
Inhibition Effects of Nippostrongylus brasiliensis and Its Derivatives against Atherosclerosis in ApoE-/- Mice through Anti-Inflammatory Response
 , , and
, , and 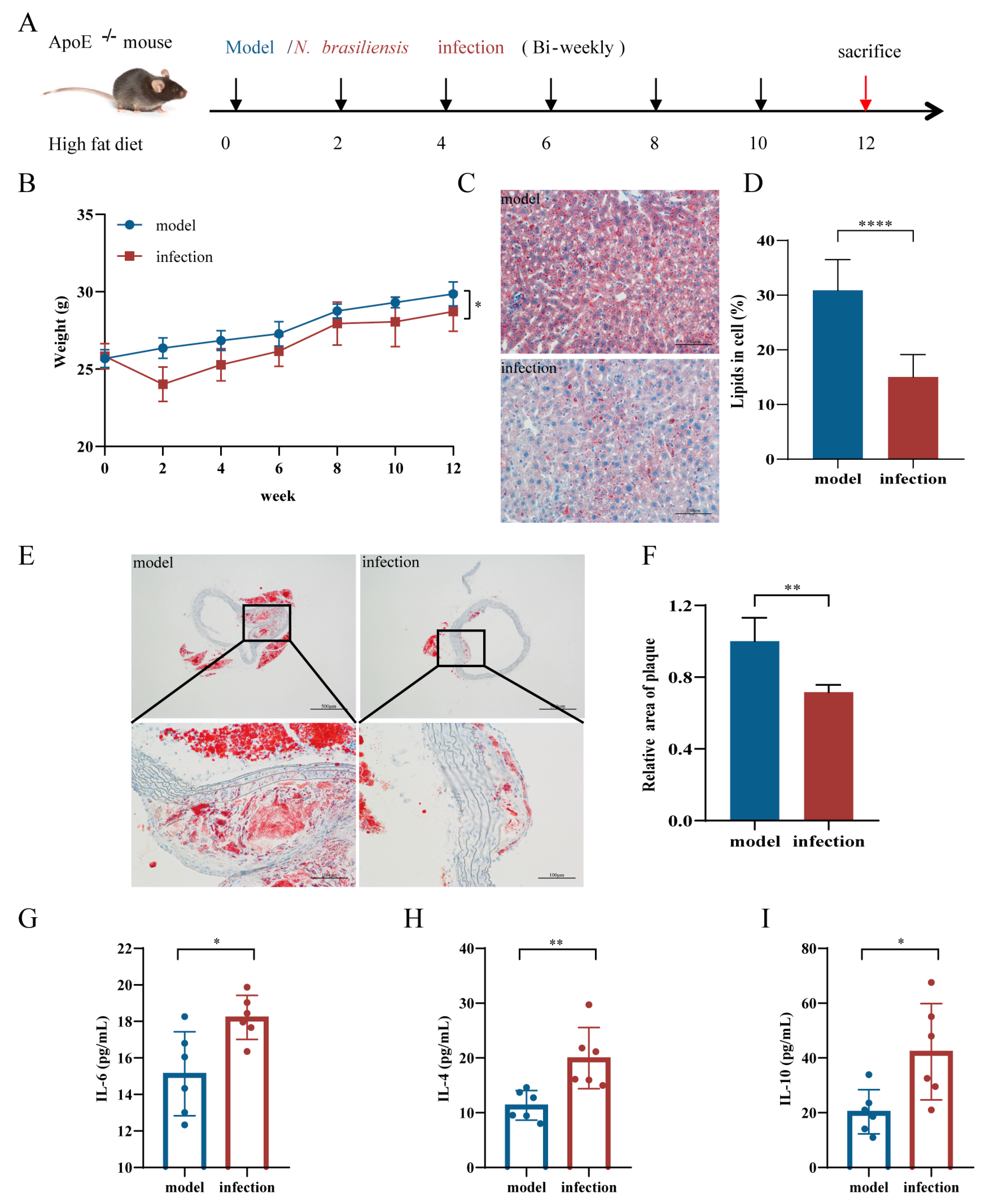{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Parasites and Animals
2.2. Preparation of Proteins Derived from N. brasiliensis
2.3. The Mouse Model of Atherosclerosis and In Vivo Interventions
2.4. Histopathological Analysis
2.5. Detection of Serum Cytokine Levels
2.6. Detection of Serum Lipid Levels
2.7. Statistical Analysis
3. Results
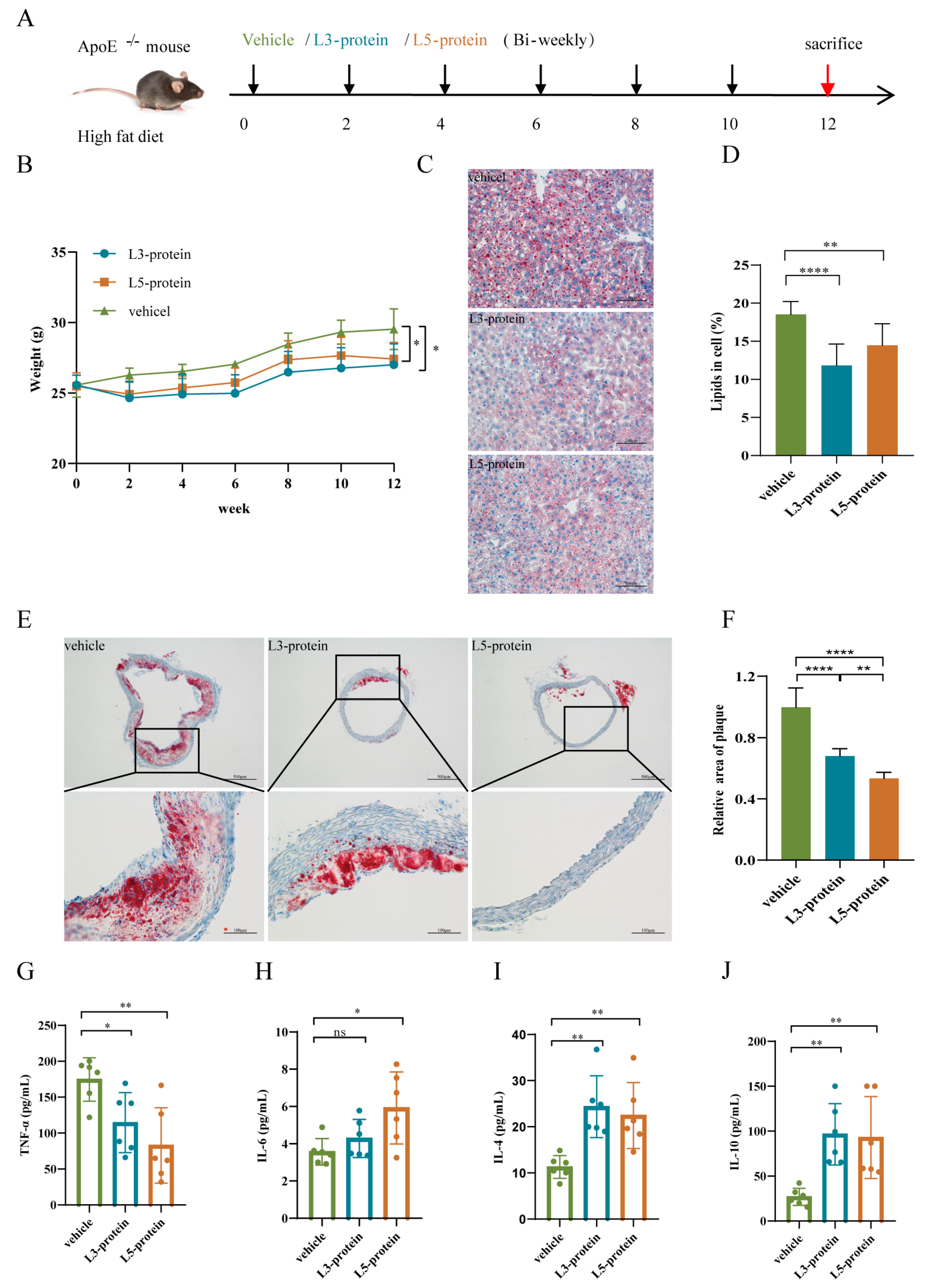
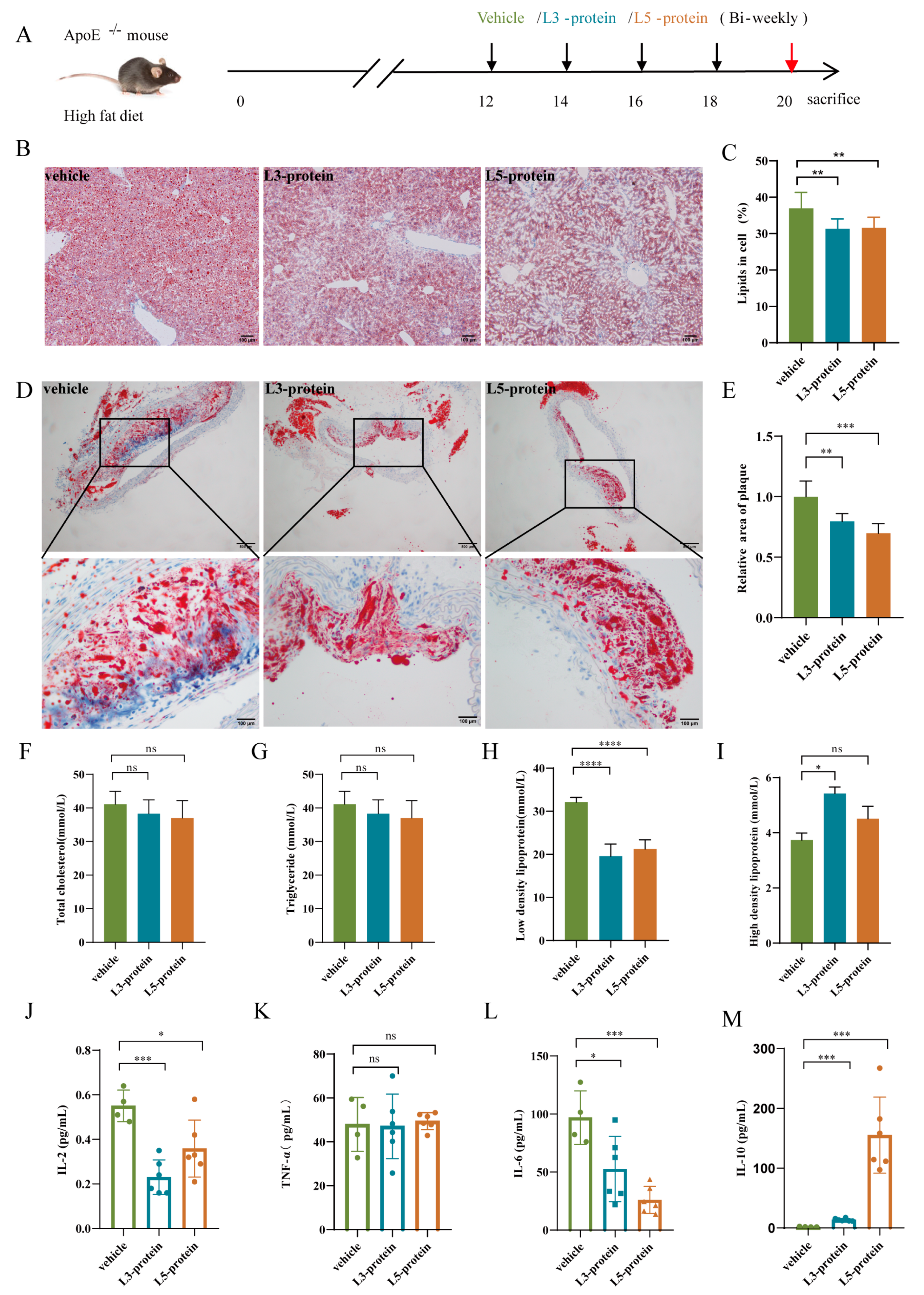
3.1. The effects of N. brasiliensis Infection and Derived Products on the Initiation of Atherosclerosis
3.1.1. Body Weight Changes
3.1.2. Histopathological Alterations in the Liver and Aortic Arch
3.1.3. Serum Cytokine Levels
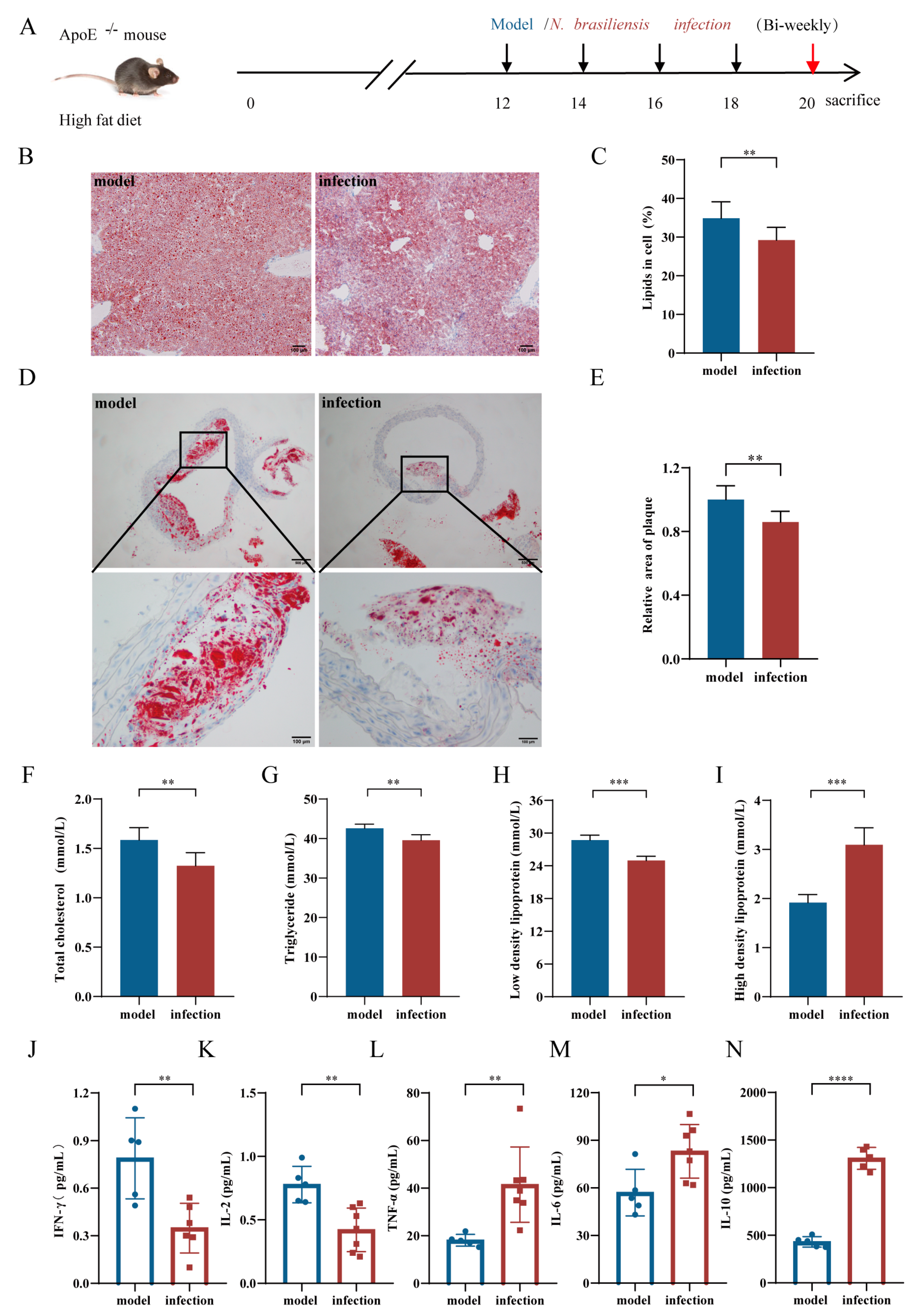
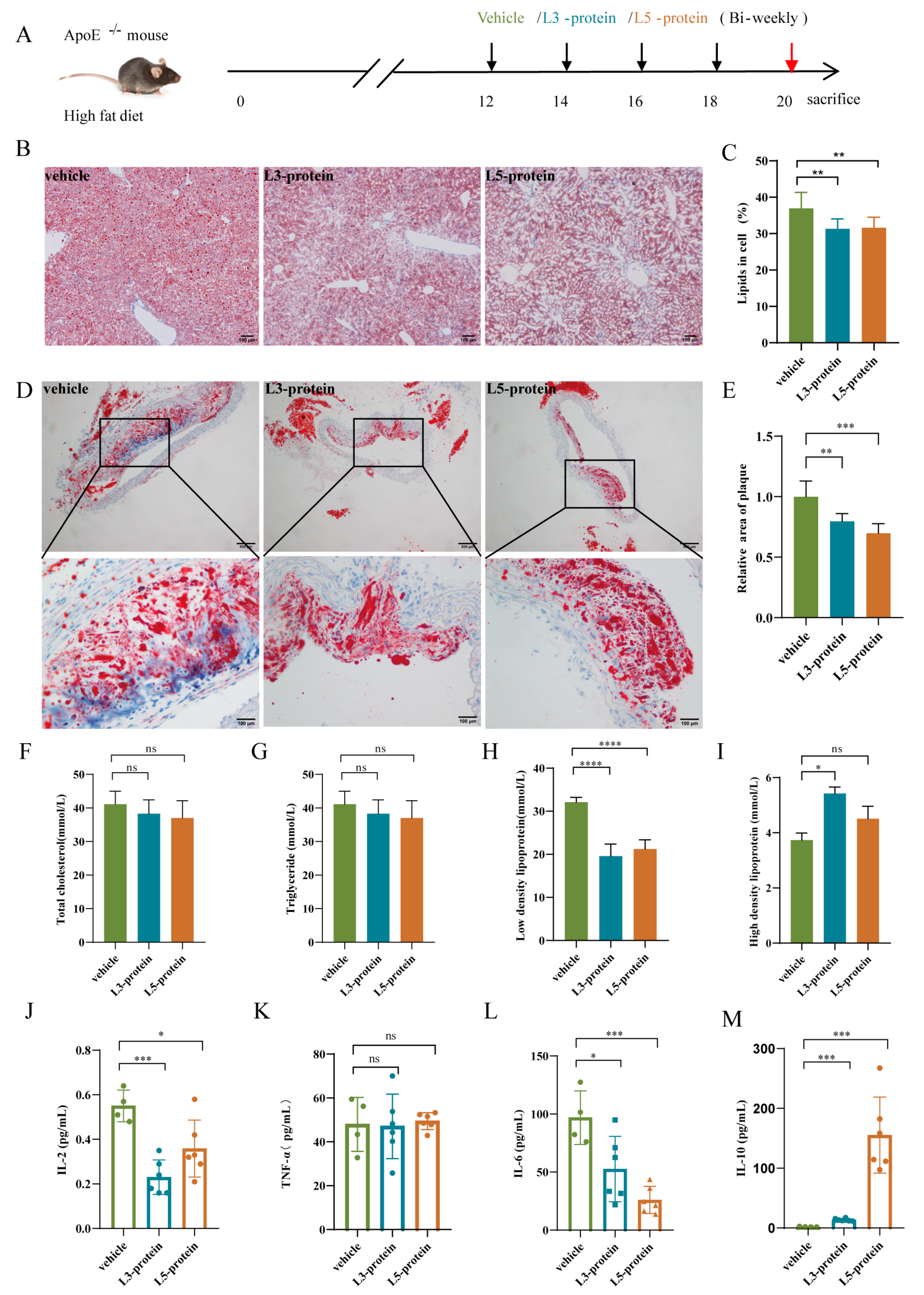
3.2. Effects of N. brasiliensis and Derived Products Intervention on the Progression of AS
3.2.1. Histopathological Alterations in the Liver and Aortic Arch
3.2.2. Serum Lipid Levels
3.2.3. Serum Cytokine Levels
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Back, M.; Yurdagul, A., Jr.; Tabas, I.; Oorni, K.; Kovanen, P.T. Inflammation and its resolution in atherosclerosis: Mediators and therapeutic opportunities. Nat. Rev. Cardiol. 2019, 16, 389–406. [Google Scholar] [CrossRef] [PubMed]
- Hedin, U.; Matic, L.P. Recent advances in therapeutic targeting of inflammation in atherosclerosis. J. Vasc. Surg. 2019, 69, 944–951. [Google Scholar] [CrossRef] [PubMed]
