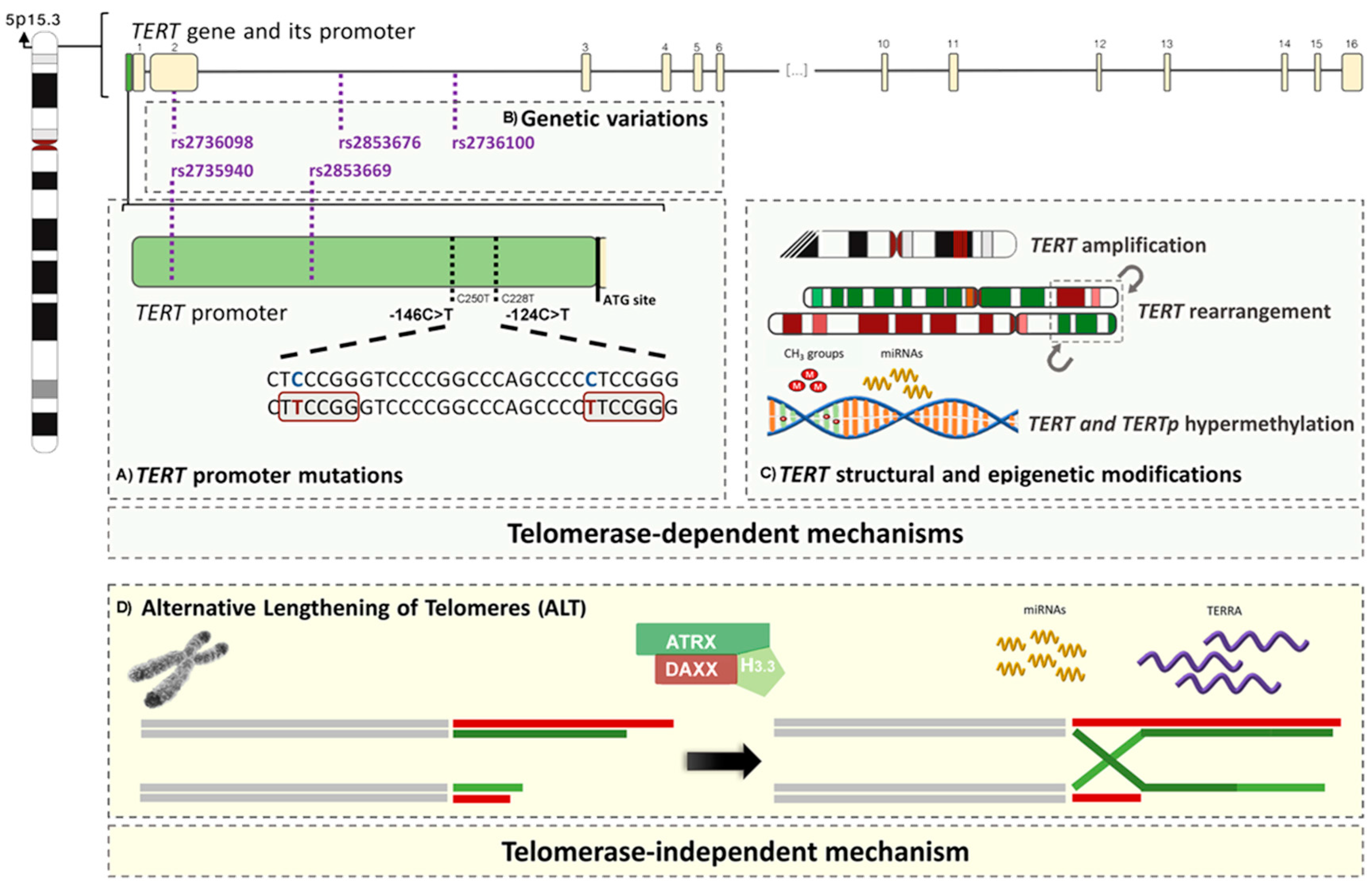{kind=link}
{kind=link}
| Tumour Type | Mutated Cases (Total) | Mutated Cases in % (Range) |
|---|---|---|
| Tumours of the breast | ||
| Fibroadenoma [58] | 4 (58) | 6.9 |
| Phyllodes tumour [58,59] | 41 (70) | 58.6 (45.8–65.2) |
| <5%: breast carcinoma, NOS (n = 88) and breast cancer, NOS (n = 122) [49,60]. | ||
| Tumours of the central nervous system | ||
| Astrocytoma (grades II-III), NOS [61,62,63] | 160 (699) | 22.9 (18.2–39.3) |
| Primary glioblastoma, NOS [50,64,65,66,67,68,69,70] | 1216 (1800) | 67.6 (46.3–85.9) |
| Secondary glioblastoma, NOS [50,64,65,67] | 25 (99) | 25.3 (5.0–35.7) |
| Paediatric glioblastoma, NOS [49,50] | 3 (51) | 5.9 (3.1–10.5) |
| Diffuse astrocytoma IDH-mutant [50,64,71,72,73] | 38 (242) | 15.7 (7.7–31.6) |
| Diffuse astrocytoma IDH-wildtype [50,64,71,72,73] | 23 (87) | 26.4 (7.1–58.3) |
| Diffuse astrocytoma, NOS [67,74,75,76,77] | 112 (574) | 19.5 (7.7–32.0) |
| Anaplastic astrocytoma, IDH-mutant [50,64,71,73,78,79] | 28 (248) | 11.3 (4.4–20.0) |
| Anaplastic astrocytoma, IDH-wildtype [50,64,71,73,78,79] | 104 (220) | 47.3 (20.0–80.0) |
| Anaplastic astrocytoma, NOS [67,74,75,76] | 29 (89) | 32.6 (10.0–40.0) |
| Glioblastoma, IDH-wildtype [61,62,63,64,67,68,71,74,77,78,79,80,81,82] | 2082 (2899) | 71.8 (27.6–91.7) |
| Giant cell [50] | 6 (17) | 35.3 |
| Gliosarcoma [50] | 21 (26) | 80.8 |
| Glioblastoma, IDH-mutant [61,62,63,64,67,68,71,74,77,78,79,80,81] | 114 (470) | 24.3 (0.0–76.9) |
| Glioblastoma, NOS a [66,75,76,83] | 328 (523) | 62.7 (46.3–84.0) |
| Oligodendroglioma, IDH-mutant and 1p/19q-codeleted [50,63] | 296 (311) | 95.2 |
| Oligodendroglioma, NOS [49,62,64,67,68,73,74,75,77] | 251 (469) | 53.5 (7.7–75.7) |
| Anaplastic oligodendroglioma, IDH-mutant and 1p/19q-codeleted [50] | 8 (38) | 21.1 |
| Anaplastic oligodendroglioma, NOS [49,50,64,67,73,74,76] | 171 (273) | 62.6 (0.0–88.5) |
| Oligodendroglioma (grades II–III), NOS [61,78] | 132 (152) | 86.8 (79.3–96.9) |
| Oligoastrocytoma, NOS b [49,50,61,62,64,67,73,74,76] | 222 (618) | 35.9 (3.8–57.9) |
| Anaplastic oligoastrocytoma, NOS [49,50,64,67,73,74,77] | 170 (415) | 41.0 (26.7–52.3) |
| Ependymoma c [49,50] | 6 (118) | 5.1 (0.0–6.9) |
| Choroid plexus carcinoma [50] | 1 (6) | 16.7 |
| Dysembryoplastic neuroepithelial tumour [49,50] | 1 (15) | 6.7 (0.0–33.3) |
| Desmoplastic infantile astrocytoma and ganglioglioma [50] | 1 (8) | 12.5 |
| Paraganglioma [50] | 1 (12) | 8.3 |
| Medulloblastoma, adult (>18 y) WNT-activated [50,84] | 5 (15) | 33.3 (30.8–50.0) |
| Medulloblastoma, adult (>18 y) SHH-MB [50,84,85,86] | 119 (134) | 88.8 (72.7–100.0) |
| Medulloblastoma, adult (>18 y), NOS [50] | 15 (23) | 65.2 |
| Medulloblastoma, paediatric SHH-MB d [50,84,86] | 49 (146) | 33.6 (20.0–31.9) |
| Medulloblastoma, paediatric, NOS [50,84,86] | 22 (121) | 18.2 (3.5–56.0) |
| Medulloblastoma, NOS [49,75,87] | 40 (166) | 24.1 (20.9–33.3) |
| Meningioma with malignant histology [88] | 5 (18) | 27.8 |
| <5%: pilocytic astrocytoma (n = 167) [50,75,76], pilomyxoid astrocytoma (n = 3) [50], subependymal giant cell astrocytoma (n = 11) [50,75], pleomorphic xanthoastrocytoma (n = 26) [50,75], mixopapillary ependymoma (n = 14) [50], subependymoma (n = 12) [50], anaplastic ependymoma (n = 48) [50], angiocentric glioma (n = 9) [50], choroid plexus papiloma (n = 13) [50], gangliocytoma (n = 2) [50], ganglioglioma (n = 40) [50], anaplastic ganglioglioma (n = 3) [50], papillary glioneuronal tumour (n = 1) [50], rosette-forming glioneuronal tumour (n = 6) [50], central neurocytoma (n = 28) [50], pineocytoma (n = 2) [50], pineal parenchymal tumours of intermediate differentiation (n = 9) [51], pineoblastoma (n = 5) [50], papillary tumours of the pineal region (n = 2) [50]), medulloblastoma, paediatric WNT-activated (n = 66) [50,84,86] and meningioma (n = 349) [49,75,89]. | ||
| Tumours of the conjunctiva and uvea | ||
