


Cord Blood Adductomics Reveals Oxidative Stress Exposure Pathways of Bronchopulmonary Dysplasia

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design and Patient Sample
2.2. Clinical Data Collection
2.3. Cord Blood and Postnatal Peripheral Blood Collection
2.4. Plasma Adductomics
2.5. Statistical Analysis
3. Results
3.1. Characteristics of the Patient Cohort
3.2. Cord Blood Adductomics
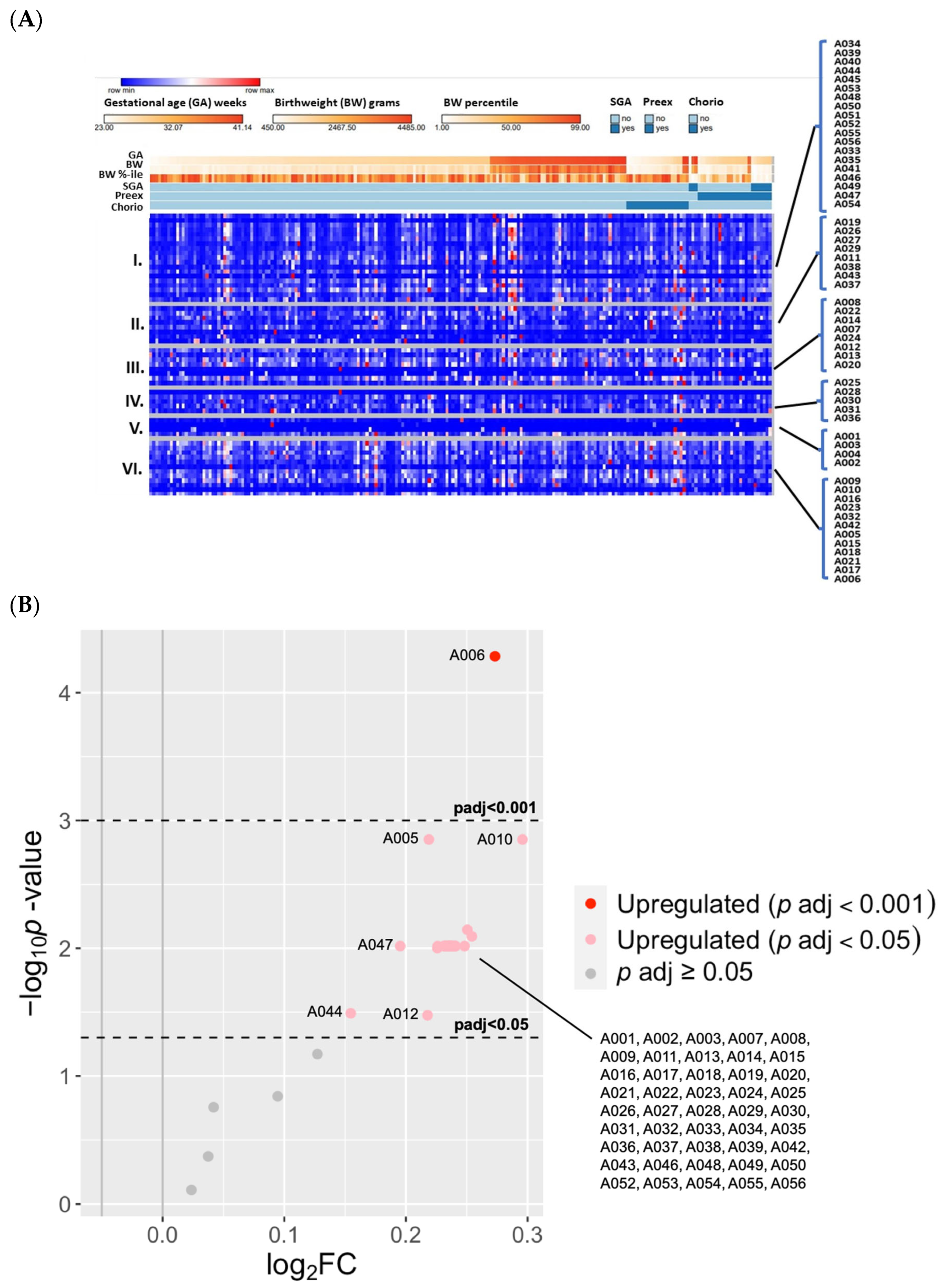
3.2.1. Known OS Adducts in Cord Blood Were Associated with Extreme Prematurity
3.2.2. Known OS Adducts Were Associated with Peripartum Exposures of Prematurity
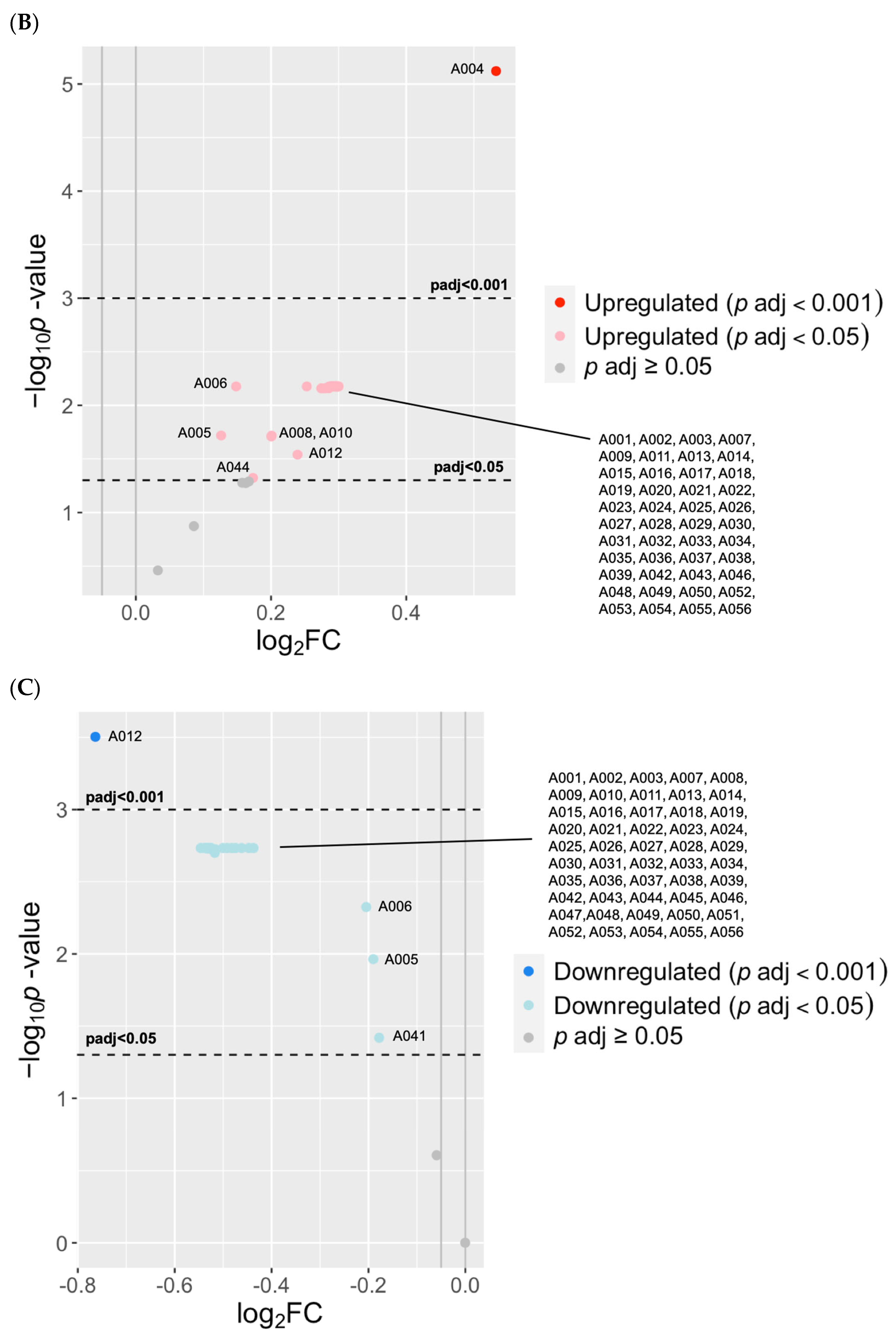
3.2.3. Known OS Adducts Were Associated with BPD and BPD-Associated Pulmonary Hypertension
3.3. Peripheral Blood (Postnatal) Adductomics
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siffel, C.; Kistler, K.D.; Lewis, J.F.M.; Sarda, S.P. Global incidence of bronchopulmonary dysplasia among extremely preterm infants: A systematic literature review. J. Matern. Fetal. Neonatal Med. 2021, 34, 1721–1731. [Google Scholar] [CrossRef] [PubMed]
- Cuestas, E.; Aguilera, B.; Cerutti, M.; Rizzotti, A. Sustained Neonatal Inflammation Is Associated with Poor Growth in Infants Born Very Preterm during the First Year of Life. J. Pediatr. 2018, 205, 91–97. [Google Scholar] [CrossRef]
- Gonçalves, D.d.M.M.; Wandalsen, G.F.; Scavacini, A.S.; Lanza, F.C.; Goulart, A.L.; Solé, D.; dos Santos, A.M.N. Pulmonary function in former very low birth weight preterm infants in the first year of life. Respir. Med. 2018, 136, 83–87. [Google Scholar] [CrossRef] [PubMed]
- Silva, L.V.; de Araújo, L.B.; Azevedo, V.M.G.d.O. Assessment of the neuropsychomotor development in the first year of life of premature infants with and without bronchopulmonary dysplasia. Rev. Bras. Ter. Intensiv. 2018, 30, 174–180. [Google Scholar] [CrossRef]
- Walsh, M.C.; Bell, E.F.; Kandefer, S.; Saha, S.; Carlo, W.A.; D′Angio, C.T.; Laptook, A.R.; Sanchez, P.J.; Stoll, B.J.; Shankaran, S.; et al. Neonatal outcomes of moderately preterm infants compared to extremely preterm infants. Pediatr. Res. 2017, 82, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Jensen, E.A.; Laughon, M.M.; DeMauro, S.B.; Cotten, C.M.; Do, B.; Carlo, W.A.; Watterberg, K.L. Contributions of the NICHD neonatal research network to the diagnosis, prevention, and treatment of bronchopulmonary dysplasia. Semin. Perinatol. 2022, 46, 151638. [Google Scholar] [CrossRef]
