
Zooplankton Temporal, Longitudinal, and Vertical Diversity Patterns in the Floodplains of the Western Amazon

Grupo de Investigación en Biodiversidad, Medio Ambiente y Salud (BIOMAS), Universidad de Las Américas, Quito 170124, Ecuador
*
Author to whom correspondence should be addressed.
Water 2024, 16(8), 1166; https://doi.org/10.3390/w16081166
Submission received: 21 February 2024
/
Revised: 27 March 2024
/
Accepted: 28 March 2024
/
Published: 20 April 2024
(This article belongs to the Special Issue Biology and Ecology of Zooplankton: Latest Advances and Prospects)
Abstract
:The Western Amazon is a highly biodiverse area. Zooplankton diversity studies in the region have been primarily conducted in Peru and Colombia, with limited research in the Ecuadorian Amazon. To address this gap, our research aimed to enhance taxonomic knowledge and understand zooplankton diversity patterns in the Napo and Pastaza lower basins at different spatial and temporal scales. Two sampling expeditions were conducted in the high waters of 2021 and rising waters of 2022. Dry conditions in 2021 led to lower-than-expected water levels. The study identified 107 zooplankton species, revealing variations in richness and composition between years, lakes, and depth strata. Grande Lake, deeper and wider than Delfincocha, exhibited significant turnover variations across strata in both seasons. Despite a relative longitudinal homogenization between channels and floodplains during high waters, beta diversity across vertical and temporal gradients highlighted complex dynamic zooplankton communities in both lakes. In addition, we include the first records of 44 taxa for the Ecuadorian Amazon and 36 for Ecuador. These findings emphasize the need for targeted research and conservation efforts in the face of escalating environmental threats to the Western Amazon.
1. Introduction
Freshwater biodiversity is driven by biotic process that include interspecific competition and predation, as well as abiotic factors such as environmental variability, water chemical conditions and productivity, and habitat type, size, and location within the river network [1,2]. Neotropical freshwater ecosystems are characterized by a high diversity of zooplankton, but this diversity is higher within the Amazon drainage [3]. Tropical floodplains, due to their temporal and environmental variability, represent a good model for the study of zooplankton diversity in its scales: alpha, beta, and gamma [4]. Floods trigger a process of homogenization of aquatic habitats, as a result of a dilution of the organic matter and chemical elements, accompanied by a reduction in beta diversity [5]. On the contrary, during the season of low waters, the local environmental conditions increase their influence on community structure [5].
For aquatic ecosystems, hydrological connectivity is of utmost importance because it mediates the structure and function of aquatic ecosystems [6], as well as the structure of their biological communities [2], by reducing beta diversity and productivity [7] or increasing alpha diversity and the density of organisms [8]. Seasonal variation in environmental parameters, such as water chemistry and the physical configuration of aquatic habitats, together with changes in the distribution, structure, and composition of aquatic animals and plants, is notable and consistent for these ecosystems [6].
The Western Amazon, recognized as one of the key areas of global diversity, harbors an unparalleled wealth of aquatic life [9]. However, knowledge about zooplankton diversity remains incipient compared to what is known for other regions. In Colombia, studies of cladocerans are still in their early stages, with scarce works [10]. As for Cladocera in Ecuador, 16 species are registered in the coastal region, 22 species for the Andes, 2 species for Galápagos, and 0 species for the Amazon [11] until our study. The Cladocerans and Protozoa reported in this study are the first records for the entire Ecuadorian Amazon. Rotifera Monogononta has been better documented for the Ecuadorian Amazon compared to the rest of the zooplankton groups, with 228 species corresponding to 57 genera from 25 families [12]. Knowledge about Copepoda remains nonexistent in this region due to the lack of studies and specialists for such a complex group. In our study, we present new registers for Rotifera and Copepoda for the Ecuadorian Amazon.
In a similar fashion, most studies of zooplankton diversity in the Western Amazon have been developed in Peru [13] and Colombia [14], where research remains scarce. Consequently, the taxonomic knowledge and the diversity patterns of zooplankton from the Napo and Pastaza lower basins is currently in very short supply.
On the other hand, anthropogenic disturbances, including disruptions in connectivity, nutrient inputs, deforestation, and overexploitation of resources, cause changes in the structure and dynamics of these freshwater ecosystems and their communities, and put their long-term conservation at risk [15]. In the Western Amazon, these disturbances are frequently related to the oil and gas industry’s development [16], as well as by illegal gold mining, and agricultural and urban development [17].
Considering the above, this study aimed to assess the structure and diversity of zooplanktonic communities. We expected to find differences in community composition between rivers and lakes due to their different lotic versus lentic conditions and between hydrological seasons because of the changes in habitat connectivity, reduced water levels, and variations in physical–chemical water parameters. The exploration of these variations in zooplankton community structure could be helpful in identifying potential risks concerning global climatic change and other environmental threats. Additionally, at a much finer temporal and spatial scale, we wanted to explore the temporal changes in community composition along the water column and across hydrological seasons.
2. Materials and Methods
2.1. Study Area
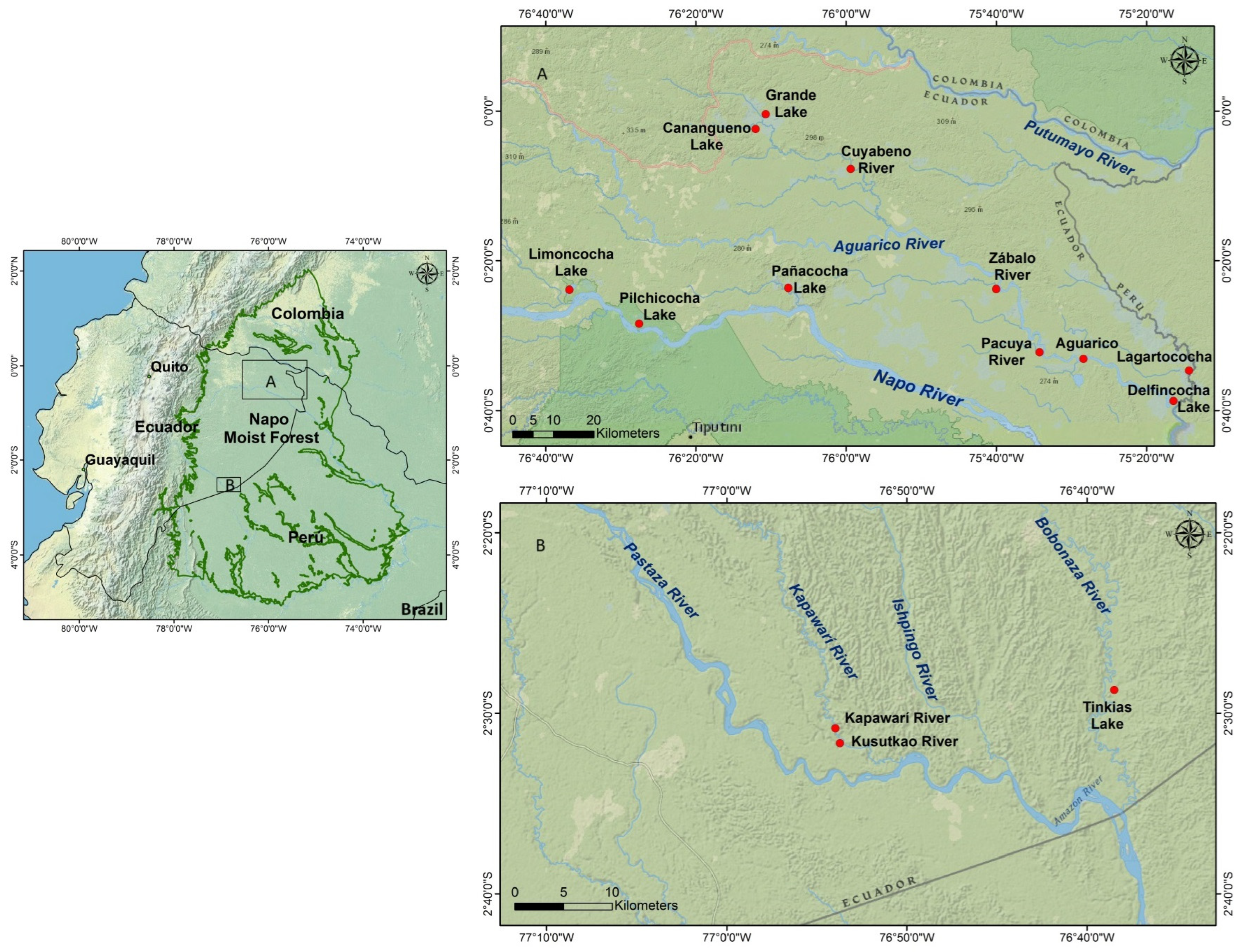
Sampling was conducted in the western Ecuadorian Amazon, within the Napo and Pastaza watersheds. Sampled water bodies within each watershed are shown in Figure 1. Both the Pastaza and the Aguarico have tributaries within the Peruvian Amazon, which, together with the rivers of the Amazon basin in northern Peru and southern Colombia, make up the Napo Moist Forest Ecoregion (Figure 1).
2.2. Sampling Protocols
Sampling journeys were conducted during June 2021 and April 2022, which corresponded to the seasons of high and rising waters, respectively. However, during 2021, conditions were particularly dry [18], and water levels were lower than expected. Zooplankton samples were collected using a 5 L van Dorn collection bottle (Royal Eijkelkamp, EN Giesbeek, The Netherlands) (collection permit No. MAAE-DBI-CM-2021-0161). For the integrated samples, 30 L of water were filtered in a 63 µm net that was collected. The integrated samples were collected at 3 sites (1 in the center and 2 at the margins of each water body) at a 50 cm depth. For the analyses of zooplankton vertical migration, we selected the Lakes Grande and Delfincocha, where 3 samples were collected in the center of the lakes at three different depths: 50 cm (surface), 150 cm (middle), and 300 cm (bottom), during a 24 h period at intervals of 3 h for each one of the depth levels. For each depth level, 10 L were filtered to collect the zooplankton. Samples were preserved using a 10% formalin solution. The identification and counting of the specimens were carried out using an Olympus CKX41 inverted microscope (Biomed Instruments, Quito, Ecuador) with magnifications between 5, 10, 20, 40× with the Utermöhl method [19]. Counts were made from subsamples ranging from 1.6 to 32 mL, depending on the densities or organisms in the samples, for all taxa. We identified the specimens using taxonomic keys and checklists of the different taxonomic groups [20,21,22,23,24], as well as general manuals [25] and specialized websites [26]. We measured total dissolved oxygen, water temperature, pH, chlorophyll, conductivity, and total dissolved solids with an In Situ Aqua Troll 500 Multiparametric Sonde (In-Situ Co., Fort Collins, CO, USA) and water depth and transparency with a Secchi disk. These water parameters are provided in Supplementary File S2.