| Primary acquired melanosis with atypia [90] | 2 (25) | 8.0 |
| Conjunctival melanoma [90,91,92] | 32 (84) | 38.1 (31.6–57.1) |
| Perilimbal squamous carcinoma [93] | 21 (48) | 43.8 |
| <5%: primary acquired melanosis without atypia (n = 13) [90], conjunctival naevi (n = 56) [90] and uveal melanomas (n = 202) [90,91,92,94]. | ||
| Tumours of the digestive system | ||
| Borderline hepatocellular adenoma/hepatocellular carcinoma [95] | 3 (18) | 16.7 |
| Hepatocellular carcinoma derived from hepatocellular adenoma [95,96] | 12 (25) | 48.0 (43.8–55.6) |
| Fibrolamellar hepatocellular carcinoma [49,97] | 1 (20) | 5.0 (0.0–12.5) |
| Hepatocellular carcinomas e [45,49,75,96,97,98,99,100,101,102,103,104,105,106,107,108,109] | 1263 (3093) | 40.8 (26.3–63.3) |
| Gallbladder carcinoma, NOS [49,75,110] | 15 (166) | 9.0 (0.0–50.0) |
| <5%: oesophageal squamous cell carcinoma (n = 313) [111], oesophageal adenocarcinoma, NOS (n = 90) [112], gastric moderately differentiated adenocarcinoma, NOS (n = 29) [113], moderately to poorly differentiated gastric adenocarcinoma, NOS (n = 39) [113], poorly differentiated gastric adenocarcinoma, NOS (n = 119) [113], gastric mucous adenocarcinoma (n = 13) [113], well-differentiated gastric cancer, NOS (n = 90) [110], poorly-differentiated gastric cancer, NOS (n = 178) [110], gastric carcinomas of both intestinal and diffuse types, NOS (n = 74) [75], colorectal adenocarcinoma, NOS (n = 22) [49], hepatoblastoma (n = 18) [49,114], hepatocellular adenoma, NOS (n = 238) [95,97], intrahepatic cholangiocarcinoma (n = 65) [97,99]), extrahepatic cholangiocarcinoma (n = 6) [75], extrahepatic cholangiosarcoma (n = 28) [49], pancreatic ductal adenocarcinoma (n = 24) [49], pancreatic acinar cell carcinoma (n = 25) [49] and pancreatic cancer, NOS (n = 46) [49]. | ||
| Tumours of the endocrine organs | ||
| Benign thyroid tumour, atypical follicular adenoma * [115] | 3 (18) | 16.7 |
| Papillary thyroid carcinoma, conventional variant [51,116,117,118,119,120,121,122,123,124,125] | 178 (1838) | 9.7 (2.9–15.2) |
| Papillary thyroid carcinoma, follicular variant [51,116,117,118,120,122,123,124,125] | 27 (481) | 5.6 (0.0–13.3) |
| Papillary thyroid carcinoma, tall cell variant [116,117,118,120,124,125,126] | 40 (214) | 18.7 (0.0–60.0) |
| Papillary thyroid carcinoma, NOS [51,118,119,120,123,125,127,128,129,130,131,132,133,134,135,136,137,138,139] | 361 (3288) | 11.0 (0.0–40.0) |
| Follicular thyroid carcinoma [51,115,116,119,120,122,125,128,129,132,135,138,139] | 102 (564) | 18.1 (13.9–50.0) |
| Hürthle cell carcinoma f [120,127,140] | 11 (146) | 7.5 (4.8–33.3) |
| Poorly differentiated thyroid carcinoma [51,116,120,122,125,127,133,135] | 88 (217) | 40.6 (21.4–51.7) |
| Anaplastic thyroid carcinoma [51,116,122,125,127,129,133,135,141,142] | 150 (326) | 46.0 (12.5–81.5) |
| Metastases of well-differentiated papillary and follicular thyroid carcinomas, NOS [118,125,138,143] | 90 (437) | 20.6 (15.0–50.0) |
| Adrenal cortical carcinoma [144] | 4 (34) | 11.8 |
| Extra-adrenal paraganglioma g [144] | 1 (9) | 11.1 |
| <5%: pituitary adenoma (n = 15) [145], follicular thyroid adenoma (n = 263) [51,115,122,128,139], benign thyroid tumour, NOS (n = 44) [146], papillary thyroid carcinoma, hobnail variant (n = 10) [147], papillary microcarcinoma (n = 435) [148,149], paediatric papillary carcinoma (n = 105) [150,151], minimally invasive Hürthle cell tumour (n = 52) [127,140], medullary thyroid carcinoma (n = 135) [49,51,116,119,129], adrenal cortical adenoma (n = 47) [144], phaechromocytoma (n = 122) [51,144], extra-adrenal benign paraganglioma * (n = 4) [144], pancreatic neuroendocrine tumour, NOS (n = 123) [49,57]. | ||
| Tumours of the female reproductive organs | ||
| Ovarian low-grade serous carcinoma [49,152] | 2 (41) | 5.0 (3.0–12.5) |
| Ovarian clear cell carcinoma [49,152,153] | 48 (301) | 15.9 (15.9–16.7) |
| Uterine endometrial carcinoma, NOS [49] | 2 (19) | 10.5 |