- Poels, K.; van Leent, M.M.T.; Boutros, C.; Tissot, H.; Roy, S.; Meerwaldt, A.E.; Toner, Y.C.A.; Reiche, M.E.; Kusters, P.J.H.; Malinova, T.; et al. Immune Checkpoint Inhibitor Therapy Aggravates T Cell-Driven Plaque Inflammation in Atherosclerosis. JACC CardioOncol. 2020, 2, 599–610. [Google Scholar] [CrossRef] [PubMed]
- Chan, Y.H.; Ramji, D.P. Key Roles of Inflammation in Atherosclerosis: Mediators Involved in Orchestrating the Inflammatory Response and Its Resolution in the Disease Along with Therapeutic Avenues Targeting Inflammation. Methods Mol. Biol. 2022, 2419, 21–37. [Google Scholar] [CrossRef] [PubMed]
- Chan, Y.H.; Ramji, D.P. A perspective on targeting inflammation and cytokine actions in atherosclerosis. Future Med. Chem. 2020, 12, 613–626. [Google Scholar] [CrossRef]
- Mendel, I.; Yacov, N.; Harats, D.; Breitbart, E. Therapies targeting innate immunity for fighting inflammation in atherosclerosis. Curr. Pharm. Des. 2015, 21, 1185–1195. [Google Scholar] [CrossRef]
- Kotlyarov, S.; Kotlyarova, A. Molecular Pharmacology of Inflammation Resolution in Atherosclerosis. Int. J. Mol. Sci. 2022, 23, 4808. [Google Scholar] [CrossRef]
- Strachan, D.P. Hay fever, hygiene, and household size. BMJ 1989, 299, 1259–1260. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, J.S.; Cardoso, L.S.; Pitrez, P.M.; Cruz, A.A. Helminths and Asthma: Risk and Protection. Immunol. Allergy Clin. 2019, 39, 417–427. [Google Scholar] [CrossRef]
- Alcantara-Neves, N.M.; de SG Britto, G.; Veiga, R.V.; Figueiredo, C.A.; Fiaccone, R.L.; da Conceicao, J.S.; Cruz, A.A.; Rodrigues, L.C.; Cooper, P.J.; Pontes-de-Carvalho, L.C.; et al. Effects of helminth co-infections on atopy, asthma and cytokine production in children living in a poor urban area in Latin America. BMC Res. Notes 2014, 7, 817. [Google Scholar] [CrossRef]