2.3. Statistical Analyses
First, based on the integrated samples, proportions of species by clades based on total richness (Rotifera, Cladocera, CO, and Amoebozoa) were compared through a Chi2 test based on 2000 simulations. We also ran a species indicator analysis using the indicspecies package [27] of R to identify indicator species of the different water bodies. For that, we first conducted K means clustering analyses based on species abundances in each water body separately for each season.
We initially estimated alpha diversity with q0, q1, and q2 Hill numbers, which represent species richness, and diversity of rare and of common species, respectively [12], for each sampled water body and for each season separately. Then, with the combined data of the main watersheds (Aguarico, Napo, and Pastaza), using the R programming language packages hillR [28] and iNEXT [29], for these main hydrological units, rarefaction curves were plotted to assess sample completeness [29] based on density (numbers of individuals by 100 mL water). Statistical comparisons of alpha diversities between watersheds were conducted through Kruskal–Wallis tests. Comparisons of zooplankton diversity were conducted using the integrated samples’ Hill numbers q0, q1, and q2, which were used to calculate alpha, beta, and gamma diversity indices for each sampled year, 2021 and 2022. The Hill number q0 is equivalent to species richness, while q1 and q2 are equivalent to the Shannon and Simpson diversities, of which the first gives more weight to rare species and the second to the most abundant species [30,31].
Then, we assessed the effects of environmental conditions on the structure of zooplankton communities through a Redundancy Analysis (RDA) [32] based on species densities in each water body during both seasons. To select the parameters to include in the RDA, first, we calculated paired Pearson correlations between chlorophyll, dissolved oxygen, water pH, depth, total dissolved solids, conductivity, and season (Supplementary File S3).
From the Lakes Grande and Delfincocha, we took subsamples of 1.6 mL and counted the zooplankton, after which we calculated the vertical and temporal shifts of community composition at three water depth levels, 50 cm, 150 cm, and 300 cm, across the different time intervals encompassing 24 h. We examined species replacement through the turnover index, which calculates the proportion of species that differ between time points [33], and comparisons were made between seasons for each lake separately, using the codyn package in R [33]. In addition, we compared zooplankton abundance over time of the day and water depth strata with two-way ANOVAs for each lake separately for 2021 and 2022. With the turnover indices, we conducted linear models with dissolved oxygen and temperature as explanatory variables for both lakes separately with the combined data of the two seasons.
3. Results
3.1. Zooplankton Diversity Patterns in the Floodplains of the Napo and Pastaza Watersheds
Overall, we found 107 zooplanktonic species, distributed as follows: 21 Cladocerans, 43 Rotifers, 15 Copepods, and 28 Ameboidae (Table S1, Supplementary File S1). Of these, 44 are new registers for the Ecuadorian Amazon and 36 for Ecuador (Table S1). Rotifers contributed the highest proportions of species richness across sampling sites, except for Grande Lake and Kusutkao River. Within Grande Lake, Copepoda and Cladocera contributed the highest proportions of species richness (Figure 2). In contrast, these clades were absent from Kusutkao, where Amoebozoa exhibited the highest proportions of zooplankton richness (Figure 2). We found that the distribution of species richness across clades was significantly different (Xi2 = 74.69, p ˂ 0.001) among water bodies.
Regarding the composition of zooplankton species in the study area, the dominant taxa within the cladocerans were Bosminopsis deitersi, Bosminopsis sp., Daphnia cf. gessneri, and Ceriodaphnia cornuta. Among the rotifers, primarily planktonic genera were dominant, with a high abundance of Filinia longiseta, Polyarthra dolichoptera, Asplanchna sp., Brachionus zahniseri, and Keratella cf. cochlearis.
Rarefaction curves indicated that samples within the Aguarico and Napo Rivers had high completeness since they reached their asymptotes at low abundances but not for the Pastaza River since the curve did not reach an asymptote (Figure 3).
The analyses of alpha, beta, and gamma diversity showed that there was a reduction in the three scales of zooplankton diversity for the entire study area during the high waters season of 2022, which had abnormally low water depths (Table 1) although more pronounced for the gamma component. Additionally, the decreases in beta diversity during the rising waters of 2022 highlight a tendency towards homogenization of zooplankton communities at higher water levels.
The groupings made for the indicator species analyses with high waters distinguished Limoncocha and Canangueno Lakes as two different groups, 1 and 3, while the other sites were clustered in group 2. Three species were identified as indicators of these two groups combined (Table 2). For rising waters, only Zancudococha and Delfincocha Lakes were separated as different groups, 2 and 3, and the other sites were clustered in group number 1. Only one species of rotifer was established as an indicator species for these two lakes (Table 2).
Comparisons of Hill numbers between seasons through Kruskal–Wallis tests resulted in non-significant differences. This implies that the decreased zooplankton alpha diversity detected during rising waters was not of a great order of magnitude for the entire study area. During rising waters, Canangueno Lake showed the highest zooplankton richness, which also resulted in higher diversity of both rare and common species (Table 3). During the abnormal high waters season, the highest zooplankton richness was observed within Delfincocha Lake, but the highest diversities of both rare and common species was detected in the Cuyabeno River (Table 3).
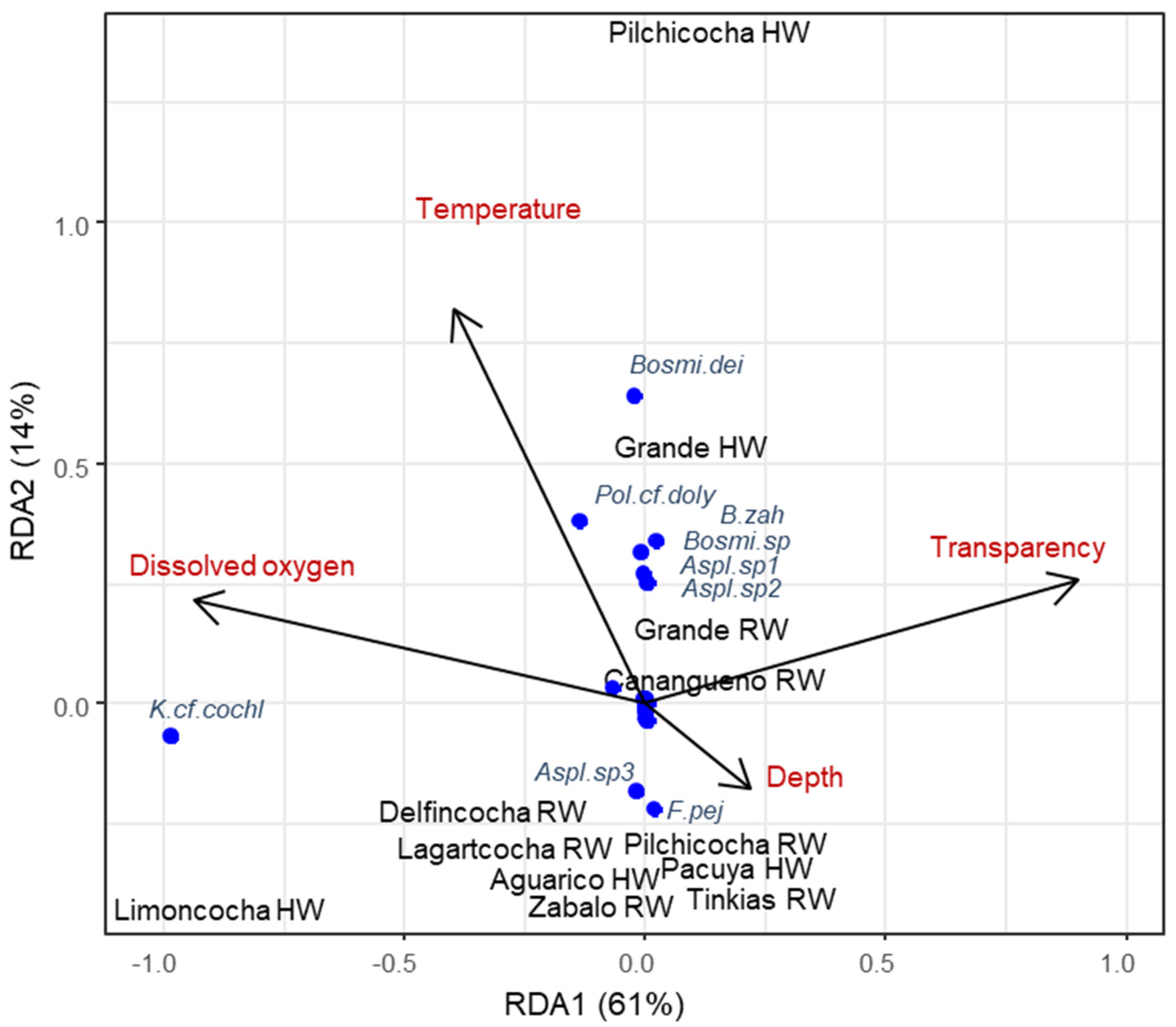
Based on the Pearson correlations (Supplementary File S3), we conducted the RDA with water temperature, dissolved oxygen, transparency, and depth as explanatory variables. The RDA indicated 0.75 constrained variance and 0.25 of unconstrained variance, and the model was significant (p = 0.049, F = 5.29, and 4 df). The first two constrained axes explained 61% and 14% of the variance, respectively. Water temperature had a significant effect (F = 3.22, p = 0.02), and dissolved oxygen had a marginally significant effect (F = 15.65, p = 0.06).
Water bodies were separated by their environmental conditions, zooplankton community structure, and by season. Limoncocha was characterized by high levels of dissolved oxygen during high waters and by high densities of Keratella cf. cochlearis (Figure 4). Canangueno Lake showed high water transparency during rising waters. The majority of water bodies, such as Zábalo River, the Lakes Tinkias and Pilchicocha, and Lagartococha during rising waters, were characterized by low levels of temperature and dissolved oxygen, with higher depths, and also by higher densities of Asplanchna sp3 and Filinia pejleri. Additionally, Pilchicocha was characterized by high temperatures during high waters. Grande Lake showed high densities of Asplanchna sp1 and sp2, Bosminiopsis sp., and Brachionus zahniseri during rising waters and of Bosminiopsis deitersi during high waters. These species, together with Polyarthra cf. dolychoptera, were associated with high water temperatures (Figure 4).