- Parsons, A.; Netsanet, A.; Seedorf, G.; Abman, S.H.; Taglauer, E.S. Understanding the role of placental pathophysiology in the development of bronchopulmonary dysplasia. Am. J. Physiol. Cell Mol. Physiol. 2022, 323, L651–L658. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Dong, W. Oxidative stress and bronchopulmonary dysplasia. Gene 2018, 678, 177–183. [Google Scholar] [CrossRef]
- Buonocore, G.; Perrone, S.; Tataranno, M. Oxidative stress and bronchopulmonary dysplasia. J. Clin. Neonatol. 2012, 1, 109–114. [Google Scholar] [CrossRef]
- Davis, J.M. Role of oxidant injury in the pathogenesis of neonatal lung disease. Acta Paediatr. Suppl. 2002, 91, 23–25. [Google Scholar] [CrossRef]
- Cheah, F.-C.; Jobe, A.H.; Moss, T.J.; Newnham, J.P.; Kallapur, S.G. Oxidative Stress in Fetal Lambs Exposed to Intra-amniotic Endotoxin in a Chorioamnionitis Model. Pediatr. Res. 2008, 63, 274–279. [Google Scholar] [CrossRef] [PubMed]
- Rappaport, S.M.; Li, H.; Grigoryan, H.; Funk, W.E.; Williams, E.R. Adductomics: Characterizing exposures to reactive electrophiles. Toxicol. Lett. 2012, 213, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Bourdon, E.; Blache, D. The importance of proteins in defense against oxidation. Antioxid. Redox Signal. 2001, 3, 293–311. [Google Scholar] [CrossRef] [PubMed]
- Grigoryan, H.; Edmands, W.; Lu, S.S.; Yano, Y.; Regazzoni, L.; Iavarone, A.T.; Williams, E.R.; Rappaport, S.M. Adductomics Pipeline for Untargeted Analysis of Modifications to Cys34 of Human Serum Albumin. Anal. Chem. 2016, 88, 10504–10512. [Google Scholar] [CrossRef] [PubMed]
- Grigoryan, H.; Li, H.; Iavarone, A.T.; Williams, E.R.; Rappaport, S.M. Cys34 adducts of reactive oxygen species in human serum albumin. Chem. Res. Toxicol. 2012, 25, 1633–1642. [Google Scholar] [CrossRef] [PubMed]
- Mestan, K.K.; Gotteiner, N.; Porta, N.; Grobman, W.; Su, E.J.; Ernst, L.M. Cord Blood Biomarkers of Placental Maternal Vascular Underperfusion Predict Bronchopulmonary Dysplasia-Associated Pulmonary Hypertension. J. Pediatr. 2017, 185, 33–41. [Google Scholar] [CrossRef]
- Voller, S.B.; Chock, S.; Ernst, L.M.; Su, E.; Liu, X.; Farrow, K.N.; Mestan, K.K. Cord blood biomarkers of vascular endothelial growth (VEGF and sFlt-1) and postnatal growth: A preterm birth cohort study. Early Hum. Dev. 2014, 90, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.M.; Birkett, R.; Lin, E.T.; Ernst, L.M.; Grobman, W.A.; Swaminathan, S.; Abdala-Valencia, H.; Misharin, A.V.; Bartom, E.T.; Mestan, K.K. Placental dysfunction influences fetal monocyte subpopulation gene expression in preterm birth. J. Clin. Investig. 2022, 7, 155482. [Google Scholar] [CrossRef]