- Hewitson, J.P.; Grainger, J.R.; Maizels, R.M. Helminth immunoregulation: The role of parasite secreted proteins in modulating host immunity. Mol. Biochem. Parasitol. 2009, 167, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Maizels, R.M.; Balic, A.; Gomez-Escobar, N.; Nair, M.; Taylor, M.D.; Allen, J.E. Helminth parasites–masters of regulation. Immunol. Rev. 2004, 201, 89–116. [Google Scholar] [CrossRef]
- Lothstein, K.E.; Gause, W.C. Mining Helminths for Novel Therapeutics. Trends Mol. Med. 2021, 27, 345–364. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, I.; Smyth, D.; Gaze, S.; Aziz, A.; Giacomin, P.; Ruyssers, N.; Artis, D.; Laha, T.; Navarro, S.; Loukas, A.; et al. Hookworm excretory/secretory products induce interleukin-4 (IL-4)+ IL-10+ CD4+ T cell responses and suppress pathology in a mouse model of colitis. Infect. Immun. 2013, 81, 2104–2111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, M.; Yang, X.; Yu, Y.; Pan, W. Helminth and Host Crosstalk: New Insight Into Treatment of Obesity and Its Associated Metabolic Syndromes. Front. Immunol. 2022, 13, 827486. [Google Scholar] [CrossRef] [PubMed]
- Stanley, R.G.; Jackson, C.L.; Griffiths, K.; Doenhoff, M.J. Effects of Schistosoma mansoni worms and eggs on circulating cholesterol and liver lipids in mice. Atherosclerosis 2009, 207, 131–138. [Google Scholar] [CrossRef]
- Loukas, A.; Hotez, P.J.; Diemert, D.; Yazdanbakhsh, M.; McCarthy, J.S.; Correa-Oliveira, R.; Croese, J.; Bethony, J.M. Hookworm infection. Nat. Rev. Dis. Prim. 2016, 2, 16088. [Google Scholar] [CrossRef]