3.2. Temporal and Vertical Zooplankton Diversity Patterns within Delfincocha and Grande Lakes
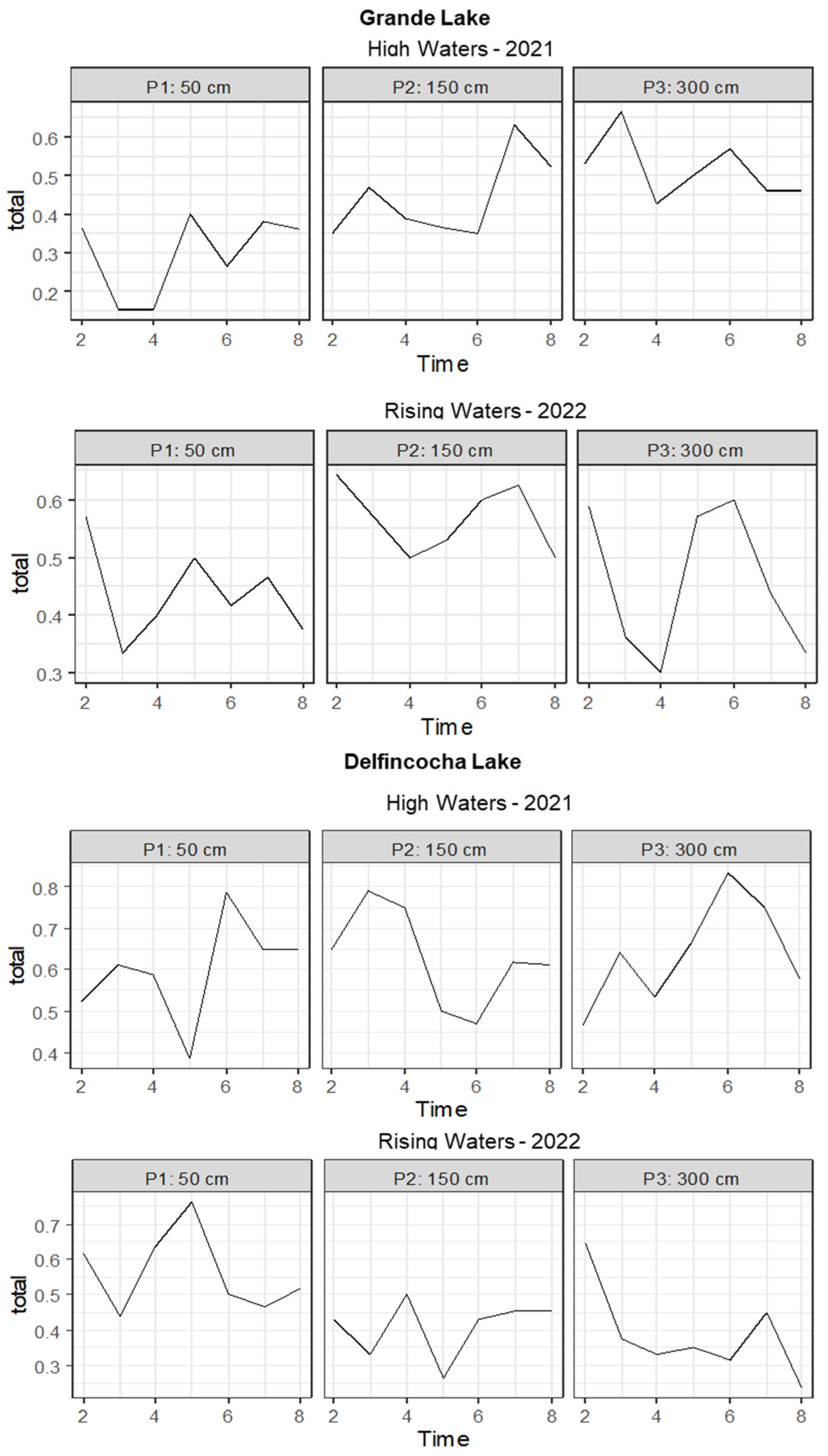
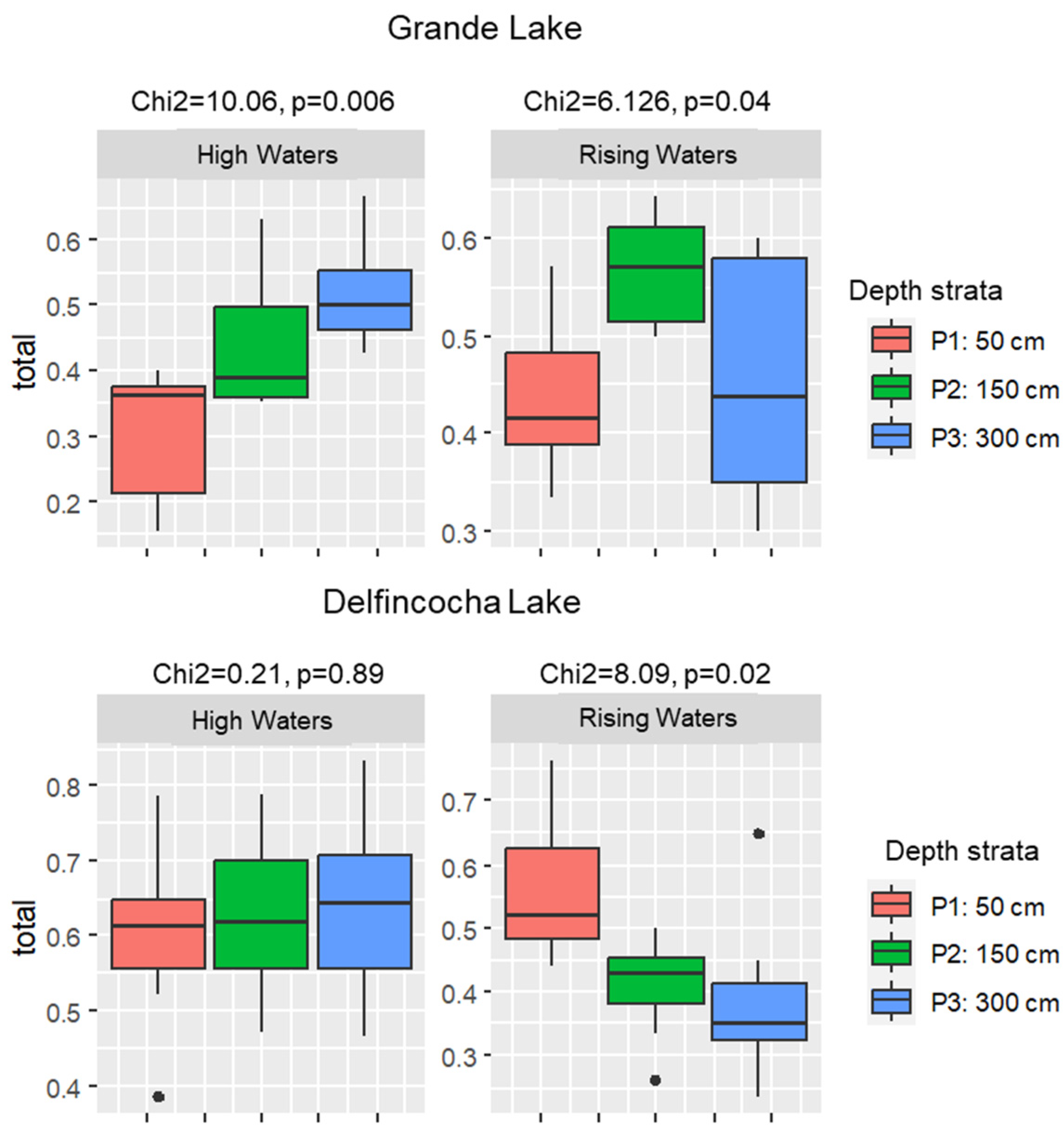
The temporal turnover across the water column varied greatly in the two studied lakes. In Grande, this index ranged from 0.15 to 0.66 during high waters and from 0.3 to 0.64 in rising waters. Significant differences in species turnover between strata were observed during both seasons (Figure 5). However, in the abnormal high waters, the turnover index progressively increased with depth, whereas in rising waters, the turnover index was higher in the mid stratum.
In Grande Lake, cladoceran abundance exhibited high daily variation across strata during high waters, which was more pronounced in the first two strata, between 50 and 150 cm (Figure S1), and between midnight, at 24:00 h, to 3:00 AM in the morning. Amoebozoans were distributed mostly in deeper waters (300 cm). Copepods were distributed indiscriminately but were more abundant at late hours (21 to 24 h). In this lake, time of the day (F = 2.88, p < 0.01) and water stratum (F = 9.05, p < 0.01) had significant effects on zooplankton abundance during high waters, but only time had a significant effect (F = 3.98, p < 0.01) during rising waters. The abundances across strata during high waters were remarkably different in Delfincocha, where Amoebozoa was the most abundant taxa in the three strata throughout the day (Figure S2). The abundances of copepods and cladocerans were very low in comparison to the other taxa. These patterns persisted during rising waters. We did not find significant differences in zooplankton abundances by depth strata or time in Delfincocha in any season. Our results provide evidence that there is a clear stratification in zooplankton communities in Grande Lake that is persistent across hydrological seasons.
In Delfincocha Lake, the turnover index ranged from 0.38 to 0.83 during high waters and from 0.23 to 0.76 during rising waters. No significant differences in species turnover between depth strata were observed in high waters, but differences were observed in rising waters, when the turnover pattern was opposite to that of Grande Lake in high waters, as the index progressively decreased with depth (Figure 6).
On the other hand, the turnover index showed significant temporal fluctuations in Grande Lake in all three depth strata during both sampled seasons (Figure 6). However, this index tended to be higher in the deepest stratum in high waters (300 cm), ranging between 4.5 to over 6.5 (Figure 6). In this lake, during rising waters, fluctuations tended to be more drastic, indicating a higher species replacement throughout the day, which was much more pronounced at a 300 m depth. Additionally, it was observed that variations in the turnover index occurred in all three strata during the same time intervals. In the surface stratum (50 cm), a peak was observed at time interval 5, corresponding to 6:00 in the morning. In the middle stratum (150 cm), this peak was observed at time interval 7, corresponding to 12:00 noon, while in the deepest stratum (300 cm), drastic declines were observed at time interval 4, corresponding to 3:00 in the morning, followed by an increase at time interval 6, corresponding to 9:00 in the morning. All these variations indicate that within Grande Lake, zooplankton execute daily predictable movements across the water column throughout the seasons.