- Jones, C.; Titus, H.; Bs, C.G.B.; Soviravong, S.; Stansfield, B.K. Evaluating definitions for maternal fever as diagnostic criteria for intraamniotic infection in low-risk pregnancies. Birth 2021, 48, 389–396. [Google Scholar] [CrossRef]
- Gestational Hypertension and Preeclampsia: ACOG Practice Bulletin, Number 222. Obstet. Gynecol. 2020, 135, e237–e260. [CrossRef]
- Kim, C.J.; Romero, R.; Chaemsaithong, P.; Chaiyasit, N.; Yoon, B.H.; Kim, Y.M. Acute chorioamnionitis and funisitis: Definition, pathologic features, and clinical significance. Am. J. Obstet. Gynecol. 2015, 213 (Suppl. S4), S29–S52. [Google Scholar] [CrossRef] [PubMed]
- Palmsten, K.; Nelson, K.K.; Laurent, L.C.; Park, S.; Chambers, C.D.; Parast, M.M. Subclinical and clinical chorioamnionitis, fetal vasculitis, and risk for preterm birth: A cohort study. Placenta 2018, 67, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Fenton, T.R. A new growth chart for preterm babies: Babson and Benda’s chart updated with recent data and a new format. BMC Pediatr. 2003, 3, 13. [Google Scholar] [CrossRef] [PubMed]
- Higgins, R.D.; Jobe, A.H.; Koso-Thomas, M.; Bancalari, E.; Viscardi, R.M.; Hartert, T.V.; Ryan, R.M.; Kallapur, S.G.; Steinhorn, R.H.; Konduri, G.G.; et al. Bronchopulmonary Dysplasia: Executive Summary of a Workshop. J. Pediatr. 2018, 197, 300–308. [Google Scholar] [CrossRef] [PubMed]
- Mestan, K.; Check, J.; Minturn, L.; Yallapragada, S.; Farrow, K.; Liu, X.; Su, E.; Porta, N.; Gotteiner, N.; Ernst, L. Placental pathologic changes of maternal vascular underperfusion in bronchopulmonary dysplasia and pulmonary hypertension. Placenta 2014, 35, 570–574. [Google Scholar] [CrossRef] [PubMed]
- Wai, K.C.; Kohn, M.A.; Ballard, R.A.; Truog, W.E.; Black, D.M.; Asselin, J.M.; Ballard, P.L.; Rogers, E.E.; Keller, R.L. Early Cumulative Supplemental Oxygen Predicts Bronchopulmonary Dysplasia in High Risk Extremely Low Gestational Age Newborns. J. Pediatr. 2016, 177, 97–102.e2. [Google Scholar] [CrossRef] [PubMed]
- Funk, W.E.; Montgomery, N.; Bae, Y.; Chen, J.; Chow, T.; Martinez, M.P.; Lurmann, F.; Eckel, S.P.; McConnell, R.; Xiang, A.H. Human Serum Albumin Cys34 Adducts in Newborn Dried Blood Spots: Associations With Air Pollution Exposure During Pregnancy. Front. Public Health 2021, 9, 730369. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.Y.; Imani, P.; Grigoryan, H.; Bassig, B.A.; Dai, Y.; Hu, W.; Blechter, B.; Rahman, M.L.; Ji, B.T.; Duan, H.; et al. Exposure to diesel engine exhaust and alterations to the Cys34/Lys525 adductome of human serum albumin. Environ. Toxicol. Pharmacol. 2022, 95, 103966. [Google Scholar] [CrossRef] [PubMed]