- Bartsch, S.M.; Hotez, P.J.; Asti, L.; Zapf, K.M.; Bottazzi, M.E.; Diemert, D.J.; Lee, B.Y. The Global Economic and Health Burden of Human Hookworm Infection. PLoS Negl. Trop. Dis. 2016, 10, e0004922. [Google Scholar] [CrossRef] [Green Version]
- Brooker, S.; Bethony, J.; Hotez, P.J. Human hookworm infection in the 21st century. Adv. Parasitol. 2004, 58, 197–288. [Google Scholar] [CrossRef] [Green Version]
- Tracey, E.F.; McDermott, R.A.; McDonald, M.I. Do worms protect against the metabolic syndrome? A systematic review and meta-analysis. Diabetes Res. Clin. Pract. 2016, 120, 209–220. [Google Scholar] [CrossRef]
- Tahapary, D.L.; de Ruiter, K.; Martin, I.; Brienen, E.A.T.; van Lieshout, L.; Cobbaert, C.M.; Soewondo, P.; Djuardi, Y.; Wiria, A.E.; Houwing-Duistermaat, J.J.; et al. Effect of Anthelmintic Treatment on Insulin Resistance: A Cluster-Randomized, Placebo-Controlled Trial in Indonesia. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2017, 65, 764–771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiria, A.E.; Wammes, L.J.; Hamid, F.; Dekkers, O.M.; Prasetyani, M.A.; May, L.; Kaisar, M.M.; Verweij, J.J.; Tamsma, J.T.; Partono, F.; et al. Relationship between carotid intima media thickness and helminth infections on Flores Island, Indonesia. PLoS ONE 2013, 8, e54855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camberis, M.; Le Gros, G.; Urban, J., Jr. Animal model of Nippostrongylus brasiliensis and Heligmosomoides polygyrus. Curr. Protoc. Immunol. 2003, 55, 19.12.1–19.12.27. [Google Scholar] [CrossRef]