In Delfincocha Lake, drastic variations in the turnover index took place in the three depth strata during the abnormal high waters season (Figure 6), implying significant species replacements at different water depths within a 24 h period. However, these variations were more pronounced at 300 cm, where the turnover index ranged between 0.4 and over 0.6. This pattern persisted and became even more pronounced during rising waters. In contrast to Grande Lake, we did not find similar zooplankton species turnover patterns between seasons, indicating that variations in this lagoon are more stochastic.
The linear models of the turnover indices with water temperature and dissolved oxygen showed contrasting results for Delfincocha and Grande Lakes. In the former, there was a significant negative relationship only with temperature. In Grande Lake, there was a positive relationship of turnover with temperature and a negative one with dissolved oxygen (Table 4).
4. Discussion
Our study encompasses spatial and temporal variation patterns of zooplankton, from the broader scale of the Ecuadorian northern and central Amazon region to the finest one of movements across the water column throughout 24 h of a day. To our knowledge, this is the first study of this kind for the Ecuadorian Amazon.
4.1. Zooplankton Diversity Patterns in the Floodplains of the Napo and Pastaza Watersheds
Species richness and diversity tended to be higher in the Aguarico Watershed in comparison to the Napo Watershed, where there is a longer history of anthropogenic intervention [34,35]. We registered a total of 107 species, which is higher than the richness of 79 species observed in 15 water bodies of the Colombian Amazon [3] or the 69 species observed in the floodplain of the Trombetas River in Brazil [36]. The observed richness is high, considering that we sampled only two hydrological seasons. For instance, in the floodplain of the Yangtze River, China, 128 zooplanktonic species were sampled in a study encompassing four hydrological seasons [37]. However, the estimated zooplankton diversity is lower than in Loktak Lake in India, which was also sampled during two seasons but where 142 to 162 species were recorded [38]. In addition, our collection of zooplankton showed high completeness in the Napo and Aguarico Watersheds and their different water bodies but not in the Pastaza.
Firstly, at a broad taxonomic resolution, we observed significant differences in clade distributions between water bodies and habitats that were sampled during the high waters of 2021 and rising waters of 2022, as indicated by the Chi2 analysis. Amoebozoa species were more diverse in Kusutkao River. Some amoebas are positively correlated to dissolved oxygen and can be dominant in these environments due to their generalist diets [39]. Also, higher turbulence can propitiate the transportation of these organisms from the sediments to the water column [40]. Rotifers were the most diverse taxa in the study area, a widespread pattern in tropical floodplains [37,41], which was favored by these organisms’ opportunistic life history strategies [42] and a trend that is typical of mesotrophic systems of low altitudes [43] and high turbidity [40]. Cladocerans and copepods, on the contrary, were more diverse in lentic water bodies, such as the Lakes Pilchicocha and Grande, which might be related to the presence of aquatic or submerged vegetation [44,45]. We found that the rofifers Keratella cf. cochlearis, Hexarthra intermedia braziliensis, and a copepod of the order Calanoidea are indicators of lentic conditions.
Floodplains are highly dynamic systems, where flooding processes can affect biodiversity in distinct ways. Flooding from hyporheic origin and from the main channel could propitiate a pulse in nutrients that could favor a rise in primary productivity [46]. In a similar fashion, for copepods and cladocerans, favorable conditions in water temperature, increased aquatic vegetation diversity, and vegetation cover during rising waters can trigger the hatching of sediment resting eggs, increasing the diversity of zooplankton both locally and regionally [47]. However, we observed higher gamma diversity during high waters when the water levels were abnormally low. Remarkably, even though gamma diversity decreased during rising waters, we did not find significant differences in alpha diversity between years. As for gamma diversity, in the floodplain of the Paraná River, reductions in alpha diversity did not result in decreased gamma diversity because the loss of some species in some localities was compensated with the maintenance of the species pool by others via dispersal processes at periods of higher connectivity [48]. For our entire study area, alpha diversity (q0) decreased in approximately five species from one season to the next in comparison to a reduction in 33 species regionally, as indicated by the results of gamma diversity (q0). Consequently, our results suggest that the changes in alpha diversity might not be the cause of the variations in gamma diversity between years, and instead, the reduction in beta diversity during rising waters could have played a substantial role in the decrease in gamma diversity observed that year. Variations in zooplankton abundances as a response to the dilution of nutrients during high water levels, together with the higher exchange of organisms between the pelagic and the littoral zones as well as between floodplain lakes and channels have been linked to decreasing beta diversity [5,49].
On the other hand, higher connectivity during the rising waters of 2022 within the floodplains of the Napo and Pastaza Rivers favored a certain degree of longitudinal homogenization between channels and lakes, manifesting in lower beta diversity during that season. On the contrary, during the high waters of 2021, beta diversity was higher for both common and rare species, as indicated by the q1 and q2 Hill numbers. These patterns are consistent with the longitudinal homogenization of communities during high waters [5,50].
Water temperature and dissolved oxygen were the variables that determined the structure of zooplankton communities in the study area [49]. Chlorophyll, total dissolved solids, or conductivity did not have significant effects on zooplankton distribution, but this might have resulted from narrow gradients of these variables in our study area [37]. Our RDA showed that there was considerable difference in zooplankton composition among water bodies during the high waters of 2021, but this season showed abnormal levels that seem to have contributed to this differentiation, while the higher levels during rising waters increased the similarity in environmental parameters and zooplankton composition [36]. The environmental conditions affected the taxa differently [51]. The rotifer Keratella cf. cochlearis was associated with high dissolved oxygen, while the cladocerans Bosminiopsis sp. and B. deitersi were associated with higher temperatures. Therefore, our research implicates that alterations of these parameters, resulting from global change, could impact the zooplankton species distribution and community structure. Likewise, understanding the ecological processes behind the species distribution is key for their future conservation.
4.2. Vertical and Temporal Zooplankton Diversity Patterns
Vertical migration of zooplankton is driven by light, temperature, predation, reproduction, and foraging conditions, as well as by organism density [52,53]. Upwards movements can result from unfavorable conditions in the deeper strata, related to decreased dissolved oxygen concentrations [54]. Water transparency can also determine migration amplitude [55]. In our study, at a coarse taxonomic scale, there was little variation in the dominant taxa between years in both Grande and Delfincocha Lakes. Nevertheless, there were differences in zooplankton abundances and composition between lakes and across depth strata that might be related to contrasting environmental conditions. For instance, Grande Lake is deeper and wider than Delfincocha, which makes it more prone to a vertical stratification in light and oxygen, especially at high water levels. In Grande Lake, we observed that species turnover varied significantly across strata during both seasons. For instance, turnover was higher between 150 to 300 cm during high waters, but only at 150 cm during rising waters. On the contrary, in Delfincocha, these variations in turnover occurred mostly during rising waters, at higher water levels. In Delfincocha, turnover was higher at 300 cm during 2021 and at 50 cm during rising waters. A strong effect of wind mixing during low waters could have prevented the stratification of zooplankton communities in Delfincocha in the high waters of 2021 [56]. These results show that although there was a homogenization of zooplankton communities among sampling sites during the rising waters season of 2022, at a local scale, beta diversity was marked across the vertical and temporal gradients within both lakes. The turnover index showed that in Grande and Delfincocha Lakes, there were remarkable daily fluctuations in the species present in the three strata [33], demonstrating that these zooplankton communities are highly dynamic, despite contrasting environmental conditions between lakes.
Water temperature was a determinant of turnover across the water column in Grande and Delfincocha Lakes, but dissolved oxygen was important only in Grande. We speculate that Delfincocha, being shallower, goes through stronger mixing due to winds, which could prevent a significant stratification in oxygen. More strikingly, temperature was related to decreased turnover in Delfincocha, but within Grande, zooplankton turnover increased at higher temperatures and decreased at higher dissolved oxygen concentrations, which suggests that low dissolved oxygen levels at the deepest strata are a driver of species turnover, as well as higher temperature at the surface. However, it is possible that other factors that we did not consider, such as water transparency [55] and energy conservation [54], might also influence zooplankton turnover.
There were differences in abundances among clades across depth strata and time, particularly within Grande Lake. During high waters, Amoebozoans were more abundant at night and at the deeper strata, but during rising waters, their abundances were higher in the shallower strata, which, considering their small size, could be the result of predation avoidance. Cladocerans occupied the shallow strata across time during the abnormal high waters season, but the next year, the higher abundances of this clade were observed during the afternoon hours, also at the shallower strata. This distribution could reflect preferences for more transparent waters as well as fish predation avoidance [57] since predator fish such as planktivore catfish tend to occupy the benthic zone [58]. Cladocerans were also associated with littoral vegetation, which represent an important refugia for this clade [59,60].
Finally, this study has some limitations that need to be acknowledged. First, due to logistic difficulties, some water bodies could not be sampled during both hydrological seasons. Additionally, the number or sampling sites was not even for the Pastaza River, and the number of sampled lakes and rivers was also uneven. Likewise, there were limitations in the taxonomic resolution for copepods and other taxa that arise from the limited knowledge status for Copepoda and Cladocera of Ecuador [11,61]. Considering these shortcomings, we are not able to make clear cut conclusions about the differences in zooplankton diversities observed between Napo and Pastaza, and thus, further efforts are needed to unveil the diversity patterns between basins. Likewise, although we found a distinctive pattern in the distribution of Amoebozoa in the Kusutkao River, this was not consistent for other rivers, such as Cuyabeno or Lagartococha. However, here, we provide the first results of zooplankton diversity patterns at different spatial and temporal scales for the Napo and Pastaza floodplains, important tributaries of the Western Amazon. We expect these results to constitute a reference baseline for future studies on zooplankton diversity of the region and of the Ecuadorian Amazon.
5. Conclusions
Zooplankton communities in the Napo and Pastaza floodplains show high alpha and beta diversity, and their dynamics are complex. Their structure is determined by variations in environmental conditions across hydrological seasons as well as by habitat type and the local conditions within the water bodies. At high water levels, the increased connectivity between the river channels and their floodplains facilitates the longitudinal homogenization of zooplankton communities, although in a vertical axis, determined by water depth, zooplankton can show a clear stratification and high daily species turnover. At a regional scale, zooplanktonic gamma diversity seems to be greatly influenced by the loss of species, rather than by decreases in local alpha diversity.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/w16081166/s1, Figure S1: Distribution of clade abundances by depth stratum and time of the day in Grande Lake; Figure S2: Distribution of clade abundances by depth stratum and time of the day in Delfincocha Lake; Table S1: Distribution of zooplankton species by water body in the Ecuadorian Western Amazon; Table S2: zooplankton densities used for the calculations of alpha, beta and gamma diversity; Table S3: zooplankton counts during 24 h in Grande and Delfincocha lakes for high waters of 2021 and rising waters of 2022; Supplementary File S1: photographic register of zooplankton species in the Napo and Pastaza floodplains. Supplementary File S2: physical-chemical water parameters used in the RDA and linear models. Supplementary File S3: Pearson correlation analysis to select the water parameters for the Redundancy Analysis.