- Petrick, L.M.; Uppal, K.; Funk, W.E. Metabolomics and adductomics of newborn bloodspots to retrospectively assess the early-life exposome. Curr. Opin. Pediatr. 2020, 32, 300–307. [Google Scholar] [CrossRef]
- Law, C.W.; Alhamdoosh, M.; Su, S.; Dong, X.; Tian, L.; Smyth, G.K.; Ritchie, M.E. RNA-seq analysis is easy as 1-2-3 with limma, Glimma and edgeR. F1000Research 2016, 5, 1408. [Google Scholar] [CrossRef]
- Carlsson, H.; Rappaport, S.M.; Törnqvist, M. Protein Adductomics: Methodologies for Untargeted Screening of Adducts to Serum Albumin and Hemoglobin in Human Blood Samples. High Throughput 2019, 8, 6. [Google Scholar] [CrossRef] [PubMed]
- Grigoryan, H.; Edmands, W.M.B.; Lan, Q.; Carlsson, H.; Vermeulen, R.; Zhang, L.; Yin, S.-N.; Li, G.-L.; Smith, M.T.; Rothman, N.; et al. Adductomic signatures of benzene exposure provide insights into cancer induction. Carcinog. 2018, 39, 661–668. [Google Scholar] [CrossRef]
- Liu, S.; Grigoryan, H.; Edmands, W.M.B.; Dagnino, S.; Sinharay, R.; Cullinan, P.; Collins, P.; Chung, K.F.; Barratt, B.; Kelly, F.J.; et al. Cys34 Adductomes Differ between Patients with Chronic Lung or Heart Disease and Healthy Controls in Central London. Environ. Sci. Technol. 2018, 52, 2307–2313. [Google Scholar] [CrossRef]
- Lu, S.S.; Grigoryan, H.; Edmands, W.M.B.; Hu, W.; Iavarone, A.T.; Hubbard, A.; Rothman, N.; Vermeulen, R.; Lan, Q.; Rappaport, S.M. Profiling the Serum Albumin Cys34 Adductome of Solid Fuel Users in Xuanwei and Fuyuan, China. Environ. Sci. Technol. 2017, 51, 46–57. [Google Scholar] [CrossRef] [PubMed]
- Yano, Y.; Grigoryan, H.; Schiffman, C.; Edmands, W.; Petrick, L.; Hall, K.; Whitehead, T.; Metayer, C.; Dudoit, S.; Rappaport, S. Untargeted adductomics of Cys34 modifications to human serum albumin in newborn dried blood spots. Anal. Bioanal. Chem. 2019, 411, 2351–2362. [Google Scholar] [CrossRef] [PubMed]
- Yano, Y.; Schiffman, C.; Grigoryan, H.; Hayes, J.; Edmands, W.; Petrick, L.; Whitehead, T.; Metayer, C.; Dudoit, S.; Rappaport, S. Untargeted adductomics of newborn dried blood spots identifies modifications to human serum albumin associated with childhood leukemia. Leuk. Res. 2020, 88, 106268. [Google Scholar] [PubMed]
- Grigoryan, H.; Imani, P.; Dudoit, S.; Rappaport, S.M. Extending the HSA-Cys34-Adductomics Pipeline to Modifications at Lys525. Chem. Res. Toxicol. 2021, 34, 2549–2557. [Google Scholar] [CrossRef] [PubMed]