- Gerbe, F.; Sidot, E.; Smyth, D.J.; Ohmoto, M.; Matsumoto, I.; Dardalhon, V.; Cesses, P.; Garnier, L.; Pouzolles, M.; Brulin, B.; et al. Intestinal epithelial tuft cells initiate type 2 mucosal immunity to helminth parasites. Nature 2016, 529, 226–230. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.M.; Zhang, Q.; Ding, X.; Mao, F.Z.; Wang, X.T.; Dai, Y.; Wang, J.H.; Cao, J. Polarization of human acute monocytic leukemia THP-1 cells-derived macprophages induced by Nippostrongylus brasiliensis proteins in vitro. Zhongguo Xue Xi Chong Bing Fang Zhi Za Zhi 2020, 32, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhang, M.; Ding, X.; Yang, Y.; Chen, Y.; Zhang, Q.; Fan, Y.; Dai, Y.; Wang, J. Mining Anti-Inflammation Molecules From Nippostrongylus brasiliensis-Derived Products Through the Metabolomics Approach. Front. Cell. Infect. Microbiol. 2021, 11, 781132. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.; Xu, N.; Wang, X.; Vallee, I.; Liu, M.; Liu, X. Helminth Therapy for Immune-Mediated Inflammatory Diseases: Current and Future Perspectives. J. Inflamm. Res. 2022, 15, 475–491. [Google Scholar] [CrossRef]
- Smallwood, T.B.; Giacomin, P.R.; Loukas, A.; Mulvenna, J.P.; Clark, R.J.; Miles, J.J. Helminth Immunomodulation in Autoimmune Disease. Front. Immunol. 2017, 8, 453. [Google Scholar] [CrossRef] [Green Version]
- Maruszewska-Cheruiyot, M.; Donskow-Lysoniewska, K.; Doligalska, M. Helminth Therapy: Advances in the use of Parasitic Worms Against Inflammatory Bowel Diseases and its Challenges. Helminthologia 2018, 55, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Conti, P.; Shaik-Dasthagirisaeb, Y. Atherosclerosis: A chronic inflammatory disease mediated by mast cells. Cent. Eur. J. Immunol. 2015, 40, 380–386. [Google Scholar] [CrossRef]