Author Contributions
Conceptualization, G.E. and F.C.; methodology, F.S.; formal analysis, G.E.; investigation, F.S. and G.E.; resources, F.C.; data curation, F.S.; writing—original draft preparation, G.E.; writing—review and editing, F.C.; project administration, F.C.; funding acquisition, F.C. All authors have read and agreed to the published version of the manuscript.
Funding
The authors report that financial support was provided by the Belgian Development Co-operation (DGD) and the World Wildlife Fund Inc. (WWF) through grants EC14507 and EC11553. Additionally, the study was funded by Universidad de Las Américas (UDLA) through grant FGE.JMC.21.01.
Data Availability Statement
Data supporting all the results are provided here as Supplementary Materials.
Acknowledgments
We thank the Cofán community of Sábalo, the Kichwa community of Zancudococha, the Siona communities of the Cuyabeno Lakes region, and the Achuar communities of Kapawi, Sharamentza, Tinkias, and Wachirpas for hosting field research teams for this study and for granting permission to sample biodiversity in their territories. Along the same line, we thank Sacha Lodge for all the logistical support in the Napo, Pilchicocha, and Pañacocha field campaigns. We also thank the indigenous guides who accompanied and assisted us with specimen collection: Ruben Shakai, Jacob Tangoy, and Enrique Moya. We thank Francis Boily for his invaluable assistance during the fish samplings and for his hospitality. The authors are grateful to the Belgian Development Cooperation (DGD) for the financial support provided and the World Wildlife Fund Inc. (WWF) for the financial, technical, and logistics support provided. We are especially grateful to Gabriela Maldonado (WWF) for all the support provided during the whole project. This article and its findings are those of the authors and do not necessarily reflect the views of the Belgian Development Cooperation. We thank Blanca Rios-Touma for her contributions and comments throughout the process of preparation of the manuscript.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Poff, N.L. Landscape filters and species traits: Towards mechanistic understanding and prediction in stream ecology. J. N. Am. Benthol. Soc. 1997, 16, 391–409. [Google Scholar] [CrossRef]
- Thorp, J.; Thoms, M.; Delong, M. The Riverine Ecosystem Synthesis; Elsevier: London, UK, 2008. [Google Scholar]
- Aranguren-Riaño, N.; Guisande, C.; Ospina, R. Factors controlling crustacean zooplankton species richness in Neotropical lakes. J. Plankton Res. 2011, 33, 1295–1303. [Google Scholar] [CrossRef]
- Simões, N.R.; Dias, J.D.; Leal, C.M.; de Souza Magalhães Braghin, L.; Lansac-Tôha, F.A.; Bonecker, C.C. Floods control the influence of environmental gradients on the diversity of zooplankton communities in a neotropical floodplain. Aquat. Sci. 2013, 75, 607–617. [Google Scholar] [CrossRef]
- Thomaz, S.M.; Bini, L.M.; Bozelli, R.L. Floods increase similarity among aquatic habitats in river-floodplain systems. Hydrobiologia 2007, 579, 1–13. [Google Scholar] [CrossRef]
- Junk, W.; Bayley, P.; Sparks, E. The Flood Pulse Concept in River—Floodplain Systems. Can. J. Fish. Aquat. Sci. 1989, 106, 110–127. [Google Scholar]
- Blackburn-Desbiens, P.; Grosbois, G.; Power, M.; Culp, J.; Rautio, M. Integrating hydrological connectivity and zooplankton composition in Arctic ponds and lakes. Freshw. Biol. 2023, 68, 2131–2150. [Google Scholar] [CrossRef]
- Napiórkowski, P.; Bąkowska, M.; Mrozińska, N.; Szymańska, M.; Kolarova, N.; Obolewski, K. The effect of hydrological connectivity on the zooplankton structure in floodplain lakes of a regulated large river (the Lower Vistula, Poland). Water 2019, 11, 1924. [Google Scholar] [CrossRef]
- Jenkins, C.N.; Pimm, S.L.; Joppa, L.N. Global patterns of terrestrial vertebrate diversity and conservation. Proc. Natl. Acad. Sci. USA 2013, 110, E2602–E2610. [Google Scholar] [CrossRef] [PubMed]
- Fuentes-Reines, J.M.; de Roa, E.Z. New additions to the cladoceran fauna of Ciénaga Grande de Santa Marta and Colombia. Check List 2013, 9, 9–24. [Google Scholar] [CrossRef]
- López, C.; Mosquera, P.V.; Hampel, H.; Neretina, A.N.; Alonso, M.; Van, D.K.; Kotov, A.A. An annotated checklist of the freshwater cladocerans (Crustacea: Branchiopoda: Cladocera) of Ecuador and the Galápagos Islands. Invertebr. Zool. 2018, 15, 277–291. [Google Scholar] [CrossRef]
- López, C.; Soto, L.M.; Lafuente, W.; Stamou, G.; Michaloudi, E.; Papakostas, S.; Fontaneto, D. Rotifers from inland water bodies of continental Ecuador and Galápagos Islands: An updated checklist. Zootaxa 2020, 4768, 551–564. [Google Scholar] [CrossRef] [PubMed]
- Araújo-Flores, J.M.; Garate-Quispe, J.; Molinos, J.G.; Pillaca-Ortiz, J.M.; Caballero-Espejo, J.; Ascorra, C.; Silman, M.; Fernandez, L.E. Seasonality and aquatic metacommunity assemblage in three abandoned gold mining ponds in the southwestern Amazon, Madre de Dios (Peru). Ecol. Indic. 2021, 125, 107455. [Google Scholar] [CrossRef]
- Andrade-Sossa, C.; Alvarez-Silva, J.P.; Aranguren-Riaño, N.; Cupitra-Gómez, O.S.; Villabona-González, S.L.; Torres-Bejarano, A.M.; López, C. Zooplankton studies in Colombian fresh and brackish water ecosystems: A review and future perspectives. Limnologica 2023, 100, 126081. [Google Scholar] [CrossRef]
- Castello, L.; McGrath, D.G.; Hess, L.L.; Coe, M.T.; Lefebvre, P.A.; Petry, P.; Macedo, M.N.; Renó, V.F.; Arantes, C.C. The vulnerability of Amazon freshwater ecosystems. Conserv. Lett. 2013, 6, 217–229. [Google Scholar] [CrossRef]
- Finer, M.; Jenkins, C.N.; Pimm, S.L.; Keane, B.; Ross, C. Oil and Gas Projects in the Western Amazon: Threats to Wilderness, Biodiversity, and Indigenous Peoples. PLoS ONE 2008, 3, e2932. [Google Scholar] [CrossRef] [PubMed]
- Mestanza-Ramón, C.; Cuenca-Cumbicus, J.; D’orio, G.; Flores-Toala, J.; Segovia-Cáceres, S.; Bonilla-Bonilla, A.; Straface, S. Gold Mining in the Amazon Region of Ecuador: History and a Review of Its Socio-Environmental Impacts. Land 2022, 11, 221. [Google Scholar] [CrossRef]
- Comité ERFEN-Ecuador. BOLETÍN ERFEN Nro. 012-2021. 2021. Available online: www.inocar.mil.ec (accessed on 1 March 2022).
- Wetzel, R.G.; Likens, G. Limnological Analyses; Springer Science & Business Media: New York, USA, 2000. [Google Scholar]
- Sousa, F.D.R.; Elmoor-Loureiro, L.M.A. Identification key for the Brazilian genera and species of Aloninae (Crustacea, Branchiopoda, Anomopoda, Chydoridae). Pap Avulsos Zool. 2019, 59, e20195924. [Google Scholar] [CrossRef]
- Elmoor-Loureiro, L.M.A. Manual de Identificação de Cladóceros Límnicos do Brasil; Universa Brasília: Brasilia, Brazil, 1997; Volume 1. [Google Scholar]
- Kotov, A.A.; Stifter, P. Notes on the genus Ilyocryptus Sars, 1862 (Cladocera: Anomopoda: Ilyocryptidae). 1. Ilyocryptus plumosus sp. n., a primitive Neotropical member of the I. spinifer-group. Arthropoda Selecta. Русский Артрoпoдoлoгический журнал 2004, 13, 193–198. [Google Scholar]
- Fuentes-Reinés, J.M.; Elmoor-Loureiro, L.M.A. Annotated checklist and new records of Cladocera from the Ciénaga El Convento, Atlántico-Colombia. Panam. J. Aquat. Sci. 2015, 10, 189–202. [Google Scholar]
- Andrade-Sossa, C.; Buitron-Caicedo, L.; Elías-Gutiérrez, M. A new species of Scapholeberis Schoedler, 1858 (Anomopoda: Daphniidae: Scapholeberinae) from the Colombian Amazon basin highlighted by DNA barcodes and morphology. PeerJ 2020, 8, e9989. [Google Scholar] [CrossRef] [PubMed]
- Witty, L.M. Practical Guide to Identifying Freshwater Crustacean Zooplankton, 2nd ed.; Cooperative Freshwater Ecology Unit: Sudbury, ON, USA, 2004. [Google Scholar]
- Siemensma, F.J. Microworld, Mundo de los Organismos Ameboides. Publicación Electrónica Mundial. Available online: https://arcella.nl/ (accessed on 1 May 2022).
- De Caceres, M.; Jansen, F.; De Caceres, M.M. Relationship between Species and Groups of Sites. Package ‘Indicspecies’. 2016, Version 1.6. Available online: https://emf-creaf.github.io/indicspecies/ (accessed on 20 February 2024).
- Li, D. hillR: Taxonomic, functional, and phylogenetic diversity and similarity through Hill Numbers. J. Open Source Softw. 2018, 3, 1041. [Google Scholar] [CrossRef]
- Hsieh, T.C.; Ma, K.H.; Chao, A. iNEXT. Interpolation and Extrapolation for Species Diversity.” cran.r-project.org, p. 18. 2016. Available online: http://chao.stat.nthu.edu.tw/blog/software-download/ (accessed on 1 July 2022).
- Gotelli, N.J.; Chao, A. Measuring and Estimating Species Richness, Species Diversity, and Biotic Similarity from Sampling Data. In Encyclopedia of Biodiversity, 2nd ed.; Academic Press: Waltham, MA, USA; Elsevier Ltd.: Amsterdam, The Netherlands, 2013; Volume 5, pp. 195–211. [Google Scholar] [CrossRef]
- Alberdi, A. Integral Analysis of Diversity Based on Hill Numbers.” cran.r-project.org, p. 34. 2022. Available online: https://github.com/anttonalberdi/hilldiv (accessed on 1 July 2022).