| Uterine clear cell carcinoma, NOS [153] | 3 (14) | 21.4 |
| Cervical squamous cell carcinoma [49,152,154,155] | 33 (225) | 14.7 (0.0–21.4) |
| Vulvar melanoma [156] | 2 (23) | 8.7 |
| <5%: high-grade ovarian serous carcinoma (n = 80) [152], ovarian endometrioid carcinoma (n = 43) [152], uterine endometrioid carcinoma (n = 24) [152], uterine leiomyosarcoma (n = 22) [152], uterine serous carcinoma (n = 12) [152], cervical adenocarcinoma, NOS (n = 25) [152] and vulvar melanoma (n = 23) [156]. | ||
| Tumours of the head and neck | ||
| Laryngeal carcinoma, NOS [157] | 64 (235) | 27.2 |
| Oral squamous cell carcinoma h [49,154,155,158] | 146 (295) | 49.5 (2.4–67.7) |
| Tongue squamous cell carcinoma [49] | 10 (28) | 35.7 |
| <5%: olfactory neuroblastoma (n = 11) [49] and salivary gland tumours (adenoid cystic carcinoma (n = 1) [159], adenocarcinoma, NOS (n = 1) [159], salivary duct carcinoma, NOS (n = 1) [159], epithelial-myoepithelial carcinoma (n = 1) [159], adenosquamous carcinoma (n = 1) [159], pleomorphic adenoma (n = 19) [159], basal cell adenoma (n = 1) [159], Warthin tumour (n = 8) [159]). | ||
| Tumours of haematopoietic and lymphoid tissues | ||
| Mantle cell lymphoma [160,161] | 8 (36) | 22.2 (0.0–33.3) |
| <5%: chronic myeloid leukaemia, NOS (n = 6) [49], acute myeloid leukaemia, NOS (n = 274) [49,162], B-cell acute lymphoblastic leukaemia, NOS (n = 12) [160], chronic lymphocytic leukaemia (n = 26) [49,160], marginal zone lymphoma (n = 16) [160], lymphoplasmacytic lymphoma (n = 7) [160], myeloma/plasmacytoma (n = 9) [160], follicular lymphoma (n = 13) [160], diffuse large B-cell lymphoma (DLBCL), NOS (n = 9) [160], plasmablastic lymphoma (n = 7) [160] and Burkitt lymphoma (n = 9) [160]. | ||
| Tumours of the lung, pleura and thymus | ||
| Epithelioid mesothelioma [163] | 10 (95) | 10.5 |
| Sarcomatoid, desmoplastic, and biphasic mesothelioma [163] | 6 (15) | 15.2 |
| <5%: lung adenocarcinoma, NOS (n = 533) [164,165,166,167], lung squamous cell carcinoma, NOS (n = 384) [164,165,166,167,168], lung adenosquamous carcinoma (n = 44) [154,164,165], mesothelioma, NOS (n = 4) [49], thymoma, NOS (n = 47) [75], type C thymic cancer * (n = 5) [75] and thymic neuroendocrine atypical carcinoma (n = 2) [75]. | ||
| Tumours of the peripheral nervous system | ||
| Absence of histotypes with a frequency of TERT promoter mutations equal or higher than 5%. | ||
| <5%: neuroblastoma, NOS (n = 153) [49,169]. | ||
| Tumours of the skin | ||
| Basal cell carcinoma [170,171,172,173] | 143 (292) | 49.0 (38.8–81.8) |
| Squamous cell carcinoma [49,154,170,172,173] | 57 (102) | 55.9 (20.0–74.1) |
| Bowen disease [172,173] | 2 (13) | 15.4 (9.1–50.0) |
| Superficial spreading melanoma [51,92,171,174,175] | 129 (380) | 33.9 (23.0–40.9) |
| Nodular melanoma [51,92,171,174,175] | 101 (184) | 54.9 (42.9–85.7) |
| Lentigo maligna [51,171,175] | 10 (41) | 24.4 (0.0–52.9) |
| Acral-lentiginous melanoma [51,92,171,174,175,176,177,178,179] | 40 (452) | 8.9 (0.0–27.0) |
| Desmoplastic melanoma i [180,181] | 43 (96) | 44.8 (22.9–85.0) |
| Cutaneous melanoma j [45,51,92,156,171,174,175,176,177,178,179,180,181,182,183,184,185,186,187,188,189,190] | 826 (1975) | 41.8 (7.1–85.0) |
| Metastatic melanoma of unknown primary site k [178,179,182] | 36 (73) | 49.3 (25.0–66.7) |
| Metastatic cutaneous melanoma l [185,188] | 168 (221) | 76.0 (58.8–81.2) |
| Metastatic melanoma of other primary locations m [188] | 7 (13) | 53.8 |
| Mucosal melanoma, NOS [178,179,182] | 21 (211) | 10.0 (8.7–13.2) |
| Atypical fibroxanthoma [191] | 25 (27) | 92.6 |
| Pleomorphic dermal sarcoma ** [191] | 26 (34) | 76.5 |
| Merkel cell carcinoma [192,193] | 6 (63) | 9.5 (6.7–10.4) |
| <5%: cutaneous naevi (n = 9) [51]. | ||
| Tumours of the soft tissues and bone | ||
| Myxoid liposarcoma [49,53,194] | 51 (76) | 67.1 (23.1–79.2) |
| Solitary fibrous tumour [49,53,194,195] | 37 (175) | 21.1 (12.5–27.7) |
| Fibrosarcoma [49] | 1 (3) | 33.3 |