- Grigoryan, H.; Schiffman, C.; Gunter, M.J.; Naccarati, A.; Polidoro, S.; Dagnino, S.; Dudoit, S.; Vineis, P.; Rappaport, S.M. Cys34 Adductomics Links Colorectal Cancer with the Gut Microbiota and Redox Biology. Cancer Res. 2019, 79, 6024–6031. [Google Scholar] [CrossRef] [PubMed]
- Perez, M.; Robbins, M.E.; Revhaug, C.; Saugstad, O.D. Oxygen radical disease in the newborn, revisited: Oxidative stress and disease in the newborn period. Free. Radic. Biol. Med. 2019, 142, 61–72. [Google Scholar] [CrossRef]
- Deng, X.; Bao, Z.; Yang, X.; Mei, Y.; Zhou, Q.; Chen, A.; Yu, R.; Zhang, Y. Molecular mechanisms of cell death in bronchopulmonary dysplasia. Apoptosis 2023, 28, 39–54. [Google Scholar] [CrossRef]
- Ferrante, G.; Montante, C.; Notarbartolo, V.; Giuffrè, M. Antioxidants: Role the in prevention and treatment of bronchopulmonary dysplasia. Paediatr. Respir. Rev. 2022, 42, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Oei, J.L.; Vento, M.; Rabi, Y.; Wright, I.; Finer, N.; Rich, W.; Kapadia, V.; Aune, D.; Rook, D.; Tarnow-Mordi, W.; et al. Higher or lower oxygen for delivery room resuscitation of preterm infants below 28 completed weeks gestation: A meta-analysis. Arch. Dis. Child. Fetal Neonatal Ed. 2017, 102, F24–F30. [Google Scholar] [CrossRef] [PubMed]
- Kapadia, V.S.; Lal, C.V.; Kakkilaya, V.; Heyne, R.; Savani, R.C.; Wyckoff, M.H. Impact of the Neonatal Resuscitation Program–Recommended Low Oxygen Strategy on Outcomes of Infants Born Preterm. J. Pediatr. 2017, 191, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Tagliaferro, T.; Jain, D.; Vanbuskirk, S.; Bancalari, E.; Claure, N. Maternal preeclampsia and respiratory outcomes in extremely premature infants. Pediatr. Res. 2019, 85, 693–696. [Google Scholar] [CrossRef] [PubMed]
- Dravet-Gounot, P.; Torchin, H.; Goffinet, F.; Aubelle, M.-S.; El Ayoubi, M.; Lefevre, C.; Jarreau, P.-H.; Zana-Taïeb, E. Bronchopulmonary dysplasia in neonates born to mothers with preeclampsia: Impact of small for gestational age. PLoS ONE 2018, 13, e0204498. [Google Scholar] [CrossRef] [PubMed]
- Elkabany, Z.A.; El-Farrash, R.A.; Shinkar, D.M.; Ismail, E.A.; Nada, A.S.; Farag, A.S.; Elsayed, M.A.; Salama, D.H.; Macken, E.L.; Gaballah, S.A. Oxidative stress markers in neonatal respiratory distress syndrome: Advanced oxidation protein products and 8-hydroxy-2-deoxyguanosine in relation to disease severity. Pediatr. Res. 2020, 87, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Longini, M.; Belvisi, E.; Proietti, F.; Bazzini, F.; Buonocore, G.; Perrone, S. Oxidative Stress Biomarkers: Establishment of Reference Values for Isoprostanes, AOPP, and NPBI in Cord Blood. Mediat. Inflamm. 