- Soehnlein, O.; Libby, P. Targeting inflammation in atherosclerosis—From experimental insights to the clinic. Nat. Rev. Drug Discov. 2021, 20, 589–610. [Google Scholar] [CrossRef] [PubMed]
- Greenwood, B.M. Autoimmune disease and parasitic infections in Nigerians. Lancet 1968, 2, 380–382. [Google Scholar] [CrossRef]
- Summers, R.W.; Elliott, D.E.; Urban, J.F., Jr.; Thompson, R.A.; Weinstock, J.V. Trichuris suis therapy for active ulcerative colitis: A randomized controlled trial. Gastroenterology 2005, 128, 825–832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapman, P.R.; Giacomin, P.; Loukas, A.; McCarthy, J.S. Experimental human hookworm infection: A narrative historical review. PLoS Negl. Trop. Dis. 2021, 15, e0009908. [Google Scholar] [CrossRef]
- Sauer, S.; Beinart, D.; Finn, S.M.B.; Kumar, S.L.; Cheng, Q.; Hwang, S.E.; Parker, W.; Devi, G.R. Hymenolepis diminuta-based helminth therapy in C3(1)-TAg mice does not alter breast tumor onset or progression. Evol. Med. Public Health 2021, 9, 131–138. [Google Scholar] [CrossRef]
- Harnett, W.; Harnett, M.M. Helminth-derived immunomodulators: Can understanding the worm produce the pill? Nat. Rev. Immunol. 2010, 10, 278–284. [Google Scholar] [CrossRef]
- Wangchuk, P.; Kouremenos, K.; Eichenberger, R.M.; Pearson, M.; Susianto, A.; Wishart, D.S.; McConville, M.J.; Loukas, A. Metabolomic profiling of the excretory-secretory products of hookworm and whipworm. Metabolomics 2019, 15, 101. [Google Scholar] [CrossRef]
- Zmyslowski, A.; Szterk, A. Current knowledge on the mechanism of atherosclerosis and pro-atherosclerotic properties of oxysterols. Lipids Health Dis. 2017, 16, 188. [Google Scholar] [CrossRef] [Green Version]