| Malignant peripheral nerve sheath tumours [49,53,194,196] | 11 (139) | 7.9 (0.0–9.6) |
| Malignant granular cell tumour [53] | 1 (2) | 50.0 |
| <5%: lipoma (n = 8) [49], atypical lipomatous tumour (n = 10) [49], dedifferentiated liposarcoma (n = 61) [194], pleomorphic liposarcoma (n = 15) [194], well differentiated liposarcoma (n = 18) [53], dermatofibrosarcoma protuberans (n = 16) [53,194], myxofibrosarcoma (n = 33) [49,53,194], low-grade fibromyxoid sarcoma (n = 18) [49,194], gastrointestinal leiomyoma (n = 5) [75], leiomyosarcoma (n = 49) [49,53,194] pleomorphic leiomyosarcoma (n = 5) [53], rhabdomyosarcoma, NOS (n = 5) [53], embryonal rhabdomyosarcoma (n = 8) [49], alveolar rhabdomyosarcoma (n = 7) [49], angiosarcoma of soft tissue (n = 9) [194], gastrointestinal stromal tumour (n = 251) [49,51,75,197], gastric schwannoma (n = 1) [75], synovial sarcoma (n = 48) [49,53,194], epithelioid sarcoma (n = 4) [194], alveolar soft part sarcoma (n = 9) [53,194], clear cell sarcoma of soft tissue (n = 6) [53,194], extraskeletal myxoid chondrosarcoma (n = 12) [49,53,194], chondrosarcoma, NOS (n = 21) [49,53], Ewing sarcoma (n = 6) [53] and undifferentiated high-grade pleomorphic sarcoma (n = 70) [49,50,53]. | ||
| Tumours of the urinary system and male genital organs | ||
| Clear cell renal cell carcinoma [198,199,200] | 39 (443) | 8.8 (0.0–12.2) |
| Chromophobe renal cell carcinoma [51,200] | 1 (12) | 8.3 (0–12.5) |
| Urothelial muscle invasive bladder carcinoma [201,202,203,204,205,206,207] | 463 (679) | 68.2 (48.8–85.2) |
| Urothelial non-muscle invasive bladder carcinoma [51,201,202,203,204,205,206,208,209] | 959 (1395) | 68.7 (44.3–85.4) |
| Urothelial bladder carcinoma [45,49,70,75,204,207,210,211] | 377 (591) | 63.8 (46.4–84.6) |
| Squamous cell carcinoma of the bladder [168,211,212] | 52 (79) | 65.8 (54.7–100.0) |
| Bladder adenocarcinoma n [210,213] | 4 (54) | 7.4 (0.0–28.6) |
| Papillary urothelial neoplasm of low malignant potential [204,214] | 26 (43) | 60.5 (28.6–75.9) |
| Urothelial carcinoma of upper urinary tract o [49,75,200,201,207,215,216] | 146 (293) | 49.8 (35.3–76.9) |
| Urothelial carcinoma of the ureter [200,214,215] | 26 (135) | 19.3 (11.1–50.0) |
| Micropapillary urothelial carcinoma p [217] | 33 (33) | 100.0 |
| Urothelial carcinoma q [154,218] | 71 (96) | 75.9 (70.0–100) |
| <5%: papillary renal cell carcinoma (n = 10) [51], nephrogenic adenoma (n = 10) [210], prostate squamous cell carcinoma (n = 26) [219], prostate carcinoma, NOS (n = 47) [49,201], ‘testicular carcinoma’ ***, NOS (n = 17) [201]. | ||
| TERT Amplifications | ||
|---|---|---|
| Tumour Type | Amplified Cases (Total) | Amplified Cases in % (Range) |
| Tumours of the breast | ||
| Breast carcinoma, NOS a [52] | 5 (19) | 26.3 |
| <5%: phyllodes tumour (n = 73) [223]. | ||
| Tumours of the central nervous system | ||
| Pineoblastoma [224] | 1 (1) | 100 |
| Classic medulloblastoma [224] | 5 (13) | 38.5 |
| Nodular medulloblastoma [224] | 1 (10) | 10.0 |
| Anaplastic medulloblastoma [224] | 2 (5) | 40.0 |
| Medulloepithelyoma [224] | 2 (2) | 100.0 |
| Medullomyoepithelyoma [224] | 1 (2) | 50.0 |
| Ewing sarcoma/peripheral primitive neuroectodermal tumour [224] | 4 (8) | 50.0 |
| <5%: medullomyoblastoma (n = 1) [224]. | ||
| Tumours of the digestive system | ||
| Colorectal carcinoma, NOS [225] | 31 (64) | 48.4 |
| Hepatocellular carcinoma, NOS b [99,103,226] | 27 (181) | 14.9 (3.4–72.2) |
| Tumours of the endocrine system | ||
| Adrenal cortical carcinoma [45] | 11 (75) | 14.7 |
| Tumours of the female reproductive organs | ||
| Ovarian serous cystadenocarcinoma [45] | 6 (27) | 22.2 |
| Cervical intraepithelial neoplasia (CIN) 2 [227] | 6 (10) | 60 |
| Cervical intraepithelial neoplasia (CIN) 3 [227] | 7 (8) | 87.5 |
| Cervical carcinoma, NOS c [52,227] | 7 (14) | 50.0 |
| <5%: cervical intraepithelial neoplasia (CIN) 1 (n = 5) [227]. | ||