- Legendre, P.; Legendre, L. Numerical Ecology, 2nd ed.; Elsevier Science B.V.: Amsterdam, The Netherlands, 1998. [Google Scholar]
- Hallett, L.M.; Jones, S.K.; MacDonald, A.A.M.; Jones, M.B.; Flynn, D.F.B.; Ripplinger, J.; Slaughter, P.; Gries, C.; Collins, S.L. codyn: An r package of community dynamics metrics. Methods Ecol. Evol. 2016, 7, 1146–1151. [Google Scholar] [CrossRef]
- Lessmann, J.; Fajardo, J.; Muñoz, J.; Bonaccorso, E. Large expansion of oil industry in the Ecuadorian Amazon: Biodiversity vulnerability and conservation alternatives. Ecol. Evol. 2016, 6, 4997–5012. [Google Scholar] [CrossRef] [PubMed]
- Wernersson, A. Aquatic ecotoxicity due to oil pollution in the Ecuadorian Amazon Aquatic ecotoxicity due to oil pollution in the Ecuadorian Amazon. Aquat. Ecosyst. Health Manag. 2004, 7, 127–136. [Google Scholar] [CrossRef]
- Bozelli, R.L.; Thomaz, S.M.; Padial, A.A.; Lopes, P.M.; Bini, L.M. Floods decrease zooplankton beta diversity and environmental heterogeneity in an Amazonian floodplain system. Hydrobiologia 2015, 753, 233–241. [Google Scholar] [CrossRef]
- Lu, Q.; Liu, X.; Qiu, X.; Liang, T.; Chen, J.; Zhao, S.; Ouyang, S.; Jin, B.; Wu, X. Changes and drivers of zooplankton diversity patterns in the middle reach of Yangtze River floodplain lakes, China. Ecol. Evol. 2021, 11, 17885–17900. [Google Scholar] [CrossRef] [PubMed]
- Sharma, B.K.; Sharma, S. Zooplankton diversity of Loktak Lake, Manipur, India. J. Threat. Taxa 2011, 3, 1745–1755. [Google Scholar] [CrossRef]
- Bagatini, I.L.; Spínola, A.L.G.; Peres, B.d.M.; Mansano, A.d.S.; Rodrigues, M.A.A.; Batalha, M.A.P.L.; de Lucca, J.V.; Godinho, M.J.L.; Tundisi, T.M.; Seleghim, M.H.R. Protozooplankton and its relationship with environmental conditions in 13 water bodies of the Mogi-Guaçu basin-SP, Brazil. Biota Neotrop. 2013, 13, 152–163. [Google Scholar] [CrossRef]
- Picapedra, P.H.D.S.; Fernandes, C.; Baumgartner, G.; Sanches, P.V. Drivers of zooplankton spatial dynamics in a small neotropical river. Acta Limnol. Bras. 2022, 34, e13. [Google Scholar] [CrossRef]
- Gabaldón, C.; Devetter, M.; Hejzlar, J.; Šimek, K.; Znachor, P.; Nedoma, J.; Seda, J. Repeated flood disturbance enhances rotifer dominance and diversity in a zooplankton community of a small dammed mountain pond. J. Limnol. 2017, 76. [Google Scholar] [CrossRef]
- Allan, J.D. Life History Patterns in Zooplankton. Am. Nat. 1976, 110, 165–180. [Google Scholar] [CrossRef]
- Pinel-Alloul, B.; Chemli, A.; Taranu, Z.E.; Bertolo, A. Using the diversity, taxonomic and functional attributes of a zooplankton community to determine lake environmental typology in the natural southern boreal lakes (Québec, Canada). Water 2022, 14, 578. [Google Scholar] [CrossRef]
- Lima, A.F.; Lansac-Tôha, F.A.; Velho, L.F.M.; Bini, L.M.; Takeda, A.M. Composition and abundance of Cladocera (Crustacea) assemblages associated with Eichhornia azurea (Swartz) Kunth stands in the Upper Paraná River floodplain. Acta Sci. Biol. Sci. 2003, 25, 41–48. [Google Scholar]
- Lima, A.F.; Lima, F.A.L.-T.; Velho, L.F.M.; Bini, L.M. Environmental Influence on Planktonic Cladocerans and Copepods in the Floodplain of the Upper River Paraná, Brazil. Stud. Neotrop Fauna Environ. 1998, 33, 188–196. [Google Scholar] [CrossRef]
- Tockner, K.; Malard, F.; Ward, J.V. An extension of the flood pulse concept. Hydrol. Process 2000, 14, 2861–2883. [Google Scholar] [CrossRef]
- Zhang, K.; Jiang, F.; Chen, H.; Dibar, D.T.; Wu, Q.; Zhou, Z. Temporal and spatial variations in zooplankton communities in relation to environmental factors in four floodplain lakes located in the middle reach of the Yangtze River, China. Environ. Pollut. 2019, 251, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Bonecker, C.C.; Simões, N.R.; Minte-Vera, C.V.; Lansac-Tôha, F.A.; Velho, L.F.M.; Agostinho, Â.A. Temporal changes in zooplankton species diversity in response to environmental changes in an alluvial valley. Limnologica 2013, 43, 114–121. [Google Scholar] [CrossRef]
- Pereira, A.L.A.; de Carvalho, P.; Granzotti, R.V.; Vieira, L.C.G.; Bini, L.M. Temporal beta diversity increases with environmental variability in zooplankton floodplain communities. Limnology 2024, 25, 1–10. [Google Scholar] [CrossRef]
- Diniz, L.P.; Petsch, D.K.; Bonecker, C.C. Zooplankton β diversity dynamics and metacommunity structure depend on spatial and temporal scales in a Neotropical floodplain. Freshw. Biol. 2021, 66, 1328–1342. [Google Scholar] [CrossRef]
- Ramos, E.A.; de Morais-Junior, C.S.; Rodrigues-Filho, C.A.S.; Sánchez-Botero, J.I.; Júnior, M.M.; Novaes, J.L.C. Influence of spatial and environmental factors on the structure of a zooplankton metacommunity in an intermittent river. Aquat. Ecol. 2022, 56, 239–249. [Google Scholar] [CrossRef]
- Gabriel, W.; Thomas, B. Vertical migration of zooplankton as an evolutionarily stable strategy. Am. Nat. 1988, 132, 199–216. [Google Scholar] [CrossRef]
- Bandara, K.; Varpe, Ø.; Wijewardene, L.; Tverberg, V.; Eiane, K. Two hundred years of zooplankton vertical migration research. Biol. Rev. 2021, 96, 1547–1589. [Google Scholar] [CrossRef] [PubMed]
- Lampert, W. The adaptive significance of diel vertical migration of zooplankton. Funct. Ecol. 1989, 3, 21–27. [Google Scholar] [CrossRef]
- Dodson, S. Predicting diel vertical migration of zooplankton. Limnol. Oceanogr. 1990, 35, 1195–1200. [Google Scholar] [CrossRef]
- Rejas, D.; De Meester, L.; Ferrufino, L.; Maldonado, M.; Ollevier, F. Diel vertical migration of zooplankton in an Amazonian várzea lake (Laguna Bufeos, Bolivia). Stud. Neotrop. Fauna Environ. 2007, 42, 71–81. [Google Scholar] [CrossRef]
- Antón-Pardo, M.; Muška, M.; Jůza, T.; Vejříková, I.; Vejřík, L.; Blabolil, P.; Čech, M.; Draštík, V.; Frouzová, J.; Holubová, M.; et al. Diel changes in vertical and horizontal distribution of cladocerans in two deep lakes during early and late summer. Sci. Total Environ. 2021, 751, 141601. [Google Scholar] [CrossRef] [PubMed]
- Echevarría, G.; Rodríguez, J.P.; Machado-Allison, A. Seasonal fluctuations in taxonomic and functional diversity in assemblages of catfishes in the Venezuelan Arauca River Floodplain. Stud. Neotrop. Fauna Environ. 2018, 53, 38–53. [Google Scholar] [CrossRef]
- Coelho, P.N.; Henry, R. Functional groups of microcrustaceans along a horizontal gradient in a Neotropical lake colonized by macrophytes. Aquat. Sci. 2020, 83, 3. [Google Scholar] [CrossRef]
- Stansfield, J.H.; Perrow, M.R.; Tench, L.D.; Jowitt, A.J.D.; Taylor, A.A.L. Submerged macrophytes as refuges for grazing Cladocera against fish predation: Observations on seasonal changes in relation to macrophyte cover and predation pressure. In Shallow Lakes ’95: Trophic Cascades in Shallow Freshwater and Brackish Lakes; Kufel, L., Prejs, A., Rybak, J.I., Eds.; Springer: Dordrecht, The Netherlands, 1997; pp. 229–240. [Google Scholar] [CrossRef]
- Corgosinho, P.H.C.; Holynska, M.; Marrone, F.; Geraldes-Primeiro, L.J.d.O.; dos Santos-Silva, E.N.; Perbiche-Neves, G.; Lopez, C. An annotated checklist of freshwater Copepoda (Crustacea, Hexanauplia) from continental Ecuador and the Galapagos Archipelago. Zookeys 2019, 871, 55. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Study area showing the distribution of the Napo Moist Forest ecoregion in Ecuador, and the location of sampling sites in the Napo and Aguarico watersheds in the north (A), and in the Pastaza watershed in the south (B).
Figure 1.
Study area showing the distribution of the Napo Moist Forest ecoregion in Ecuador, and the location of sampling sites in the Napo and Aguarico watersheds in the north (A), and in the Pastaza watershed in the south (B).

Figure 2.
Proportions of species richness by clade within each sampling site for the high and rising waters of 2021 and 2022 combined.
Figure 2.
Proportions of species richness by clade within each sampling site for the high and rising waters of 2021 and 2022 combined.