- Deroissart, J.; Porsch, F.; Koller, T.; Binder, C.J. Anti-inflammatory and Immunomodulatory Therapies in Atherosclerosis. Handb. Exp. Pharmacol. 2022, 270, 359–404. [Google Scholar] [CrossRef]
- Khan, R.; Spagnoli, V.; Tardif, J.C.; L’Allier, P.L. Novel anti-inflammatory therapies for the treatment of atherosclerosis. Atherosclerosis 2015, 240, 497–509. [Google Scholar] [CrossRef]
- Ridker, P.M.; Rane, M. Interleukin-6 Signaling and Anti-Interleukin-6 Therapeutics in Cardiovascular Disease. Circ. Res. 2021, 128, 1728–1746. [Google Scholar] [CrossRef] [PubMed]
- Hartman, J.; Frishman, W.H. Inflammation and atherosclerosis: A review of the role of interleukin-6 in the development of atherosclerosis and the potential for targeted drug therapy. Cardiol. Rev. 2014, 22, 147–151. [Google Scholar] [CrossRef] [PubMed]
- Summerhill, V.I.; Grechko, A.V.; Yet, S.F.; Sobenin, I.A.; Orekhov, A.N. The Atherogenic Role of Circulating Modified Lipids in Atherosclerosis. Int. J. Mol. Sci. 2019, 20, 3561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mushenkova, N.V.; Bezsonov, E.E.; Orekhova, V.A.; Popkova, T.V.; Starodubova, A.V.; Orekhov, A.N. Recognition of Oxidized Lipids by Macrophages and Its Role in Atherosclerosis Development. Biomedicines 2021, 9, 915. [Google Scholar] [CrossRef]
- Kattoor, A.J.; Kanuri, S.H.; Mehta, J.L. Role of Ox-LDL and LOX-1 in Atherogenesis. Curr. Med. Chem. 2019, 26, 1693–1700. [Google Scholar] [CrossRef]
- Fuhrman, B.; Partoush, A.; Volkova, N.; Aviram, M. Ox-LDL induces monocyte-to-macrophage differentiation in vivo: Possible role for the macrophage colony stimulating factor receptor (M-CSF-R). Atherosclerosis 2008, 196, 598–607. [Google Scholar] [CrossRef]