2017, 2017, 1758432. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Tian, F.-J.; Lin, Y. Oxidative Stress in Placenta: Health and Diseases. BioMed Res. Int. 2015, 2015, 293271. [Google Scholar] [CrossRef] [PubMed]
- Dizdar, E.A.; Uras, N.; Oguz, S.; Erdeve, O.; Sari, F.N.; Aydemir, C.; Dilmen, U. Total antioxidant capacity and total oxidant status after surfactant treatment in preterm infants with respiratory distress syndrome. Ann. Clin. Biochem. 2011, 48, 462–467. [Google Scholar] [CrossRef]
- Chung, M.-K.; Grigoryan, H.; Iavarone, A.T.; Rappaport, S.M. Antibody enrichment and mass spectrometry of albumin-Cys34 adducts. Chem. Res. Toxicol. 2014, 27, 400–407. [Google Scholar] [CrossRef]
- Behl, T.; Rachamalla, M.; Najda, A.; Sehgal, A.; Singh, S.; Sharma, N.; Bhatia, S.; Al-Harrasi, A.; Chigurupati, S.; Vargas-De-La-Cruz, C.; et al. Applications of Adductomics in Chemically Induced Adverse Outcomes and Major Emphasis on DNA Adductomics: A Pathbreaking Tool in Biomedical Research. Int. J. Mol. Sci. 2021, 22, 10141. [Google Scholar] [CrossRef] [PubMed]
- Ogihara, T.; Hirano, K.; Morinobu, T.; Kim, H.-S.; Hiroi, M.; Ogihara, H.; Tamai, H. Raised concentrations of aldehyde lipid peroxidation products in premature infants with chronic lung disease. Arch. Dis. Child. Fetal Neonatal Ed. 1999, 80, F21–F25. [Google Scholar] [CrossRef] [PubMed]
- Stevens, R.; Pass, K.; Fuller, S.; Wiznia, A.; Noble, L.; Duva, S.; Neal, M. Blood spot screening and confirmatory tests for syphilis antibody. J. Clin. Microbiol. 1992, 30, 2353–2358. [Google Scholar] [CrossRef] [PubMed]
- Worthman, C.M.; Stallings, J.F. Hormone measures in finger-prick blood spot samples: New field methods for reproductive endocrinology. Am. J. Phys. Anthropol. 1997, 104, 1–21. [Google Scholar] [CrossRef]
- McDade, T.W.; Stallings, J.F.; Angold, A.; Costello, E.J.; Burleson, M.; Cacioppo, J.T.; Glaser, R.; Worthman, C.M. Epstein-Barr Virus Antibodies in Whole Blood Spots: A Minimally Invasive Method for Assessing an Aspect of Cell-Mediated Immunity. Psychosom. Med. 2000, 62, 560–568. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, T.A.; Kler, J.S.; Bae, Y.; Chen, J.; Ladror, D.T.; Iyer, R.; Nunes, D.A.; Montgomery, N.D.; Pleil, J.D.; Funk, W.E. A state-of-the-science review and guide for measuring environmental exposure biomarkers in dried blood spots. J. Expo. Sci. Environ. Epidemiol. 2022, 33, 505–523. [Google Scholar] [CrossRef]