| Tumours of the head and neck | ||
| Pharyngeal/laryngeal squamous cell carcinoma, NOS [228] | 8 (81) | 9.9 |
| <5%: oral squamous cell carcinoma (n = 191) [228]. | ||
| Tumours of the lung | ||
| Lung carcinoma, NOS d [52] | 8 (21) | 38.1 |
| Lung adenocarcinoma, NOS [45,229,230] | 97 (529) | 18.3 (13.2–75.0) |
| Lung squamous cell carcinoma, NOS [45,229,230] | 59 (240) | 24.6 (13.8–63.9) |
| Lung large cell carcinoma [230] | 2 (5) | 40.0 |
| Mixed histology lung tumours [230] | 1 (5) | 20.0 |
| Tumours of the peripheral nervous system | ||
| Neuroblastoma, NOS [52] | 1 (8) | 12.5 |
| Tumours of the skin | ||
| Acral-lentiginous melanoma [231,232] | 14 (60) | 23.3 (20.1–29.4) |
| Melanoma e [233] | 4 (10) | 40.0 |
| Merkel cell carcinoma [192] | 11 (14) | 78.6 |
| <5%: desmoplastic melanoma (n = 62) [181]. | ||
| Tumours of the urinary system | ||
| Urothelial invasive bladder carcinoma [234] | 2 (3) | 66.7 |
| <5%: urothelial non-invasive bladder cancer (n = 15) [234]. | ||
| TERC amplifications | ||
| Tumours of the digestive system | ||
| Oesophageal carcinoma, NOS [45] | 36 (168) | 21.4 |
| Tumours of the female reproductive organs | ||
| Ovarian carcinoma f [45,235] Cervical intraepithelial neoplasia (CIN) 1 [227,236,237] Cervical intraepithelial neoplasia (CIN) 2 [227,236,237] Cervical intraepithelial neoplasia (CIN) 3 [227,236,237] Cervical carcinoma g [227,236,237,238,239,240] | 13 (35) 10 (41) 29 (44) 51 (58) 53 (90) | 37.1 (22.2–87.5) 24.4 (21.1–40.0) 69.0 (50.0–90.0) 87.9 (81.5–100.0) 58.9 (6.1–100.00) |
| Tumours of head and neck | ||
| Absence of histotypes with a frequency of TERC amplifications equal or higher than 5%. | ||
| <5%: squamous cell carcinoma of the head and neck, site not-specified (n = 31) [238]. | ||
| Tumours of the lung | ||
| Lung carcinoma, NOS [238] Lung squamous cell carcinoma, NOS [45] | 1 (9) 68 (167) | 11.1 40.7 |
| TERT Polymorphism | Cancers with Higher Risk of Development |
| rs2736100 | Acute lymphoblastic leukaemia (paediatric); myeloproliferative neoplasms; bladder, cervical, colorectal, gastric, lung and pancreas (exocrine) cancers, NOS; gliomas; oral squamous cell and papillary thyroid carcinomas [241,242,243,244,245,246,247,248,249,250,251,252,253,254,255,256,257,258,259,260,261,262,263,264,265,266,267,268,269,270] |
| rs2736098 | Bladder, breast, cervical, colorectal, lung, pancreas (exocrine) and prostate cancers, NOS; basal cell, hepatocellular and nasopharyngeal carcinomas, NOS [252,270,271,272,273,274,275,276,277] |
| rs2853676 | Breast, gastric, lung, prostate and ovary cancers, NOS; gliomas, NOS; melanomas, NOS [243,247,278,279,280,281,282,283,284,285,286,287] |
| TERT Promoter Polymorphism | Cancer Types with Higher Risk of Development |
| rs2853669 | Bladder, breast, gastric, lung and prostate cancers, NOS; gliomas, NOS; hepatocellular carcinomas, NOS; melanomas, NOS [68,69,76,80,162,188,222,265,275,288,289,290,291,292,293] |
| rs2735940 | Acute lymphoblastic leukaemia (paediatric); gastric and lung cancers, NOS [256,265,294] |
| Tumour Type | Positive Cases (Total) | Positive Cases in % (Range) |
|---|---|---|
| Tumours of the breast | ||
| Absence of histotypes with a frequency of ALT equal or higher than 5%. | ||
| <5%: invasive lobular carcinoma (n = 27) [55], tubular carcinoma (n = 9) [55], carcinoma with medullary features (n = 55) [55], mucinous carcinoma (n = 15) [55] and ductal carcinoma (n = 271) [55]. | ||
| Tumours of the central nervous system | ||
| Diffuse astrocytoma, NOS [55,295,296] | 22 (42) | 52.4 (27.3–63.0) |
| Anaplastic astrocytoma, NOS [55,295,296,297,298] | 63 (143) | 53.8 (26.4–100.0) |
| Astrocytoma, NOS [299] | 17 (50) | 34.0 |
| Anaplastic paediatric astrocytoma, NOS [296] | 26 (88) | 29.5 |
| Paediatric glioblastoma, NOS [55,300] | 17 (57) | 29.8 (12.0–43.8) |
| Glioblastoma, NOS a [55,295,297,298,301,302,303] | 147 (953) | 15.4 (11.4–50.0) |
| Oligodendroglioma, NOS [55] | 8 (40) | 20.0 |