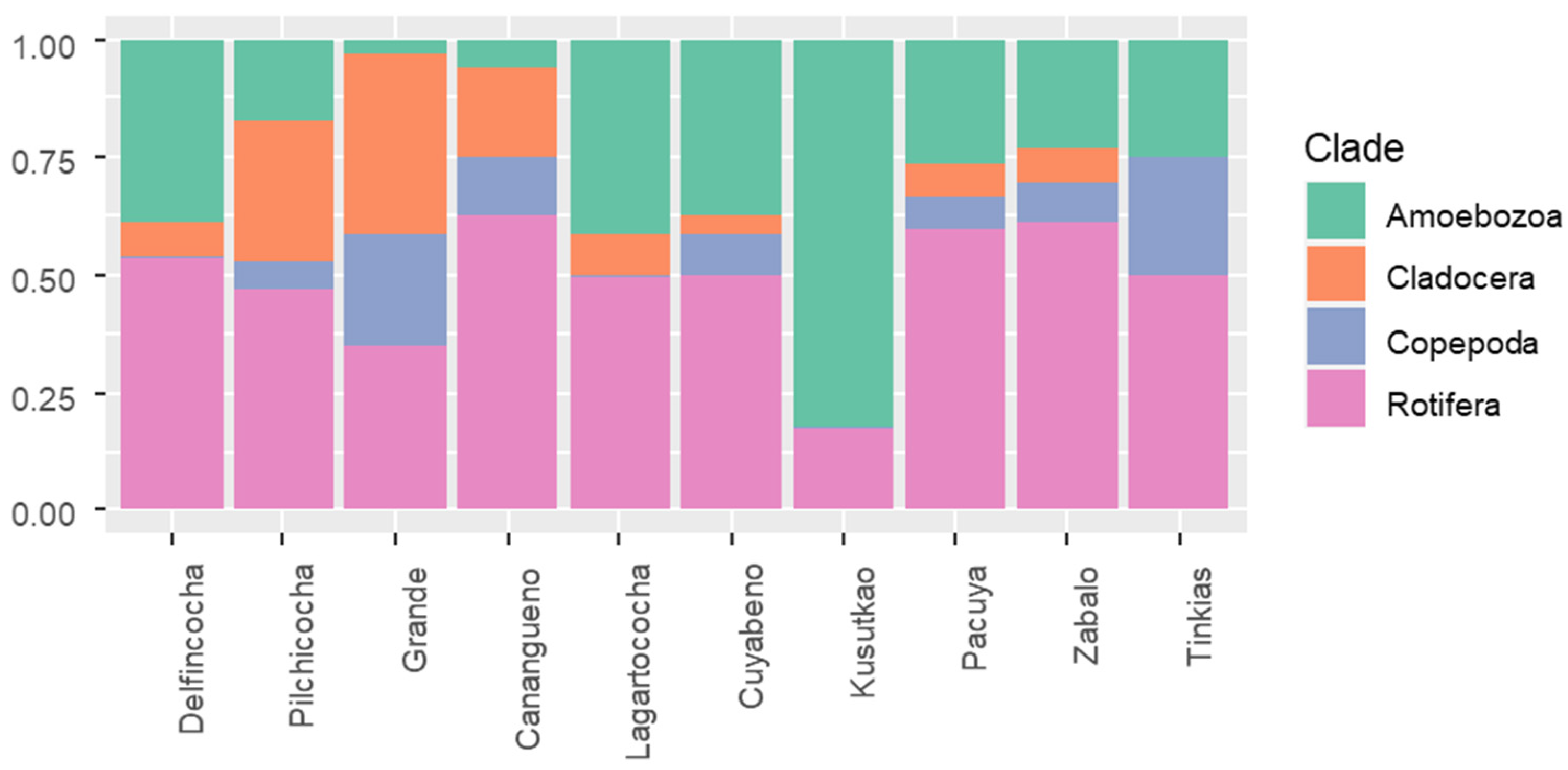
Figure 3.
Rarefaction curves of zooplankton diversity of the Aguarico, Napo, and Pastaza watersheds during the high waters of 2021 (top) and rising waters of 2022 (bottom).
Figure 3.
Rarefaction curves of zooplankton diversity of the Aguarico, Napo, and Pastaza watersheds during the high waters of 2021 (top) and rising waters of 2022 (bottom).
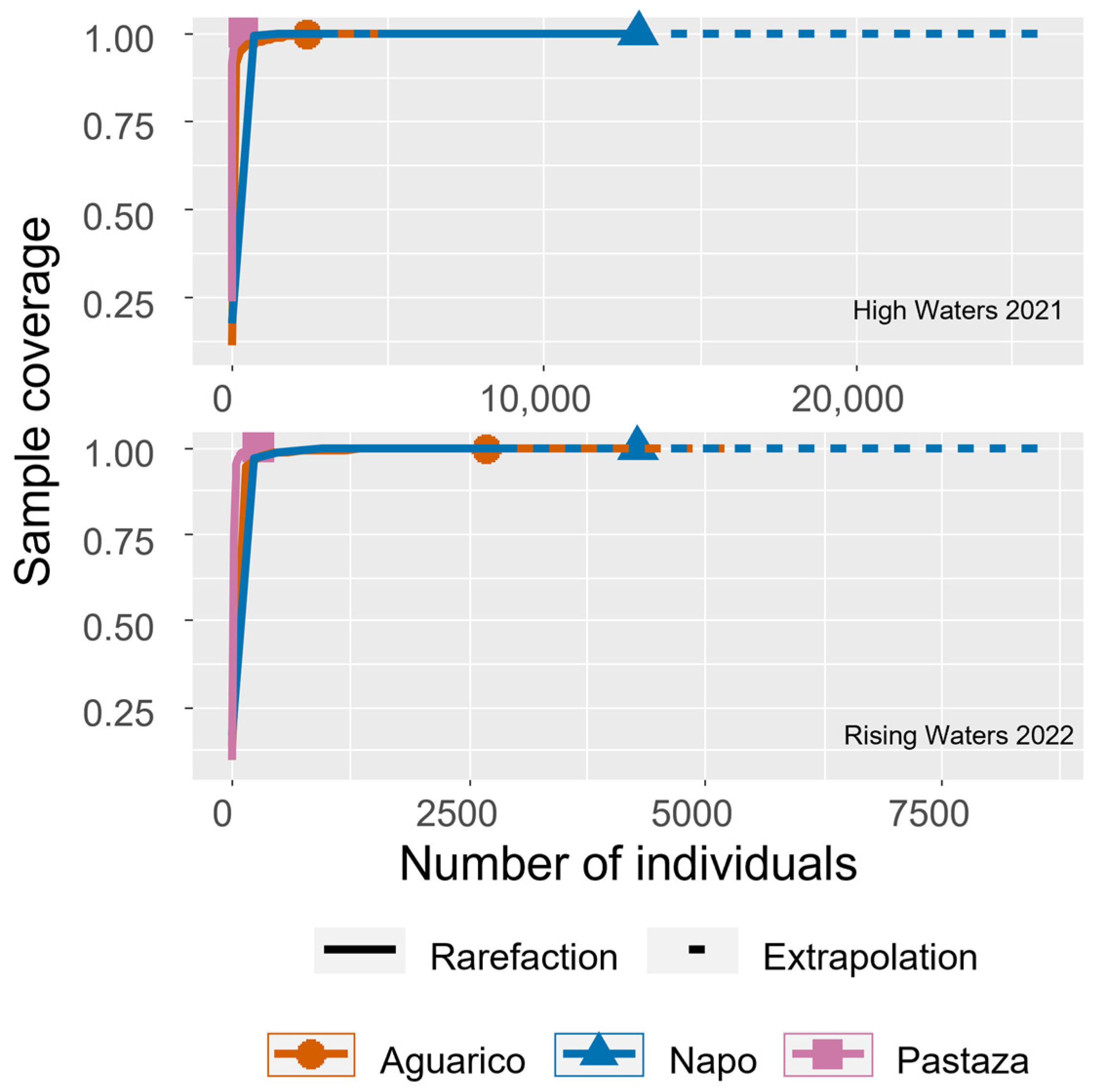
Figure 4.
RDA plot of zooplankton species densities by water body and season, and their relationships with water parameters.
Figure 4.
RDA plot of zooplankton species densities by water body and season, and their relationships with water parameters.
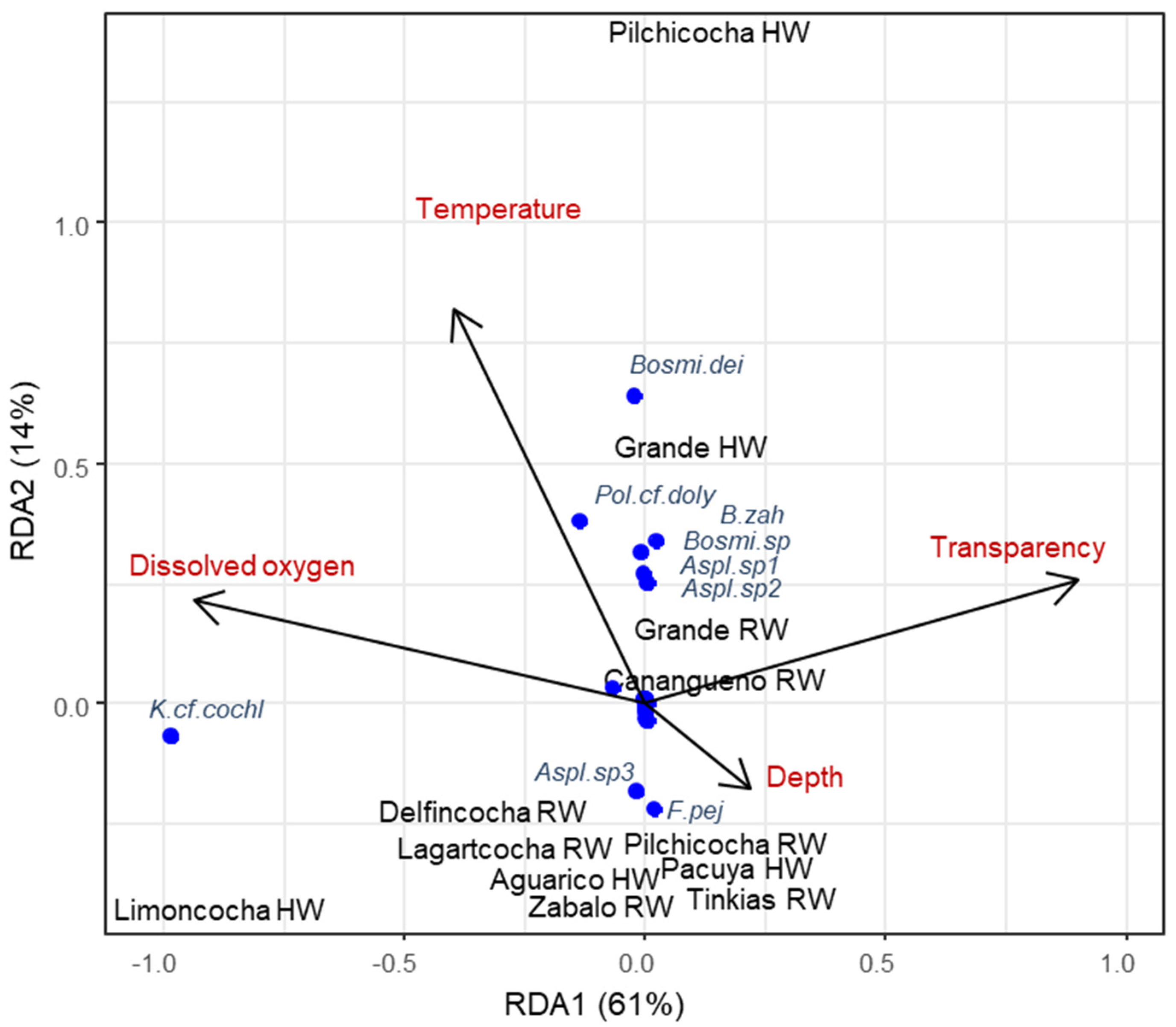
Figure 5.