- Jacobson, T.A.; Bae, Y.; Kler, J.S.; Iyer, R.; Zhang, R.; Montgomery, N.D.; Nunes, D.; Pleil, J.D.; Funk, W.E. Advancing Global Health Surveillance of Mycotoxin Exposures using Minimally Invasive Sampling Techniques: A State-of-the-Science Review. Environ. Sci. Technol. 2024, 58, 3580–3594. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Full Term (n = 51) | No BPD (n = 103) | BPD (n = 102) | p * | |
|---|---|---|---|---|
| MATERNAL | ||||
| Maternal age, years, (mean ± SD) | 34.6 ± 4.1 | 30.5 ± 5.6 | 32.3 ± 5.6 | 0.02 |
| Maternal race, n (%) | ||||
| White | 40 (78.4) | 41 (39.8) | 42 (41.2) | 0.1 |
| Black/African American | 3 (5.9) | 30 (29.1) | 24 (23.5) | |
| Asian | 1 (2.0) | 5 (4.9) | 7 (6.9) | |
| American Indian/Alaska Native | 0 (0) | 2 (1.9) | 2 (2.0) | |
| Other | 5 (9.8) | 19 (18.4) | 16 (15.7) | |
| Declined/Unknown | 2 (3.9) | 6 (5.8) | 11 (10.8) | |
| Ethnicity, n (%) | ||||
| Hispanic or Latino | 10 (19.6) | 25 (24.3) | 18 (17.6) | 0.47 |
| Not Hispanic or Latino | 41 (80.4) | 73 (70.9) | 75 (73.5) | |
| Declined/Unknown | 0 (0) | 5 (4.9) | 9 (8.8) | |
| Smoking status, n (%) | ||||
| Current smoker | 2 (3.9) | 4 (3.9) | 0 (0) | 0.05 |
| Former smoker | 4 (7.8) | 14 (13.6) | 8 (7.8) | |
| Never smoker | 42 (82.4) | 43 (41.7) | 38 (37.3) | |
| Unknown | 3 (5.9) | 42 (40.8) | 56 (54.9) | |
| Rupture of membranes, n (%) | ||||
| Artificial | 42 (82.4) | 50 (48.5) | 49 (48) | 1 |
| Spontaneous | 9 (17.6) | 52 (50.5) | 53 (52) | |
| Delivery method, n (%) | ||||
| Cesarean | 7 (13.7) | 58 (56.3) | 68 (66.7) | 0.17 |
| Vaginal | 44 (86.3) | 45 (43.7) | 34 (33.3) | |
| Twin delivery, n (%) | 2 (3.9) | 33 (32) | 30 (29.4) | 0.80 |
| Antenatal steroids, n (%) | ||||
| None | 51 (100) | 6 (5.8) | 6 (5.9) | 0.32 |
| Complete | 0 (0) | 83 (80.6) | 74 (72.5) | |
| Incomplete | 0 (0) | 14 (13.6) | 22 (21.6) | |
| Preterm labor | 0 (0) | 75 (72.8) | 64 (62.7) | 0.16 |
| Preterm premature rupture of membranes | 0 (0) | 51 (49.5) | 48 (47.1) | 0.83 |
| Prolonged rupture of membranes | 3 (5.9) | 31 (30.1) | 31 (30.4) | 1 |
| Pre-eclampsia/HELLP | 1 (2) | 17 (16.5) | 27 (26.5) | 0.14 |
| Antenatal IUGR | 0 (0) | 2 (1.9) | 16 (15.7) | 0.001 |
| Non-reassuring fetal heart tones | 3 (5.9) | 14 (13.6) | 19 (18.6) | 0.43 |
| Chorioamnionitis | 49 (96.1) | 73 (70.9) | 68 (66.7) | 0.62 |
| Diabetes, n (%) | ||||
| None | 49 (96.1) | 98 (95.1) | 94 (92.2) | 0.67 |