| Anaplastic pleomorphic xanthoastrocytoma [295] | 2 (2) | 100.0 |
| Choroid plexus carcinoma b [300] | 7 (31) | 22.6 |
| Ewing sarcoma/peripheral primitive neuroectodermal tumour b [300] | 5 (43) | 11.6 |
| <5%: paediatric anaplastic astrocytoma (n = 24) [296], pilocytic astrocytoma (n = 45) [295,300], pleomorphic xanthoastrocytoma (n = 8) [300], ependymoma (n = 95) [300], choroid plexus papilloma (n = 24) [300], ganglioglioma (n = 8) [300], medulloblastoma, NOS [55,300], atypical teratoid/rhabdoid tumour (n = 38) [300], schwannoma (n = 44) [55], meningioma (n = 46) [55]. | ||
| Tumours of the digestive system | ||
| Hepatocellular carcinoma, NOS [55] | 8 (121) | 6.6 |
| Chromophobe hepatocellular carcinoma with abrupt anaplasia [304] | 11 (12) | 91.7 |
| Gastric adenocarcinoma, NOS [55,305] | 16 (197) | 8.1 (0.0–38.1) |
| <5%: oesophageal adenocarcinoma (n = 106) [55], oesophageal small cell neuroendocrine carcinoma (n = 1) [55], oesophageal squamous cell carcinoma (n = 29) [55], adenocarcinoma of the small intestine, NOS (n = 215) [55], colon adenoma, NOS (n = 136) [55], colon adenocarcinoma, NOS (n = 126) [55], hepatocellular adenoma, NOS (n = 17) [55], extrahepatic cholangiocarcinoma (n = 33) [55], adenocarcinoma of the gallbladder, NOS (n = 60) [55], pancreatic ductal adenocarcinoma (n = 448) [55], gastrointestinal carcinoid tumours, NOS/SNS (n = 47) [306] and carcinoid tumour, NOS/SNS (n = 32) [55]. | ||
| Tumours of the endocrine organs | ||
| Medullary thyroid carcinoma [307] | 11 (42) | 26.2 |
| Adrenal cortical carcinoma [308] | 3 (24) | 12.5 |
| Extra-adrenal paranganglioma [55] | 1 (8) | 12.5 |
| Pancreatic neuroendocrine tumour, NOS [306,309,310,311,312,313,314] | 255 (849) | 30.0 (14.9–61.0) |
| <5%: thyroid adenoma (n = 34) [55], papillary thyroid carcinoma, NOS (n = 47) [55,299], follicular thyroid carcinoma (n = 52) [55], parathyroid adenoma (n = 38) [55], adrenal adenoma (n = 14) [55], and phaeochromocytoma (n = 67) [55]. | ||
| Tumours of the female reproductive system | ||
| Diffuse malignant peritoneal mesothelioma c [315] | 10 (38) | 26.3 |
| Uterine carcinosarcoma [316] | 8 (16) | 50.0 |
| Uterine leiomyosarcoma [316] | 7 (8) | 87.5 |
| Uterine serous carcinoma [55] | 3 (41) | 7.3 |
| Uterine stromal sarcoma [316] | 4 (17) | 23.5 |
| <5%: ovarian serous carcinoma, NOS (n = 205) [55], ovarian mucinous carcinoma (n = 21) [55], ovarian endometrioid carcinoma (n = 72) [55], ovarian clear cell carcinoma (n = 56) [55], uterine endometrioid carcinoma (n = 64) [55], uterine clear cell carcinoma (n = 3) [55], uterine mixed mesodermal tumour (n = 4) [55], cervical squamous cell carcinoma (n = 152) [55] and cervical adenocarcinoma, NOS (n = 19) [55]. | ||
| Tumours of the head and neck | ||
| Absence of histotypes with a frequency of ALT equal or higher than 5%. | ||
| <5%: laryngeal squamous cell carcinoma (n = 29) [55], oral squamous cell carcinoma, NOS (n = 41) [55], salivary gland cylindroma (n = 28) [55], salivary gland carcinoma, NOS (n = 98) [55], pleomorphic adenoma (n = 45) [55] and Warthin tumour (n = 23) [55]. | ||
| Tumours of haematopoietic and lymphoid tissues | ||
| Absence of histotypes with a frequency of ALT equal or higher than 5%. | ||
| <5%: diffuse large B-cell lymphoma (DLBCL), NOS (n = 10) [55], nodular sclerosis classic Hodgkin lymphoma (n = 23) [55], mixed-cellularity classic Hodgkin lymphoma (n = 17) [55] and other subtypes of non-Hodgkin lymphoma, NOS (n = 54) [55]. | ||
| Tumours of the lung, pleura, thymus and heart | ||
| Absence of histotypes with a frequency of ALT equal or higher than 5%. | ||
| <5%: lung tumours (adenocarcinoma, NOS (n = 153) [55], papillary carcinoma (n = 15) [55], bronchoalveolar carcinoma, NOS (n = 40) [55], squamous cell carcinoma (n = 100) [55], small cell neuroendocrine carcinoma (n = 63) [55], carcinoid tumour, NOS (n = 3) [55], large cell carcinoma (n = 35) [55], other hystotypes, NOS (n = 15) [55] and thymoma, NOS (n = 37) [55]. | ||