- Ali, A.H.; Younis, N.; Abdallah, R.; Shaer, F.; Dakroub, A.; Ayoub, M.A.; Iratni, R.; Yassine, H.M.; Zibara, K.; Orekhov, A.; et al. Lipid-Lowering Therapies for Atherosclerosis: Statins, Fibrates, Ezetimibe and PCSK9 Monoclonal Antibodies. Curr. Med. Chem. 2021, 28, 7427–7445. [Google Scholar] [CrossRef]
- Ruscica, M.; Ferri, N.; Santos, R.D.; Sirtori, C.R.; Corsini, A. Lipid Lowering Drugs: Present Status and Future Developments. Curr. Atheroscler. Rep. 2021, 23, 17. [Google Scholar] [CrossRef]
- Bandeali, S.; Farmer, J. High-density lipoprotein and atherosclerosis: The role of antioxidant activity. Curr. Atheroscler. Rep. 2012, 14, 101–107. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.; Ding, X.; Chen, F.; Wu, X.; Chen, Y.; Zhang, Q.; Cao, J.; Wang, J.; Dai, Y. Inhibition Effects of Nippostrongylus brasiliensis and Its Derivatives against Atherosclerosis in ApoE-/- Mice through Anti-Inflammatory Response. Pathogens 2022, 11, 1208. https://doi.org/10.3390/pathogens11101208
Yang Y, Ding X, Chen F, Wu X, Chen Y, Zhang Q, Cao J, Wang J, Dai Y. Inhibition Effects of Nippostrongylus brasiliensis and Its Derivatives against Atherosclerosis in ApoE-/- Mice through Anti-Inflammatory Response. Pathogens. 2022; 11(10):1208. https://doi.org/10.3390/pathogens11101208
Chicago/Turabian StyleYang, Yougui, Xin Ding, Fuzhong Chen, Xiaomin Wu, Yuying Chen, Qiang Zhang, Jun Cao, Junhong Wang, and Yang Dai. 2022. "Inhibition Effects of Nippostrongylus brasiliensis and Its Derivatives against Atherosclerosis in ApoE-/- Mice through Anti-Inflammatory Response" Pathogens 11, no. 10: 1208. https://doi.org/10.3390/pathogens11101208