Boxplots and Kruskal–Wallis test results of the turnover index comparisons across depth strata in Grande Lake (top) and Delfincocha Lake (bottom). P1 to P3 refer to depth strata.
Figure 5.
Boxplots and Kruskal–Wallis test results of the turnover index comparisons across depth strata in Grande Lake (top) and Delfincocha Lake (bottom). P1 to P3 refer to depth strata.
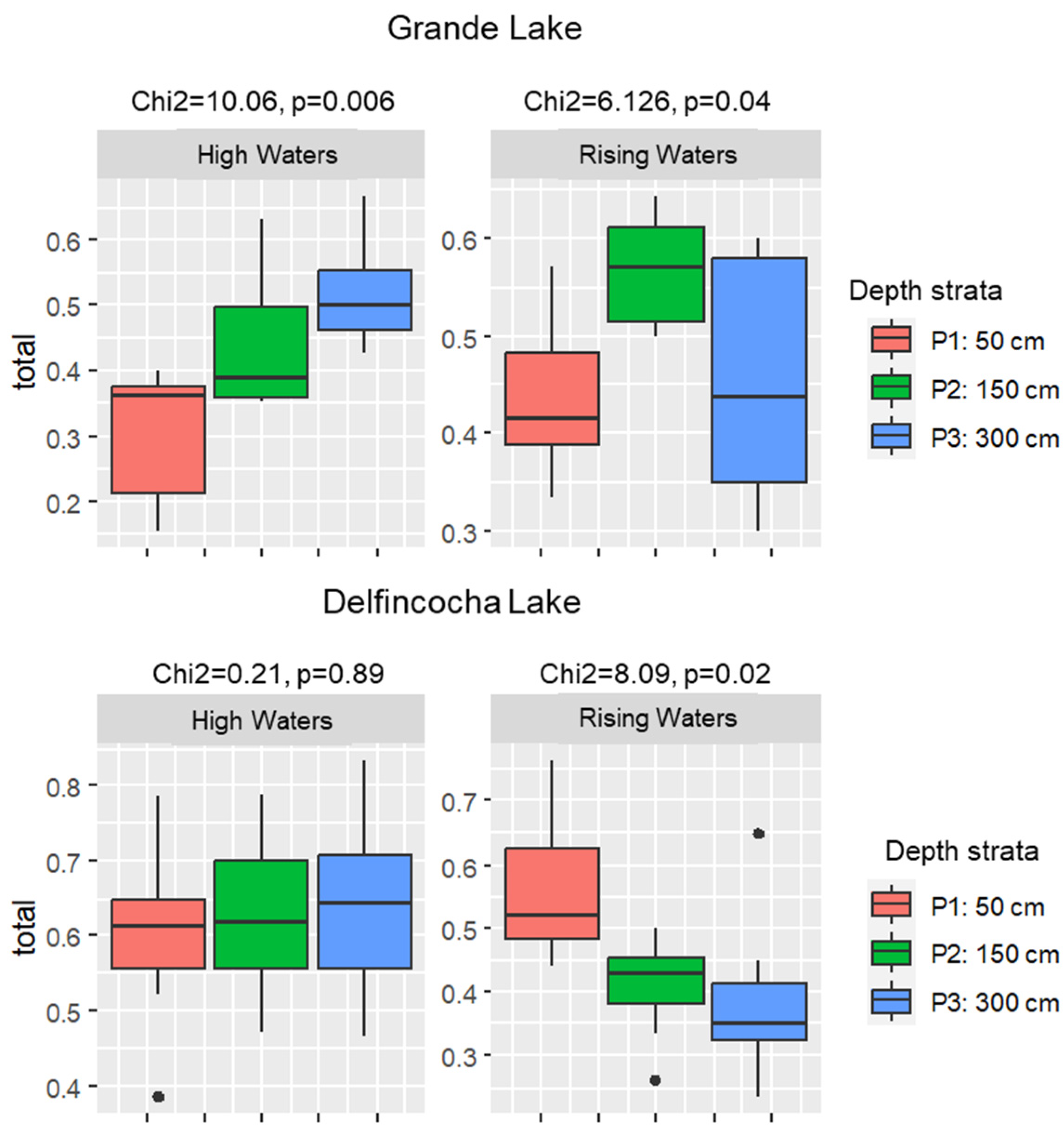
Figure 6.
Fluctuations of the zooplankton turnover index over 24 h in three depth strata in Grande and Delfincocha Lakes.
Figure 6.
Fluctuations of the zooplankton turnover index over 24 h in three depth strata in Grande and Delfincocha Lakes.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Partition of zooplankton diversity in its alpha, beta, and gamma components for the entire study area.
Table 1.
Partition of zooplankton diversity in its alpha, beta, and gamma components for the entire study area.
| Hill Number | Diversity | ||
|---|---|---|---|
| High Waters—2021 | |||
| q | Gamma | Alpha | Beta |
| 0 | 81.00 | 19.40 | 4.17 |
| 1 | 22.9 | 6.54 | 3.50 |
| 2 | 15.64 | 3.62 | 4.28 |
| Rising Waters—2022 | |||
| q | Gamma | Alpha | Beta |
| 0 | 48 | 14.27 | 3.36 |
| 1 | 21.5 | 6.65 | 3.23 |
| 2 | 14.95 | 4.02 | 3.72 |
Table 2.
Zooplankton species selected as indicators of sites during the high waters of 2021 and rising waters of 2022.
Table 2.
Zooplankton species selected as indicators of sites during the high waters of 2021 and rising waters of 2022.
| Indicator Species | A | B | Stat | p Value |
|---|---|---|---|---|
| High Waters—2021: Group 1 + 3: Limoncocha and Canangueno Lakes | ||||
| Keratella cf. cochlearis | 0.9942 | 1 | 0.997 | 0.049 |
| Hexarthra intermedia braziliensis | 0.9764 | 1 | 0.988 | 0.049 |
| Copepoda Calanoida.sp3 | 0.9446 | 1 | 0.972 | 0.026 |
| Rising Waters—2022: Group 2 + 3: Zancudococha and Delfincocha Lakes | ||||
| Hexarthra intermedia braziliensis | 0.9985 | 1 | 0.999 | 0.033 |
Table 3.
Zooplankton q0, q1, and q2 diversity within the sampled water bodies during the two sampled seasons. NAs refer to sites not sampled during that season.
Table 3.
Zooplankton q0, q1, and q2 diversity within the sampled water bodies during the two sampled seasons. NAs refer to sites not sampled during that season.
| Watershed | Water Body | High Waters—2021 | Rising Waters—2022 | ||||
|---|---|---|---|---|---|---|---|
| q0 | q1 | q2 | q0 | q1 | q2 | ||
| Aguarico | Lagartococha River | 22 | 12.34 | 8.23 | 21.00 | 10.90 | 6.49 |
| Delfincocha Lake | 40 | 19.30 | 11.20 | 28.00 | 8.06 | 3.75 | |
| Canangueno Lake | 16 | 2.92 | 1.83 | 14.00 | 8.19 | 7.05 | |
| Grande Lake | 33 | 9.26 | 5.68 | 16.00 | 7.48 | 5.13 | |
| Cuyabeno River | 22 | 14.64 | 11.05 | 17.00 | 13.20 | 11.08 | |
| Zancudococha Lake | NA | NA | NA | 6.00 | 3.05 | 2.32 | |
| Aguarico River | 4 | 2.92 | 2.47 | NA | NA | NA | |
| Zabalo River | NA | NA | NA | 11.00 | 4.92 | 2.91 | |
| Napo | Pilchicocha Lake | 16 | 5.64 | 4.37 | 15.00 | 5.63 | 3.03 |
| Pacuya River | NA | NA | NA | 13.00 | 6.71 | 4.06 | |
| Limoncocha Lake | 16 | 3.91 | 2.30 | NA | NA | NA | |
| Pañacocha Lake | 16 | 4.43 | 2.35 | NA | NA | NA | |
| Pastaza | Kusutkao River | 9 | 5.37 | 4.13 | 13.00 | 10.84 | 10.04 |
| Nueva Tinkias Lake | NA | NA | NA | 3.00 | 2.59 | 2.27 | |
Table 4.
Results of linear models of zooplankton turnover with water temperature and dissolved oxygen in Delfincocha and Grande Lakes, with the data of both seasons combined.
Table 4.
Results of linear models of zooplankton turnover with water temperature and dissolved oxygen in Delfincocha and Grande Lakes, with the data of both seasons combined.
| Delfincocha Lake | |||
| Estimate | t value | p value | |
| Temperature | −0.171 | −3.190 | 0.003 |
| Dissolved oxygen | 0.034 | 0.257 | 0.799 |
| Grande Lake | |||
| Estimate | t value | p value | |
| Temperature | 0.068 | 2.064 | 0.046 |
| Dissolved oxygen | −0.077 | −3.302 | 0.002 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Sánchez, F.; Cuesta, F.; Echevarría, G. Zooplankton Temporal, Longitudinal, and Vertical Diversity Patterns in the Floodplains of the Western Amazon. Water 2024, 16, 1166. https://doi.org/10.3390/w16081166
AMA Style
Sánchez F, Cuesta F, Echevarría G. Zooplankton Temporal, Longitudinal, and Vertical Diversity Patterns in the Floodplains of the Western Amazon. Water. 2024; 16(8):1166. https://doi.org/10.3390/w16081166
Chicago/Turabian StyleSánchez, Fernando, Francisco Cuesta, and Gabriela Echevarría. 2024. "Zooplankton Temporal, Longitudinal, and Vertical Diversity Patterns in the Floodplains of the Western Amazon" Water 16, no. 8: 1166. https://doi.org/10.3390/w16081166
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.