| Tumours of the peripheral nervous system | ||
| Neuroblastoma, NOS [55,317] | 62 (124) | 50.0 (9.1–58.8) |
| <5%: ganglioneuroma (n = 3) [55]. | ||
| Tumours of the skin | ||
| Malignant melanoma, NOS [55] | 7 (106) | 6.6 |
| <5%: basal cell carcinoma (n = 57) [55], squamous cell carcinoma (n = 56) [55], benign naevus, NOS (n = 12) [55] and benign adnexal tumour (n = 15) [55]. | ||
| Tumours of the soft tissues | ||
| Liposarcoma, NOS [55,299,318] | 21 (84) | 25.0 (23.5–33.3) |
| Dedifferentiated liposarcoma [319,320,321] | 35 (106) | 33.0 (26.9–47.6) |
| Myxoid liposarcoma d [319,320,321] | 20 (107) | 18.5 (5.0–30.0) |
| Pleomorphic liposarcoma [320] | 21 (26) | 80.8 (72.7–100) |
| Fibrosarcoma and variants [55,299] | 5 (23) | 21.7 (14.3–100.0) |
| Myxofibrosarcoma [322] | 19 (25) | 76.0 |
| Leiomyosarcoma [55,299,323] | 91 (161) | 56.5 (33.3–61.5) |
| Rhabdomyosarcoma, NOS [55,299] | 2 (39) | 5.1 (0.0–5.7) |
| Embryonal rhabdomyosarcoma [322,324] | 7 (24) | 29.2 (12.5–37.5) |
| Epithelioid haemangioendothelioma [325] | 1 (7) | 14.3 |
| Angiosarcoma [55,325] | 18 (79) | 22.8 (11.1–24.3) |
| Neurofibroma [55] | 2 (2) | 100.0 |
| Malignant peripheral nerve sheath tumours [55,320,326] | 47 (75) | 62.7 (0.0–79.2) |
| Epithelioid sarcoma [55,299,322] | 2 (14) | 14.3 (0.0–33.3) |
| Alveolar soft part sarcoma [299,322] | 1 (8) | 12.5 (0.0–25.0) |
| Chondrosarcoma, NOS [55,299] | 1 (7) | 14.3 (0.0–33.3) |
| Osteosarcoma e [299,327,328] | 109 (173) | 63.0 (46.6–79.6) |
| Malignant fibrous histiocytic tumour * [299,329] | 39 (75) | 52.0 (32.6–80.0) |
| Undifferentiated pleomorphic sarcoma f [55,322] | 55 (86) | 64.0 (63.5–64.7) |
| Radiation-associated sarcoma, NOS [322] | 3 (15) | 20.0 |
| <5%: lipoma (n = 1) [55], solitary fibrous tumours (n = 7), dermatofibrosarcoma protuberans (n = 9) [322], benign fibrous hystiocytoma (n = 16) [55], giant cell tumour of the tendon sheath (n = 22) [55], alveolar rhabdomyosarcoma (n = 23) [322,324], capillary haemangioma (n = 32) [55], Kaposi’s sarcoma (n = 55) [55], gastrointestinal stromal tumours (n = 50) [55,322], Ewing sarcoma (n = 63) [55,322,327], synovial sarcoma (n = 24) [299,322], clear cell sarcoma (n = 5) [55], extraskeletal myxoid chondrosarcoma (n = 2) [322]. | ||
| Tumours of the urinary system and male genital system | ||
| Chromophobe renal cell carcinoma [55] | 4 (47) | 8.5 |
| Nephroblastoma [330] | 26 (32) | 81.3 |
| Sarcomatoid renal carcinoma [55] | 2 (27) | 7.4 |
| Bladder small cell neuroendocrine carcinoma [55] | 3 (13) | 23.1 |
| Non-seminomatous germ cell tumour, NOS [55] | 7 (46) | 15.2 |
| <5%: clear cell renal cell carcinoma (n = 117) [55], papillary renal cell carcinoma (n = 86) [55], renal oncocytoma (n = 18) [55], urothelial muscle invasive (n = 150) [55] and non-muscle invasive bladder carcinoma (n = 38) [55], non-invasive papillary urothelial carcinoma (n = 5) [55], sarcomatoid bladder carcinoma (n = 1) [55], squamous cell carcinoma of the bladder (n = 2) [55], prostate adenocarcinoma (n = 1152) [55], prostate small cell neuroendocrine carcinoma (n = 24) [55] and seminoma (n = 48) [55]. | ||
| Hepatocellular Carcinomas |
| TERTp mutations: marker of malignant progression [95,331,332]. |
| Well-differentiated thyroid carcinomas |
| TERTp mutations: association with larger tumours, older patient age, higher tumour stage, tumour recurrence, and distant metastases [51,116,119,126,333]; association with BRAFV600E mutations [117,126,141,146]. |
| Pancreatic neuroendocrine tumours |
| TERTp mutations: association with hereditary syndromes [57]. |
| ALT: association with protein loss and mutation of ATRX/DAXX [311,312,314]; indicator of more aggressive disease, metastases and worse patient survival [309,311,312,334]. |
| Diffuse astrocytic and oligodendroglial tumours |