| Gestational | 2 (3.9) | 4 (3.9) | 6 (5.9) | |
| Insulin-dependent | 0 (0) | 1 (1) | 2 (2) | |
| INFANT | ||||
| Gestational age, weeks, (mean ± SD) | 39.5 ± 1.0 | 27.7 ± 1.2 | 26.1 ± 1.6 | <0.001 |
| Gestational age category, n (%) | ||||
| 22–24 weeks | - | 1 (1.0) | 25 (24.5) | <0.001 |
| 25–26 weeks | - | 25 (24.3) | 39 (38.2) | |
| 27–29 weeks | - | 77 (74.8) | 38 (37.3) | |
| Infant sex, n (%) | ||||
| Female | 25 (49) | 45 (43.7) | 52 (51) | 0.37 |
| Male | 26 (51) | 58 (56.3) | 50 (49) | |
| Birth weight (g), (mean ± SD) | 3430 ± 452 | 1110 ± 201 | 854 ± 202 | <0.001 |
| Birth-weight-for-GA percentile (%) | 52.8 ± 26.3 | 64.3 ± 21.2 | 56.3 ± 28.4 | 0.02 |
| Small-for-gestational-age status, n (%) | 2 (3.9) | 1 (1) | 9 (8.8) | 0.02 |
| APGAR score, median [min, max] | <0.001 | |||
| 1-min | 8 [2, 9] | 6 [0, 9] | 4 [0, 8] | |
| 5-min | 9 [8, 9] | 8 [1, 9] | 7 [1, 9] | |
| BPD grade, n (%) | ||||
| Grade I | - | 0 (0) | 33 (32.4) | -- |
| Grade II | - | 0 (0) | 26 (25.5) | |
| Grade III/IIIA | - | 0 (0) | 43 (42.2) | |
| Other neonatal complications, n (%) | ||||
| Necrotizing enterocolitis (any) | 0 (0) | 6 (5.8) | 24 (23.5) | <0.001 |
| Intraventricular hemorrhage (any) | 0 (0) | 24 (23.3) | 42 (41.2) | <0.001 |
| Retinopathy of prematurity (stage 2+) | 0 (0) | 9 (8.7) | 32 (31.4) | <0.001 |
| Pulmonary hypertension @ 36 weeks | 0 (0) | 8 (7.8) | 28 (27.5) | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, E.T.; Bae, Y.; Birkett, R.; Sharma, A.M.; Zhang, R.; Fisch, K.M.; Funk, W.; Mestan, K.K. Cord Blood Adductomics Reveals Oxidative Stress Exposure Pathways of Bronchopulmonary Dysplasia. Antioxidants 2024, 13, 494. https://doi.org/10.3390/antiox13040494
Lin ET, Bae Y, Birkett R, Sharma AM, Zhang R, Fisch KM, Funk W, Mestan KK. Cord Blood Adductomics Reveals Oxidative Stress Exposure Pathways of Bronchopulmonary Dysplasia. Antioxidants. 2024; 13(4):494. https://doi.org/10.3390/antiox13040494
Chicago/Turabian StyleLin, Erika T., Yeunook Bae, Robert Birkett, Abhineet M. Sharma, Runze Zhang, Kathleen M. Fisch, William Funk, and Karen K. Mestan. 2024. "Cord Blood Adductomics Reveals Oxidative Stress Exposure Pathways of Bronchopulmonary Dysplasia" Antioxidants 13, no. 4: 494. https://doi.org/10.3390/antiox13040494
APA StyleLin, E. T., Bae, Y., Birkett, R., Sharma, A. M., Zhang, R., Fisch, K. M., Funk, W., & Mestan, K. K. (2024). Cord Blood Adductomics Reveals Oxidative Stress Exposure Pathways of Bronchopulmonary Dysplasia. Antioxidants, 13(4), 494. https://doi.org/10.3390/antiox13040494