| TERTp mutations: correlation with older patient age, higher tumour grade, tumour progression and worse overall survival [62,65,80,98]; combination of TERTp and IDH mutations for survival assessment: TERTp and IDH concomitant mutations confer the longest overall patient survival; TERTp mutations alone confer the lowest [61,71,73,74,78,79]. |
| TERT and TERTp polymorphisms: rs2736100 [335] and rs2853676 [336] affect the risk of tumour development; association of TERTp rs2853669 status with worse prognosis and worse survival in GB patients [68,76,80]; modifying effect of rs2853669 on TERTp mutations [65,68,76,80]. |
| ALT: association with ATRX loss [295,296,298,337]; identification of less aggressive GBs with longer patient survival [298,301,303]; mutual exclusivity with TERTp mutations [49,50]. |
| Epigenetic mechanisms: methylation of TERT promoter as a potential biomarker for malignancy in paediatric gliomas [338]. |
| Neuroblastomas |
| TERT rearrangements: indicator of poor prognosis, particularly in combination with MYCN amplification [54,339,340,341]. |
| ALT: association with chemoresistant tumours with unfavourable prognosis [342]. |
| Uterine cervical lesions |
| TERT and TERC amplifications: early identification of patients with low-grade lesions and higher progression risk in routinely liquid based cytology by Pap smears [227,236,237]. |
| Cutaneous melanomas |
| TERTp mutations: association with male gender, older patient age, tumour ulceration, higher Breslow’s thickness, and worse overall survival [92,175,179,182,184,221]; association with BRAFV600E mutations; combination used to identify tumours with aggressive behaviour [175,179,186]. |
| TERT and TERTp polymorphisms: association of TERTp polymorphism rs2853669 with TERTp mutations identify patients at risk of aggressive disease [175,188,293]. |
| Liposarcomas |
| ALT: association with ATRX loss, disease progression and poor clinical outcome [319,320]. |
| Urothelial bladder carcinomas |
| TERTp mutations: association with increased disease recurrence and reduced patient survival [209,215,222]; useful biomarker for patient screening as a non-invasive diagnostic and follow-up tool [206,209]; combination with FGFR3 mutations to identify tumours with poor prognosis [222]. |
| TERT amplification: potential biomarker to identify high-risk patients with disease progression [234]. |
| TERT and TERTp polymorphisms: association of TERTp rs2853669 with tumour recurrence and worse patient survival [204]. |
| TERT Promoter Mutations, % (Total) | TERT and TERC Amplifications % (Total) | ALT, % (Total) | |
|---|---|---|---|
| Tumours of the breast | |||
| 0 (210) | 26 (19) | 0–3.7 (377) | |
| Tumours of the digestive system | |||
| Stomach | 0 (543) | N.A. | 0–8 (197) |
| Small intestine | N.A. | N.A. | 0 (215) |
| Colon and rectum | 0 (22) | 48 (64) | 0 (126) |
| Exocrine pancreas | 0 (95) | N.A. | 0 (448) |
| Tumours of haematopoietic and lymphoid tissues | |||
| 2 (424) | N.A. | 0 (104) | |
| Tumours of the lung | |||
| 1 (611) | 9 (976) | 0 (424) | |
| Tumours of the male genital organs | |||
| Prostate | 0 (99) | N.A. | 0 (1176) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gaspar, T.B.; Sá, A.; Lopes, J.M.; Sobrinho-Simões, M.; Soares, P.; Vinagre, J. Telomere Maintenance Mechanisms in Cancer. Genes 2018, 9, 241. https://doi.org/10.3390/genes9050241
Gaspar TB, Sá A, Lopes JM, Sobrinho-Simões M, Soares P, Vinagre J. Telomere Maintenance Mechanisms in Cancer. Genes. 2018; 9(5):241. https://doi.org/10.3390/genes9050241
Chicago/Turabian StyleGaspar, Tiago Bordeira, Ana Sá, José Manuel Lopes, Manuel Sobrinho-Simões, Paula Soares, and João Vinagre. 2018. "Telomere Maintenance Mechanisms in Cancer" Genes 9, no. 5: 241. https://doi.org/10.3390/genes9050241
APA StyleGaspar, T. B., Sá, A., Lopes, J. M., Sobrinho-Simões, M., Soares, P., & Vinagre, J. (2018). Telomere Maintenance Mechanisms in Cancer. Genes, 9(5), 241. https://doi.org/10.3390/genes9050241